

Палеонтологический журнал, 2019, № 6, стр. 105-110
Находки жвачных (Artiodactyla, Ruminantia) и новые данные по стратиграфии позднемиоценового местонахождения Фортепьянка 2 (Северный Кавказ, Республика Адыгея)
И. А. Вислобокова a, *, К. К. Тарасенко a, **
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: ivisl@paleo.ru
** E-mail: tarasenkokk@gmail.com
Поступила в редакцию 01.02.2019
После доработки 07.02.2019
Принята к публикации 15.04.2019
Аннотация
Находка остатков жвачных родов Euprox Stehlin, 1928 (Dicrocerini, Muntiacinae, Cervidae) и Protragocerus Depéret, 1887 (Boselaphini, Bovinae, Bovidae) в 2018 г. и фаунистический анализ позволили уточнить возраст наземной фауны сводного разреза на р. Фортепьянке. Показано, что фауна соответствует раннему валлезию, зоне млекопитающих MN 9 Европы и относится к первой половине тортона, верхнему сармату (херсонию) Восточного Паратетиса.
Местонахождение позднемиоценовой фауны позвоночных Фортепьянка 1, открытое в 2003 г. в долине р. Фортепьянки, приток р. Белой (Россия, Республика Адыгея, Майкопский р-н), содержит ископаемые остатки наземных и морских животных и представляет особый интерес в связи с возможностью корреляции наземных и морских фаун. В блиновской подсвите гавердовской свиты этого местонахождения найдены бобр Trogontherium (Euroxenomys) minutum (von Meyer, 1838), позднемиоценовые представители подсемейства Monachinae [в том числе тюлени Monachopsis pontica (Eichwald, 1850)], Kurdalagonus cf. maicopicus (Spasskii, 1951), Kurdalagonus sp., представители “Pithanodelphini” (в том числе предположительно зубатые киты рода Sarmatodelphis), гиппарионы Cormohipparion sp. 1 и sp. 2 и другие позвоночные, вместе с остатками двустворчатых моллюсков Mactra (Chersonimactra) caspia (Eichwald, 1841) (Тарасенко и др., 2014). Фрагменты зубов и нижнечелюстных костей жвачных из слоя 10 были определены как Cervidae indet. (Тарасенко и др., 2014). Предварительное изучение фауны Фортепьянки 1 показало, что по составу наземных позвоночных она относится к валлезию Европы и, в верхах разреза, возможно, – к раннему туролию, а по составу морских млекопитающих и моллюсков (M. caspia) соответствуeт позднему сармату Восточного Паратетиса (Тарасенко и др., 2014).
В ходе работ Крымско-Кавказского отряда ПИН РАН в 2018 г. были собраны новые материалы из местонахождения Фортепьянка 2 (охватывает основание и среднюю часть сводного разреза на р. Фортепьянке, в то время как местонахождение Фортепьянка 1 включает в себя также кровлю сводного разреза). В слое 12 были найдены ископаемые остатки мелкого оленя рода Euprox Stehlin, 1928 из трибы Dicrocerini подсемейства Muntiacinae и небольшого полорогого рода Protragocerus Depéret, 1887 из трибы Boselaphini подсемейства Bovinae. Присутствие представителей этих родов вместе с ранними гиппарионами позволяет уточнить возраст наземной фауны позвоночных из сводного разреза на р. Фортепьянке, используя при этом данные биохронологии, палеогеографии и палеоклиматологии, а также палеомагнитные датировки фаун млекопитающих из разных регионов Европы и Азии.
ОПИСАНИЕ МАТЕРИАЛОВ
Олень Еuprox sp. из местонахождения Фортепьянка 2 по морфологии и размерам лобной кости, рогов и верхнего моляра M3 (рис. 1, а–е) более всего соответствует Е. furcatus (Hensel, 1859) из среднего и верхнего миоцена Eвропы, зоны млекопитающих MN 7–MN 9/MN 10 (см. Stehlin, 1928; Thenius, 1948, 1950; Czyżewska, Stefaniak, 1994; Azanza, 2000; Aiglstorfer et al., 2014). У экз. ПИН, № 5462/65 пенeк длинный, лежит в плоскости лба, спереди переходит в мощный продольный валик. Розетка хорошо развита, слабо наклонена вперед и в сторону; поверхность рогов бороздчатая. Отростки обломаны, но, очевидно, расходились под острым углом. Промеры: переднезадний и медиолатеральный (поперечный) диаметры (DAP×DT) основания рогов у экз. ПИН № 5462/65 и № 5462/66 – 21 × 19.5 мм и 17 × 16.5 мм, соответственно; длина и ширина (L × W) M3 у экз. ПИН, № 5462/77 – 11.7 × 15 мм.
Рис. 1.
Остатки жвачных из местонахождения Фортепьянка 2, Северный Кавказ; поздний миоцен, херсоний: а–e – Еuprox sp. (Cervidae): а–в – экз. ПИН, № 5462/65, фрагмент правой лобной кости с основанием рога спереди (а), сзади (б) и его реконструкция (в); г – экз. ПИН, № 5462/66, основание правого рога с медиальной стороны; д, е – экз. ПИН, № 5462/77, правый верхний моляр M3 с окклюзиальной (д) и лабиальной (е) сторон; ж–и – Protragocerus sp. (Bovidae), экз. ПИН, № 5462/78, фрагмент левой ветви нижнечелюстной кости с m2 и m3 с окклюзиальной (ж), лабиальной (з) и лингвальной (и) сторон.

В раннем валлезии E. furcatus присутствует в Испании (Асамбухейра, MN 9) и Германии (Эппельсгейм, MN 9) (Gentry et al., 1999), а E. аff. furcatus – в среднем сармате (бессарабии) Молдавии (Калфа и Бужор 1) (Лунгу, 1984). Род Euprox был широко распространен в среднем миоцене и начале позднего миоцена от Испании до Монголии и Китая и исчез в середине тортона на рубеже валлезия и туролия в Европе и, по-видимому, в конце бахо в Китае. К последним представителям рода в Европе относятся E. dicranocerus (Kaup, 1839) из местонахождений Вена III (Wien III, MN 9/10, 10.5 млн л. н.), Австрия (Gentry et al., 1999) и Euprox sp. из Кофидиш (Kohfidisch, Pannonian Zone H, начало MN11, <8.7 млн л. н.), Австрия (Вислобокова, 2007). В Монголии эупроксы известны только из среднего миоцена свиты Ошин Котловины Больших Озер – E. margaritae Vislobokova, 1983 (Хиргис-Нур II, MN 7–MN 8) и Euprox sp. (Наран-Булак) (Вислобокова, 1983). В Китае самые древние эупроксы найдены в местонахождении Тунгур (12.6–11.8 млн л. н., MN 7/8) Внутренней Монголии (Qiu et al., 2013), а самые молодые – на юго-западе Китая (пров. Юннань) в бассейне Юаньмоу в формации Сяохе (8.2–7.2 млн л. н.) и в бассейне Луфен в Шихуибе (~6.9–6.2 млн л. н.) (Dong et al., 2003; Dong, Qi, 2013; Qiu et al., 2013).
По морфологии и размерам моляров m2 и m3 (L × W m2 – 18 × 12.2 мм, L × W m3 – 24 × 11.2 мм) фрагмент левой ветви нижнечелюстной кости Protragocerus sp., экз. ПИН, № 5462/78, из Фортепьянки 2 (рис. 1, ж–и) близок к паратипу P. chantrei Deperét, 1887 (фрагмент нижней челюсти с p4–m3: Deperét, 1887, табл. XII , фиг. 6 ; из местонахождения Ла Грив Сент Альбан; серравалий, MN 7/8, Франция). Этот образец также имеет сходство с молярами P. glutens (Pilgrim, 1937) (=Strepsiportax glutens Pilgrim, 1937) из Сиваликов Индии: в частности, из местонахождения Hурпур (Gaur et al., 1985, табл. II , фиг. 2). Примитивные признаки моляров [брахиодонтность, слабое развитие лингвальных элементов, сильная струйчатость (“rugosities”) эмали] в сочетании с узостью коронок и их небольшими размерами отличают Protragocerus от других базальных бозеляфин, в том числе ранних Miotragocerus Stromer, 1928 и Tragoportaх Pilgrim, 1937. Вид P. chantrei был распространeн в Европе в серравалии и раннем тортоне, MN 7–MN 9, возможно, вплоть до середины валлезия (Sommerein, MN 9/MN 10, Австрия) (Deperét, 1887; Thenius, 1956; Gentry et al., 1999; Fuss et al., 2015). В Сиваликах Индии Protragocerus существовал в интервале 13.6–9.8 млн л. н., а в Сиваликах Пакистана – 13.9–10.8 млн л. н., в Непале – в Нижних Сиваликах (Dang Valley, ~MN 7/8) (Pilgrim, 1937; Patnaik, 2013). В Индии самые ранние Protragocerus известны из местонахождения Рамнагар (13.8–13.2 млн л. н.), а самые поздние – из Нурпура (10.1–9.7 млн л. н.).
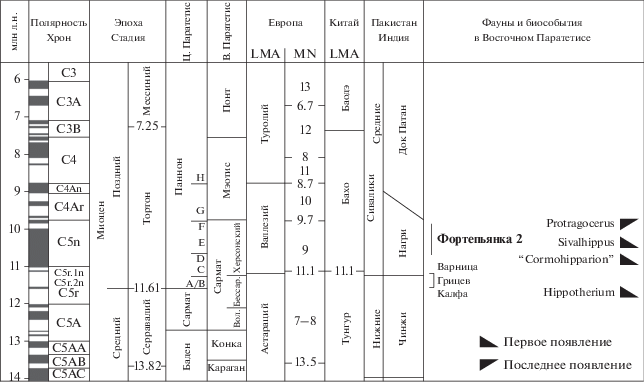
ОБСУЖДЕНИЕ
Данные по фаунам наземных позвоночных позволяют предполагать, что представители родов Euprox и Protragocerus могли впервые прийти на Кавказ в среднем миоцене. При этом они не исключают возможности инвазий этих форм на Кавказ и в сарматское время. Так, в состав фауны Беломечетской (MN 6, чокрак, Северный Кавказ, правобережье Кубани) входит ряд таксонов Ruminantia (представители родов Dorcatherium, Hispanomeryx, Hipsodontus, Giraffokeryx), которые отражают связи этой северокавказской фауны с фаунами Европы и Азии, в том числе Внутренней Монголии Китая и Сиваликов Индийского субконтинента (Pickford et al., 2000). Из Беломечетской (MN 6) описан фрагмент верхнего моляра Heteroprox sp. (=Paradicrocerus flerovi Gabunia, 1959: Pickford et al., 2000), не отличимый от Euprox. Остатки ранних бозеляфин из Беломечетской отнесены к Paratragocerus caucasicus Sokolov, 1945 (Габуния, 1973; Pickford et al., 2000). По строению рогов этот вид сходен с P. chantrei из Ла Грив, и Paratragocerus Sokolov, 1945, возможно, тождествен Protragocerus (см. Thenius, 1956). В сарматской фауне Эльдара (MN9, Восточная Грузия) представлена антилопа, близкая к Miotragocerus leskewitschi (Borissiak, 1914) [=Protragocerus leskewitschi: Korotkevitch, 1988; Tragoportax leskewitschi: Gentry et al., 1999; Gentry, Kaiser, 2009] конца среднего сармата (верхи MN9) из Севастополя (Крым) и, возможно, Варницы (Молдавия). Этa антилопа присутствует в Эльдаре вместе с гиппарионом Hippotherium ex gr. primigenium (v. Meyer, 1829).
Первое появление гиппарионов (т. н. “Hipparion” datum) является важным биохронологическим маркером для начала позднего миоцена. Распространение гиппарионов в Западную Палеарктику, включая Кавказ, связано с биогеографическими и ландшафтно-климатическими изменениями. Усиление аридизации способствовало расширению открытых пространств и вселению в регионы Восточного Паратетиса элементов азиатских фаун, в том числе гиппарионов. Здесь прослеживаются две основные волны расселения ранних гиппарионов: “северная” с ранними Hippotherium и “южная” с “Cormohipparion”. По данным палеомагнитного анализа, в Северное Причерноморье гиппарионы рода Hippotherium могли распространиться 11.9 млн л. н. или 11.53–11.47 млн л. н. (Vangengeim et al., 2006). В Молдавии в калфинском (C5r2n или C5An.1n) комплексе они представлены видом H. sarmaticum Lungu, 1973, очень похожим на Н. primigenium (Лунгу, 1984), а на Украине в Грицеве – Н. primigenium (Krakhmalnaya, 1996). В регионах Центрального Паратетиса первое появление гиппарионов рода Hippotherium фиксируется в горизонте С паннона Венского бассейна и относится к интервалу 11.4–11.0 млн л. н.; ранний вид H. primigenium был наиболее характерен для горизонтов D–E паннона (Bernor et al., 2017). Появление гиппарионов на Северном Кавказе, по-видимому, связано с “южной” волной расселения гиппарионов. В Фортепьянке в слое 12 и ниже, по данным В.И. Жегалло, присутствуют остатки гиппариона Cormohipparion sp. 2, близкого по морфотипу к сиваликскому C. nagriensis (Hussain, 1971) [=Sivalhippus nagriensis, ~10.7–10.4 млн л. н. (Wolf et al., 2013)], а в слое 20 – Cormohipparion sp. 1, близкий к C. theobaldi (Lydekker, 1877) [=Sivalhippus theobaldi, 9.3–7.8 млн л. н. (Wolf et al., 2013)]. В Сиваликах и Западной Азии (Турции) первые “Cormohipparion” sp. появились 10.8 млн л. н., а наиболее ранние Sivalhippus – S. nagriensis – в Сиваликах датируются 10.7 млн л. н. (Wolf et al., 2013; Bernor et al., 2017). Cormohipparion theobaldi был адаптирован к влажным стациям, а S. nagriensis – к более сухим. На Таманском полуострове и в Закавказье, судя по палеомагнитным данным из разрезов морских отложений сармата, эти даты соответствуют низам верхнего сармата (херсонию) Восточного Паратетиса. В Восточном Паратетисе возраст границы между средним и верхним сарматом – 11.2 млн л. н. (ниже субхрона С5r.1n), а между сарматом и мэотисом ~9.7 млн л. н. (Vangengeim et al., 2006). По другим данным, возраст этих границ более молодой, ~9.8 млн л. н. и ~7.65 млн л. н., соответственно (Popov et al., 2006). Совместное присутствие в фауне Фортепьянки Euprox sp. и Protragocerus sp. с гиппарионом, близким к S. nagriensis, предполагает ранневаллезийский возраст фауны из слоeв 12 и ниже. Нижний возрастной рубеж фауны Фортепьянки 2 определяет первое возможное появление в регионе “Сormohipparion” (<10.8 млн л. н.) и S. nagriensis (<10.7 млн л. н.), а верхний – верхний предел распространения Protragocerus, который в Европе и Азии не превышал 9.8 млн л. н. Этот временной интервал охватывает большую часть зоны MN 9 (рис. 2).
Ассоциация жвачных Euprox–Protragocerus, а также присутствие бобра в сводном разрезе на р. Фортепьянке, свидетельствуют о сохранении довольно гумидных условий окружающей среды и соответствующих биотопов. Эупрокс и протрагоцерус – обитатели влажных, залесeнных пространств. Они питались в основном листьями и побегами деревьев и кустарников, а также травянистыми растениями. Оба рода появились в среднем миоцене, и их представители были распространены в серрaвалии и начале тортона в гумидных условиях от Испании до центральных и южных районов Азии. В валлезии эупроксы и протрагоцеросы были обычны в Центральном и Восточном Паратетисе. Euprox sp. представлен в фауне Атцельсдорф (Atzelsdorf, Pannonian Zone C, основание MN9, ~11.2–11.1 млн л. н.) (Hillenbrand et al., 2009), где зафиксировано первое появление Hippotherium в Венском бассейне (Bernor et al., 2017). На Пиренейском полуострове Protragocerus исчез на границе раннего и позднего валлезия, зон MN 9 и MN 10 (C4Ar.3r), во время так называемого средневаллезийского кризиса, вместе с Amphiprox, Miotragocerus и др. (Agustí et al., 2001). В регионах Центрального Паратетиса ассоциация Euprox–Protragocerus была характерна для начала тортона и также существовала до рубежа MN 9/MN 10. Процессы аридизации в этой части Европы были выражены слабее, чем на востоке, и среднемиоценовые влаголюбивые формы растительноядных животных сохранялись дольше. В Северном Причерноморье Восточного Паратетиса количество таких форм заметно сократилось в конце среднего сармата (бессарабии), с расширением открытых пространств лесостепного и степного типов.
Подобные преобразования фауны прослежены в среднем сармате Молдавии при переходе от калфинской к варницкой фаунам (Лунгу, 1984). В фауне Варницы (поздний бессарабий), в частности, отсутствует Euprox. На Украине самая поздняя ассоциация Euprox и Protragocerus известна из Грицева и Климентовичей (Короткевич, 1988), близких по возрасту к калфинскому комплексу (Vangengeim et al., 2006). Все эти среднесарматские фауны древнее 11.2 млн л. н. В Центральном Паратетисе выше этой границы заметный сдвиг к более гумидным условиям приурочен к интервалу ~10.4–9.8 млн л. н. (Böhme et al., 2011). Вполне вероятно, что ассоциация жвачных Euprox–Protragocerus в разрезе на р. Фортепьянке также относится к этому интервалу между морскими регрессиями в конце среднего сармата и на рубеже сармата и мэотиса. Такая датировка допускает приход Protragocerus на Кавказ с “Cormohipparion” theobaldi из Южной Азии, так как этот вид гиппарионов, адаптированных к влажным стациям, и P. gluten, присутствуют в Нурпуре (10.1–9.7 млн л. н.) в Сиваликах Индии. Возможно, усиление гумидности привело к расширению ареалов азиатских форм в южные районы Западной Палеарктики, прилегающие к Сарматскому морю.
Таким образом, новые находки жвачных и фаунистический анализ позволили уточнить возраст наземной фауны сводного разреза на р. Фортепьянке. Эта фауна соответствует раннему валлезию, зоне MN 9 Европы и относится к первой половине тортона, верхнему сармату (херсонию) Восточного Паратетиса. Новые данные о возрасте этой группы местонахождений по наземным млекопитающим хорошо согласуются с возрастом, установленным ранее по морским млекопитающим и моллюскам из этого местонахождения.
Исследование выполнено при финансовой поддержке РНФ18-74-10081 “Эволюция сообществ позвоночных в позднем кайнозое Восточной Европы”.
Список литературы
Вислобокова И.А. Ископаемые олени Монголии. М.: Наука, 1983. 79 с.
Вислобокова И.А. Новые данные о млекопитающих позднемиоценовой фауны Кофидиш, Австрия // Палеонтол. журн. 2007. № 4. С. 96–103.
Габуния Л.К. Беломечетская фауна ископаемых позвоночных. Тбилиси: Мецниереба, 1973. 140 с.
Короткевич Е.Л. История формирования гиппарионовой фауны Восточной Европы. Киев: Наук. думка, 1988. 162 с.
Лунгу А.Н. Гиппарионовая фауна среднего сармата Молдавии. Кишинев: Штиинца, 1984. 160 с.
Тарасенко К.К., Лопатин А.В., Зеленков Н.В. и др. Новые местонахождения наземных и морских млекопитающих позднего миоцена в Адыгее (Россия, Северный Кавказ) // Докл. АН. 2014. Т. 459. № 6. С. 726–728.
Aiglstorfer M., Rössner G.E., Böhme M. Dorcatherium naui and pecoran ruminants from the late Middle Miocene Gratkorn locality (Austria) // Palaeobiodiv. Palaeoenv. 2014. V. 94. P. 83–123.
Agustí J., Cabrera L., Gracés M. A calibrated mammal scale for the Neogene of Western Europe. State of the art // Earth-Sci. Rev. 2001. V. 52. P. 247–260.
Azanza B. Los Cervidae (Artiodactyla, Mammalia) del Mioceno de las cuencas del Duero, Tajo, Calatayud-Teruel y Levante // Mem. Mus. Paleontol. Univ. Zaragosa. 2000. V. 8. P. 1–376.
Bernor R.L., Göhlich U.B., Harzhauser M. et al. The Pannonian C hipparions from the Vienna Basin // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2017. V. 476. P. 28–41.
Böhme M., Winklhofer M., Ilg A. Miocene precipitation in Europe: temporal trends and spatial gradients // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2011. V. 304. P. 212–218.
Czyżewska T., Stefaniak K. Euprox furcatus (Henzel, 1859) (Cervidae, Mammalia) from Przeworno (Middle Miocene, Lower Silesia, Poland) // Acta. zool. cracov. 1994. V. 33. № 1. P. 55–74.
Depéret Ch. Recherches sur la succession des faunes de vertébrés miocènes de la vailée du Rhône // Arch. Mus. Hist. Natur. Lyon. 1887. T. 4. P. 45–313.
Dong W., Liu J., Pan Y. A new Euprox from the Late Miocene of Yuanmou, Yunnan Province, China, with interpretation of its paleoenvironment // Chin. Sci. Bull. 2003. V. 48. P. 485–491.
Dong W., Qi G.-Q. Hominoid-producing localities and biostratigraphy in Yunnan // Fossil mammals of Asia: Neogene biostratigraphy and chronology / Eds. Wang X., Flynn L.J., Fortelius M. N.Y.: Columbia Univ. Press, 2013. P. 293–313.
Fuss J., Prieto J., Böhme M. Revision of the boselaphin bovid Miotragocerus monacensis Stromer, 1928 (Mammalia, Bovidae) at the Middle to Late Miocene transition in central Europe // N. Jb. Geol. Paläontol. Abh. 2015. Bd 276. Hf. 3. S. 229–265.
Gaur R., Vasishat R.N., Chopra S.R.K. New and some additional fossil mammals from the Siwaliks exposed at Nurpur, Kangra District (H.P.), India // J. Paleontol. Soc. India. 1985. V. 30. P. 42–48.
Gentry A.W., Rössner G.E., Heizmann E.P.J. Suborder Ruminantia // The Miocene land mammals of Europe / Eds. Rössner G.E., Heissing K. München: Verlag Dr. F. Pfeil, 1999. P. 225–258.
Hillenbrand V., Göhlich U.B., Rössner G.E. The early Vallesian vertebrates of Atzelsdorf (Late Miocene, Austria) // Ann. Naturhist. Mus. Wien. 2009. Bd 111 A. P. 519–556.
Krakhmalnaya T. Hipparions of the Northern Black Sea coast area (Ukraine and Moldova): Species composition and stratigraphic distribution // Acta Zool. Cracov. 1996. V. 39. № 1. P. 261–267.
Patnaik R. Indian Neogene Siwalik mammalian biostratigraphy: An overview // Fossil mammals of Asia: Neogene biostratigraphy and chronology / Eds. Wang X., Flynn L.J., Fortelius M. N.Y.: Columbia Univ. Press, 2013. P. 423–444.
Pickford M., Gabunia L., Mein P. et al. The Middle Miocene mammalian site of Belomechetskaya, North Caucasus: an important biostratigraphic link between Europe and China // Geobios. 2000. V. 33. № 2. P. 257–267.
Pilgrim G.E. Siwalik antelopes and oxen in the American Museum of Natural History // Bull. Amer. Mus. Natur. Hist. 1937. V. 72. P. 729–874.
Popov S.V., Shcherba I.G., Ilyina L.B. et al. Late Miocene to Pliocene palaeogeography of the Paratethys and its relation to the Mediterranean // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2006. V. 238. P. 91–106.
Qiu Zh., Qiu Zh., Deng T. et al. Neogene Land Mammal Stages/Ages of China: Toward the goal to establish an Asian Land Mammal Stage/Age scheme // Fossil mammals of Asia: Neogene biostratigraphy and chronology / Eds. Wang X., Flynn L.J., Fortelius M. N.Y.: Columbia Univ. Press, 2013. P. 29–90.
Stehlin H. Bemerkungen über die Hirsche von Steinheim am Aalbuch // Eclog. Geol. Helv. 1928. Bd 21. P. 245–256.
Thenius E. Zur Kenntnis der fossilen Hirsche des Wiener Beckens, unter besonderer Berücksichtigung ihrer stratigraphischen Bedeutung // Ann. Naturhist. Mus. Wien 1948. Bd 56. P. 262–308.
Thenius E. Die tertiären Lagomeryciden und Cerviden der Steiermark // S.-Ber. Österr. Akad. Wiss. Math.-Naturwiss. Kl. 1950. Bd 159. P. 219–254.
Thenius E. Zur Entwicklung des Knochenzapfens von Protragocerus Depéret (Bovidae, Mammalia) aus dem Miozän // Geologie. 1956. V. 5. № 4–5. P. 308–318.
Vangengeim E.A., Lungu A.N., Tesakov A.S. Age of the Vallesian lower boundary (Continental Miocene of Europe) // Stratigr. Geol. Corr. 2006. V. 14. № 6. P. 655–667.
Wolf D., Bernor R.L., Hussain S.T. Evolution, biostratigraphy and geochronology of Siwalik hipparionine horses // Palaeontogr. Abt. A. 2013. V. 300. P. 1–115.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал