

Палеонтологический журнал, 2019, № 6, стр. 16-24
Ископаемые следы жизнедеятельности из переходных отложений венда и кембрия Игарского поднятия (северо-запад Сибирской платформы)
И. В. Коровников a, b, *, В. В. Марусин a, b, Д. А. Токарев a, b, О. Т. Обут a, b
a Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
b Новосибирский государственный университет
Новосибирск, Россия
* E-mail: Korovnikoviv@ipgg.sbras.ru
Поступила в редакцию 05.12.2018
После доработки 19.12.2018
Принята к публикации 17.04.2019
Аннотация
Переходные отложения венда и кембрия Игарского поднятия (северо-запад Сибирской платформы), выделяемые в объеме сухарихинской свиты, являются одним из ключевых объектов для реконструкции развития экосистем в позднем венде. Из немакит-далдынской части разреза (средняя часть сухарихинской свиты) впервые приводится описание ископаемых следов жизнедеятельности Skolithos и Arenicolites, представляющих собой открытые норы обитания организмов в осадке. Высокая плотность и локальная распространенность этих следов в сухарихинской свите свидетельствуют о том, что уже в позднем венде вертикально зарывающимися бентосными организмами активно осваивались мелководно-морские обстановки седиментации и формировались типичные для фанерозойских отложений ихноассоциации (ихнофация Skolithos).
ВВЕДЕНИЕ
Кембрийские отложения Игарского поднятия (северо-запад Сибирской платформы) характеризуются весьма представительной палеонтологической летописью (Розанов и др., 1969, 1992; Репина, 1972; Лучинина и др., 1997; Rowland et al., 1998; Kouchinsky et al., 2010 и др.) и потому являются одним из ключевых объектов, как в региональном, так и в глобальном масштабе, для реконструкции экосистем раннего фанерозоя. Однако выделяемые в пределах Игарского поднятия в объеме сухарихинской свиты переходные отложения от венда к кембрию, за исключением ее верхних 2 метров, весьма скудно охарактеризованы палеонтологически. Отсутствие сменяемости в вендской части разреза типичных для этого интервала ассоциаций мелких скелетных остатков (комплексные зоны Anabarites trisulcatus и Purella antiqua) и ископаемых следов жизнедеятельности делает дискуссионной как внутрирегиональную, так и межрегиональную корреляцию (Розанов и др., 1992; Kouchinsky et al., 2007; Maloof et al., 2010), а также выявление этапности становления бентосных сообществ в позднем венде и раннем кембрии на северо-западе Сибирской платформы (в современных координатах).
Описываемые в данной работе остатки ранее были отмечены в сухарихинской свите в восточном крыле антиклинальной складки, расположенной в 15 км выше по течению от устья р. Сухариха, и предыдущими исследователями указывались без детального описания и фототаблиц (Лучинина и др., 1997). В 2015 г. в ходе полевых работ авторами статьи в западном крыле этой складки в средней части сухарихинской свиты обнаружен представительный комплекс остатков. Детальное изучение морфологии и тафономии позволило не только установить их принадлежность к ископаемым следам жизнедеятельности, но также выявить этологические (поведенческие) особенности и экологические условия обитания поздневендских организмов, их оставлявших.
ГЕОГРАФИЧЕСКОЕ И СТРАТИГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ
Описываемые остатки обнаружены в береговых обнажениях на левом берегу р. Сухарихи в 45 км выше по течению от устья (N 67°12′39.5′′, E 87°19′47.3′′) (рис. 1, а, б). В структурном плане находки расположены в западном крыле антиклинальной складки (рис. 1, в; 2, б). Складка сложена породами переходного интервала венда–кембрия. В пределах антиклинали вскрывается разрез, представленный верхами излучинской, сухарихинской, краснопорожской и шумнинской свитами (рис. 2, б). Излучинская и сухарихинская свита (за исключением верхних 2 м) относятся к венду (Розанов и др., 1992). Находки в верхней части сухарихинской свиты микрофитолитов Nubecularites antis, N. varius, известковых цианобактериальных форм Renalcis sp., Girvanella sp. (Розанов и др., 1992), а также единичные находки трубчатых мелких скелетных остатков Anabarites sp. (Rowland et al., 1998) позволяют отнести ее к немакит-далдынскому региоярусу верхнего венда. Верхние 2 м сухарихинской свиты, а также вышележащие краснопорожская и нижняя часть шумнинской свиты соответствуют томмотскому, атдабанскому и низам ботомского яруса нижнего кембрия на основании представительных комплексов мелкой скелетной фауны, археоциат и трилобитов (Розанов и др., 1969; Rowland et al., 1998) (рис 2, а).
Рис. 1.
а – Сибирская платформа и местоположение изученных находок (треугольником показан фрагмент, детально изображенный на рис. 1, б); б – географическое положение разреза, из которого происходит описываемый комплекс ихнофоссилий (серым прямоугольником выделен фрагмент, детально показанный на рис. 1, в); в – фрагмент геологической карты Игарского поднятия с географическим положением изученных находок. Oбозначения: 1 – обнажение, из которого происходит описываемый комплекс остатков; 2 – разлом; 3 – излучинская свита; 4 – сухарихинская свита; 5 – краснопорожская свита (стратиграфические взаимоотношения свит приведены на рис. 2).
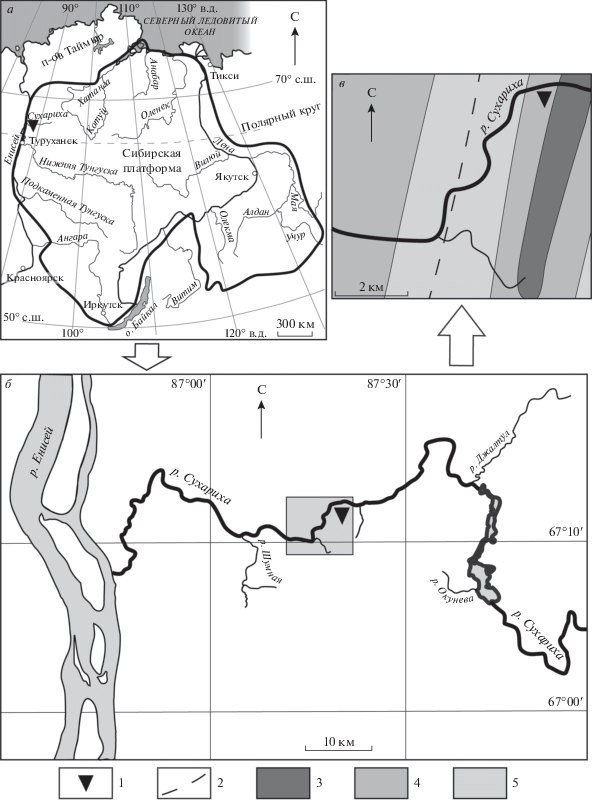
Рис. 2.
а – Сводный разрез переходного интервала венда и кембрия Игарского поднятия и кривая вариаций изотопного состава углерода (δ13C) и индексы изотопных экскурсов (по Kouchinsky et al., 2007); границы подразделений Общей стратиграфической шкалы России (ОСШ) и Международной хроностратигафической шкалы (ICC) приведены по: Лучинина и др., 1997 и Kouchinsky et al., 2007; Maloof et al., 2010, соответственно; их сопоставление приводится на основании данных: Жамойда, 2008; б – геологический профиль по р. Сухариха с местоположением находок ихнофоссилий; используемые для свит цвета соответствуют приведенным на рис. 1, б; в – схематичное изображение взаимоотношений ихнофоссилий с вмещающими бугорчатослоистыми доломитами. Oбозначения: 1 – тонкослоистые алевролиты и аргиллиты; 2 – песчаники горизонтально- и косослоистые; 3 – чередование доломитов и известняков (а также их переходных разностей); 4 – известняки тонкослоистые; 5 – уровни первого появления в сухарихинской свите мелких скелетных остатков Anabarites (240 м выше основания) (а) и комплекса мелких скелетных остатков и археоциат комплексной зоны Nochoroicyathus sunnaginicus томмотского яруса кембрия ОСШ (2 м ниже кровли) (б) по: Лучинина и др., 1997; Rowland et al., 1998. Сокращения: izl – излучинская свита; kp – краснопорожская свита; tom – томмотский ярус; atd – атдабанский ярус.

В настоящее время мировой геологической общественностью активно ведутся исследования, направленные на детализацию геологической летописи в пограничном интервале докембрия и фанерозоя (Peng et al., 2012; Babcock et al., 2014 и др.). Ввиду продолжающейся разработки критериев определения глобальных границ ярусов и отделов кембрия в Международной хроностратиграфической шкале, сопоставление ее с подразделениями раннего кембрия Общей стратиграфической шкалы России является во многом дискуссионным (Розанов и др., 1997; Parkhaev et al., 2011 и др.). На основании сопоставления данных о вариациях изотопного состава углерода в карбонатах в сухарихинской свите с модельной кривой значений δ13Сcarb (Peng et al., 2012) предыдущими исследователями предполагался фортунский (Kouchinsky et al., 2007) (в терминах Международной хроностратиграфической шкалы) возраст как минимум нижних 370 м свиты, ввиду сходного смещения значений изотопного состава углерода от резко отрицательных к положительным значениям вверх по разрезу (Maloof et al., 2010). Поскольку представляемый нами в рамках настоящего исследования материал не позволяет уточнить ни одну из существующих моделей сопоставления международной и отечественной стратиграфических шкал, нами используется схема, утвержденная в 2008 г. Межведомственным стратиграфическим комитетом России (Жамойда, 2008).
Изученные в рамках данной работы остатки обнаружены в средней части сухарихинской свиты, в 284 м выше ее основания, и приурочены к темно-серым бугорчатослоистым доломитам (рис. 2, а). Сухарихинская свита (570 м) представлена преимущественно светло- и темно-серыми доломитами, известняками и доломитовыми известняками, в разной степени глинистыми, с локально распространенными маломощными куполообразными биогермами. В нижней части свиты встречаются прослои и пласты среднезернистых косослоистых песчаников, в верхней встречаются трещины усыхания на кровлях пластов. Формирование сухарихинской свиты происходило в мелководных шельфовых обстановках, в пределах проксимальной зоны карбонатного рампа, ниже приливно-отливной зоны (Rowland et al., 1998).
МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ОСТАТКОВ
Ранее указывались аналогичные единичные находки из средней части сухарихинской свиты в береговых обнажениях по р. Сухариха на восточном крыле антиклинали (Лучинина и др., 1997). Они были интерпретированы как следы жизнедеятельности мягкотелых организмов и сопровождались схематичным изображением найденных остатков. Находки, описываемые в данной работе, найдены в западном крыле антиклинальной складки и демонстрируют как значительно большее морфологическое разнообразие, так и количественную представительность. Было обнаружено большое количество остатков, локализованных в пределах изолированного выхода доломитов средней части свиты, имеющего мощность несколько метров. Слои, в которых встречены изучаемые остатки, представлены темно-серыми доломитами с бугорчатой слоистостью.
Остатки преимущественно представляют собой прямые, слабо изогнутые и редкие U-образные трубчатые формы, ориентированные субвертикально и слабо наклонно. В поперечном сечении остатков четко выделяются две зоны: внутренняя (1–3 мм в диаметре), заполненная крупнокристаллическим кальцитом и внешняя (ширина зоны может достигать 8 мм), представленная мелкокристаллическим доломитом, более светлым, в сравнении с вмещающей породой (табл. III , фиг. 1, 3, 6 ). На некоторых экземплярах внутренняя трубка заполнена кальцитом не до конца, и в ней наблюдается цилиндрическая внутренняя полость (табл. III , фиг. 4 ). Максимальный диаметр внешней зоны достигает 20 мм. Внешняя зона может иметь нечеткий, “размытый” внешний контакт с вмещающей породой (табл. III , фиг. 3–6 ), либо быть отделенной от нее скоплением темных зерен (табл. III , фиг. 7 ). Видимая длина остатков варьирует от первых сантиметров до 15–20 см, однако ввиду крупноплитчатой отдельности вмещающих отложений, установить реальную длину в большинстве случаев не представляется возможным. В продольном сечении периодически встречаются искривления центральной трубки, а также локальные пережимы. Остатки не ветвятся. В пределах слоя они характеризуются высокой частотой встречаемости, однако располагаются изолированно – пересечений, наложений и слияний центральных цилиндрических зон не наблюдается. На некоторых экземплярах отмечается “слияние” внешних зон двух близкорасположенных параллельных трубок.
Объяснение к таблице III
Все образцы из одного местонахождения: северо-запад Сибирской платформы, Игарское поднятие, р. Сухариха; сухарихинская свита, 284 м от основания свиты, немакит-далдынский региоярус верхнего венда.
Фиг. 1. Поперечные сечения Skolithos verticalis (показаны стрелками) на выветренной поверхности напластования бугорчатослоистых доломитов в негативном эпирельефе. Пунктирными линиями показаны внешние контуры диагенетического ореола вокруг нор; экз. ЦСГМ, № 2091/2.
Фиг. 2. Продольные сечения вертикальных и наклонных Skolithos verticalis на выветренном сколе бугорчатослоистых доломитов. Стрелками показаны заполненные крупнокристаллическим кальцитом норы, пунктирными линиями – внешние контуры диагенетического ореола вокруг нор; экз. ЦСГМ, № 2091/3.
Фиг. 3 . Изолированные и со слившимися диагенетическими ореолами (светлые круглые зоны) норы Skolithos verticalis в поперечном сечении (субпараллельно поверхности напластования). Прямоугольником выделен фрагмент, показанный в увеличенном виде на фиг. 4 ; экз. ЦСГМ, № 2091/4.
Фиг. 4 . Увеличенный фрагмент фиг. 3 с поперечными сечениями Skolithos verticalis. Стрелкой указано сечение норы с центральной полостью, не заполненной кальцитом. Белыми прямоугольниками показаны зоны с постепенным переходом от дигенетического ореола к вмещающей породе; экз. ЦСГМ, № 2091/4.
Фиг. 5 . Параллельные, прямые и слабо изгибающиеся субвертикальные норы Skolithos verticalis (показаны стрелками) и окружающие их светлые диагенетические ореолы с нечеткой внешней границей (пунктирные линии) с вмещающей породой в продольном сечении; экз. ЦСГМ, № 2091/5.
Фиг. 6 . Шлиф с поперечным сечением субвертикальной норы Skolithos verticalis. Прямоугольником показана зона постепенного перехода от мелкокристаллического доломита, выполняющего диагенетический ореол вокруг заполненной хорошо раскристаллизованным кальцитом цилиндрической норы (черная стрелка), к вмещающему кристаллическому доломиту; экз. ЦСГМ, № 2091/6.
Фиг. 7 . Шлиф с сечением субпараллельно слоистости наклонно ориентированной норы Skolithos verticalis. Черная стрелка – цилиндрическая нора, заполненная хорошо раскристаллизованным кальцитом. Белый прямоугольник – контакт окружающего нору диагенетического ореола (светлый мелкокристаллический доломит) и вмещающего кристаллического доломита, подчеркнутый скоплением мелких темных зерен; экз. ЦСГМ, № 2091/7.
Фиг. 8 . Поперечное сечение U-образной вертикальной норы Arenicolites isp. на выветренном сколе бугорчатослоистых доломитов. Стрелкой показан короткий фрагмент заполненной кальцитом цилиндрической норы. Пунктирная линия очерчивает границы диагенетического ореола; экз. ЦСГМ, № 2091/1.

Морфологическая простота, частота встречаемости, закономерность пространственного расположения, а также демонстрируемые морфологические признаки позволяют интерпретировать эти остатки как ископаемые следы жизнедеятельности, оставленные бентосными организмами в осадке. Присутствие литологической дифференциации вещества внутри и вне остатка, а также особенности морфологии позволяют предположить, что центральные цилиндрические образования представляют собой открытые норы обитания организмов в субстрате. Это подтверждается перекристаллизацией центральной части норы, что является достаточно распространенной тафономической особенностью для открытых нор в карбонатном осадке (Kemp, 1995; Myrow, 1995). Внешняя же зона, часто с неровной внешней границей, представляет собой ореол – результат диагенетического изменения осадка близ норы, стенки которой были скреплены слизью обитающего в ней организма (Myrow, 1995; Pemberton, Gingras, 2005; Buatois, Mangano, 2011). Принципиально выделяются две морфологические категории: прямые (либо слабоизогнутые) субвертикальные остатки и вертикально ориентированные U-образные формы. Первые отнесены нами к ихнороду Skolithos Haldeman, 1840, вторые – к ихнороду Arenicolites Salter, 1857.
ОПИСАНИЕ МАТЕРИАЛА
Описываемый материал хранится в Центральном Сибирском геологическом музее (ЦСГМ, г. Новосибирск), коллекция № 2091.
В сухарихинской свите норы Arenicolites сравнительно редки – обнаружен единственный экземпляр (ЦСГМ № 2091/1) плохой сохранности. К сожалению, отсутствие представительной выборки экземпляров и неудовлетворительная сохранность материала не позволяют установить его ихновидовую принадлежность в составе ихнорода Arenicolites. Описываемый экземпляр представляет собой U-образную вертикальную нору в негативном эндорельефе, сохранившуюся в виде желобка, оконтуривающего внешнюю зону (диагенетический ореол) вокруг норы, в темно-серых доломитах (табл. III , фиг. 8 ). Диаметр ореола достигает 1 см, переход от ореола к вмещающей породе неровный, постепенный. Видимая глубина норы 3 см, расстояние между осевыми линиями – 2.5 см. Из-за специфики выветривания и сохранности, на экземпляре практически не представлена собственно цилиндрическая нора – присутствует только короткий (7 мм) фрагмент диаметром 3 мм. Она заполнена хорошо раскристаллизованным кальцитом, без признаков пережимов или структурированного заполнения (табл. III , фиг. 5–7 ). Пространство между вертикальными сегментами норы не несет признаков переработки осадка, текстурно и литологически идентично вмещающей породе.
Отсутствие признаков активной переработки пространства между вертикальными сегментами норы (spreite-структур) и бесструктурное ее заполнение позволяют с достаточной уверенностью отличить изучаемый материал от U-образных нор с активной переработкой осадка Diplocraterion Torell, 1870 и отнести их к ихнороду Arenicolites. Следы Arenicolites представляют собой субвертикальные U-образные норы обитания в субстрате (domichnia) бентосных организмов – фильтраторов взвеси (напр., Fürsich, 1975; Pickerill et al., 1984; Bromley, 1996; MacEachern et al., 2012). Они широко распространены в терригенных и карбонатных мелководно-морских шельфовых отложениях с активной гидродинамикой и являются одним из индикаторных таксонов ихнофации Skolithos (Seilacher, 1964; Howard, Frey, 1984; MacEachern et al., 2012; Knaust et al., 2012). В качестве организмов, оставляющих норы Arenicolites, рассматриваются питающиеся взвесью аннелиды и ракообразные (Pickerill et al., 1984). В геологической летописи эти следы получают наиболее широкое распространение с раннего кембрия [безымянный второй ярус кембрия Международной стратиграфической шкалы (МСШ) (Cambrian Stage 2)] (Mangano, Buatois, 2014), однако редкие их находки известны и из верхневендских отложений (Oji et al., 2018).
Многочисленные вертикальные и слабонаклонные цилиндрические норы диаметром 1–3 мм и видимой длиной от первых сантиметров до 15 см (отдельные экземпляры достигают 20 см в длину), встречающиеся в средней части сухарихинской свиты, отнесены к ихновиду Skolithos verticalis (Hall, 1843) (табл. III , фиг. 1–7 ). Норы преимущественно прямые, иногда полого изгибающиеся, либо с флексурными перегибами (табл. III , фиг. 5 ). Норы заполнены крупнокристаллическим кальцитом бесструктурно, иногда центральная часть норы может быть полой (табл. III , фиг. 4 ). Цилиндрические норы повсеместно окружены зоной (диаметр до 2 см), сложенной светлым мелкокристаллическим доломитом (диагенетический ореол). Граница между диагенетическим ореолом и вмещающим темно-серым доломитом преимущественно неровная, постепенная. Однако на некоторых экземплярах граница прослеживается четко и подчеркнута скоплением темных зерен (табл. III , фиг. 7 ). У близкорасположенных нор диагенетические ореолы могут сливаться в один (табл. III , фиг. 1, 3, 4 ). Норы в пределах слоя располагаются изолированно (не соприкасаются), но достаточно плотно. На поверхностях напластования диагенетические ореолы выветриваются сильнее, чем вмещающая порода, и представлены в виде округлых углублений. В центральной части этих углублений незначительно выдается вверх цилиндрическая нора (табл. III , фиг. 1).
От наиболее сходного по морфологии ихновида Skolithos linearis Haldeman, 1840, типового вида ихнорода, S. verticalis отличаются меньшим диаметром (1–4 мм, в сравнении с 3–12 мм у S. linearis) и меньшей глубиной нор. Также диагностическим признаком S. verticalis является их преимущественно субвертикальная ориентировка и минимальная изогнутость.
Наибольшее морфологическое сходство представители ихнорода Skolithos имеют с простыми вертикальными норами Monocraterion Torell, 1870, отличаясь от последних отсутствием воронкообразного расширения в верхней части норы. Вопрос о валидности таксона Monocraterion неоднократно поднимался разными исследователями (Alpert, 1974; Bromley, 1996; Jensen, 1997), поскольку наличие главного диагностического признака этого ихнорода – воронкообразного расширения – во многих случаях может зависеть от условий сохранности изучаемого материала. Ввиду специфики сохранности, изученный материал из сухарихинской свиты не позволяет решить эту проблему. Однако отсутствие в образцах, на которых представлены выходы вертикальных нор на поверхностях напластования (табл. III , фиг. 1, 2), воронкообразных расширений позволяет уверенно отнести эти норы к ихнороду Skolithos. Сходные по типу сохранности остатки были описаны В.А. Асташкиным (1985) из нижнего кембрия (нохоройская пачка, куторгиновая и кетеменская свиты) Анабаро-Синского фациального региона (среднее течение р. Лены и р. Ботомы) как норы Aulophycus repens Fenton et Fenton, 1939. Однако, в отличие от изученного нами материала из сухарихинской свиты Игарского поднятия, описанные Асташкиным формы характеризуются значительно более высокой морфологической вариативностью: помимо прямых и изгибающихся субвертикальных и наклонных сегментов, трубчатые остатки широко распространены и на поверхностях напластования (Асташкин, 1985, табл. LIII , фиг. 2, 3 ), где формируют полихотомически ветвящиеся системы цилиндрических туннелей. Основываясь на морфологии, описанные из нижнекембрийских отложений юго-востока Сибирской платформы остатки, наиболее вероятно, представляют собой трехмерные системы открытых туннелей Thalassinoides Ehrenberg, 1944. В работе А.Ю. Журавлева c соавт. (Zhuravlev et al., 2014) было сделано предположение, что эти остатки могут быть отнесены к ихнороду Ophiomorpha Lundgren, 1891, однако, во-первых, приведенные в работе иллюстрации не позволяют достоверно установить наличие инкрустации стенок нор сферическими пеллетами, являющейся диагностическим признаком этого ихнорода; во-вторых, представители ихнорода Ophiomorpha появляются в ископаемой летописи, лишь начиная с пермского периода (Droser, Bottjer, 1993; Buatois, Mangano, 2011).
Следы Skolithos интерпретируются как пассивно заполненные осадком норы обитания в субстрате (domichnia) питающихся взвесью организмов (suspension feeders) (Alpert, 1974; Häntzschel, 1975; Bromley, 1996). Ввиду морфологической простоты нор, они могут быть образованы чрезвычайно широким спектром морских бентосных организмов. В палеозойских осадочных последовательностях вертикальные норы Skolithos преимущественно приурочены к мелководно-морским отложениям с активной гидродинамикой и постоянным привносом обломочного материала и питательных веществ, являясь ключевым таксоном одноименной ихнофации (Seilacher, 1964; Buatois, Mangano, 2011).
Норы Skolithos чрезвычайно широко распространены в ископаемой летописи бентосных ихносообществ, начиная с основания кембрия (Cambrian Stage 2) (Mangano, Buatois, 2014), однако первые представители известны в отложениях редкинского горизонта верхнего венда Восточно-Европейской платформы (Федонкин, 1985) и немакит-далдынского региояруса верхнего венда (фортунский ярус кембрия МСШ) (McIlroy, Brasier, 2017).
ОБСУЖДЕНИЕ
Локальное обнаружение столь многочисленных нор в сухарихинской свите в пределах монофациального интервала разреза, наиболее вероятно, обусловлено, с одной стороны, фрагментированной обнаженностью этого интервала в пределах западного крыла антиклинали, в то время как характер обнаженности свиты на восточном крыле (относительно непрерывная обнаженность круто и субвертикально залегающих слоев, часто амальгамированных в мощные пакеты и пласты) и ограниченный доступ к поверхностям напластования крайне затрудняет поиск и диагностику ископаемых следов жизнедеятельности. Во-вторых, локальное распространение столь многочисленных Skolithos и Arenicolites в сухарихинской свите может быть связано с увеличивающейся степенью перекристаллизации карбонатных отложений вверх по разрезу, ухудшающей сохранность нор. Наиболее вероятно, это ихносообщество распространено во всей свите, что подтверждается единичными находками предыдущих исследователей, приведенных без описания и иллюстраций (Лучинина и др., 1997).
Находки Skolithos и Arenicolites в сухарихинской свите согласуются с ранее сформулированной фациальной интерпретацией этого интервала (Rowland et al., 1998): в фанерозойских отложениях в пределах карбонатного рампа его проксимальную часть характеризуют сообщества ихнофации Skolithos, представленные разнообразными морфологически простыми норами обитания питающихся взвесью организмов (Skolithos, Arenicolites, Ophiomorpha, Psilonichnus, Rhizocorallium и др.), сформированными в обстановках с высокой гидродинамической активностью среды (Knaust et al., 2012). Развитие эндобентосных сообществ и взаимоотношений организмов и субстрата в пограничном интервале позднего докембрия и кембрия носит название “Первой агрономической революции” (Seilacher, Pflüger, 1994; Seilacher, 1999). В череде усложнений типов переработки осадка роющими организмами поздневендский этап (немакит-далдынский региоярус), приблизительно отвечающий фортунскому ярусу кембрия МСШ, характеризуется широким распространением разнообразных горизонтальных нор переработки осадка. Вертикальные же норы обитания сравнительно редки, их распространение постепенно увеличивается ближе к началу второго яруса кембрия (~530 млн лет) (Mangano, Buatois, 2017), условно отвечающему началу томмотского века Общей стратиграфической шкалы России (Parkhaev et al., 2011; Zhuravlev, Wood, 2018). Дальнейшая колонизация питающихся взвесью седентарных организмов мелководно-морских обстановок в кембрии привело к формированию осадочных пород со специфическими текстурными характеристиками, т. н. “трубчатых пород” (pipe-rocks) (Desjardins et al., 2010). Находки многочисленных Skolithos в сухарихинской свите Игарского поднятия показывают, что уже в немакит-далдынское время в мелководно-морских обстановках с карбонатной седиментацией началось формирование типичных фанерозойских рекуррентных ихноассоциаций мелководного шельфа (ихнофация Skolithos), пусть и в ограниченном масштабе, в сравнении с их более молодыми аналогами.
Список литературы
Асташкин В.А. Проблематические организмы – породообразователи в нижнекембрийских отложениях Сибирской платформы // Проблематики позднего докембрия и палеозоя. М.: Наука, 1985. С. 144–149 (Тр. ИГиГ СО АН СССР. Вып. 632).
Жамойда А.И. Состояние изученности стратиграфии докембрия и фанерозоя России. Задачи дальнейших исследований. СПб.: ВСЕГЕИ, 2008. 131 с. (Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Вып. 38.).
Лучинина В.А., Коровников И.В., Сипин Д.П., Федосеев А.В. Биостратиграфия верхнего венда – нижнего кембрия разреза р. Сухариха (Сибирская платформа) // Геол. и геофиз. 1997. Т. 38. № 8. С. 1346–1358.
Репина Л.Н. К вопросу о развитии трилобитов семейства Protolenidae (Сибирь) // Проблемы биостратиграфии и палеонтологии нижнего кембрия Сибири. М: Наука, 1972. С. 15–30.
Розанов А.Ю., Миссаржевский В.В., Волкова Н.А. и др. Томмотский ярус и проблема нижней границы кембрия. М.: Наука, 1969. 380 с. (Тр. Геол. ин-та АН СССР. Вып. 206).
Розанов А.Ю., Репина Л.Н., Аполлонов М.К. и др. Кембрий Сибири. Новосибирск: Наука, 1992. 134 с.
Розанов А.Ю., Семихатов М.А., Соколов Б.С. и др. Решение о выборе стратотипа границы докембрия и кембрия: прорыв в проблеме или ошибка? // Стратигр. Геол. корреляция. 1997. Т. 5. № 1. С. 21–31.
Федонкин М.А. Палеоихнология вендских Metazoa // Вендская система. Историко-геологическое и палеонтологическое обоснование. Т. 1. Палеонтология. М.: Наука, 1985. С. 112–117.
Alpert S.P. Systematic review of the genus Skolithos // J. Paleontol. 1974. V. 48. P. 661–669.
Babcock L.E., Peng S., Zhu M. et al. Proposed reassessment of the Cambrian GSSP // J. Afr. Earth Sci. 2014. V. 98. P. 3–10.
Bromley R.G. Trace fossils. Biology, taphonomy and applications. 2nd ed. L.: Chapman & Hall, 1996. 361 p.
Buatois L.A., Mángano M.G. Ichnology. Organism–substrate interactions in space and time. Cambridge: Cambridge Univ. Press, 2011. 358 p.
Desjardins P.R., Mángano M.G., Buatois L.A., Pratt B.R. Skolithos pipe rocks and associated ichnofabrics from the southern Rocky Mountains, Canada: colonization trends and environmental controls in an early Cambrian sand-sheet complex // Lethaia. 2010. V. 43. P. 507–528.
Droser M.L., Bottjer D.J. Trends and patterns of Phanerozoic ichnofabrics // Annu. Rev. Earth Planet. Sci. 1993. V. 21. P. 205–225.
Fürsich F.T. Trace fossils as environmental indicators in the Corallian of England and Normandy // Lethaia. 1975. V. 8. P. 151–172.
Häntzschel W. Trace fossils and problematica // Treatise on Invertebrate Paleontology. Part W. Miscellanea. Suppl. 1. 2nd ed. (revised and enlarged) / Ed. Teichert C. Lawrence: Geol. Soc. Amer.; Univ. Kansas Press, 1975. 269 p.
Howard J.D., Frey R.W. Characteristic trace fossils in nearshore to offshore sequences, Upper Cretaceous of east-central Utah // Can. J. Earth Sci. 1984. V. 21. P. 200–219.
Jensen S. Trace fossils from the Lower Mickwitzia sandstone, south-central Sweden // Fossils and Strata. 1997. № 42. 110 p.
Kemp A.E.S. Variation of trace fossils and ichnofacies in Neogene and Quaternary pelagic sediments from the eastern equatorial Pacific Ocean (Leg 138) // Proc. Ocean Drilling Program, Sci. Results. Texas: College Station, 1995. V. 138. P. 177–190.
Knaust D., Curran H.A., Dronov A.V. Shalow-marine carbonates // Trace Fossils as Indicators of Sedimentary Environments. Amsterdam: Elsevier, 2012. P. 705–750 (Developments in Sedimentology. V. 64).
Kouchinsky A., Bengtson S., Murdock D.J.E. A new tannuolinid problematic from the lower Cambrian of the Sukharikha River in northern Siberia // Acta Palaeontol. Pol. 2010. V. 55. № 2. P. 321–331.
Kouchinsky A., Bengtson S., Pavlov V. et al. Carbon isotope stratigraphy of the Precambrian–Cambrian Sukharikha River section, northwestern Siberian platform // Geol. Mag. 2007. V. 144. P. 609–618.
MacEachern J.A., Bann K.L., Gingras M.K. et al. The ichnofacies paradigm // Trace Fossils as Indicators of Sedimentary Environments. Amsterdam: Elsevier, 2012. P. 103–138 (Developments in Sedimentology. V. 64).
Maloof A.C., Porter S.M., Moore J.L. et al. The earliest Cambrian record of animals and ocean geochemical change // GSA Bull. 2010. V. 122. P. 1731–1774.
Mángano M.G., Buatois L.A. Decoupling of body-plan diversification and ecological structuring during the Ediacaran–Cambrian transition: evolutionary and geobiological feedbacks // Proc. Roy. Soc. Lond. Ser. B. 2014. V. 281. 20140038.
Mángano M.G., Buatois L.A. The Cambrian revolutions: Trace-fossil record, timing, links and geobiological impact // Earth-Sci. Rev. 2017. V. 173. P. 96–108.
McIlroy D., Brasier M.D. Ichnological evidence for the Cambrian Explosion in the Ediacaran to Cambrian of Tanafjord, Finnmark, northern Norway // Earth System Evolution and Early Life: a Celebration of the Work of Martin Brasier / Eds. Brasier A.T., McIlroy D., McLoughlin N.L., 2017. P. 351–368 (Geol. Soc. Lond. Spec. Publ. № 448).
Myrow P.M. Thalassinoides and the enigma of Early Paleozoic open-framework burrow systems // Palaios. 1995. V. 10. P. 58–74.
Oji T., Dornbos S.Q., Yada K. et al. Penetrative trace fossils from the late Ediacaran of Mongolia: early onset of the agronomic revolution // Roy. Soc. Open Sci. 2018. V. 5. 172250.
Parkhaev P.Yu., Karlova G.A., Rozanov A.Yu. Taxonomy, stratigraphy and biogeography of Aldanella attleborensis – a possible candidate for defining the base of Cambrian Stage 2 // Bull. Mus. Northern Arizona. 2011. № 76. P. 298–300.
Pemberton S.G., Gingras M.K. Classification and characterizations of biogenically enhanced permeability // AAPG Bull. 2005. V. 89. P. 1493–1517.
Peng S.C., Babcock L.E., Cooper R.A. The Cambrian Period // The Geological Time Scale. Amsterdam: Elsevier, 2012. P. 437–488.
Pickerill R.K., Fillion D., Harland T.L. Middle Ordovician trace fossils in carbonates of the Trenton Group between Montreal and Quebec City, St. Lawrence Lowland, eastern Canada // J. Paleontol. 1984. V. 58. P. 416–439.
Rowland S.M., Luchinina V.A., Korovnikov I.V. et al. Biostratigraphy of the Vendian-Cambrian Sukharikha River section, northwestern Siberian Platform // Can. J. Earth Sci. 1998. V. 35. P. 339–352.
Seilacher A. Biogenic sedimentary structures // Approaches to Paleoecology. N. Y.: Wiley, 1964. P. 296–316.
Seilacher A. Biomat-related lifestyles in the Precambrian // Palaios. 1999. V. 14. P. 86–93.
Seilacher A., Pflüger F. From biomats to benthic agriculture: A biohistoric revolution // Biostabilization of Sediments. Odenburg: Bibliotheks und Informationssystem der Carl von Ossietzky Universität, 1994. P. 97–105.
Zhuravlev A.Yu., Gámez Vintaned J.A., Tabachnick K.R. Comment on “An enigmatic, possibly chemosymbiotic, hexactenellid sponge from the early Cambrian of South China” by Joseph P. Botting, Lucy A. Muir, Xiang-Feng Li, and Jin-Pai Lin // Acta Palaeontol. Pol. 2014. V. 59. P. 475–476.
Zhuravlev A.Yu., Wood R.A. The two phases of the Cambrian Explosion // Nature Sci. Reports. 2018. V. 8. 16656.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал