

Палеонтологический журнал, 2019, № 6, стр. 79-90
Первая находка двоякодышащей рыбы (Dipnoi, Sarcopterygii) в фаменских отложениях (верхний девон) Тверской области
О. А. Лебедев a, *, Н. И. Крупина b, **, В. В. Линкевич c, ***
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Научно-учебный музей Землеведения, МГУ им. М.В. Ломоносова
Москва, Россия
c Андреапольский районный краеведческий музей им. Э.Э. Шимкевича
Андреаполь, Россия
* E-mail: elops12@yandex.ru
** E-mail: n.krupina@mail.ru
*** E-mail: linkevichvalerijj@rambler.ru
Поступила в редакцию 08.11.2018
После доработки 27.12.2018
Принята к публикации 29.03.2019
Аннотация
Описывается нижняя челюсть двоякодышащей рыбы Anchidipterus dariae Krupina, gen. et sp. nov. (Dipteridae, Dipnoi) из фаменского (верхний девон) местонахождения Билово (Тверская обл., Торопецкий р-н). Новый род и вид характеризуется коротким и широким симфизным отделом, большим углом расхождения гребней зубных пластинок и ростролатеральной ориентировкой мезиальных гребней. Платисимфизная конструкция нижней челюсти в сочетании с относительно небольшими, широко расставленными зубными пластинками позволяют предположить, что анхидиптеры питались слабо армированными или мягкотелыми, малоподвижными беспозвоночными и занимали в трофической структуре сообщества нишу неспециализированных омнифагов.
ВВЕДЕНИЕ
Двоякодышащие рыбы – склерофаги, омнифаги и даже хищники – играли важную роль в трофических цепях девонских сообществ позвоночных, но грацильность черепа и, чаще всего, слабое окостенение его костей приводили к тому, что остатки дипной в ископаемом состоянии чаще всего представлены только крепкими изолированными зубными пластинками. Каждая новая находка даже фрагмента черепа, в особенности, если она ассоциирована с зубными пластинками, несет важную информацию как о морфологии рыбы, так и о характере ее питания и положении в трофических структурах сообществ.
Местонахождение Билово – одно из немногочисленных фаменских местонахождений в восточной части Главного девонского поля. Оно богато остатками как беспозвоночных, так и разнообразных позвоночных животных. Впервые остатки рыб из двух аналогичных разрезов на левом (у дер. Билово) и правом берегу (под хут. Билово) р. Малый Тудер (приток р. Куньи, бассейн р. Ловати) были собраны Б.П. Марковским (Геолком) в 1929–1930 гг., а в 1930–1931 гг. – экспедицией Девонской литологической партии Ленинградского геолого-разведочного треста под руководством Р.Ф. Геккера [хранятся в колл. Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН)].
В работе Р.Ф. Геккера и М.Ф. Филипповой (1935) Д.В. Обручевым приведены следующие определения рыб из “сводного” биловского разреза: Bothriolepis n. sp., Bothriolepis sp., Coccosteus sp., Holoptychius flemingi Agassiz, 1839, Holoptychius sp., Osteolepis n. sp. Только в 1958 г. палеоихтиологический отряд под руководством этого автора посетил Билово и собрал небольшое количество остатков антиарх и остеолепиформов.
Э.И. Воробьева (1977) описала отико-окципитальный блок остеолепиформной рипидистии Megapomus heckeri Vorobyeva, 1977 по материалу из этих коллекций. На этикетке упомянуто, что экземпляр обнаружен Геккером в 1930 г. в слое 4 (по описанию Геккера и др., 1935, с. 75). Э. Лукшевич (Lukševičs, 2001) описал новый вид антиарха Bothriolepis heckeri Lukševičs, 2001, основанный на остатках из этих же сборов и из того же слоя. Находок двоякодышащих рыб в XX в. сделано не было.
Заново привлекли внимание палеоихтиологов к этому местонахождению регулярные полевые работы (с 2011 г.) по мониторингу некоторых береговых обнажений малых рек – притоков р. Ловати – в Андреапольском и Торопецком районах Тверской обл., проводимые сотрудниками Андреапольского районного краеведческого музея им. Э.Э. Шимкевича (КМА). Основная цель этих работ – сбор остатков ископаемых девонских позвоночных и беспозвоночных животных.
В коллекции из местонахождения Билово, хранящейся в КМА, С.В. Молошников (Музей землеведения МГУ им. М.В. Ломоносова) и В.В. Линкевич (КМА) определили остатки следующих позвоночных: Bothriolepis heckeri Lukševičs, 2001, Bothriolepis sp., Dunkleosteoidea indet., Holoptychius sp., Osteolepididae indet., Dipteridae indet. (Молошников, Линкевич, 2017а, б). В этих двух работах описаны новые находки антиарх; вид Bothriolepis heckeri отнесен к ранее установленному роду Livnolepis Moloshnikov, 2008.
С 2011 по 2015 гг. на биловских разрезах удалось собрать несколько зубных пластинок двоякодышащих рыб, а в 2016 г. была найдена нижняя челюсть с хорошо сохранившимися зубными пластинками. Находка сделана сотрудником Торопецкой биостанции “Чистый лес” (Тверская обл.) Д.Г. Бондарь.
Эта нижняя челюсть, описываемая ниже, отнесена к новому роду и виду Anchidipterus dariae Krupina, gen. et sp. nov. Новый таксон отличается от других членов семейства Dipteridae Owen, 1846 относительно короткой и широкой нижней челюстью, симфизной частью своеобразной формы, а также строением зубных пластинок.
ГЕОЛОГИЧЕСКОЕ СТРОЕНИЕ ВМЕЩАЮЩИХ ОТЛОЖЕНИЙ
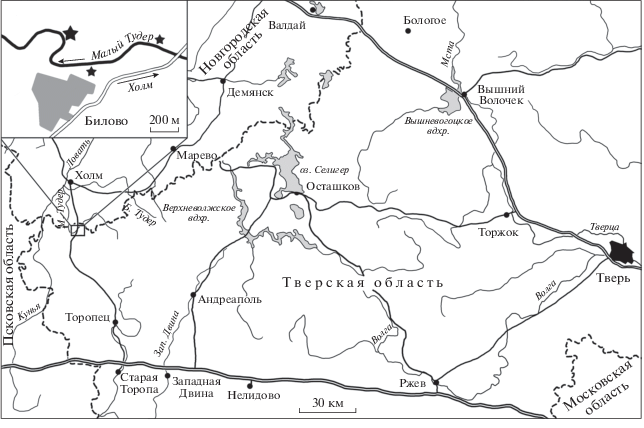
Местонахождение Билово располагается на северо-западе Тверской области, вблизи ее границы с Новгородской. Местонахождение объединяет по крайней мере три разреза, расположенных на крутой излучине р. Малый Тудер и выше нее по обоим берегам около д. Билово (рис. 1). Здесь вскрываются пачки чередующихся песчаных, карбонатных и глинисто-песчаных пород тудерской (сопоставляемой с елецким горизонтом), биловской (с лебедянским горизонтом) и нижней части льнянской (с оптуховским горизонтом) свит верхней части нижнего – среднего фамена (Вербицкий и др., 2012).
Рис. 1.
Расположение местонахождения Билово. На основном поле – западная часть Тверской области, на врезке – положение местонахождений относительно дер. Билово. Большой звездочкой отмечено обнажение, откуда происходит описываемый материал, маленькими – два остальных разреза.
Тудерская свита распространена в юго-восточной и восточной частях Изборско-Ильменской структурной зоны и выходит на дочетвертичную поверхность вдоль западной границы своего распространения (Вербицкий и др., 2012).
Песчаники тудерской свиты несогласно перекрываются известняками биловской свиты. В биловском разрезе нижняя граница последней принимается по подошве карбонатной пачки, налегающей на желто-бурые песчаники (Вербицкий и др., 2012). Сама биловская свита представлена переслаивающимися пестроцветными мергелями, известковистыми глинами, светло-серыми и зеленовато-серыми песчанистыми мелкозернистыми известняками.
Учитывая присутствие карбонатных прослоев внутри в целом терригенной тудерской свиты (Саммет, 1973), расчленение разреза в Билове не представляется очевидным. В разрезе, описанном Геккером и Филипповой (1935), брахиоподы Cyrtospirifer (Spirifer) cf. lebedjanicus Nalivkin, 1947, датирующие лебедянский горизонт, появляются только со слоя 7. Все нижележащие карбонатные слои, таким образом, должны быть отнесены к елецкому горизонту, что противоречит принимаемому ныне уровню кровли тудерской свиты по литологическим данным (Вербицкий и др., 2012). При этом уже из слоя 4 в нижней части разреза известны Megapomus heckeri и Bothriolepis heckeri, считавшиеся в соответствующих публикациях (Воробьева, 1977; Lukševičs, 2001) происходящими из биловской свиты. Найденный в осыпи экземпляр, описываемый в настоящей работе, может происходить из костеносных горизонтов как биловской, так и тудерской свит, то есть, иметь как елецкий, так и лебедянский возраст. Для уточнения этого вопроса необходимы продолжение исследований разреза и сбор новых органических остатков. Льнянский возраст описываемых экземпляров маловероятен, поскольку разрез льнянской свиты не карбонатный, а песчанисто-глинистый. Во всяком случае, дальнейшее сопоставление этих свит бассейна Ловати с горизонтами Центрального девонского поля требует уточнения и проверки, в особенности, с использованием палеонтологического материала.
Определения позвоночных в палеонтологической коллекции КМА, сделанные Молошниковым и Линкевичем (2017а, б) и перечисленные выше, уточнены и дополнены по материалам новых сборов. А.О. Иванов (СПбГУ) переопределил остатки Dunkleosteoidea indet. как Dunkleosteus sp. (личн. сообщ., 2016). Просмотр коллекции первым автором этой работы показал также присутствие акантод Devononchus (?) sp., фрагментов костей панциря Bothriolepis sp. и чешуй Holoptychius sp. из самого высокого, льнянского уровня (верхний фамен). Более ясную и полную информацию о послойном таксономическом составе рыб может дать выделение микроостатков позвоночных из старых, привязанных к разрезу Геккера образцов, и сравнение с полученными в результате новых сборов, а также переописание стратотипического разреза.
СИСТЕМАТИКА
ПОДКЛАСС SARCOPTERYGII
ОТРЯД DIPNOIFORMES
СЕМЕЙСТВО DIPTERIDAE OWEN, 1846
Род Anchidipterus Krupina, gen. nov.
Название рода от ἀνχῐ древнегреч. – сходный, близкий к роду Dipterus.
Типовой вид – Anchidipterus dariae Krupina, sp. nov.
Диагноз. Длина симфизной пластины нижней челюсти вдвое меньше ее ширины, так же как и симфизной медиальной впадины. Угол расхождения ветвей челюсти примерно 60°. Апекс преартикулярной зубной пластинки развернут дорсально. Общий угол пластинок составляет 120°, угол расхождения мезиального-латерального рядов – 110°. Мезиальный гребень пластинки короткий, достигает мезиально заднего угла dentale, ориентирован ростролатерально.
Видовой состав. Типовой вид.
Сравнение. Н.И. Крупина (2004) включила в состав семейства Dipteridae Owen, 1846 следующие роды: Dipterus Sedgwick et Murchison, 1828, Rhinodipterus Gross, 1956, Orlovichthys Krupina, 1980, Tarachnomylax Barwick, Campbell et Mark-Kurik, 1997 и, условно, Grossipterus Obruchev, 1964. Здесь мы также включаем в состав семейства описанные позднее роды Harajicadipterus Clement, 2009 и Sinodipterus Qiao et Zhu, 2009, первоначально не отнесенные авторами этих родов ни к одному семейству. При этом у представителей только трех первых родов известно строение нижних челюстей, поэтому сравнение нашего материала проводится только с ними.
От рода Grossipterus (Крупина, 2004, табл. I , фиг. 9, 10 ), условно отнесенного к подсемейству Dipterinae, новый род отличается зубными пластинками перетирающего, а не перемалывающего типа (“Dipterus – тип”).
Угол схождения ветвей нижней челюсти у Anchidipterus gen. nov. близок к наблюдаемому у Dipterus, слегка более тупой, чем у Rhinodipterus, и гораздо более тупой, чем у Orlovichthys, у которого сильно вытянуто dentale. Анхидиптер сближается с этими родами отсутствием симфизного смыкания преартикулярных зубных пластинок, но отличается от них гораздо более коротким и широким симфизным отделом, изометричными зубными пластинками с гораздо более коротким мезиальным рядом зубов на них, количеством гребней на пластинках и углом их расхождения.
В отличие от Rhinodipterus, у которого мезиальный гребень преартикулярной пластинки занимает парасагиттальное положение или направлен ростромедиально, а передний конец гребня не достигает уровня заднего угла dentale, у Anchidipterus gen. nov. мезиальный гребень ориентирован ростролатерально, а его передний конец достигает уровня заднего угла dentale, но не заходит далее рострально. У Dipterus мезиальный гребень протягивается далеко рострально.
Угол расхождения рядов преартикулярных зубных пластин у анхидиптера примерно соответствует наблюдаемому у Dipterus и близок к углу у Rhinodipterus. Апекс зубной пластинки виден с дорсальной стороны, тогда как у представителей родов Dipterus и Rhinodipterus апекс значительно развернут вентромезиально и с дорсальной стороны не виден. Не исключено, что эта особенность является не родовым (или видовым?) признаком, а отражает ювенильную стадию развития описываемого экземпляра.
Anchidipterus dariae Krupina, sp. nov.
Название вида – в честь автора находки Дарьи Бондарь.
Голотип – КМА/4213, неполная нижняя челюсть (без артикулярной области); Тверская обл., Торопецкий р-н, прав. берег р. Малый Тудер, у хут. Билово; верхний девон, нижний–средний фамен, возможно, тудерская или биловская свита.
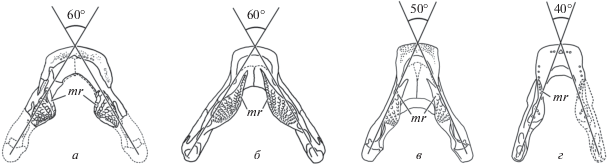
Описание (рис. 2; 3, а; 4; 5, в; 6, в). Нижняя челюсть широкая, массивная, с ветвями, сходящимися под углом примерно 60° (рис. 2, а, в; 3, а). Этот угол сходен с наблюдаемым у Dipterus (60°) (Jarvik, 1967) (рис. 3, б), слегка более тупой, чем у Rhinodipterus (50°) (Jarvik, 1967) (рис. 3, в), и гораздо более тупой, чем у длиннорылого Orlovichthys (40°) (Крупина, 1980; Krupina et al., 2001) (рис. 3, г). Судя по состоянию четко выраженных и прослеживаемых от самых ранних генераций зубов в рядах на зубных пластинках, по отсутствию следов истирания как на зубах, так и на вершинной части пластин, нижняя челюсть принадлежала ювенильной особи. Артикулярные концы обеих ветвей челюсти обломаны сразу позади задних краев зубных пластин, поэтому измерения длины ветвей (17 мм) и ширины расхождения ветвей на уровне заднего края зубных пластин (20 мм) относятся только к сохранившемуся остатку. Инфрадентальные кости срослись без швов (рис. 2, в, д), и определение соотношений spleniale и postspleniale с supraangulare не представляется возможным. Медиальная впадина для размещения передней части гипобранхиального аппарата широкая, ее длина меньше ширины в два раза.
Рис. 2.
Anchidipterus dariae Krupina, gen. et sp. nov., голотип КМА/4213, неполная нижняя челюсть: а, б – дорсально, в, г – вентрально, д, е – латерально слева. Стрелка на (д) направлена рострально. Oбозначения: af – аддукторная яма; antf – передняя борозда, ограничивающая задний край dentale; dp – зубная пластинка; idvl – вертикальная инфрадентальная пластина; ip – горизонтальная инфрадентальная пластина; jd – зубчики на dentale; lp – лабиальная ямка; Mc – меккелев хрящ; mi – медиальная вырезка на dentale; mlPra – мезиальные пластины praearticulare; oc – отверстия выхода наружных ветвей оральной ветви сенсорного канала; Pra – praearticulare с преартикулярными зубными пластинками; psl – передние (симфизные) пластины praearticulare; rd – участок зубной пластинки, сформированный репаративным дентином; slmc – симфизная пластина меккелева хряща. Масштабная линейка 10 мм.
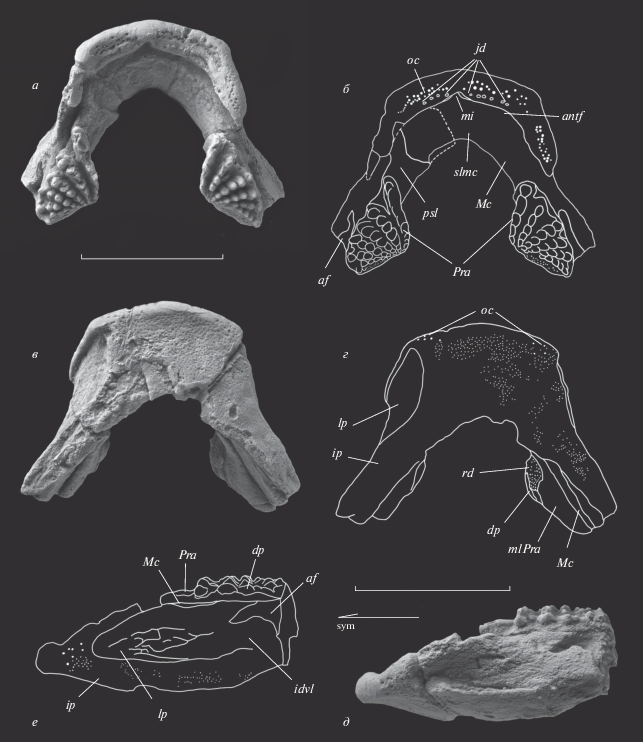
Рис. 3.
Углы расхождения ветвей нижней челюсти и ориентировка первого (мезиального) зубного ряда преартикулярных зубных пластин у различных Dipteridae: а – Anchidipterus dariae Krupina, gen. et sp. nov., задние отделы ветвей нижней челюсти реконструированы; б – Dipterus valenciennesi Sedgwick et Murchison, 1828; в – Rhinodipterus ulrichi Ørvig, 1961; г – Orlovichthys limnatis Krupina, 1980, задний отдел правой ветви нижней челюсти реконструирован [б, в – из Э. Ярвика (Jarvik, 1967); г – из Крупиной (1980)]. Oбозначениe: mr – первый (мезиальный) зубной ряд преартикулярной зубной пластины.
Меккелев хрящ (Mc, рис. 2, а–г) окостенел слабо. Длина симфизной пластины меккелева хряща (slmc) приблизительно в два раза меньше ее ширины. Эта пластина вскрыта дорсально за счет недоокостенения и/или неполной сохранности выстилавших ее прижизненно тонких и широких передних (симфизных) пластин praearticulare (psl, рис. 2, а). Помимо этой области, меккелев хрящ открывается на мезиальную поверхность челюсти между мезиальным краем инфрадентальных костей и вентральным краем praearticulare широкой полосой, клиновидно сходящей на нет в ростральном направлении, до смыкания этих костных элементов у задней части симфизной пластинки (рис. 2, г). Эта площадка служила для крепления интермандибулярной мускулатуры (Miles, 1977; Bemis, 1986). На левой ветви челюсти внутри антеровентрального отростка praearticulare располагается узкое и длинное, ориентированное косо постеродорсально/антеровентрально окно, также позволяющее наблюдать меккелев хрящ, при этом на правой ветви такое окно отсутствует. На внешней стороне челюсти меккелев хрящ открывается на латеральной стенке щелевидной, глубокой и длинной аддукторной ямы, протягивающейся вперед до уровня третьего гребня зубной пластинки (af, рис. 2, a, б, д, е). Р. Майлз (Miles, 1977) называет эту структуру с продолжением ее рострально у Chirodipterus australis Miles, 1977 супрамеккелевой ямой, Крупина и др. у Orlovichthys limnatis Krupina, 1980 (Krupina et al., 2001) – меккелевой ямкой. Здесь мы принимаем термин Э. Ярвика (Jarvik, 1967) “аддукторная яма”, поскольку гомология этой структуры, обоснованная этим автором, основана на изучении крепления челюстной мускулатуры у современного Neoceratodus, и позволяет морфологическое сравнение.
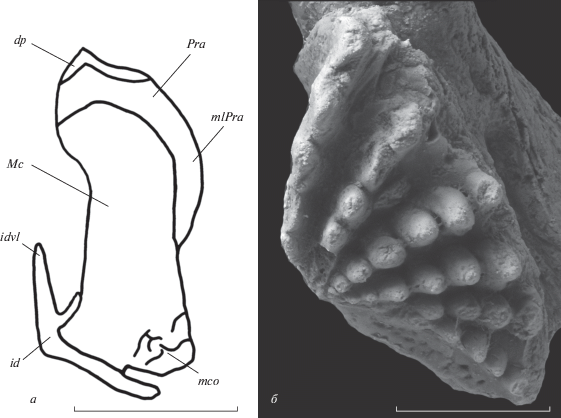
Латеральная стенка аддукторной ямы целиком образована инфрадентальными костями. Это наблюдается на пришлифованной поверхности вертикального отлома артикулярного отдела челюсти с левой стороны, (id, рис. 4, а). Вентрально дно аддукторной ямы образовано карманом между наружной пластиной инфраденталий (idvl) и их особой продольной латеральной пластиной. Дорсальный край вертикальной пластины инфрадентальных элементов, не контактируя в передней части с латеральным краем praearticulare, открывает короткий участок меккелева хряща, протягивающийся от задней оконечности передней борозды до переднего края аддукторной ямы (рис. 2, д, е). На пришлифовке, сделанной на обломанном торце левой ветви челюсти, внутри заполняющей меккелеву полость породы различаются слабо сформированные участки эндохондрального окостенения (mco, рис. 4, а).
Рис. 4.
Anchidipterus dariae Krupina, gen. et sp. nov., голотип КМА/4213: а – схема вертикального сечения левой ветви нижней челюсти на уровне заднего края зубной пластинки, по естественному обломанному краю; б – правая преартикулярная зубная пластинка. Oбозначения: dp – зубные пластины; id – инфрадентальные кости; idvl – вертикальная инфрадентальная пластина; Mc – меккелев хрящ; mco – участок эндохондрального окостенения в пределах меккелева хряща; mlPra – мезиальные пластины praearticulare; Pra – praearticulare. Масштабная линейка 3 мм.
Швы между dentale и соседними с ним костями, за исключением praearticulare, не распознаются, так же как и трасса мандибулярного сенсорного канала. Мезиальные участки орального сенсорного канала маркируются несколькими крупными порами на постеролатеральных углах dentale (oc) и одним-двумя хорошо выраженными рядами пор на дорсальной (оральной) поверхности кости, но отсутствуют в медиальной части вентральной стороны (рис. 2, а–г). В симфизной части дорсальной стороны ряды пор прерываются на коротком расстоянии, поскольку разделены симфизным бугорком. Каудально (лингвально) от этого бугорка край dentale образует симметричную узкую вырезку (mi), которая могла вмещать маленькую непарную adsymphysiale, как у Chirodipterus australis (Miles, 1977) или Ch. wildungensis Säve-Söderbergh, 1952 (Jarvik, 1967), хотя у последнего вида эта структура несколько отделена от заднего края dentale. На мальковых стадиях развития Neoceratodus в этом же положении присутствует маленькая непарная, несущая один или два зуба кость MdY (Jarvik, 1967).
С обеих сторон от вырезки на лингвальной стороне dentale располагаются короткие ряды из 4–6 мелких округлых бугорков-зубчиков (jd) в том же положении, что и описанные у Dipterus oervigi Gross, 1964 (Gross, 1964), D. valenciennesi Sedgwick et Murchison, 1828 (Jarvik, 1967) и Orlovichthys limnatis (Krupina et al., 2001), и известные у современного Neoceratodus на мальковой стадии (Jarvik, 1967; Kemp, 1995). Они могут быть гомологичны зубчикам на “лабиальных” (дентальных) зубных пластинках, описанным у мальков Andreyevichthys epitomus Krupina, 1987 (Krupina, 1992; Krupina, Reisz, 1999, с.106, рис. 2A, 4B).
Лингвальный край dentale (рис. 2, а, б) нависает над передней бороздой (antf), отделяющей его от реконструированного на иллюстрации (рис. 2, б) с левой стороны антеродорсального края мезиальных пластин praearticulare; постеромезиально от заостренных задних оконечностей dentale в дне борозды на поверхность открывается окостенение меккелева хряща.
Лабиовентральный край dentale срастается с костями инфрадентальной серии без следов шва (рис. 2, в–е). Сросшиеся без швов splenialia и postsplenialia подстилают вентральную сторону меккелева хряща единой, покрытой космином с крупными порами горизонтальной инфрадентальной пластиной (ip), а также образуют общую латеральную вертикальную пластину (idvl, рис. 2, е), которая каудально образует латеральную стенку аддукторной ямы. Выстилает ли она лабиальную ямку (lp) в ростральной части черепа, визуально определить трудно.
Praearticularia (Pra) образованы дорсальными ламинами, несущими умеренно сдвинутые назад (по сравнению с Chirodipteridae и Holodontidae) зубные пластины (dp), и передними мезиальными, частично выстилающими внутреннюю сторону ветви челюсти (mlPra, рис. 2, в, г; 4, а). Симфизные части мезиальных пластин praearticulare обломаны на правой ветви челюсти полностью, на левой – в значительной степени, поэтому проследить, как далеко вперед они простирались, и как выстилали дорсальную сторону симфизной пластинки меккелева хряща, можно только приблизительно (рис. 2, а, б).
Преартикулярные зубные пластинки (рис. 4, б) широкие. Их ширина составляет 8 мм, длина – 4 мм. На пластинках располагается шесть рядов зубчиков, четыре из которых начинаются от апекса, а два последних – от уровня третьего ряда каждой из пластинок. Общий угол пластинок составляет 120°, угол расхождения рядов – примерно 110°. Последний примерно соответствует наблюдаемому у Dipterus valenciennesi (Jarvik, 1967) и близок к углу у Rhinodipterus secans Gross, 1956 (Gross, 1956). Вершина ясно выражена, апекс расположен на уровне начала первого ряда, хорошо виден с дорсальной стороны.
В каждом из рядов насчитывается по шесть зубов, размер которых увеличивается в лабиальном направлении. Желобки между рядами глубокие и отчетливые, начинаются сразу от вершины пластинки. Наиболее глубокими, широкими и протяженными являются желобки между первым–вторым и вторым–третьим рядами. Наиболее длинный ряд – первый (мезиальный), его длина составляет 6 мм. Второй ряд составляет 0.75, третий–четвертый ряды составляют 0.625, пятый – 0.5, а последний, шестой ряд – 0.375 длины первого. Мезиальный ряд ориентирован ростролатерально, его передний конец только достигает уровня заднего угла dentale, но отделен от него бороздой (рис 2, а, б), в отличие от Rhinodipterus ulrichi Ørvig, 1961 (Jarvik, 1967) (mr) и Rh. kimberleyensis Clement, 2012 (Clement, 2012), у которых мезиальный ряд зубов занимает парасагиттальное положение или направлен ростромедиально, а передний зубчик гребня не достигает этого уровня (рис. 3, а, в). У Dipterus valenciennesi (Jarvik, 1967) передний гребень протягивается далеко рострально и отделен от заднего угла dentale широкой бороздой (рис. 3, б).
К основанию мезиального и постеро-мезиального краев зубных пластинок примыкает неширокое поле, образованное отдельными “блистерами”, состоящими из “косминовой”, несущей крупные поры, или гладкой ткани (rd, рис. 2, в, г). “Блистеры” неправильной формы, могут накладываться друг на друга, их края неровные. Это дает основания предполагать, что ткань в этих зонах попеременно резорбировалась и нарастала заново. Сходная ткань, прилегающая к зубным пластинкам, была описана Э. Уайтом (White, 1965) и Р. Дэнисоном (Denison, 1974) у Dipterus valenciennesi. К. Кэмпбелл и Р. Барвик (Campbell, Barwick, 1998), описавшие эту ткань у Adololopas moyasmithiae Campbell et Barwick, 1998, назвали ее “репаративным дентином”. В отличие от обычного космина, под верхним слоем “зубного паркета” в этой ткани отсутствует система горизонтальных каналов (den Blaauwen et al., 2005).
Размеры в мм. Длина сохранившейся части ветвей челюсти – 17.
Материал. Только голотип.
ОБСУЖДЕНИЕ
Помимо морфологического описания нового материала и установления нового таксона, задачей, решаемой в настоящей статье, является определение трофической роли двоякодышащих рыб, входящих в биловское сообщество позвоночных. Для ее решения мы постараемся выяснить наиболее вероятный способ обработки добычи анхидиптерами.
Девонские двоякодышащие рыбы обладали двумя принципиально отличавшимися типами строения зубных систем. К первому типу относятся рыбы с обширными полями сменяющихся зубчиков шагреневого типа, выстилавших кости стенок ротовой полости. Ко второму – дипнои, верхне- и нижнечелюстные кости которых несли зубные пластинки (Campbell, Barwick, 1983, 1990). Эти типы устройства зубного аппарата отражают различия в способе переработки добычи: простое схватывание и удержание с последующим заглатыванием без обработки зубными элементами в первом типе и раздавливание во втором (Schultze, 1992). Промежуточное положение занимает род Fleurantia (Graham-Smith, Westoll, 1937; Bystrow, 1944, Smith et al., 1987), у которого рассеянные дентиновые зубчики на птеригоидах сочетаются с зубами, организованными в слабо сформированные ряды.
У дипной, относящихся ко второму типу, полное срастание ветвей нижней челюсти в симфизе в сочетании с особенностями строения гленоида исключали ротацию ветвей нижней челюсти и позволяли только вертикальное смыкание пластин верхней и нижней челюсти при аддукции, как показано У. Бимисом (Bemis, 1986) для современных лепидосиренид со сходной структурой челюстей. Такая конструкция позволяла воздействовать на пищевой объект только путем его вертикального сдавливания и делала невозможным перетирание и размалывание.
Второй тип может быть, в свою очередь, подразделен на две морфо-функциональные группы (рис. 5): 1. Двоякодышащие с контактирующими или почти контактирующими передними краями в симфизной плоскости зубными пластинками давящего типа; пластинки крупные относительно длины челюсти (члены семейств Dipnorhynchidae, Chirodipteridae, Holodontidae), и 2. Двоякодышащие с далеко не доходящими до симфиза небольшими относительно общих размеров челюсти пластинками (напр., семейство Dipteridae). Во второй группе симфизной области может достигать только передняя пластина (отросток) предсочленовной кости, но не ее зубная пластинка.
Рис. 5.
Соотношения общей длины нижних челюстей, длины их ветвей, межгленоидной ширины и плеча рычага воздействия зубной пластинки на пищевой объект: а – Chirodipterus australis Miles, 1977; б – Rhinodipterus ulrichi Ørvig, 1961; в – Anchidipterus dariae Krupina, gen. et sp. nov. Задние отделы ветвей нижней челюсти, общая длина нижних челюстей, длина их ветвей и межгленоидная ширина Anchidipterus dariae Krupina, gen. et sp. nov. реконструированы [а – из Майлса (Miles, 1977); б – из Ярвика (Jarvik, 1967)]. Oбозначения: igw – межгленоидная ширина; l – длина нижних челюстей; l' – длина ветвей нижних челюстей; ll – плечо рычага воздействия на пищевой объект; lp – длина преартикулярных зубных пластинок; pc – геометрический центр преартикулярных зубных пластинок.
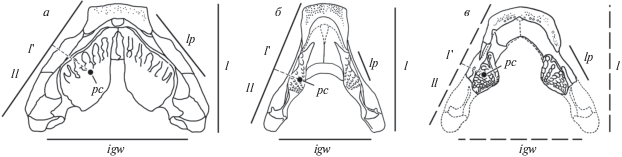
Для первой группы характерны укороченные относительно межгленоидного расстояния ветви челюсти (рис. 5, а). Межгленоидная ширина (igw) больше общей длины челюсти (l), и больше или равна длине ее ветви (l '). Длина пластинок (lp)может превышать половину длины ветви челюсти. Геометрический центр преартикулярных пластинок (pc) располагается в передней половине длины ветви челюсти. Вследствие этого и, несмотря на общее укорочение челюсти, плечо рычага (ll) оказывается относительно длинным, что делает силу воздействия на пищевой объект не такой уж большой, какой она могла бы быть при заднем расположении геометрического центра пластинки.
Этот дисбаланс компенсируется общим укорочением челюстей, что увеличивает силу, прикладываемую к пищевому объекту при аддукции челюсти, и приводит к медиальному смыканию преартикулярных зубных пластинок в симфизной области. Тесному контакту противолежащих нижнечелюстных пластинок соответствовало в разной степени выраженное слияние верхнечелюстных, что позволяло укрепить свод неба за счет не только птеригоидных костей, но и толстого слоя дентина самих зубных пластинок. Это увеличивало эффективность склерофагии. С другой стороны, медиальное смыкание пластинок нижней челюсти, например, у хиродиптерид (рис. 5, а), у которых пластинки занимали большую часть площади дна ротовой полости, сужало и укорачивало медиальное пространство для размещения basihyale и хондроидно-соединительнотканного “языка”, поддерживаемого этим скелетным элементом.
Во второй группе (рис. 5, б, в) межгленоидная ширина равна или меньше как общей длины челюсти, так и ее ветвей. Длина зубной пластинки значительно меньше половины длины несущей ее ветви. Геометрический центр преартикулярных пластинок располагается в середине длины ветви челюсти, или даже несколько позади нее. Несмотря на относительно длинные ветви челюстей, плечо рычага оказывается относительно более коротким, чем у членов первой группы, что делает силу воздействия на пищевой объект большей, чем если бы геометрический центр пластинки располагался более рострально.
Как показано У. Бимисом и Г. Лодером (Bemis, Lauder, 1986), в обработке пищевого объекта у современных двоякодышащих (Neoceratodus, Protopterus и Lepidosiren) гипобранхиальный аппарат совершает вертикальные движения, возвратно-поступательно прокачивающие воду через ротовое отверстие. Этот механизм назван авторами “гиоидным насосом”. Ток воды, создаваемый этим “насосом”, передвигает пищевой комок внутри ротовой полости и даже частично выводит его наружу изо рта в процессе “пережевывания”. Сходство морфологии висцерального аппарата современных и ископаемых дипной позволяет предположительно распространить наши представления о его функционировании и на последних.
У каждой из двух описанных выше конструкций есть свои морфо-функциональные преимущества и недостатки. Уменьшение площади “языка” и укорочение передней части челюсти у девонских дипной в первой группе (рис. 6, а) со слитыми медиально зубными пластинками уменьшало поверхность гиоидного насоса, приводившее, соответственно, к уменьшению амплитуды гидравлического внутриротового давления и ослаблению возможности манипуляции пищевым комком. Возможно, впрочем, что в этой группе девонских дипной таких манипуляций и не осуществлялось, поскольку большая часть дна ротовой полости была занята зубными пластинками, и необходимости передвигать пищу для более полного измельчения не возникало, то есть размеры добычи должны были быть сравнимы с размером суммарной площади зубных пластин в каждой челюсти.
Рис. 6.
Реконструкция площади “поршня гиоидного насоса” (обозначена серой заливкой) и конфигурация симфизной пластины нижних челюстей у различных девонских двоякодышащих (стеносимфизный и платисимфизный типы); крестовидные диаграммы показывают соотношение ширины рта и длины симфизной пластины: а, б, г – стеносимфизный тип, в, д – платисимфизный тип: а – Chirodipterus australis Miles, 1977; б – Dipterus valenciennesi Sedgwick et Murchison, 1828; в – Anchidipterus dariae Krupina, gen. et sp. nov.; г – Rhinodipterus ulrichi Ørvig, 1961; д – Orlovichthys limnatis Krupina, 1980 [а – из Майлса (Miles, 1977); б, г – из Ярвика (Jarvik, 1967); д – из Крупиной (1980)].
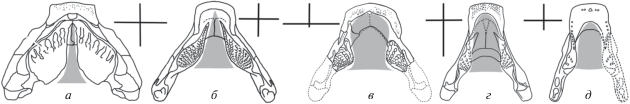
У диптерид площадь пластинок невелика, поэтому пространство между ними, занятое “языком”, пропорционально больше, чем в первой группе (рис. 6, б–д). Эффективная, по сравнению с первой группой, площадь “поршня” гиоидного насоса обеспечивала улавливание объекта питания за счет всасывания воды в ротовую полость и давала возможность значительного манипулирования им. Линейные размеры добычи могли превышать площадь самих зубных пластин, и ее обработка могла проводиться поэтапно с помощью передвижения пищевого комка внутри ротовой полости и за ее пределами, как это и происходит у современных Protopterus и Lepidosiren (Bemis, Lauder, 1986).
Функциональное значение губных складок у современных двоякодышащих описано Бимисом (Bemis, 1986), который показал, что эти мягкие структуры определяют размер и форму ротового отверстия, что важно при засасывающем типе поимки добычи. У современных дипной отсутствуют кости передней части рыла, поэтому для них хондроидно-соединительнотканные губы формируют не только латеральные, но также дорсальный и вентральный края рта. У девонских дипной последние сформированы костями верхней и нижней челюстей. Вероятно, что мышцы и соединительные ткани губных складок, крепившиеся в лабиальной ямке нижней челюсти постеро-латерально от симфиза у девонских дипной, ограничивали ротовое отверстие по бокам и, таким образом, регулировали приточное отверстие гиоидного насоса. Соотношения ширины и длины симфизной пластины во второй группе варьируют в значительной степени (рис. 6, а–д; 7, а–в). Ее ширина определяла размер вентрального края ротового отверстия. У девонских дипной мы выделяем стено- и платисимфизный варианты конфигурации симфизных пластин. Стеносимфизная конструкция указывает на ротовое отверстие меньшего, платисимфизная – большего размера. Новый род и вид Anchidipterus dariae gen. et sp. nov. относится, без сомнения, к платисимфизному типу (рис. 6, в; 7, а), тогда как Dipterus (рис. 6, б; 7, б) и Rhinodipterus (рис. 6, г) – к стеносимфизному. Неожиданно, Orlovichthys (рис. 6, д; 7, в) попадает в платисимфизную группу, несмотря на удлиненные челюсти и узкое ротовое отверстие; это связано с его относительно короткой симфизной пластиной. У стеносимфизных форм, согласно закону Бернулли, увеличенная скорость потока воды (позволявшая эффективнее затягивать пищевой объект) сопровождалось снижением давления в нем (что уменьшало мощность всасывания). Стеносимфизный рот, таким образом, был лучше приспособлен к захвату более мелких, но быстро плавающих объектов, например, таких, как мелкие членистоногие. У платисимфизных ротовое отверстие позволяло заглатывать более крупные, но менее подвижные пищевые объекты.
Длина симфизной пластинки могла быть коррелятивно связана и с формой пищевого объекта. Для удержания удлиненных, червеобразных объектов было достаточно короткого симфиза, который способствовал перехватыванию объекта поперек на любом участке его длины (рис. 7, а), а удлиненный был более эффективен при захвате более коротких, но изометричных организмов, для удержания которых была необходима бóльшая поверхности контакта, зачастую ложковидная (рис. 7, б, в).
Рис. 7.
Возможные способы захвата пищевых объектов различным диптеридными двоякодышащими: а – захват удлиненного объекта с помощью короткого симфиза у Anchidipterus; б, в – захват изометричного объекта с помощью значительной поверхности контакта удлиненного симфиза: б – Dipterus (из: Jarvik, 1967), в – Orlovichthys (из: Крупина, 1980). Эффективность захвата увеличивали челюстные зубчики на оральной поверхности нижней челюсти. Обозначение: jd – челюстные зубчики.
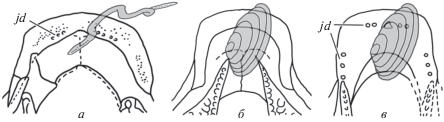
Длина ветвей нижней челюсти спереди от переднего края зубных пластин во второй группе могла достигать значительных размеров (напр., у Rhinodipterus kimberleyensis: Clement, 2012, рис. 3a, в). Удлинение верхней и нижней челюстей создавало дополнительный инструмент добычи и фиксации пищевого объекта, работавший как пинцет. Эффективность захвата этого “пинцета” должны были увеличивать мелкие зубчики на гладких, покрытых космином оральных поверхностях верхней и нижней челюсти (jd), известные, например, у Anchidipterus dariae gen. et sp. nov. (рис. 7, а), Dipterus oervigi (Gross, 1964), D. valenciennesi (Jarvik, 1967), Orlovichthys limnatis (Krupina et al., 2001) (рис. 7, в) и у мальков Andreyevichthys epitomus (Krupina, 1992; Krupina, Reisz, 1999; Крупина, 2004). Удлиненные челюсти также давали возможность копать придонные осадки в поисках мелких, неглубоко зарывающихся животных.
Сочетание таких признаков, как платисимфизная конструкция рта, гиоидный насос с пониженной скоростью, но большей мощностью всасывания, невозможность перемалывания пищевого комка зубными пластинками, а только его сжатие, дают возможность предположить, что анхидиптеры питались в биловском бассейне слабо армированной или мягкотелой, малоподвижной добычей относительно крупного размера. По этим параметрам в качестве объектов питания для этих рыб можно предположить разнообразных червей, мелких беззамковых брахиопод, тонкораковинных или лишенных раковин моллюсков и мелких членистоногих. Из этих животных в биловском сообществе известны все группы, кроме червей, наличие которых в любом бассейне легко предположить. Таким образом, анхидиптеры, вероятнее всего, занимали в трофической структуре сообщества нишу неспециализированных омнифагов.
ЗАКЛЮЧЕНИЕ
Местонахождение Билово – одно из немногочисленных фаменских местонахождений в восточной части Главного девонского поля (ГДП). Оно богато палеонтологическими находками – как беспозвоночных, так и разнообразных позвоночных животных. Первая описанная из фаменских отложений ГДП двоякодышащая рыба, отнесенная нами к семейству Dipteridae Owen, 1846, Anchidipterus dariae Krupina, gen. et sp. nov., характеризуется коротким и широким симфизным отделом нижней челюсти, ростролатеральной ориентировкой мезиальных зубных рядов зубных пластинок и значительным углом их расхождения.
Девонские двоякодышащие рыбы с зубными пластинками разделены на две морфо-функциональные группы. К первой относятся склерофагические формы с относительно короткими челюстями и крупными, сближенными или смыкающимися медиально зубными пластинками. Ко второй – преимущественно омнифаги с относительно небольшими, раздвинутыми латерально и сдвинутыми назад зубными пластинками. В первой группе тесный контакт крупных противостоящих нижнечелюстных пластинок увеличивал эффективность склерофагии. Во второй межчелюстное пространство, занятое “языком”, было относительно больше. Эффективная по сравнению с первой группой площадь “поршня” гиоидного насоса обеспечивала более активное улавливание подвижных объектов питания в полость и давала возможность значительного манипулирования пищевым комком.
Ширина нижнечелюстного симфиза во второй группе определяла размер ротового отверстия. Мы выделяем стено- и платисимфизный варианты конфигурации симфизных пластин. Стеносимфизный рот был лучше приспособлен к захвату более мелких, но быстро плавающих объектов. У платисимфизных форм ротовое отверстие позволяло заглатывать более крупные, но менее подвижные пищевые объекты. К этой морфофункциональной группе отнесен описываемый здесь Anchidipterus dariae Krupina, gen. et sp. nov.
Длина симфизной пластины могла быть коррелятивно связана с формой пищевого объекта. Для удержания удлиненных, червеобразных объектов было достаточно короткого симфиза, длинный был более эффективен при захвате коротких, но изометричных организмов.
Платисимфизная конструкция нижней челюсти в сочетании с относительно небольшими, широко расставленными зубными пластинами позволяют предположить, что добычей анхидиптеров в биловском бассейне были слабо армированные или мягкотелые беспозвоночные. Анхидиптеры могли занимать в трофической структуре сообщества нишу неспециализированных омнифагов.
* * *
Авторы статьи благодарны сотруднику Торопецкой биостанции “Чистый лес” (Тверская обл.) Д.Г. Бондарь, которая нашла экземпляр, описанный в настоящей статье, и передала его в Андреапольский краеведческий музей им. Э.Э. Шимкевича. Мы признательны А.О. Иванову за рецензирование, которое значительно улучшило качество данной работы. Работа выполнена в рамках Программы фундаментальных научных исследований Президиума РАН № 17 “Эволюция органического мира. Роль и влияние планетарных процессов” по проекту “Перестройка экологической структуры сообществ позвоночных в периоды крупнейших экосистемных кризисов в позднем палеозое и начале мезозоя”. Фотографии выполнены в ПИН РАН С.В. Багировым.
Список литературы
Воробьева Э.И. Морфология и особенности эволюции кистеперых рыб. М.: Наука, 1977. 239 с. (Тр. Палеонтол. ин-та АН СССР. Т. 163).
Геккер Р.Ф., Филиппова М.Ф. VII. Разрез по р. Ловати и ее притокам – рр. Б. и М. Тудрам, Кунье и Смоте // Отложения Главного девонского поля. Части V–VII. Л., 1935. С. 40–82 (Тр. Лен. геол.-гидро-геодез. треста. Вып. 9).
Вербицкий В.Р., Вербицкий И.В., Васильева О.В. и др. Государственная геологическая карта Российской Федерации. Масштаб 1 : 1 000 000 (третье поколение). Серия Центрально-Европейская. Листы O-35 – Псков, (N‑35), O-36 – Санкт-Петербург. Объяснительная записка. СПб.: Картографическая фабрика ВСЕГЕИ, 2012. 508 с.
Крупина Н.И. Новый род дипной из фамена Центрального девонского поля // Палеонтол. журн. 1980. № 3. С. 140–143.
Крупина Н.И. Подкласс Dipnoi. Двоякодышащие // Ископаемые позвоночные России и сопредельных стран. Бесчелюстные и древние рыбы / Ред. Новицкая Л.И. М.: ГЕОС, 2004. С. 373–422.
Молошников С.В., Линкевич В.В. Новые данные по позднедевонской ихтиофауне Тверской области (Билово, р. Малый Тудер) // Интегративная палеонтология: перспективы развития для геологических целей. Матер. LXIII сессии Палеонтол. об-ва при РАН (3–7 апреля 2017 г., Санкт-Петербург). СПб., 2017а. С. 196–198.
Молошников С.В., Линкевич В.В. Систематическое положение фаменских антиарх (Vertebrata, Placodermi) Тверской области // Изв. ВУЗов. Геол. и разведка. 2017б. № 2. С. 8–14.
Саммет Э.Ю. Восточная часть Главного девонского поля // Стратиграфия СССР. Девонская система / Ред. Наливкин Д.В. и др. М.: Недра, 1973. Т. 1. С. 90–100.
Bemis W.E. Feeding systems of living Dipnoi: anatomy and function // Bemis W.E., Buggren W.W., Kemp N.E. The biology and evolution of lungfishes // J. Morphol. 1986. Centennial Suppl. P. 249–275.
Bemis W.E., Lauder G.V. Morphology and function of the feeding apparatus of the lungfish, Lepidosiren paradoxa (Dipnoi) // J. Morphol. 1986. V. 187. № 1. P. 81–108.
Blaauwen J.L. den, Barwick R.E., Campbell K.S.W. Structure and function of the tooth plates of the Devonian lungfish Dipterus valenciennesi from Caithness and the Orkney Islands // Rec. West. Austral. Mus. 2005. V. 23. P. 91–113.
Bystrow A.P. On the dentition of Fleurantia denticulata // Докл. АН СССР. 1944. Т. 44. С. 31–32.
Campbell K.S.W., Barwick R.E. Early evolution of dipnoan dentitions and a new genus Speonesydrion // Mem. Assoc. Australas. Pa1aeonto1. 1983. V. 1. P. 17–49.
Campbell K.S.W., Barwick R.E. Paleozoic dipnoan phylogeny: functional complexes and evolution without parsimony // Paleobiology. 1990. V. 16. P. 143–169.
Campbell K.S.W., Barwick R.E. A new tooth-plated dipnoan from the Upper Devonian Gogo Formation and its relationships // Mem. Queensl. Mus. 1998. V. 42. № 2. P. 403–437.
Clement A.M. A new species of long-snouted lungfish from the Late Devonian of Australia, and its functional and biogeographical implications // Palaeontology. 2012. V. 55. P. 51–71.
Denison R.H. The structure and evolution of teeth in lungfishes // Fieldiana, Geol. 1974. V. 33. P. 31–58.
Graham-Smith W., Westoll T.S. On a new long-headed dipnoan fish from the Upper Devonian of Scaumenac Bay, P.Q., Canada // Trans. Roy. Soc. Edinburgh. 1937. V. 59. P. 241–266.
Gross W. Über Crossopterygier und Dipnoer aus dem baltischen Oberdevon im Zusammenhang einer vergleichenden Untersuchung des Porenkanalsystems paläozoischer Agnathen und Fische // Kungl. Svenska Vetenskapsakad. Handl. Fjarde Ser. 1956. Bd 5. № 6. P. 3–140.
Gross W. Über die Randzähne des Mundes, die Ethmoidalregion des Schädels und die Unterkiefersymphyse von Dipterus oervigi n. sp. // Paläontol. Z. 1964. Bd 38. H. 1–2. P. 7–25.
Jarvik E. On the structure of the lower jaw in dipnoans: with a description of an early Devonian dipnoan from Canada, Melanognathus canadensis gen. et sp. nov. // J. Linn. Soc. (Zool.). 1967. V. 47. № 311. P. 155–183.
Kemp A. Marginal tooth bearing bones in the lower jaw of the Recent Australian lungfish, Neoceratodus forsteri (Osteichthyes:Dipnoi) // J. Morphol. 1995. V. 225. P. 345–355.
Krupina N.I. Some data on the ontogeny of Devonian dipnoans // Academia. 1992. V. 1. P. 215–222.
Krupina N.I., Reisz R.R. Reconstruction of the dentition in hatchlings of Andreyevichthys epitomus, a Late Famennian dipnoan from Russia // Modern Geol. 1999. V. 24. P. 99–108.
Krupina N.I., Reisz R.R., Scott D. The skull and tooth system of Orlovichthys limnatis, a Late Devonian dipnoan from Russia // Canad. J. Earth Sci. 2001. V. 38. № 9. P. 1301–1311.
Lukševičs E. Bothriolepid antiarchs (Vertebrata, Placodermi) from the Devonian of the north-western part of the East European Platform // Geodiversitas. 2001. V. 23. № 4. P. 489–609.
Miles R.S. Dipnoan (lungfish) skulls and the relationships of the group: a study based on new species from the Devonian of Australia // Zool. J. Linn. Soc. 1977. V. 61. P. 1–328.
Schultze H.-P. A new long-headed dipnoan (Osteichthyes) from the Middle Devonian of Iowa, USA // J. Vert. Paleontol. 1992. V. 12. № 1. P. 42–58.
Smith M.M., Smithson T.R., Campbell K.S.W. The relationships of Uronemus: a Carboniferous dipnoan with highly modified tooth plates // Phil. Trans. Roy. Soc. B. Biol. Sci. 1987. V. 317. P. 299–327.
White E.I. The head of Dipterus valenciennesi Sedgwick & Murchison // Bull. Brit. Mus. (Natur. Hist.) Geol. 1965. V. 11. P. 1–45.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал