

Палеонтологический журнал, 2019, № 6, стр. 91-104
Черепахи Protestudo bessarabica из позднего миоцена Украины
Е. В. Сыромятникова a, b, *, Т. В. Крахмальная c, В. М. Чхиквадзе d, И. Г. Данилов b
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Зоологический институт РАН
Санкт-Петербург, Россия
c Национальный научно-природоведческий музей НАН Украины
Киев, Украина
d Институт палеобиологии Национального музея Грузии
Тбилиси, Грузия
* E-mail: esyromyatnikova@gmail.com
Поступила в редакцию 13.02.2019
После доработки 15.02.2019
Принята к публикации 18.02.2019
Аннотация
В статье описаны три неполных панциря сухопутных черепах Protestudo bessarabica (Testudinidae) из позднего миоцена (мэотис, биозона MN 12) местонахождения Белка (Украина). Это первое описание данного вида с территории Украины. Материалы из Белки дополняют данные по морфологии и изменчивости панциря P. bessarabica в отношении формулы невральных пластинок, формы прецентрального щитка и гулярного выступа. Кроме того, они подтверждают ранее выявленную изменчивость панциря P. bessarabica по ряду других признаков. Этот вид, по-видимому, имел широкое географическое распространение в Восточной Европе (Молдова, Россия, Украина) в достаточно узком временном интервале позднего сармата–мэотиса, что определяет его важное значение для биостратиграфической корреляции.
ВВЕДЕНИЕ
Protestudo (изначально Testudo) bessarabica (Riabinin, 1918) – типовой вид рода Protestudo Chkhikvadze, 1970 семейства Testudinidae Batsch, 1788. Черепахи этого вида отличаются от других видов Protestudo наличием прецентрального щитка и сильно вытянутым вперед, но не остроконечным гулярным выступом (Данилов и др., 2017). Они известны из мэотиса (=среднего туролия) Молдовы, России и Украины. Впервые этот вид был описан по нескольким неполным панцирям и одному внутреннему ядру из мэотиса Молдовы (местонахождения Тараклия и Тудорово; Рябинин, 1918). В той же работе из Тараклии по еще одному панцирю был описан Testudo tarakliensis Riabinin, 1918. Позднее к T. bessarabica был отнесен отдельный карапакс из местонахождения Чобручи (мэотис Молдовы; Macarovici, 1930). Изображение этого карапакса и пластрона, возможно, от того же экземпляра было опубликовано позже в другой работе (Macarovici, Vancea, 1960). Еще один карапакс и фрагмент передней части панциря были подробно описаны из местонахождения Чимишлия (мэотис Молдовы; Хозацкий, Тарабукин, 1970). В то же время T. tarakliensis был сведен в синонимы T. bessarabica (Хозацкий, Тофан, 1970), а сам этот вид был выделен в самостоятельный род Protestudo, к которому в серии публикаций было отнесено более 20 видов черепах из неогена Азии, Европы и Северной Америки (Чхиквадзе, 1970; и др.). Впоследствии к Protestudo bessarabica были отнесены дополнительные материалы из местонахождений Чобручи (карапакс; Хозацкий, Редкозубов, 1989) и Недвиговка [?сармат (=поздний арагоний); Ростовская область, пластрон; Чхиквадзе, 1989], однако пластрон из Недвиговки был только изображен, но не описан. Помимо этого, в литературе были упомянуты и другие панцирные материалы по этому виду из перечисленных и других местонахождений позднего миоцена Молдавии и Украины [Чекур-Менжир, Ченак, Белка, Гребеники, Новая Эметовка 2 (=Костев овраг), Новоелизаветовка, Новоукраинка, Черевичное; см. Данилов и др., 201711]. Недавно было опубликовано новое изображение одного из типовых экземпляров (лектотип: Danilov, 2005, рис. 82A, B ; Данилов и др., 2017, рис. 103 , табл. XXX ), однако этот и другие экземпляры типовой серии не переописывались. При этом, несмотря на свою полноту, типовые материалы малоинформативны, так как границы многих их костных и роговых элементов неразличимы, что не позволяет наблюдать многие важные признаки строения панциря. Многочисленность упомянутых в литературе находок P. bessarabica свидетельствует о широком распространение этих черепах в Восточной Европе в конце миоцена (позднем сармате – мэотисе). Однако отсутствие качественных описаний и изображений большей части перечисленных материалов делают актуальным их детальное изучение. Помимо самостоятельной ценности, это важно для дополнения характеристики рода Protestudo, статус которого остается спорным (см., напр., Lapparent de Broin, 2001; Perälä, 2002; Lapparent de Broin et al., 2006; Rhodin et al., 2008; Gnetneva et al., 2018).
Данная статья посвящена описанию материалов по P. bessarabica из местонахождения Белка, которые ранее только упоминались в литературе, но не описывались и не изображались (Чхиквадзе, 1983; Короткевич, 1988; Данилов и др., 2017). Их совместное изучение было начато в 2010 г. и продолжено в 2016 г.
Местонахождение Белка располагается на окраине с. Белка Ивановского р-на Одесской области, Украина (46°53′24′′ с. ш.; 30°25′28′′ в. д.; рис. 1). Оно расположено на высоком склоне оврага (рис. 2). Раскопки местонахождения проводились с 1960 по 1966 гг. сотрудниками Отдела палеозоологии Ин-та зоологии АН УССР (Короткевич, 1988). Костеносный горизонт залегает на глубине 4 м от поверхности и представлен суглинками и супесями (часто слоистыми), внизу горизонта их сменяет мелкозернистый песок. Верхняя часть костеносного горизонта содержит мелкие фрагменты костей. Средняя часть горизонта в основном образована брекчией из деформированных костей, вжатых друг в друга, хотя здесь же были обнаружены целые кости и отдельные анатомические группы, принадлежащие крупным млекопитающим. В нижней части костеносного горизонта также находили целые кости. Из покрывающих и подстилающих его слоев известны раковины речных моллюсков Unio sp. (Короткевич, 1988).
Рис. 2.
Общий вид местонахождения Белка (Одесская обл., Украина; место раскопок обозначено стрелками). Фото Д.В. Иванова (2003).

В 2003 г. раскопки местонахождения были возобновлены Отделом палеозоологии позвоночных и Палеонтологическим музеем (с 2016 г. – Отдел палеонтологии) Национального научно-природоведческого музея НАН Украины, при участии одного из авторов (Т.В. Крахмальная), в результате которых удалось пополнить сборы ископаемого материала прошлых лет. За все годы раскопок количество костных остатков превысило 7000 единиц, большинство из которых принадлежит крупным млекопитающим, тогда как остатки рептилий и птиц немногочисленны. Видовой состав Белки по состоянию изученности на сегодняшний день представлен следующими таксонами (по: Короткевич, 1988; Semenov, 2001; Krakhmalnaya, 2008, с уточнениями): черепахи Protestudo bessarabica; птицы Miophasianus sp., Miopica paradoxa Kurochkin et Sobolev, 2004 и Struthio asiaticus Milne-Edwards, 1871; млекопитающие Palinhyaena bessarabica (Simionescu, 1938), Machairodus giganteus (Wagner, 1848), Ictitherium viverrinum Roth et Wagner, 1854, Hyaenictitherium venator Semenov, 1989, Adcrocuta eximia (Roth et Wagner, 1854), Zygolophodon turicensis (Vacek, 1877), Deinotherium giganteum Kaup, 1829, Hipparion moldavicum Gromova, 1952, Aceratherium incisivum (Cuvier, 1822), Stephanorhinus orientalis (Schlosser, 1921), Microstonyx major (Gervais, 1848), ?Procapreolus ucrainicus Korotkevich, 1965, Palaeotragus rouenii Gaudry, 1861, Samotherium sp., Procapra rodleri (Pilgrim et Hopwood, 1928), Graecoryx bonus (Korotkevich, 1971).
Белкинская фаунистическая группировка по систематическому составу хорошо отличается от позднесарматских и близка к типичным мэотическим. Она относится к Белкинскому фаунистичекому комплексу и рассматривается как его средняя стадия (белкинский подкомплекс, средний мэотис; Короткевич, 1988; Крахмальная, 1996, 2008; Krakhmalnaya, 1996; Semenov, 2001). По мнению Е.Л. Короткевич (1988), вся толща песков и суглинков образовалась в результате деятельности древней реки, медленное течение которой способствовало захоронению почти целых скелетов небольших позвоночных, оказавшихся на периферийных участках костеносного слоя.
Из трех изученных панцирей P. bessarabica из Белки ниже подробно описывается наиболее полный экземпляр – экз. NMNHU-P, № 48-7118, а для остальных (экз. NMNHU-P, №№ 48-7116 и 48-7117) указываются только отличительные особенности строения. В тексте использовалась анатомическая терминология И.Г. Данилова с соавт. (2017). Изученный материал хранится в фондах отдела палеозоологии Национального научно-природоведческого музея НАН Украины, Киев (NMNHU-P). Другие материалы, упоминаемые в статье, хранятся в следующих учреждениях: ГКМ – Национальный музей этнографии и естественной истории, Кишинев (Молдова); ГЛЯУ – Геологическая лаборатория Ясского ун-та им. Ал.И. Кузы, Бухарест (Румыния); ПИН – Палеонтологический ин-т им. А.А. Борисяка РАН, Москва (Россия); ТКМ – Таганрогский краеведческий музей, Таганрог (Россия).
Работа выполнена в рамках государственной темы АААА-А19-119020590095-9 и при поддержке гранта РФФИ 19-04-00514. Мы благодарим рецензентов А.О. Аверьянова (Зоологический ин-т РАН) и Е.А. Звонка (Луганский национальный ун-т им. Тараса Шевченко) за ценные замечания, а также Д.В. Иванова за предоставленные фото местонахождения.
ОПИСАНИЕ НОВОГО МАТЕРИАЛА
Экз. NMNHU-P, № 48-7118 (OF-1169) (рис. 3, 4) представлен неполным панцирем без правых III–IX периферальных пластинок. Длина панциря составляет около 190 мм. Панцирь высокий, с наибольшей высотой на уровне IV костальных пластинок.
Рис. 3.
Protestudo bessarabica (Riabinin, 1918), экз. NMNHU-P, № 48-7118, неполный панцирь; местонахождение Белка, Украина, поздний миоцен, мэотис (MN 12): а, б – сверху; в, г – снизу; а, в – фото; б, г – поясняющие рисунки. Обозначения: abd – абдоминальный щиток; an – анальный щиток; epi – эпипластрон; ent – энтопластрон; fe – феморальный щиток; gu – гулярный щиток; hu – гумеральный щиток; hyo – гиопластрон; hypo – гипопластрон; n1 – I невральная пластинка; nu – нухальная пластинка; p1 – I периферальная пластинка; pe – пекторальный щиток; sp1 и 2 – супрапигальные пластинки; xi – ксифипластрон; порода показана серым цветом; сломы костей – штриховкой, шовные поверхности – точками; обломанные и предполагаемые части показаны пунктиром.
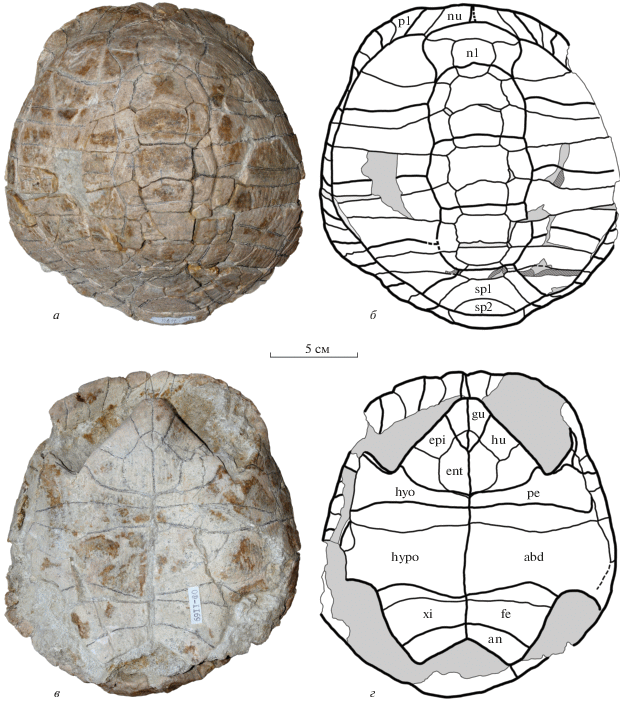
Рис. 4.
Protestudo bessarabica (Riabinin, 1918), экз. NMNHU-P, № 48-7118, неполный панцирь; местонахождение Белка, Украина; поздний миоцен, мэотис (MN 12): a, б – спереди; в, г – сзади; д, е – сбоку; а, в, д – фото; б, г, е – поясняющие рисунки. Обозначения: ce – прецентральный щиток; n8 – VIII невральная пластинка; py – пигальная пластинка; см. также подписи к рис. 3.
Карапакс округлый, расширен сзади (наибольшая ширина находится на уровне VI–VIII периферальных пластинок). Ширина карапакса составляет около 90% от его длины. В задней части карапакс круто изогнут книзу, что может указывать на принадлежность панциря самцу. Нухальная вырезка хорошо развита, образована нухальной пластинкой и передними частями I периферальных пластинок. Нухальная пластинка широкая, с почти прямым передним краем. Невральных пластинок восемь, их формула 4 < 8 > 4 < 8 > 4 < 6 < 6 > 6. I невральная пластинка удлиненная (длина больше ширины), последующие невральные широкие (ширина больше длины). Задние пластинки (VI–VIII) шестиугольные: VI и VII – с короткими переднебоковыми сторонами, VIII – c одинаковыми по длине передне- и заднебоковыми краями. Супрапигальных пластинoк две. I супрапигальная пластинка крупная трапециевидная, суженная спереди. Ее боковые края почти прямые. II супрапигальная – маленькая, округленно-треугольная, ее ширина в 2 раза превышает длину. Пигальная пластинка трапециевидная, суженная сзади, ее длина примерно равна ее максимальной ширине (вдоль переднего края). Костальных пластинок восемь пар. Они характерной для тестудинид попеременно-клиновидной формы. Периферальные пластинки представлены I–XI элементами на левой стороне панциря и I, II, VIII–XI – на правой стороне. Все периферальные пластинки четырехугольные (кроме XI пары, которая трапециевидная). Мостовые периферальные пластинки высокие.
Прецентральный щиток очень узкий трапециевидный (чуть более широкий сзади). Его длина составляет около половины от длины нухальной пластинки. На внутренней поверхности нухальной пластинки видна только передняя часть прецентрального щитка, который также трапециевидный и немного шире, чем снаружи. Центральные щитки относительно широкие. I щиток четырехугольный, с S-образно изогнутыми боковыми краями, более широкий спереди. Его передний край достигает правой I периферальной пластинки, но не достигает левой. Последующие центральные щитки уже I центрального. IV центральный щиток слегка сужен сзади. V центральный щиток длинный, трапециевидный, расширен сзади. Плевральные щитки широкие (ширина превышает длину). Маргинальные щитки по ширине (высоте) соответствуют периферальным пластинкам, плевро-маргинальная борозда проходит по косто-периферальному шву. На VII‒VIII периферальных пластинках слева плевро-маргинальная борозда немного отдалена от косто-периферального шва. XII маргинальные щитки слиты в один щиток с дорсальной стороны пигальной пластинки, а их состояние на вентральной стороне пластинки неизвестно, так как эта часть скрыта породой.
Пластрон сохранился почти полностью. Спереди и сзади он не достигает уровня переднего и заднего краев карапакса. Длина пластрона составляет 80% от длины карапакса. Ширина пластрона примерно равна его длине. Передняя и задняя доли пластрона относительно короткие, составляют около трети от длины пластрона каждая. Передняя доля пластрона треугольной формы, ее длина составляет около 60% от ширины в основании (расстояние между центрами подмышечных вырезок). Задняя доля пластрона шире передней в основании и слабее сужается кзади, ее длина составляет около 40% от ширины. Анальная вырезка хорошо развита, в форме широкого треугольника. Длина мостов составляет около 40% от длины пластрона. Эпипластроны с хорошо развитой и вытянутой вперед эпипластральной губой. Гулярный выступ округлый спереди. Эпипластральный симфиз высокий, массивный и длинный. Медиальная длина эпипластрального симфиза меньше, чем его максимальная высота. Верхнезадний отдел симфиза не нависает над передним краем энтопластрона и не образует карманоподобное углубление. Энтопластрон относительно крупный (его длина составляет более половины длины передней доли пластрона и около 1/3 ее ширины), шестиугольный, спереди сужен, а сзади широкий и округлый. Его задний край располагается каудальнее линии, соединяющей центры подмышечных вырезок. Гиопластроны короткие медиально, их длина не превышает 10% от длины пластрона. Вклад гио- и гипопластронов в длину моста примерно равный. Ксифипластроны короткие и широкие (их длина составляет 80% от ширины), с немного выпуклыми боковыми краями.
Щиткование пластрона состоит из парных гулярных, гумеральных, пекторальных, абдоминальных, феморальных и анальных щитков. Гулярные щитки в виде узких треугольников, заходят задними углами на энтопластрон примерно на треть его длины. Гумеро-пекторальная борозда прямая в медиальной части (образует прямой угол со средней линией пластрона), проходит по заднему краю энтопластрона (слева) и слегка на него заходит (справа). В латеральной части гумеро-пекторальная борозда поворачивает вперед и располагается спереди от угла аксиллярной вырезки. Пекторальные щитки очень короткие в медиальной части (около половины длины гиопластрона) и широкие в латеральной. Абдоминальные щитки длинные в медиальной части (длиннее остальных щитков). Aбдомино-феморальная борозда расположена вблизи от гипо-ксифипластрального шва в ее медиальной части и значительно отдалена от него в латеральной части. Феморальные щитки короткие медиально и более длинные вдоль свободного края задней доли пластрона. Феморо-анальная борозда почти прямая, слегка выгнута вперед. Анальные щитки маленькие, их длина по средней линии меньше половины длины ксифипластронов. Срединная борозда пластрона почти прямая, со слабым изгибом в области абдоминальных щитков.
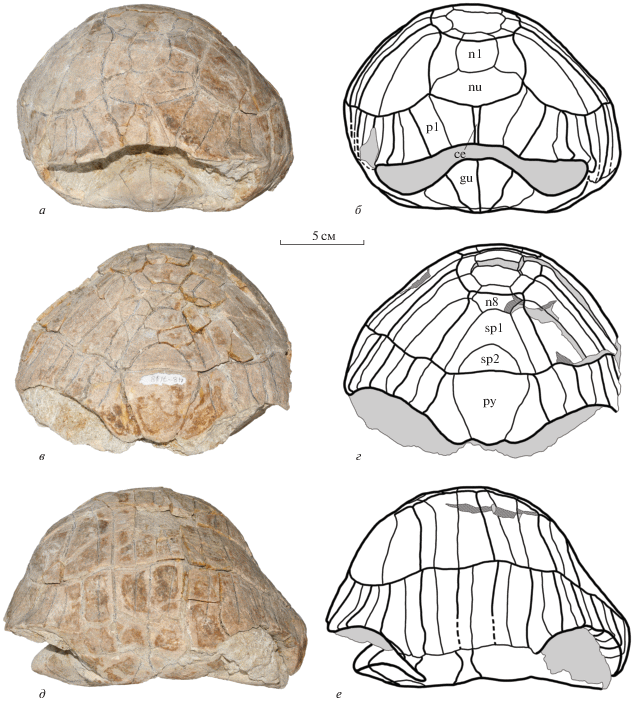
Экз. NMNHU-P, № 48-7116 (OF-1167) (рис. 5) представлен полным карапаксом, с вентральной стороны полностью скрытым породой. Отдельные фрагменты пластрона, возможно от этого экземпляра, скрыты породой под карапаксом и не позволяют дать их описание. Длина карапакса составляет около 250 мм. Карапакс высокий, не расширен сзади (наибольшая ширина на уровне V периферальных пластинок), в отличие от экз. NMNHU-P, № 48-7118. Ширина карапакса составляет около 85% от его длины. В задней части карапакс изогнут книзу, однако изгиб менее выражен, чем у экз. NMNHU-P, № 48-7118. Нухальная вырезка немного шире, чем у экз. NMNHU-P, № 48-7118. Нухальная пластинка вогнутая спереди. Формула невральных пластинок 4 < 8 > 4 < 8 > > 4 < 6 < 6–4. I невральная пластинка удлиненная, тогда как последующие невральные (кроме VIII) широкие (ширина больше длины). В отличие от экз. NMNHU-P, № 48-7118, VIII невральная маленькая, четырехугольная, с округлыми краями. Супрапигальные пластинки сходны с таковыми у экз. NMNHU-P, № 48-7118, однако I супрапигальная пластинка немного короче, чем у экз. NMNHU-P, № 48-7118. Пигальная пластинка не отличается от таковой экз. NMNHU-P, № 48-7118. Костальных пластинок семь пар, но последняя пара пластинок образована слитыми VII и VIII костальными. Периферальных пластинок 11 пар. Мостовые периферальные пластинки снизу закрыты породой, что не позволяет оценить их ширину (высоту).
Рис. 5.
Protestudo bessarabica (Riabinin, 1918), экз. NMNHU-P, № 48-7116, карапакс, местонахождение Белка, Украина; поздний миоцен, мэотис (MN 12): а, б – сверху; в, г – спереди; д, е – сзади; ж, з – сбоку; а, в, д, ж – фото; б, г, е, з – поясняющие рисунки. Обозначения: см. подписи к рис. 3, 4.
Прецентральный щиток трапециевидный, сильно расширен сзади. Висцеральная часть прецентрального щитка не видна. Центральные щитки сходны с таковыми экз. NMNHU-P, № 48-7118, однако отличаются чуть более широкими спереди I и V щитками. Передний край I щитка достигает правой и левой I периферальных пластинок. Плевральные и маргинальные щитки не отличаются от таковых экз. NMNHU-P, № 48-7118, за исключением XII маргинальных щитков, передний край которых уже (равен ширине переднего края пигальной пластинки), чем у экз. NMNHU-P, № 48-7118 (шире переднего края пигальной пластинки).
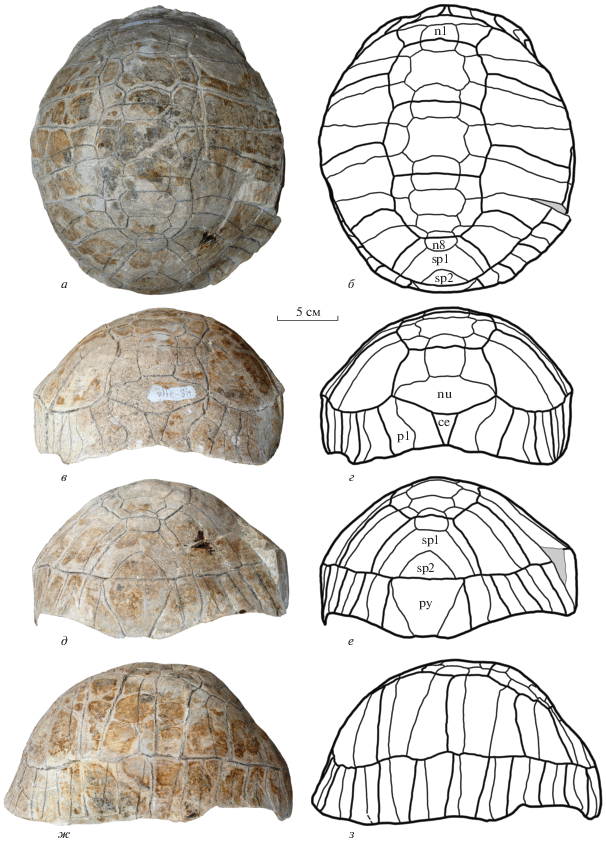
Экз. NMNHU-P, № 48-7117 (OF-1168) (рис. 6) представлен неполным пластроном сравнительно небольшого размера (предположительно, молодой особи) без левого и части правого эпипластронов, а также левого и части правого мостов. Реконструируемая длина пластрона составляла около 120 мм, ширина – около 130 мм. Передняя доля пластрона относительно широкая. Задняя доля сходна по ширине с передней долей, а ее длина составляет около половины от ширины. Анальная вырезка хорошо развита, но заметно уже, чем у экз. NMNHU-P, № 48-7118. Энтопластрон крупный, шестиугольный, не закруглен сзади, по сравнению с экз. № 48-7118. Гиопластроны немного длиннее медиально, чем у экз. NMNHU-P, № 48-7118 (15% от длины пластрона). Хотя мостовые части пластрона сохранились не полностью, видно, что вклад гио- и гипопластронов в длину моста был примерно равный. Ксифипластроны широкие, их длина составляет 70% от ширины.
Рис. 6.
Protestudo bessarabica (Riabinin, 1918), экз. NMNHU-P, № 48-7117, неполный пластрон сверху; местонахождение Белка, Украина; поздний миоцен, мэотис (MN 12): а – фото; б – поясняющий рисунок. Обозначения: см. подписи к рис. 3, 4.
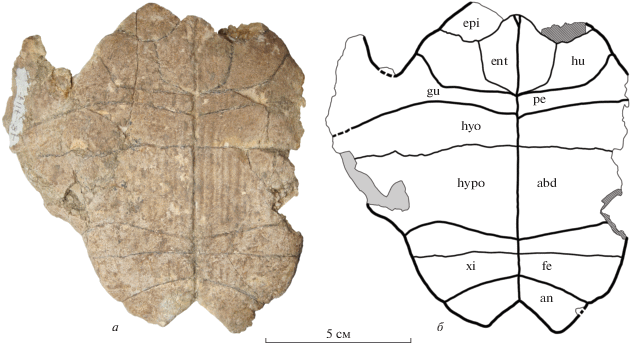
Щитки пластрона представлены неполно, отсутствуют гулярные, передние части гумеральных, латеральные части левого пекторального и абдоминальных щитков. Гумеро-пекторальная борозда проходит по заднему краю энтопластрона, но не заходит на него. В латеральной части гумеро-пекторальная борозда изогнута вперед и больше смещена вперед, в отличие от экз. NMNHU-P, № 48‑7118. Пекторальные щитки очень короткие в медиальной части (около трети длины гиопластрона), но длиннее чем у экз. NMNHU-P, № 48-7118. Aбдомино-феморальная борозда расположена немного дальше от гипо-ксифипластрального шва в ее медиальной части, по сравнению с экз. NMNHU-P, № 48-7118. Анальные щитки относительно маленькие, их длина по средней линии составляет около половины от длины ксифипластронов. Срединная борозда прямая.
Размеры см. табл. 1.
Таблица 1.
Промеры панцирей (в мм) Protestudo bessarabica (Riabinin, 1918) из местонахождения Белка, Украина, поздний миоцен, мэотис (MN 12). Обозначения: прочерк – измерение невозможно; d – правый элемент панциря, s – левый элемент панциря
| Элементы панциря и их промеры | Экз. NMNHU-P, № 48-7118 |
Экз. NMNHU-P, № 48-7116 |
Экз. NMNHU-P, № 48-7117 |
|---|---|---|---|
| Карапакс (длина/ширина/высота) | 189/172/117 | ~250/~215/– | – |
| Нухальная пластинка (длина/ширина наибольшая/ширина по переднему краю/ширина по заднему краю) | ~40/50.3/30.4/20 | 47/64.4/41.5/19.5 | – |
| Невральные пластинки (длина/ширина) | |||
| I | 30.2/22.8 | 37/26.8 | – |
| II | 23.4/33.5 | 26.6/43.6 | – |
| III | 21.2/29 | 25/32 | – |
| IV | 23/31.7 | 25.7/45 | – |
| V | 18.5/28 | 23/33 | – |
| VI | 16.5/30 | 19.3/38 | – |
| VII | 28.8/16.6 | 14/30 | – |
| VIII | 17/28 | 18.8/22.8 | – |
| I cупрапигальная пластинка (длина/ ширина по переднему краю/ширина по заднему краю) | 42.2/17/52 | 38/18.2/63.5 | – |
| II cупрапигальная пластинка (длина/ширина) | 17.5/34.8 | 20.5/34 | – |
| Пигальная пластинка (длина/ширина по переднему краю/ширина по заднему краю) | 41/41.8/17.4 | 34.8/43.8/14.2 | – |
| Костальные пластинки (длина/медиальная ширина/дистальная ширина) | |||
| I | 67.7/26.3/57.5 d | 72/32.7/47.6 s | – |
| II | 73.8/15/30.5 d | 80/21.2/32.5 s | – |
| III | 84.5/19.4/9.7 d | 85.8/20.7/~13 s | – |
| IV | 80.5/13.5/26.6 d | 79.2/16.5/36.7 s | – |
| V | 78.0/16.5/11.2 d | 73.3/17/15.5 s | – |
| VI | 67.2/13/27.6 d | 57.3/14.5/32.5 s | – |
| VII | 55.8/13/12.5 d | 39.3/9.8/34.8 s | – |
| VIII | 41.5/0.9/32 | – | – |
| Периферальные пластинки (ширина по свободному краю/ширина по медиальному краю/длина по заднему краю) | |||
| I | 32.2/23.8/31.2 s | 33.7/24.3/34 | – |
| II | 25/12.5/26.4 s | 29.3/13.9/33.8 | – |
| III | 23/17.5/33 s | 22/19.7/– | – |
| IV | 19.5/18/47.5 s | 22.2/22.4/– | – |
| V | 19/20.4/46 s | 20.8/25.5/– | – |
| VI | 16.6/20.6/~50 s | 20/~22/– | – |
| VII | 19.2/~19/40.8 s | 20.8/22.3/– | – |
| VIII | 21.2/15.6/35.5 s | 23.8/23.3/30.3 | – |
| IX | 20/19.4/26.2 s | 20/7/20.3/28 | – |
| X | 21.7/20.8/~26 s | 24.3/18/30 | – |
| XI | 40.3/15.4/~40 s | 30.7/11.3/38.4 | – |
| Прецентральный щиток (длина/ширина по переднему краю/ширина по заднему краю) | 21.7/–/– | 21.3/2.8/15.8 | – |
| Центральные щитки (длина/ширина) | |||
| I | 41.7/54.4 | 48.3/67.5 | – |
| II | 42.5/52.8 | 53.7/67.6 | – |
| III | 36.3/55.8 | 48.9/72.8 | – |
| IV | 46.7/50.5 | 49.8/60 | – |
| V | 53.5/67.2 | 61.6/68.7 | – |
| Плевральные щитки (длина/медиальная ширина/дистальная ширина) | |||
| I | 60.7/16.2/60 s | 65.5/24.8/65.6 s | – |
| II | 67.7/38.3/41.2 s | 65.2/49.3/48.9 s | – |
| III | ~55/~39/41 s | 53/45.7/50.3 s | – |
| IV | 45.7/22.6/42.5 s | 45/27.7/39.6 s | – |
| Маргинальные щитки (ширина по свободному краю/медиальная ширина/длина по заднему краю) | |||
| I | ~27/15.7/34.3 s | 32.3/14/38.2 s | – |
| II | 31.5/15.5/~26 s | 32.6/16.9/34.5 s | – |
| III | 24.6/15.8/26.2 s | 27/19/~34 s | – |
| IV | 24/21.6/45.6 s | 23.3/23.4/– s | – |
| V | 21.5/20/44 s | 23.4/28.4/– s | – |
| VI | 21.6/~19/46.2 s | 23.7/25/– s | – |
| VII | –/–/42.7 s | 20.7/23/– s | – |
| VIII | 16.6/21.2/37.6 s | 20.6/15.8/– s | – |
| IX | 16.5/21.1/27.5 s | 23.2/25.3/– s | – |
| X | ~20/~20/25 s | 21/18.3/– s | – |
| XI | ~25/~25/26.3 s | 25/20.8/– s | – |
| Пластрон (длина/ширина) | 135.2/149.3 | – | –/~100 |
| Мосты пластрона (медиальная длина) | 65.8 | – | 45.5 |
| Эпипластроны (медиальная длина/длина по заднему краю/толщина эпипластрального симфиза) | 20/17.5/29 | – | –/13.6/~15 |
| Энтопластрон (длина/ширина) | 35.7/33.6 | – | 23.6/22.5 |
| Гиопластрон (медиальная длина) | 16.2 | – | 14.4 |
| Гипопластрон (медиальная длина) | 42 | – | 27.4 |
| Ксифипластрон (медиальная длина) | 22.6 | – | 15.1 |
| Гумеральные щитки (медиальная длина) | 25.3 | – | – |
| Пекторальные щитки (медиальная длина) | 7.7 | – | 5.7 |
| Абдоминальные щитки (медиальная длина) | 47.3 | – | 35.7 |
| Феморальные щитки (медиальная длина) | 17.1 | – | 10.8 |
| Анальные щитки (медиальная длина) | 8.7 | – | 8.3 |
Описываемые материалы из местонахождения Белка отнесены к роду Protestudo на основе следующих признаков: нухальная вырезка хорошо развита; невральных пластинок восемь; эпипластральная губа хорошо развита и выступает вперед; эпипластроны очень массивные; медиальная длина эпипластрального симфиза меньше, чем его максимальная высота; медиальная часть абдомино-феморальной борозды располагается вблизи гипо-ксифипластрального шва, но не достигает его. Описываемые материалы отнесены к виду P. bessarabica по наличию хорошо развитой нухальной вырезки и сильно вытянутого вперед, но не остроконечного гулярного выступа (Данилов и др., 2017). От других Protestudo этот вид отличается формой гулярного выступа. Помимо этого, P. bessarabica отличается от европейских Protestudo: от P. csakvarensis (Szalai, 1934) из сармата Венгрии и Молдовы – наличием прецентрального щитка; от P. moldavica Chkhikvadze et Lungu, 1979 из среднего сармата Молдовы – прямой феморо-анальной бороздой; от P. chisinauensis Redkozubov, 2007 из среднего сармата Молдовы – более длинными эпипластронами и широкой второй супрапигальной пластинкой.
Материалы из Белки являются одними из наиболее полных среди опубликованных материалов по P. bessarabica и дополняют данные по морфологии и изменчивости панциря этого вида. Экземпляры NMNHU-P, №№ 48-7116 и 48-7118 имеют по восемь невральных пластинок. Из них последняя (VIII) невральная пластинка может быть шестиугольной (NMNHU-P, № 48-7118) или четырехугольной (NMNHU-P, № 48-7116), тогда как у всех остальных P. bessarabica последняя пластинка шестиугольная. У большинства других описанных экземпляров P. bessarabica, за исключением экз. из Чобручей (экз. ГЛЯУ, б/н), невральных пластинок семь. У экз. из Чобручей (экз. ГЛЯУ, б/н) невральных пластинок восемь и VIII невральная пластинка шестиугольная. Необходимо отметить, что у экз. из Чимишлии (экз. ГКМ, № 8027) в описании указано наличие восьми невральных пластинок с формулой последних четырех пластинок 6 < 6 < 6 < 6, тогда как на рисунке их показано семь и формула последних пластинок 5 < 6 < 6 (Хозацкий, Тарабукин, 1970, рис. 1). Эти и другие вариации строения панциря P. bessarabica перечислены в табл. 2.
Таблица 2.
Вариации строения панциря Protestudo bessarabica (Riabinin, 1918)
| Признаки | Экз. NMNHU-P, № 48-7118 |
Экз. NMNHU-P, № 48-7116 |
Экз. NMNHU-P, № 48-7117 | Экз. ПИН, № 1451, Тудорово | Экз. ГЛЯУ, без №, Чобручи | Экз. ГКМ, № 8027, Чимишлия | Экз. ТКМ, № КЛ 8659/ЕИ-I, Недвиговка |
|---|---|---|---|---|---|---|---|
| Количество невральных пластинок | 8 | 8 | – | 7 | 8 | 7 | – |
| Формула невральных пластинок | 4 < 8 >4 < 8 > 4 < < 6 < 6–6 | 4 < 8 > 4 < 8 > > 4 < 6 < 6–4 | – | 4 < 8 > 4 < 8 > 4 < < 6 < 6 | 4 < 8 > 4 < 8 > 4 < < 6 < 6–6 | 4 < 8 > 4 < 8 > 5 < < 6 < 6 | – |
| I периферальные пластинки (отношение длины медиального края к длине свободного края) | 0.7 | 0.7 | – | 0.6 | 0.8 | 0.5 | – |
| Форма прецентрального щитка | очень узкий трапециевидный, слабо расширен сзади | нормально развитый трапециевидный, сильно расширен сзади | – | нормально развитый трапециевидный, сильно расширен сзади | нормально развитый трапециевидный, сильно расширен сзади | нормально развитый трапециевидный, сильно расширен сзади | – |
| Анальная вырезка | широкая | – | узкая | широкая | узкая | широкая | широкая |
| Срединная борозда | слабоизвилистая | – | прямая | прямая | слабоизвилистая | прямая | прямая |
Помимо этого, у P. bessarabica из Белки наблюдается изменчивость формы прецентрального щитка, который может быть более (экз. NMNHU-P, № 48-7116) или менее (экз. NMNHU-P, № 48-7118) широким сзади. При этом у остальных описанных экземпляров P. bessarabica прецентральный щиток заметно расширен сзади.
Различия в высоте панциря, наблюдаемые у описанных экземпляров (высокий у экз. NMNHU-P, № 48-7118 и низкий у экз. NMNHU-P, № 48-7116) подтверждают изменчивость P. bessarabica по этому признаку, ранее указанную и для других экземпляров (см., напр., Рябинин, 1918). Первые периферальные пластинки у экземпляров из Белки и у большинства остальных P. bessarabica четырехугольные, слабоклиновидные (отношение длины медиального края к длине свободного края пластинки – 0.7 у экз. NMNHU-P, №№ 48-7118 и 48-7116), тогда как у экз. из Чимишлии (Хозацкий, Тарабукин, 1970) они демонстрируют ярко выраженную клиновидность (то же отношение – 0.5). Анальная вырезка может быть более (NMNHU-P, № 48-7118) или менее (NMNHU-P, № 48-7117) широкой. Срединная борозда пластрона у P. bessarabica из Белки может быть прямой (экз. NMNHU-P, № 48-7118) или слабоизвилистой (экз. NMNHU-P, № 48-7117). Для остальных P. bessarabica характерна прямая срединная борозда, хотя есть указание на ее слабую извилистость (Хозацкий, Редкозубов, 1989).
Среди других проявлений изменчивости P. bessarabica следует указать изменчивость в форме супрапигальных и пигальной пластинок, I центрального щитка и положении гумеро-пекторальной борозды. Супрапигальные пластинки у большинства P. bessarabica имеют обычную для тестудинид форму – крупная трапециевидная I супрапигальная и маленькая треугольная II супрапигальная пластинки. Однако у экз. из Чобручей (экз. ГЛЯУ, б/н) обе супрапигальные пластинки трапециевидные, из них I пластинка маленькая, а II – более крупная. Такое строение супрапигальных пластинок известно у Agrionemys (Protagrionemys) kegenica (Khosatzky in Bazhanov et Pigulevskiy, 1955) из среднего–позднего миоцена Казахстана и, возможно, может встречаться и у Protestudo.
Пигальная пластинка у большинства P. bessarabica трапециевидная четырехугольная, однако у экз. из Чобручей (экз. ГЛЯУ, б/н) она имеет скошенные переднебоковые стороны (Macarovici, 1930). Такое строение характерно для представителей Chersine Merrem, 1820 (=Eurotestudo Lapparent de Broin et al., 2006), однако встречается и у других тестудинид, например, Protestudo darewskii Chkhikvadze, 1971 (Хозацкий, Нармандах, 1975; Lapparent de Broin et al., 2006).
Первый позвоночный щиток у P. bessarabica немного варьирует по ширине, достигает (экз. ГКМ, № 8027, из Чимишлии) или не достигает (экз. ПИН, № 1451, экз. ГЛЯУ, б/н, из Чобручей) I периферальной пластинки. Гумеро-пекторальная борозда у большинства P. bessarabica проходит по заднему краю энтопластрона, но в некоторых случаях отдалена от него (экз. ГКМ, № 8027, из Чимишлии и экз. ГЛЯУ, б/н, из Чобручей).
Из перечисленных вариаций различия по высоте панциря, форме гулярного выступа и анальной вырезки, по-видимому, связаны с половым диморфизмом, тогда как остальные являются вариантами индивидуальной изменчивости (см., напр., McRae et al., 1981; Kaddour et al., 2008).
Экземпляры P. bessarabica из местонахождения Белка являются первыми описанными и изображенными материалами по этому виду из Украины. Находки черепах этого вида, упоминавшиеся из других местонахождений Украины [Гребеники, Новая Эметовка 2 (=Костев овраг), Новоелизаветовка, Новоукраинка, Черевичное; Данилов и др., 2017], остаются неописанными. P. bessarabica была, по-видимому, широко распространенной в Восточной Европе в достаточно узком временном интервале (конец сармата – мэотис). Здесь необходимо отметить, что возраст местонахождения Новоукраинка (далее Новоукраинка 1 – отложения с костями крупных млекопитающих, птиц и рептилий; Новоукраинка 2 – местонахождения мелких млекопитающих; см. Топачевский и др., 1998) первоначально был определен ранним мэотисом (Короткевич, 1961). При упоминании находок черепах из этого местонахождения их возраст также определили как мэотический (Чхиквадзе, 1983). Позднее Короткевич (1972, 1988) на основании общности видового состава крупных млекопитающих из Гребеников, Старой Кубанки и Новоукраинки 1 и его отличиям от видового состава крупных млекопитающих раннего мэотиса отнесла все указанные местонахождения к позднему сармату. Учитывая, что находки P. bessarabica приурочены к костеносным горизонтам Новоукраинки 1, не вызывает сомнения, что и остатки черепах имеют позднесарматский возраст. Таким образом, P. bessarabica из Новоукраинки представляет особый интерес, являясь наиболее древней находкой этих черепах. Все остальные находки P. bessarabica – мэотического возраста. Указание на возможно сарматский возраст местонахождения Недвиговка (Ростовская обл., Россия) не подтверждается (Тесаков А.С., Титов В.В., устн. сообщ.). Хорошая сохранность крупных и массивных элементов панциря, а также особенности морфологии P. bessarabica позволяют легко диагностировать остатки этих черепах в комплексах позвоночных, делая P. bessarabica важным таксоном для биостратиграфических корреляций.
Список литературы
Данилов И.Г., Сыромятникова Е.В., Суханов В.Б. Подкласс Testudinata // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 4 / Ред. Лопатин А.В., Зеленков Н.В. М.: ГЕОС, 2017. С. 27–395.
Короткевич О.Л. До вивчення гіпаріонової фауни долини р. Куяльника // Зб. праць Зоол. музею. 1961. № 130. С. 122–128.
Короткевич О.Л. Основные этапы развития позднемиоценовой фауны копытных и хоботных Северного Причерноморья // Вестн. зоол. 1972. № 4. С. 19–34.
Короткевич Е.Л. История формирования гиппарионовой фауны Восточной Европы. Киев: Наук. думка, 1988. 160 с.
Крахмальная Т.В. Гиппарионовая фауна древнего мэотиса Северного Причерноморья. Киев: Наук. думка, 1996. 224 с.
Крахмальная Т.В. История изучения, стратиграфическое положение и состав фаунистических комплексов крупных млекопитающих позднего миоцена Украины // Біостратиграф. основи побудови стратиграф. схем фанерозою України. Київ: Ин-т геол. наук НАН України, 2008. С. 290–296.
Топачевский В.А., Несин В.А., Топачевский И.В. Биозональная микротериологическая схема (стратиграфическое распределение мелких млекопитающих – Insectivora, Lagomorpha, Rodentia) неогена северной части Восточного Паратетиса // Вестн. зоол. 1998. Т. 32. № 1-2. С. 76–87.
Рябинин А.Н. О черепахах из меотических отложений Бессарабии // Тр. Геол. минерал. музея Росс. АН. 1918. Т. 1. Вып. 1. С. 1–16.
Хозацкий Л.И., Нармандах П. Миоценовая черепаха из Западной Монголии // Тр. Совм. Сов.-Монгол. палеонтол. экспед. 1975. Вып. 2. С. 110–121.
Хозацкий Л.И., Редкозубов О.И. Неогеновые черепахи Молдавии. Кишинэу: Штиинца, 1989. 94 с.
Хозацкий Л.И., Тарабукин Б.А. О панцирях мэотических черепах, найденных в МССР // Уч. зап. Тираспол. гос. пед. ин-та. 1970. Вып. 17. С. 64–70.
Хозацкий Л.И., Тофан В.Е. Прошлое и современное состояние герпетофауны Молдавии // Уч. зап. Тираспол. гос. пед. ин-та. 1970. Вып. 20. С. 157–181
Чхиквадзе В.М. О происхождении современных сухопутных черепах Палеарктики // Сообщ. АН Груз. ССР. 1970. Т. 57. № 1. С. 245–247.
Чхиквадзе В.М. Ископаемые черепахи Кавказа и Северного Причерноморья. Тбилиси: Мецниереба, 1983. 149 с.
Чхиквадзе В.М. Неогеновые черепахи СССР. Тбилиси: Мецниереба, 1989. 104 с.
Danilov I.G. Die fossilen Schildkröten Europas // Handbuch der Reptilien und Amphibien Europas. Bd. 3/IIIB: Schildkroten II / Hrsg. Aula: U. Fritz. Wiebelsheim, 2005. P. 329–441.
Gnetneva A.N., Chkhikvadze V.M., Syromyatnikova E.V., Danilov I.G. Tortoises of the genus Agrionemys: phylogenetic position, systematics and fossil record // Turtle Evolution Symposium. Tübingen: Scidinge Hall Verlag, 2018. P. 41–47.
Kaddour K.B., Mouden E.H.E., Slimani T. et al. Sexual dimorphism in the Greek tortoise: a test of the body shape hypothesis // Chelon. Conserv. Biol. 2008. V. 7. № 1. P. 21–27.
Krakhmalnaya T. Hipparions of the Northern Black Sea coast area (Ukraine and Moldova): species composition and stratigraphic distribution // Acta Zool. Cracov. 1996. V. 39. № 1. P. 261–267.
Krakhmalnaya T.V. Proboscideans and ungulates of late Miocene fauna of Ukraine // 6th meeting of the European assoc. of vertebrate palaeontologists / Ed. Krempaská Z. Spišská Nová Ves: Museum Spisa, 2008. P. 51–55.
Lapparent de Broin F. de. The European turtle fauna from the Triassic to the Present // Dumerilia. 2001. V. 4. P. 155–217.
Lapparent de Broin F. de, Bour R., Perälä J. Morphological definition of Eurotestudo (Testudinidae, Chelonii): first part // Ann. Paléontol. 2006. V. 92. P. 255–304.
Macarovici N. Sur une Testudo bessarabica Riab. trouvée dans les couches méotiques de Cioburciu (Dép. Tighina-Roumanie) // Bull. Sect. Sci. Acad. Roum. 1930. № 1/2. P. 1–5.
Macarovici N., Vancea S. Sur les restes de tortues de la faune de Mallisteni de la Moldavie meridionale // Ann. Sci. Univ. Jasi. 2. Sci. Natur. 1960. T. 6. P. 377–386.
McRae W.A., Landers J.L., Cleveland G.D. Sexual dimorphism in the gopher tortoise (Gopherus polyphemus) // Herpetologica. 1981. V. 34. P. 46–57.
Perälä J. The genus Testudo (Testudines: Testudinidae): phylogenetic inferences // Chelonii. 2002. V. 3. P. 32–39.
Rhodin A.G.J., van Dijk P.P., Parham J.F. Turtles of the World: annotated checklist of taxonomy and synonymy // Conservation Biology of Freshwater Turtles and Tortoises: A Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle Specialist Group / Eds. Rhodin A.G.J., Pritchard P.C.H., van Dijk P.P. et al. Chelonian Res. Monogr., 2008. № 5. P. 000.1–000.38.
Semenov Yu.A. Stratigraphic distribution of terrestrial carnivores in the Vallesian and Turolian of Ukraine // Beitr. Paläontol. 2001. № 136. P. 139–144.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал