

Палеонтологический журнал, 2019, № 6, стр. 25-30
Раннекембрийские томмотииды разреза Хаирхан (Центрально-Тувинский прогиб)
Н. В. Новожилова a, b, *, М. Штайнер c, И. В. Коровников a, b, Д. А. Токарев a, b
a Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
b Новосибирский государственный университет
Новосибирск, Россия
c Свободный университет Берлина
Берлин, Германия
* E-mail: NovozhilovaNV@ipgg.sbras.ru
Поступила в редакцию 14.12.2018
После доработки 31.05.2019
Принята к публикации 31.05.2019
Аннотация
Нижний кембрий разреза Хаирхан, расположенного в пределах Центрально-Тувинского прогиба, впервые охарактеризован находками томмотиид. Комплекс мелкораковинной фауны здесь представлен видами таких типичных для вехнеатдабанского века родов, как Tannuolina, Kelanella, Lugoviella и Microdictyon. Установлено, что склеритом вида Lugoviella ojmuranica составляли различные морфотипы склеритов.
Томмотииды – одна из групп кембрийской проблематичной фауны, в ископаемом состоянии встречающейся, большей частью, в виде разрозненных симметричных и асимметричных, колпачковидных и седловидных склеритов фосфатного состава. В основном их сравнивают с животными, наружный скелет которых был образован склеритами различных морфотипов. Наиболее широко известны с основания кембрия и до начала среднего кембрия, именно в этот период они имели высокие темпы эволюции и практически повсеместное глобальное распространение, что обусловило их высокое стратиграфическое значение.
До сих пор не существует единой концепции о систематике томмотиид, несмотря на то, что известны и описаны находки сросшихся или сочлененных вместе склеритов (Демиденко, 2004; Skovsted et al., 2008, 2011; Larsson et al., 2014). Наиболее общепринятым является предположение о родстве томмотиид с брахиоподами и форонидами (Holmer et al., 2008; Skovsted et al., 2008, 2009, 2011; Balthasar et al., 2009; Murdock et al., 2012). В данной работе принимается систематика Н.В. Есаковой (Есакова, Жегалло, 1996), которая, на наш взгляд, является наиболее удобной и обоснованной, где в составе отряда Tommotiida Missarzhevsky, 1970 выделяется шесть семейств: Tommotiidae Missarzhevsky in Bengtson, 1970, Tannuolinidae Fonin et Smirnova, 1967, Lapworthellidae Missarzhevsky in Rozanov et Missarzhevsky, 1966, Sunnaginiidae Landing, 1984, Kelanellidae Missarzhevsky et Grigorieva, 1981, Kennardidae Laurie, 1986. Следует отметить, что данная классификация является достаточно условной, но может быть использована так же, как и другие систематики этой группы (Landing, 1984; Dzik, 1986; Миссаржевский, 1989; Conway Morris, Chen, 1990), пока не будут определены единые критерии для обособления семейств в пределах отряда Tommotiida.
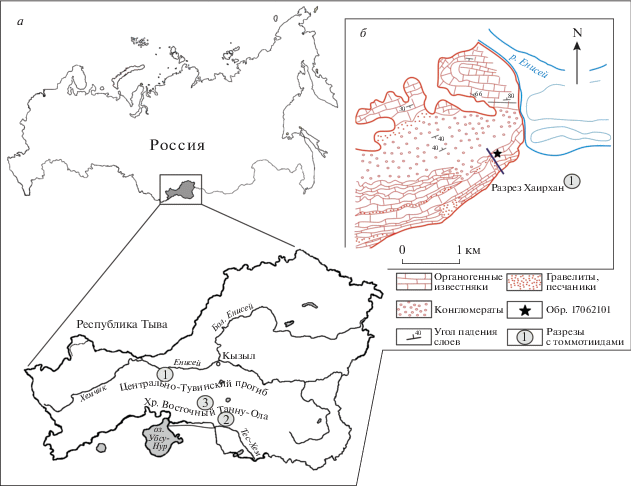
Находки раннекембрийских томмотиид на территории Тывы ранее были известны из шанганской свиты (рис. 1) разрезов, расположенных на водоразделе хребта Восточный Танну-Ола в верховьях р. Шевелиг-Хем и в низовьях р. Улуг-Шанган (Фонин, Смирнова, 1967; Журавлева и др., 1967), и представлены одним видом Tannuolina multifora Fonin et Smirnova, 1967. В результате экспедиционных работ в 2017-2018 гг. был отобран материал из баянкольской свиты нижнего кембрия разреза г. Хаирхан, расположенного в пределах Центрально-Тувинского прогиба (рис. 1, Б; координаты точки 17062101: 51°34′22.8′′ с.ш., 93°03′44.3′′ в.д.), где впервые обнаружены томмотииды родов Tannuolina, Kelanella и Lugoviella. Ранее из этого разреза были описаны лишь находки археоциат и трилобитов (Покровская, 1959; Журавлева и др., 1967).
Рис. 1.
Местоположение района работ: A – схематическая карта республики Тыва, цифрами обозначены разрезы нижнего кембрия с томмотиидами: 1 – разрез г. Хаирхан; 2 – разрез в верховьях р. Шевелиг-Хем; 3 – разрез в низовьях р. Улуг-Шанган; Б – план-схема геологического строения изученного района.
Томмотииды рода Tannuolina представлены видами Tannuolina fonini Esakova in Esakova et Zhegallo, 1996 (табл. IV , фиг. 1–5 , см. вклейку) и Tannuolina sp. (табл. IV , фиг. 5 ). Для T. fonini выделяют два типа склеритов (Есакова, Жегалло, 1996): митральных (табл. IV , фиг. 1а, 2) и селлатных (табл. IV , фиг. 3, 4 а). Митральные склериты имеют вид уплощенной пирамиды с практически овальным основанием. От вершины до основания проходит один слабовыраженный киль. Практически по всей наружной поверхности склеритов наблюдаются многочисленные поры (табл. IV , фиг. 1б) различного диаметра. Селлатные склериты удлиненные, асимметричные, с уплощенной вершиной, изогнутой к дубликатуре, что отличает их от Tannuolina multifora. С внутренней стороны одного из седловидных склеритов впервые обнаружены шипообразные выросты (табл. IV , фиг. 4 б), в поле зрения насчитывается 20 шипов (диаметр шипов варьирует от 0.01 до 0.025 мм, высота до 0.05 мм).
Рис. 2.
Lugoviella ojmuranica Gigorieva in Grigorieva et al., 1983: а–в – общий вид склеритов типа А, экз. ИНГГ, №№ 17062102/19–21; г, д – тип С, общий вид склеритов, экз. ИНГГ, №№ 17062102/22–23; е–и – общий вид склеритов типа В, экз. ИНГГ, №№ 17062102/24–27; к–м – склериты типа D, экз. ИНГГ, №№ 17062102/28–30. Обозначения: H – длина склерита, L – максимальная ширина основания, Т – высота основания склерита, в – вершина, ос – основание, п – внутренние перегородки. Шкала – 200 мкм.

На нашем материале был обнаружен асимметричный склерит рода Tannuolina (табл. IV , фиг. 5 ), который может являться как представителем другого вида этого рода, так и асимметричным типом селлатных склеритов T. fonini. Такое предположение основано на описании двух типов селлатных склеритов (симметричных и асимметричных) для T. maroccana Skovsted et Clausen, 2014 (Skovsted et al., 2014). К сожалению, пока обнаружен только один экземпляр, и этого недостаточно для установления таксономической принадлежности этого склерита.
Представители рода Kelanella (табл. V , фиг. 1–8 , см. вклейку) в изученном материале представлены несколькими типами склеритов, которые составляли единый склеритом вида Kelanella altaica Missarzhevsky in Rozanov et Missarzhevsky, 1966. Детальное описание различных морфотипов склеритов для вида K. altaica (A, B, C, D, E) ранее было дано Л. Дэвар с соавт. (Devaere et al., 2014). Мы согласны, что склериты, ранее рассматриваемые не только как разные виды, но и как представители различных родов томмотиид, могли образовывать единый склеритом. Несмотря на то, что в предложенной ими классификации имеются некоторые спорные на наш взгляд моменты, для определения различных типов склеритов K. altaica в данной работе использовалась именно эта (Devaere et al., 2014) терминология и типизация. Но род Kelanella считаем целесообразным рассматривать в составе семейства Kelanellidae Missarzhevsky et Grigorieva, 1981, и часть склеритов типа А (Devaere et al., 2014, фиг. 7, H–L ) очень сходны с представителями типа D, а симметричные склериты того же типа А (Devaere et al., 2014, фиг. 7, N, O ) сходны с типом В. Таким образом, в баянкольской свите разреза Хаирхан установлены склериты K. altaica типа А (табл. V , фиг. 2), типа В (табл. V , фиг. 4 ), типа E (табл. V , фиг. 1) и типа D (табл. V , фиг. 3, 5, 6 ). Дополнительно определены склериты не установленного у Дэвар с соавт. типа (табл. V , фиг. 7, 8 ), которые идентичны типовому виду K. altaica (Розанов, Миссаржевский, 1966). К сожалению, всего обнаружено 14 экз., и пока этого оказалось недостаточно для проведения ревизии видовых типов раннекембрийских форм келанеллид Тывы.
Установлено, что склеритом вида Lugoviella ojmuranica Grigorieva, 1983 составляли склериты четырех различных морфотипов (табл. V , фиг. 9–11 ; рис. 2, а–м), детальное описание которых приведено ниже (см. описание рода Lugoviella). Все выделенные типы были обнаружены с одного стратиграфического уровня, и их объединяет присутствие внутренних перегородок, морфология и характерная только для рода Lugoviella микроструктура наружной поверхности склеритов.
Комплекс мелкораковинной фауны изученного разреза представлен томмотиидами родов Tannuolina, Kelanella и Lugoviella и склеритами Microdictyon effusum Matthews, Bengtson et Missarzhevsky, 1981 (табл. IV , фиг. 6, 7 ). Это позволяет говорить о позднеатдабанском возрасте изученного интервала разреза. Большинство томмотиид имеет внутренние перегородки, которые начали появляться у них только в атдабанское время.
Используемая нами методика химического препарирования позволила извлечь из серых массивных известняков баянкольской свиты целые склериты хорошей сохранности. Образцы помещались в стаканы емкостью 1 л и заливались раствором уксусной кислоты, при этом чередовалась концентрация раствора 3% и 10%.
Палеонтологические коллекции хранятся в Лаб. палеонтологии и стратиграфии палеозоя Ин-та нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН под № 17062101. Работа выполнена при поддержке гранта РФФИ (проект № 16-05-00196) и программы ФНИ (проект IX.126.1.2).
ТИП И КЛАСС НЕ УСТАНОВЛЕНЫ
ОТРЯД TOMMOTIIDA
СЕМЕЙСТВО KELANELLIDAE MISSARZHEVSKY ET GRIGORIEVA, 1981
Род Lugoviella Grigorieva, 1983
Lugoviella: Григорьева и др., 1983а, с. 57.
Типовой вид – Lugoviella ojmuranica Grigorieva in Grigorieva et al., 1983; нижний кембрий, ботомский ярус Якутии (Григорьева и др., 1983).
Диагноз. Фосфатные асимметричные и симметричные склериты. Микроструктура внешней поверхности всех типов склеритов одинакова на всех сторонах склеритов и представлена в виде отчетливых, неравномерно расположенных поперечных ребер, между которыми расположены менее выраженные, более тонкие и многочисленные “струящиеся” продольные ребра (табл. V , фиг. 10 б). Для всех типов характерно наличие внутренних перегородок. Склеритом Lugoviella объединял в себе склериты четырех различных морфотипов:
1. Тип A. Асимметричные пирамидальные или конусовидные склериты, прямые или слабоизогнутые вбок. Поперечное сечение округлой, овальной, округло-прямоугольной или трапециевидной формы.
2. Тип B. Широкие и сильноуплощенные склериты в виде округленно-треугольной пластины, плавно расширяющиеся от вершины к основанию. Поперечное сечение очень узкое, слабоизогнутое, дугообразное. Внешняя и внутренняя стороны склерита практически плотно прилегают друг к другу. Вершина округлая, притупленная.
3. Тип C. Асимметричные колпачковидные склериты, резко расширяющиеся от вершины к основанию. Вершина округленно-треугольной формы, смещена к боковому краю. Поперечное сечение удлиненно-овальной, прямоугольной или трапециевидной формы.
4. Тип D. Прямые билатерально-симметричные склериты, с широким основанием, расширяющиеся от вершины к основанию. Вершина удлиненная, округлая. Поперечное сечение овальной, округленно-прямоугольной формы по всей длине склерита.
Сравнение. В составе семейства рассматриваются роды Kelanella и Lugoviella. Описываемый род отличается от представителей другого рода формой склеритов и характерной скульптурой наружной поверхности, проявленной на всех сторонах склерита.
Замечания. Терминология и измерения различных морфотипов склеритов, составляющих склеритом рода Lugoviella (и типового вида Lugoviella ojmuranica), показаны на рис. 2.
Lugoviella ojmuranica Grigorieva in Grigorieva et al., 1983
Табл. V , фиг. 9–11
Lugoviella ojmuranica: Григорьева и др., 1983, с. 57, табл. V , фиг. 4–6 ; Григорьева, 1983, с. 164, табл. XII , фиг. 8, 9 ; Миссаржевский, 1989, табл. XIX , фиг. 4 ; Есакова в Есакова, Жегалло, 1996, с. 115, табл. XIV , фиг. 1–9 ; Розанов и др., 2010, с. 88, табл. 59 , фиг. 1–3 .
Lugoviella chizhovi: Есакова в Есакова, Жегалло, 1996, с. 115, табл. XIV , фиг. 10, 11 .
Голотип – ПИН, № 3848/203, ядро; Якутия, р. Лена, разрез Ой-Муран; нижний кембрий, ботомский ярус.
Описание (рис. 2, а–м). Склеритом составляют склериты фосфатного состава различной морфологии, которые разделены на четыре типа в зависимости от размеров и формы. Все типы склеритов объединяет характер наружной поверхности (см. описание рода), присутствие внутренних перегородок и совместное нахождение. Краткое описание основных морфотипов склеритов Lugoviella ojmuranica дано при описании рода, ниже рассмотрим каждый тип более детально.
Наиболее многочисленными являются представители типа A (табл. V , фиг. 9–11 ; рис. 2, а–в). Это асимметричные пирамидальные или конусовидные склериты, прямые (рис. 2, а) и слабоизогнутые вбок (рис. 2, б, в) или неправильно изогнутые (табл. V , фиг. 10 а, 11). Поперечное сечение округлой, овальной, округло-прямоугольной или трапециевидной формы. Апикальная часть у всех изученных склеритов этого типа обломана. Максимальная ширина основания составляет 1.1 мм, длина склеритов 1.5–2.6 мм, высота основания 0.4–0.6 мм.
Плоские и широкие, крупные склериты типа В менее многочисленны и имеют вид округленно-треугольной “пластины”, плавно расширяющейся от вершины к основанию (рис. 2, е–и). Поперечное сечение очень узкое и слабоизогнутое. Внешняя и внутренняя стороны склерита практически плотно прилегают друг к другу, особенно у вершины склерита. Вершина плоская (рис. 2, е, ж) или округло-треугольной формы (рис. 2, з, и). Некоторые отличия апикальной части склеритов обусловлены внутривидовой изменчивостью. Максимальная ширина основания составляет 2.4 мм. Высота основания склеритов 0.05 мм, длина 1.2–2.5 мм.
У колпачковидных склеритов типа С (рис. 2, г, д) вершина смещена к боковому краю, поперечное сечение удлиненно-овальной, прямоугольной или трапециевидной формы. Скульптура наружной поверхности менее ярко выражена по сравнению с остальными типами, особенно на внутренней стороне. Внутри имеются перегородки. Максимальная длина склеритов составляет 0.9 мм, высота основания 0.1 мм, ширина основания составляет 0.8–1.2 мм.
Билатерально-симметричные склериты типа D (рис. 2, к–м) отличаются от остальных типов прямой формой, широким основанием и плавным расширением от вершины к основанию. Вершина имеет округлую форму. Поперечное сечение удлиненное, от овального до округленно-прямоугольного по всей длине склерита. Стороны не примыкают друг к другу, в отличие от склеритов типа B. Скульптура внешней поверхности наиболее ярко выражена с внешней стороны склеритов. Внутри имеются поперечные перегородки (рис. 2, к, л). Длина склеритов составляет 1.1 мм, высота основания 0.1 мм, ширина основания 1 мм.
Замечания. Установленные морфотипы склеритов, на наш взгляд, принадлежали одному склеритому вида L. ojmuranica и были обнаружены в образцах, отобранных с одного стратиграфического уровня. Кроме того, их объединяет присутствие внутренних перегородок, морфология и характерная только для рода Lugoviella структура наружной поверхности склеритов.
Распространение. Нижний кембрий, ботомский ярус – Якутия, Северо-Западная Монголия; атдабанский ярус – Республика Тыва.
Материал. Около 35 экз., среди которых 20 склеритов типа A, 6 склеритов типа В, 3 экз., относящиеся к типу С, и 6 склеритов типа D из баянкольской свиты атдабанского яруса нижнего кембрия, Республика Тыва (Центрально-Тувинский прогиб, разрез г. Хаирхан).
Список литературы
Григорьева Н.В. Скелетные проблематичные организмы // Ярусное расчленение нижнего кембрия Сибири. Атлас окаменелостей. М.: Наука, 1983. С. 155–169 (Тр. ИГиГ СО АН СССР. Вып. 558).
Григорьева Н.В., Мельникова Л.М., Пельман Ю.Л. Брахиоподы, остракоды (брадорииды) и проблематика из стратотипического района ярусов нижнего кембрия // Палеонтол. журн. 1983. № 3. С. 54–58.
Демиденко Ю.Е. Новые данные по морфологии склеритов томмотиид Lapworthella fasciculatа // Палеонтол. журн. 2004. № 2. С. 23–27.
Есакова Н.В., Жегалло Е.А. Биостратиграфия и фауна нижнего кембрия Монголии. М.: Наука, 1996. 216 с. (Тр. Совм. Росс.-Монгол. палеонтол. экспед. Вып. 46).
Журавлева И.Т., Задорожная Н.М., Осадчая Д.В. и др. Фауна нижнего кембрия Тувы (опорный разрез р. Шивелиг-Хем). M.: Наука, 1967. 249 с.
Миссаржевский В.В., Григорьева Н.В. Новые представители отряда Tommotiida // Палеонтол. журн. 1981. № 4. С. 91–97.
Миссаржевский В.В. Древнейшие скелетные окаменелости и стратиграфия пограничных толщ докембрия и кембрия. М.: Наука, 1989. 238 с. (Тр. ГИН АН СССР. Вып. 443).
Покровская Н.В. Трилобитовая фауна и стратиграфия кембрийских отложений Тувы. М.: Изд-во АН СССР, 1959. 199 с. (Тр. ГИН АН СССР. Вып. 27).
Розанов А.Ю., Миссаржевский В.В. Биостратиграфия и фауна нижних горизонтов кембрия. М.: Наука, 1966. 126 с. (Тр. ГИН АН СССР. Вып. 148).
Розанов А.Ю., Пархаев П.Ю., Демиденко Ю.Е. и др. Ископаемые стратотипов ярусов нижнего кембрия. М.: ПИН РАН, 2010. 228 с.
Фонин В.Д., Смирнова Т.Н. Новая группа проблематичных раннекембрийских организмов и некоторые методы их препарирования // Палеонтол. журн. 1967. № 2. С. 15–27.
Balthasar U., Skovsted C.B., Holmer L.E., Brock G.A. Homologous skeletal secretion in tommotiids and brachiopods // Geology. 2009. V. 37. P. 1143–1146.
Conway Morris S., Chen M. Tommotids from the Lower Cambrian of China // J. Paleontol. 1990. V. 64. P. 169–184.
Devaere L., Clausen S., Monceret E. et al. The tommotiid Kelanella and associated fauna from the early Cambrian of southern Montagne Noire (France): implication for camenellan phylogeny // Palaeontology. 2014. V. 57. P. 979–1002.
Dzik J. Turrilepadida and other Machaeridia // Problematic fossil taxa / Eds. Hoffman A., Nitecki M.H. Oxford: Oxford Univ. Press, 1986. P. 116–134.
Holmer L.E., Skovsted C.B., Brock G.A. et al. The Early Cambrian tommotiid Micrina, a sessile bivalved stem group brachiopod // Biol. Letters. 2008. V. 4. P. 724–728.
Landing E. Skeleton of lapworthellids and the suprageneric classification of tommotiids (Early and Middle Cambrian phosphatic problematica) // J. Paleontol. 1984. V. 58. P. 1380–1398.
Larsson C.M., Skovsted C.B., Brock G.A. et al. Paterimitra pyramidalis from South Australia: scleritome, shell structure and evolution of a lower Cambrian stem group brachiopod // Palaeontology. 2014. V. 57. P. 417–446.
Murdock D.J.E., Donoghue P.C.J., Bengtson S., Marone F. Ontogeny and microstructure of the enigmatic Cambrian tommotiid Sunnaginia Missarzhevsky, 1969 // Palaeontology. 2012. V. 55. P. 661–676.
Skovsted C.B., Balthasar U., Brock G.A., Paterson J.R. The tommotiid Camenella reticulosa from the early Cambrian of South Australia: morphology, scleritome reconstruction, and phylogeny // Acta Palaeontol. Pol. 2009. V. 54. P. 525–540.
Skovsted C.B., Brock G.A., Paterson J.R. et al. The scleritome of Eccentrotheca from the Lower Cambrian of South Australia: lophophorate affinities and implications for tommotiid phylogeny // Geology. 2008. V. 36. P. 171–174.
Skovsted C.B., Brock G.A., Topper T.P. et al. Scleritome construction, biofacies, biostratigraphy and systematics of the tommotiid Eccentrotheca helenia sp. nov. from the early Cambrian of South Australia // Palaeontology. 2011. V. 54. P. 253–286.
Skovsted C.B., Clausen S., Álvaro J.J., Ponlevé D. Tommotiids from the early Cambrian (Series 2, Stage 3) of Morocco and the evolution of the tannuolinid scleritome and setigerous shell structures in stem group brachiopods // Palaeontology. 2014. V. 57. P. 171–192.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал