

Палеонтологический журнал, 2019, № 6, стр. 31-39
Об устрицах рода Rhynchostreon Bayle (Bivalvia, Gryphaeidae) из верхнего маастрихта Горного Крыма
a Нанкинский институт геологии и палеонтологии Китайской академии наук
Нанкин, Китай
b Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
* E-mail: KosenkoIN@ipgg.sbras.ru
Поступила в редакцию 09.01.2019
После доработки 14.03.2019
Принята к публикации 07.06.2019
Аннотация
Изучена морфология и микроструктура раковины гладких экзогироидных устриц из верхнего маастрихта Горного Крыма. Микроструктура раковины этих устриц состоит из простой правильной листоватой, неправильной перекрещенно-пластинчатой и переходной от неправильной простой призматической к неправильной сложной перекрещенно-пластинчатой структур. Последний тип микроструктуры характерен только для рода Rhynchostreon, что наряду с формой раковин обосновывает отнесение изученных устриц к виду R. aralensis (Arkhangelsky). Находки устриц Rhynchostreon в верхнем маастрихте Горного Крыма позволяют уточнить верхнюю границу стратиграфического распространения рода.
ВВЕДЕНИЕ
Микроструктура раковин устриц является важным признаком для систематики надсемейства Ostreoidea, и в связи с этим ее изучению уделяется большое внимание (Чельцова, 1969; Siewert, 1971; Stenzel, 1971; Carter, 1980, 1990; Freneix, 1982; Malchus, 1990, 1998; Aqrabawi, 1993; Якушин, Иванов, 2001; Koppka, 2015; Kosenko, 2018; и др.).
Среди надсемейства Ostreoidea наиболее разнообразна микроструктура раковины у представителей семейства Gryphaeidae, расцвет которого пришелся на юру и мел (до настоящего времени из четырех подсемейств, Gryphaeinae, Exogyrinae, Gryphaeostreinae и Pycnodonteinae, дожили представители только последнего). За счет многообразия и сложности микроструктура раковины именно для этого семейства имеет наибольшее таксономическое значение. Особенно это касается вымерших таксонов, когда отсутствует возможность привлечения данных о строении мягкого тела. Наибольшее значение у грифеид имеет микроструктура левых (обычно более выпуклых и массивных) створок; кроме того, они больше подходят для изготовления шлифов.
В некоторых случаях изучение микроструктуры раковины является единственным способом идентифицировать таксономическую принадлежность того или иного вида устриц. Ниже такой случай рассматривается на примере устриц рода Rhynchostreon Bayle из верхнего маастрихта Горного Крыма.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Изученный материал представлен более чем 120 створками (главным образом левыми) устриц Rhynchostreon, собранными совместно с проф. В.В. Аркадьевым из разрезов верхнего маастрихта Баклинской и Корабельной куэст. Сбор раковин в Баклинской куэсте производился непосредственно из слоя, в Корабельной куэсте – из осыпи. Для изучения микроструктуры раковины было изготовлено 10 шлифов из левых и правых створок. При морфологическом анализе особое внимание уделялось изменчивости антимаргинальной скульптуры в выборке и на различных стадиях онтогенеза, а также хоматам. Перед фотографированием раковины напылялись хлоридом аммония (NH4Cl). Для фотографирования использовалась камера Canon EOS 60D. Изучение микроструктуры раковины проводилось на световом микроскопе Olympus BX60. Биометрический анализ раковин выполнялся в программе Past 3 (Hammer et al., 2001). Изученная коллекция раковин устриц и шлифы переданы на хранение в Центр коллективного пользования “Геохрон” (ИНГГ СО РАН) (колл. № 2087). В качестве сравнительного материала использовались раковины типового вида R. suborbiculatum (Lamarck) из сеномана Юго-Восточной Франции, хранящиеся в Музее естественной истории г. Лилля (Франция).
ГЕОЛОГИЧЕСКИЙ ОЧЕРК
Верхняя часть верхнего маастрихта Баклинской и Корабельной куэст (рис. 1, 2) представлена пачкой XXIII – алевролитами известковистыми глауконитовыми и песчаниками мелкозернистыми с ячеистой текстурой выветривания (Алексеев, 1989). Возраст пачки установлен (Алексеев, 1989) по находкам белемнитов Neobelemnella kazimiroviensis (Skołozdrówna), стратиграфическое распространение которого ограничено верхним маастрихтом (Christensen, 1990, 1996, 1997; Christensen et al., 1996; Keutgen et al., 2017). Двустворчатые моллюски крайне многочисленны в пачке и представлены главным образом устрицами и, в меньшей степени, пектинидами, неравномерно распространенными вертикально и латерально (рис. 2). Двустворчатые моллюски встречаются в виде разрозненных створок, в Баклинской куэсте иногда наблюдается сортировка раковин экзогир по размеру, что свидетельствует об аллохтонном характере захоронения.

ТЕРМИНОЛОГИЯ
Терминология, используемая при описании микроструктуры раковины, заимствована из ревизованных “Treatise …” (Carter et al., 2012). Названия микроструктур, предложенные в этой работе, были по возможности сопоставлены с терминологией, разработанной Н.А. Чельцовой (1969), которая широко используется в русскоязычной литературе (табл. 1). Приведем сопоставления микроструктур по Чельцовой (1969) и Дж. Картеру и др. (Carter et al., 2012) (эквивалент по Картеру и др. приводится в скобках). Структуры эктостракума по Чельцовой представлены тремя типами: нормальной призматической (эквивалент – призматическая [prismatic]), субконической (эквивалент отсутствует) и фурциллятной (эквивалент отсутствует). Структуры эндостракума у устриц по Чельцовой представлены правильной листоватой (эквивалент – простая правильная листоватая [simple regularly foliated, simple RF]), перекрещенно-пластинчатой (эквивалентна неправильной сложной перекрещенно-пластинчатой с маленькими углами [low angle irregularly complex crossed foliated, аббревиатура la ICCF]), субромбической (по всей видимости, эквивалентна неправильной сложной перекрещенно-пластинчатой с большими углами [high angle irregularly complex crossed foliated, аббревиатура ha ICCF]) и вакуолярной (эквивалентна везикулярной [vesicular]). Кроме того, у Картера с соавт. выделяются неправильная перекрещенно-пластинчатая структура (irregular cross foliated, ICF) и переходная от неправильной простой призматической к неправильной сложной перекрещенно-пластинчатой (transitional irregular simple prismatic/irregular complex crossed foliated, ISP/ICCF), эквивалентов которых в работе Чельцовой нет.
Таблица 1.
Соотношение названий микроструктур, принятых в работах Чельцовой (1969) и Картера и др. (Carter et al., 2012)
| Названия структур по Чельцова, 1969 | Названия структур по Carter et al., 2012 | |||
|---|---|---|---|---|
| русский перевод | оригинальное название | аббревиатура | ||
| эктостракума | нормальная призматическая | призматическая | prismatic | нет |
| субконическая | нет эквивалента | |||
| фурциллятная | нет эквивалента | |||
| эндостракума | правильная листоватая | простая правильная листоватая | simple regularly foliated | simple RF |
| перистая | правильная листоватая “селедочная кость” | heringbone regularly foliated | herringbone RF | |
| субромбическая | нет эквивалента | |||
| вакуолярная | везикулярная | vesicular | нет | |
| нет эквивалента | неправильная перекрещенно-пластинчатая | irregular crossed foliated | ICF | |
| перекрещенно-пластинчатая | неправильная сложная перекрещенно-пластинчатая (бывает двух типов – low angle и high angle) | irregular complex crossed foliated (low angle и high angle) | ICCF | |
| нет эквивалента | коническая сложная перекрещенно-пластинчатая | cone complex crossed foliated | cone CCF | |
| нет эквивалента | переходная от неправильной простой призматической к неправильной сложной перекрещенно-пластинчатой | transitional irregular simple prismatic/irregular complex crossed foliated | ISP/ICCF | |
ИСТОРИЯ ИССЛЕДОВАНИЯ
Позднемаастрихтские устрицы Горного Крыма изучались многими авторами. Установлено присутствие родов Agerostrea Vialov, 1936 и Acutostrea Vialov, 1936 (Flemingostreidae, Liostreinae), Pycnodonte Fisher de Waldheim, 1835 и Hyotissa Stenzel, 1971 (Gryphaeidae, Pycnodonteinae), и Gryphaeostrea Conrad, 1865 (Gryphaeidae, Gryphaeostreinae) (Чельцова, 1969; Собецкий, 1977; Донт, 2004). Относительно представителей подсемейства Exogyrinae (Gryphaeidae) данные разных авторов противоречивы. Это связано с разным пониманием таксонов надвидового ранга различными авторами, трудностью идентификации видов в связи с широкими пределами фенотипической изменчивости, а также с гомеоморфизмом, свойственным большинству устриц. Так, В.А. Собецкий (1977) описал три вида в составе рода Exogyra Say, 1820 – E. (E.) decussata (Goldfuss, 1833), E. (Costagyra) auricularis (Wahlenberg, 1821) и E. (C.) goldfussiana (Sobetsky, 1977), и один вид рода Ceratostreon Bayle, 1878 – C. spinosum (Matheron, 1842). Чельцова (1969) указывала присутствие вида Amphidonte aralensis (Arkhangelsky, 1912) в верхнем маастрихте Горного Крыма. А.В. Донт (2004) в списке фауны указывала присутствие Amphidonte decussata и под вопросом присутствие Ceratostreon pliciferum (Dujardin, 1837).
Собранные автором экзогиры по морфологии раковин делятся на две группы. Первая объединяет вытянутые в высоту гладкие раковины с хоматами, развитыми вдоль всего периметра внутренней поверхности створок. Эти устрицы должны быть отнесены к роду Amphidonte (в составе которого в качестве подрода в настоящее время рассматривается Ceratostreon – см. Malchus, 1990). Судя по всему, именно эти раковины определялись в работах предшественников как виды рода Ceratostreon. Вторая группа объединяет экзогир с сильно выпуклыми чашеобразными левыми створками, геликоидальным закручиванием макушки, и с хоматами, развитыми только вблизи замочной площадки. По всей видимости, эти экзогиры описывались ранее Собецким (1977) как Exogyra auricularis (Wahlenberg, 1821), Чельцовой (1969) – как Amphidonte aralensis (Arkhangelsky), а Донт (2004) были определены как Amphidonte decussata (Goldfuss, 1833).
Автором была исследована морфология и микроструктура раковин этих устриц с целью уточнения их таксономического положения. Ниже приводятся результаты этих исследований.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
НАДСЕМЕЙСТВО OSTREOIDEA RAFINESQUE, 1815
СЕМЕЙСТВО GRYPHAEIDAE VIALOV, 1936
ПОДСЕМЕЙСТВО EXOGYRINAE VIALOV, 1936
Триба Exogyrini Vialov, 1936
Род Rhynchostreon Bayle, 1878
Подрод Rhynchostreon s. str.
Rhynchostreon aralensis (Arkhangelsky, 1912)
Табл. VI , фиг. 1–6 (см. вклейку)
Рис. 3.
Микроструктура раковины устрицы Rhynchostreon aralensis (Arkhangelsky) (микрофотографии сделаны при скрещенных николях; положение микрофотографий на шлифе показано на фотографии шлифа в центре рисунка): simple RF – простая правильная листоватая структура; simple RF/ICF – переходная структура от простой правильной листоватой к неправильной перекрещенно-пластинчатой структуре; ICF – неправильная перекрещенно-пластинчатая структура; ISP/ICCF – переходная структура от неправильной простой призматической к неправильной сложной перекрещенно пластинчатой; ChM –камера, заполненная мелоподобным веществом; шлиф № К-216.

Рис. 4.
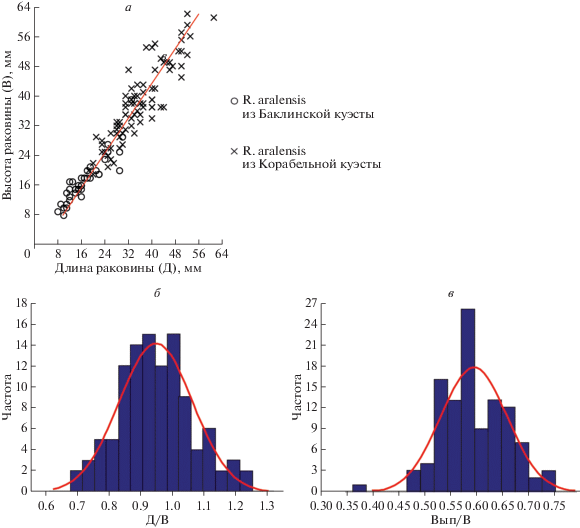
График зависимости высоты раковины (В) от длины (Д) (а) и гистограммы распределения коэффициента удлинения (Д/В) (б) и коэффициента выпуклости (Вып/В) (в) в выборке устриц Rhynchostreon aralensis (Arkhangelsky) из верхнего маастрихта Горного Крыма.
Exogyra aralensis: Архангельский, 1912, с. 45, табл. 2 , фиг. 14–18 .
Amphidonta aralensis: Ренгартен, 1964, с. 47, табл. VI , фиг. 8 а, б; Чельцова, 1969, с. 74, табл. XIII , фиг. 2–6 .
Exogyra (Costagyra) auricularis: Собецкий, 1977, с. 154, табл. XIV , фиг. 4–5 .
Лектотип – экз., изображенный Архангельским (1912, табл. 2 , фиг. 14 ); Узбекистан, Аральское море, о-в Возрождения; верхний маастрихт, зона Belemnitella americana; обозначен Чельцовой (1969, с. 74); место хранения неизвестно.
Описание. Левые створки сильно выпуклые, вместительные, треугольных очертаний, с округлым килем, смещенным к заднему краю створки. Киль делит створку на большую переднюю и меньшую круто падающую заднюю поверхности. Макушка округлая, ось завивания макушки расположена под углом около 45° к плоскости смыкания створок. Правые створки плоские или слабовогнутые (convex-concave по: Malchus, 1990), обычно с передним спирально закрученным килем. Изученные экземпляры имеют размеры от 8 до 62 мм в высоту (в среднем 32.7 мм) и от 8 до 61 мм в длину (в среднем 30.7 мм). В коллекции имеется одна правая створка, имеющая все морфологические признаки этого вида, достигающая очень большого размера – высоты в 147 мм и длины в 140 мм. Измерения этой створки не учитывались при подсчете средних значений. Площадка прирастания обычно очень маленькая, расположена вблизи макушки. Поверхность левой створки покрыта слабыми линиями роста. Некоторые экземпляры имеют тонкую антимаргинальную струйчатость вблизи макушки. Правые створки покрыты концентрическими чешуями роста. Замочная площадка узкая, экзогироидного типа. Резилифер левой створки слабовогнутый, его ширина сильно превышает ширину переднего валика. Задний валик редуцирован. Примакушечная полость хорошо выражена. Отпечаток мускула-замыкателя овальный, слегка вытянутый в высоту, смещен к заднему и верхнему краям створки, слабо вдавлен в поверхность створок. Отпечаток мускула Квенштедта не наблюдался. Спаечный край узкий, на внутренней поверхности правых створок ограничен резким перегибом, на внутренней поверхности левых створок ограничен вдавленной бороздой. Короткие прямые хоматы расположены вблизи замочной площадки.
Микроструктура. Левые створки показывают последовательность сменяющих друг друга типов микроструктур (от внешней поверхности левой створки к внутренней): simple regularly foliated – переходная от simple RF к irregular cross foliated – irregular cross foliated – transitional irregular simple prismatic/irregular complex crossed foliated (иногда члены последовательности могут быть сильно редуцированы) (рис. 3). При этом у наиболее толстых створок наблюдается несколько циклов из вышеуказанной последовательности микроструктур, что может быть объяснено сезонностью роста раковины. Количество камер с мелоподобным веществом также варьирует от полного отсутствия до присутствия нескольких, но все они приурочены к внешнему краю раковины.
Размеры в мм и их соотношения (по результатам замеров 121 экз. левой створки):
| В | Д | Вып. | Д/В | Вып/В | |
|---|---|---|---|---|---|
| Среднее | 31.24 | 29.55 | 18.6 | 0.96 | 0.59 |
| Максимум | 62 | 61 | 39 | 1.45 | 0.75 |
| Минимум | 8 | 8 | 5 | 0.68 | 0.36 |
Изменчивость. Проявляется в первую очередь в разной степени удлиненности раковин. Подсчет коэффициента отношения длины раковины к высоте Д/В в выборке из n раковин показал, что его значения изменяются от 0.67 до 1.25 при среднем значении 0.96 (рис. 4). Коэффициент выпуклости Вып/В изменяется в пределах от 0.37 до 0.75 при среднем значении 0.59. Изменчивости также подвержена скульптура раковины – большинство раковин характеризуется полным отсутствием радиальной скульптуры, в то время как некоторые экземпляры имеют антимаргинальную струйчатость вблизи макушек.
Сравнение. От типового вида рода R. suborbiculatum (Lamarck, 1801, с. 398, табл. 23 , фиг. 11–13 ; здесь, табл. VI , фиг. 7 ) из сеномана–турона Европы отличается резко несимметричными вытянутыми в длину очертаниями раковины (сравниваемый вид имеет почти симметричные раковины, похожие по форме на раковины юрских грифей). От R. langlosi (Coquand, 1869, с. 122, табл. XI , фиг. 11–16 ) из сантона Франции и Средней Азии отличается менее широкой раковиной, более вытянутой в длину. От R. mermeti (Coquand, 1862, с. 234, табл. 23 , фиг. 3–5 ; Malchus, 1990, с. 128, табл. 8 , фиг. 15–17 , табл. 9 , фиг. 1–4, табл. 9 , фиг. 5–21 ), распространенного в сеномане–туроне Северной Африки и на Ближнем Востоке, отличается отсутствием задне-верхней депрессии (см. Malchus, 1990, с. 94, фиг. 31 ) вблизи макушки правой створки, а также более вытянутыми в длину раковинами.
От R. asiaticum (Arkhangelsky) (Архангельский, 1912, с. 44, табл. 2 , рис. 9, 10, 11–13 ) из сеномана Средней Азии отличается более выпуклыми несимметричными вытянутыми в длину раковинами.
Замечания. Род Rhynchostreon в настоящее время рассматривается в составе трибы Exogyrini подсем. Exogyrinae (Malchus, 1990; Aqrabawi, 1993). Разными авторами объем рода и, как следствие, его стратиграфическое распространение понимается по-разному. Х.Х. Миркамалов (1966, с. 38–39) рассматривает в составе рода 28 видов, распространенных от неокома до маастрихта. Другие авторы (Malchus, 1990; Aqrabawi, 1993) понимают объем рода более узко (с распространением: ?верхний альб, сеноман–сантон, ?кампан), при этом отдельно рассматривается род Laevigyra Malchus, 1990 (?альб, сеноман–?верхний маастрихт), отличающийся большей вытянутостью раковин в высоту. М. Акрабави (Aqrabawi, 1993, рис. 23 ) показал, что для Laevigyra так же, как и для Rhynchostreon, характерно наличие структуры ISP/ICCF в микроструктуре раковины. Позднее Н. Мальхус стал рассматривать Laevigyra в качестве подрода Rhynchostreon (Malchus, 1996). При этом, давая диагноз рода, никто из авторов не приводил сравнение с широко распространенном в верхнем мелу родом Exogyra. Это, по-видимому, связано с тем, что типичные экземпляры рода Rhynchostreon, чьи изображения приводятся в литературе, начиная с XIX в., обладают относительной симметрией раковин, отсутствием радиальной скульптуры и слабо завернутой макушкой, что придает им внешнее сходство с юрскими грифеями и резко отличает от типичных экзогир с развитой радиальной скульптурой. Кроме того, Г. Стензель (Stenzel, 1971) отмечал отсутствие у представителей рода Rhynchostreon хомат, что также отличает их от экзогир, которые характеризуются наличием хомат вблизи замочной площадки. Однако, по мере накопления материала, стало ясно, что и для ринхостреонов характерны хоматы, а в выборках одного вида присутствуют экземпляры как с отсутствием радиальной скульптуры, так и с развитыми антимаргинальными ребрами (Malchus, 1990; Aqrabawi, 1993; Videt, Neradeau, 2003; Abdel-Gawad et al., 2006; и др.). Более того, не у всех экзогир развита радиальная или антимаргинальная скульптура. Как следствие, границы между родами Rhynchostreon и Exogyra нивелируются, и единственным способом достоверно различить два этих рода становится изучение микроструктуры их раковин. Род Exogyra характеризуется простой листоватой микроструктурой и развитием многочисленных линзовидных камер, заполненных мелоподобным веществом, реже пустых (Aqrabawi, 1993). Микроструктура рода Rhynchostreon слагается несколькими типами структур, из которых наиболее специфичной является ISP/ICCF (transitional irregular simple prismatic/irregular complex crossed foliated), которая среди всех Exogyrinae встречается только у этого рода; камеры с мелоподобным веществом развиты слабо. Таким образом, при большом внешнем сходстве раковин, микроструктура раковин родов Exogyra и Rhynchostreon очень сильно различается, что позволяет разграничивать два этих рода. Большое внешнее сходство между экзогирами и ринхостреонами объясняется гомеоморфизмом, свойственным большинству устриц (Malchus, 1998).
Распространение. Маастрихт Крыма, Кавказа, Туркмении и Приаралья.
Материал. Более 120 разрозненных преимущественно левых створок из Баклинской и Корабельной куэст.
ОБСУЖДЕНИЕ
Полученные данные по микроструктуре устриц Rhynchostreon aralensis из верхнего маастрихта Горного Крыма в целом согласуются с описанием микроструктуры этого вида, выполненным Чельцовой (1969, с. 74), и с данными по микроструктуре рода Rhynchostreon Мальхуса (Malchus, 1990) и Акрабави (Aqrabawi, 1993).
Характерной особенностью микроструктуры этого вида является присутствие структуры ISP/ICCF. Чельцова (1969) рассматривала этот вид в составе рода Amphidonta, к которому относила всех экзогир с гладкими раковинами (в настоящее время относимых к родам Amphidonte Fisher de Waldheim, 1829, Ilymatogyra Stenzel, 1971, Rhynchostreon и Aetostreon Bayle, 1878), и не признавала самостоятельность рода Rhynchostreon. В настоящее время самостоятельность рода Rhynchostreon не подвергается сомнению (Stenzel, 1971; Malchus, 1990; Aqrabawi, 1993; и др.), причем микроструктура рода сильно отличается от микроструктуры других представителей Exogyrinae наличием структуры ISP/ICCF (Malchus, 1990; Aqrabawi, 1993). При этом присутствие структуры ISP/ICCF в микроструктуре раковины является единственным надежным диагностическим признаком, позволяющим отличать представителей Rhynchostreon от Exogyra со схожей скульптурой раковин (для некоторых экзогир характерно практически полное отсутствие радиальной скульптуры, в то же время некоторые ринхостреоны имеют выраженную радиальную скульптуру). Таким образом, наличие ISP/ICCF в микроструктуре раковины R. aralensis доказывает его принадлежность к роду Rhynchostreon.
Присутствие R. aralensis в верхнем маастрихте Горного Крыма позволяет уточнить стратиграфическое распространение рода. Как было отмечено выше, стратиграфическое распространение рода Rhynchostreon оставалось дискуссионным, вследствие различного понимания объема рода разными исследователями. Полученные данные по микроструктуре R. aralensis позволяют расширить верхнюю границу стратиграфического распространения рода до верхнего маастрихта.
ЗАКЛЮЧЕНИЕ
Изучена микроструктура раковины устрицы Rhynchostreon aralensis (Arkhangelsky, 1912) из верхнего маастрихта Горного Крыма. Микроструктура левых створок этого вида состоит из последовательности сменяющих друг друга типов микроструктур (от внешней поверхности левой створки к внутренней): simple regularly foliated – переходная от simple RF к irregular cross foliated – irregular cross foliated – transitional irregular simple prismatic/irregular complex crossed foliated. Наличие структуры ISP/ICCF в микроструктуре раковины этого вида позволило уточнить его таксономическое положение в составе подсемейства Exogyrinae и включить в род Rhynchostreon. Присутствие рода Rhynchostreon в верхнем маастрихте Горного Крыма позволило уточнить верхнюю границу стратиграфического распространения рода.
* * *
Автор благодарен В.В. Аркадьеву (СПбГУ, С.‑Петербург) за помощь в организации и проведении полевых работ в Горном Крыму, П.А. Яну (ИНГГ СО РАН, Новосибирск) за помощь при подготовке микрофотографий, Н. Мальхусу (ICP Catalan Institute of Paleontology Miquel Crusafont, Барселона, Испания) за плодотворные обсуждения микроструктуры раковин устриц и ее значения для систематики, С.В. Попову (ПИН РАН, Москва) за конструктивные замечания при рецензировании статьи. Работа выполнена при поддержке проекта ФНИ № 0331-2019-0004 “Палеонтология, стратиграфия, биогеография бореальных и смежных с ними палеобассейнов и комплексное обоснование усовершенствования региональных стратиграфических схем мезозоя и кайнозоя Сибири”.
Список литературы
Алексеев А.С. Меловая система. Верхний отдел // Геологическое строение Качинского поднятия Горного Крыма (стратиграфия мезозоя) / Ред. Мазарович О.А., Милеев В.С. М.: МГУ, 1989. С. 123–157.
Архангельский А.Д. Ископаемая фауна берегов Аральского моря // Изв. Туркестанск. отд. Рус. геогр. об-ва. 1912. Т. 8. Вып. 2. С. 1–79.
Вялов О.С. О классификации устриц // Докл. АН СССР. 1936. Т. 4. № 1. С. 19–22.
Донт А.В. Кампанские и маастрихтские двустворчатые моллюски Горного Крыма: стратиграфическое распространение и палеобиогеография // Бюлл. Моск. об-ва испыт. прир. Отд. геол. 2004. Т. 79. Вып. 2. С. 60–67.
Миркамалов Х.Х. Экзогиры, их систематика и значение для стратиграфии меловых отложений юго-западных отрогов Гиссарского хребта. Ташкент: ФАН УзССР, 1966. 135 с.
Собецкий В.А. Двустворчатые моллюски позднемеловых платформенных морей юго-запада СССР. М.: Наука, 1977. 256 с.
Чельцова Н.А. Значение микроструктуры раковины меловых устриц для их систематики. М.: Наука, 1969. 83 с.
Якушин Л.Н., Иванов А.В. Краткий атлас позднемеловых двустворчатых моллюсков юго-востока Восточно-Европейской платформы. Саратов: Науч. книга, 2001. 116 с.
Abdel-Gawad G.I., El-Qot G.M., Mekawy M.S. Cenomanian-Turonian macrobiostratigraphy of Abu-Darag area, northern Galala, Eastern Desert, Egypt // Intern. Conf. Geology of the Arab World (GAW8), Cairo University. Giza, Egypt, 2006. P. 553–568.
Aqrabawi M. Oysters (Bivalvia-Pteriomorphia) of the Upper Cretaceous rocks of Jordan. Palaeontology, stratigraphy and comparison with the Upper Cretaceous oysters of northwest Europe // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 1993. V. 75. P. 1–135.
Carter J.G. Bivalve shell mineralogy and microstructure, parts A + B // Skeletal growth of aquatic organisms / Eds. Rhoads D.C., Luitz R.A. N.Y.: Plenum Press, 1980. P. 627–673.
Carter J.G. Shell microstructural data for the Bivalvia. Part IV. Order Ostreoida // Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. V. I / Ed. Carter J.G. N.Y.: Van Nostrand Reinhold, 1990. P. 347–362.
Carter J.G., Harries P.J., Malchus N., Sartori A.F. Treatise Online № 48: Pt N, Revised. V. 1. Chapt. 31: Illustrated Glossary of the Bivalvia. 2012. P. 1–209.
Christensen W.K. Upper Cretaceous belemnite stratigraphy of Europe // Cret. Res. 1990. V. 11. № 4. P. 371–386.
Christensen W.K. A review of the Upper Campanian and Maastrichtian belemnite biostratigraphy of Europe // Cret. Res. 1996. V. 17. № 6. P. 751–766.
Christensen W.K. Palaeobiogeography and migration in the Late Cretaceous belemnite family Belemnitellidae // Acta Palaeontol. Pol. 1997. V. 42 № 4. P. 457–495.
Christensen W.K., Hancock J.M., Peake N.B., Kennedy W.J. The base of the Maastrichtian // Bull. Geol. Soc. Denmark. 1996. V. 47. P. 81–85.
Coquand H. Géologie et paléontologie de la region sud de la province de Constantine. Marseille: Impr. Arnaud, 1862. 320 p.
Coquand H. Monographie du genre Ostrea. Terrain Crétacé. Marseille: Seren, 1869. 215 p.
Freneix S. Disparité microstructurale du test entre Gryphaeostrea et Gyrostrea (Huîtres du Crétacé-Miocène). Nouvelle classification des genres // Ann. Paléontol. 1982. V. 68. № 3. P. 223–240.
Hammer O., Harper D.A.T., Ryan P.D. PAST: Paleontological Statistics software package for education and data analysis // Palaeontol. Electron. 2001. V. 4. № 1. P. 1–9.
Keutgen N., Remin Z., Jagt J.W.M. The late Maastrichtian Belemnella kazimiroviensis group (Cephalopoda, Coleoidea) in the Middle Vistula valley (Poland) and the Maastricht area (the Netherlands, Belgium) – taxonomy and palaeobiological implications // Palaeontol. Electron. 2017. № 20.2.38A. P. 1–29. https://doi.org/10.26879/671
Koppka J. Revision of the Bivalvia from the Upper Jurassic Reuchenette Formation, Northwest Switzerland – Ostreoidea // Zootaxa. 2015. V. 3927. № 1. P. 1–117.
Kosenko I. The origin of the Pycnodonteinae and relationship between gryphaeas and true pycnodontes // Acta Palaeontol. Pol. 2018. V. 63. № 4. P. 769–778.
Lamarck J.B. de. Système des animaux sans vertèbres, ou tableau général des classes, des ordres et des genres de ces animaux. P.: Chez Deterville, 1801. 432 p.
Malchus N. Revision der Kreide-Austern (Bivalvia: Pteriomorphia) Ägyptens (Biostratigraphie, Systematik) // Berlin. Geowiss. Abh. Reihe A. 1990. Bd 125. 231 S.
Malchus N. Palaeobiogeography of Cretaceous oysters (Bivalvia) in the Western Tethys // Mitt. Geol.-Paläontol. Inst. Univ. Hamburg. 1996. V. 77. P. 165–181.
Malchus N. Multiple parallel evolution and phylogenetic significance of shell chambers and chomata in the Ostreoidea (Bivalvia) // Bivalves: An Eon of Evolution. Kansas: Univ. Calgary Press, 1998. P. 393–407.
Siewert W. von. Schalenbau und Stammesgeschichte von Austern // Stuttg. Beitr. Naturk. Ser. B. 1972. V. 1. P. 1–57.
Stenzel H.B. Oysters // Treatise on Invertebrate Paleontology. Pt N. Mollusca 6 (Bivalvia). V. 3 / Ed. Moore R.C. Boulder: Geol. Soc. Amer.; Univ. Kansas Press, 1971. P. 953–1124.
Videt B., Neraudeau D. Variabilité et hétérochronies chez l’exogyre Rhynchostreon suborbiculatum (Lamarck, 1801) (Bivalvia: Ostreoidea: Gryphaeidae) du Cénomanien et du Turonien inférieur des Charentes (SW France) // C. R. Palevol. 2003. V. 2. № 6–7. P. 563–576.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал