

Палеонтологический журнал, 2019, № 6, стр. 73-78
Новые виды конодонтов из фамена (верхний девон) Урала
А. Н. Плотицын a, *, Ю. А. Гатовский b, **
a Институт геологии им. академика Н.П. Юшкина Коми НЦ УрО РАН
Сыктывкар, Россия
b Московский государственный университет имени М.В. Ломоносова
Москва, Россия
* E-mail: anplotitzyn@rambler.ru
** E-mail: ustas62@bk.ru
Поступила в редакцию 03.12.2018
После доработки 21.01.2019
Принята к публикации 21.01.2019
Аннотация
Из верхнего фамена Урала и Приуралья описаны новые виды конодонтов рода Polygnathus. Po. tsygankoi sp. nov. распространен в зонах expansa–praesulcata и занимает промежуточное положение в филетической линии Polygnathus atasuensis–Po. tsygankoi sp. nov.–Po. lobatus sensu Kononova. Вид Po. serriformis sp. nov. встречен в зоне praesulcata. Эволюционные связи вида Po. serriformis sp. nov. не установлены.
Пограничным отложениям девонской и каменноугольной систем в настоящее время уделяется пристальное внимание в связи с выбором нового лимитотипа границы и, как следствие, пересмотром распространения основных групп фауны (в том числе конодонтов) в пределах этого стратиграфического интервала (Kaiser, Becker, 2007; Kaiser, Corradini, 2008; Corradini et al., 2016). Столь высокая актуальность предопределила повышение степени детальности биостратиграфических исследований в пограничных отложениях как в стратотипической местности, так и в серии разрезов различных регионов мира.
На западном склоне Северного Урала и в Приуралье граница между девонской и каменноугольной системами с различной степенью детальности охарактеризована конодонтами в ряде разрезов на реках Кожим, Сывъю, Вангыр, Подчерем (ур. Кузьяма), Изъяю, Каменка и Изъяель (Плотицын и др., 2018). Детализация биостратиграфических исследований в этом регионе привела не только к уточнению стратиграфического распространения уже известных представителей ряда конодонтовых родов (Плотицын, Груздев, 2016), но и к выделению новых видов Polygnathus tsygankoi sp. nov. и Po. serriformis sp. nov., описания которых приведены в настоящей работе.
Находки конодонтов Po. tsygankoi sp. nov. отмечены только в разрезе на р. Сывъю (Приполярный Урал) и ограничены зонами Late expansa и praesulcata фаменского яруса (пробы C-22, C-23, C-28 и C-40) (Плотицын и др., 2014). В пределах зоны Late expansa совместно с новым видом встречены Apatognathus varians Branson et Mehl, Bispathodus stabilis (Branson et Mehl) M1, B. stabilis (Branson et Mehl) M2, Branmehla disparilis (Branson et Mehl), Br. inornata (Branson et Mehl), Br. aff. werneri (Ziegler), Neopolygnathus communis (Branson et Mehl), Palmatolepis gracilis expansa Ziegler et Sandberg, Pa. gracilis gracilis Branson et Mehl, Pa. gracilis sigmoidalis Ziegler, Polygnathus vogesi Ziegler, Pseudopolygnathus marburgensis trigonicus Ziegler, Rhodalepis polylophodontiformis Wang et Yin, а также представители рода Mitrellataxis. В зоне praesulcata основу комплекса (слой с пробой С-40) слагают представители рода Neopolygnathus – N. carinus (Hass), N. communis и N. dentatus (Druce), в подчиненном количестве присутствуют Bispathodus aculeatus (Branson et Mehl), Protognathodus collinsoni Ziegler, Siphonodella praesulcata Sandberg и переходящие из зоны expansa Apatognathus varians, Bispathodus stabilis M1, Branmehla inornata, Palmatolepis gracilis gracilis, Pa. gracilis sigmoidalis, Polygnathus serriformis sp. nov. и Po. vogesi.
На Южном Урале Po. tsygankoi sp. nov. обнаружен в лытвинском горизонте в разрезе Сиказа (слои 4–6) (Артюшкова и др., 2011). Новый вид встречен в комплексе конодонтов, характеризующем верхнефаменские зоны Middle‑Late expansa: Apatognathus varians, Bispathodus aculeatus, Bi. stabilis, Neopolygnathus communis, Palmatolepis gracilis expansa, Pa. gracilis gracilis, Pa. gracilis sigmoidalis, Polygnathus delicatulus Ulrich et Bassler, Po. inornatus, Po. vogesi, Po. znepolensis, Pseudopolygnathus brevipennatus, Ps. marburgensis trigonicus и Rhodalepis polylophodontiformis.
В коллекционном материале В.Н. Пазухина (ИГ УФИЦ РАН, Уфа) Po. tsygankoi sp. nov. встречается в верхнефаменских отложениях (зоны Middle‑Late expansa) западного склона Юж. Урала в разрезах Сиказа, Зиган, Ряузяк, Усуйли, Мурадымово, а также на восточном склоне в разрезе Дзержинка-Поповский и Магнитогорской скважине 2016.
Новый вид Po. serriformis sp. nov. встречен в разрезах на реках Сывъю на Приполярном Урале (проба С-40) и Изъяель в южной части гряды Чернышева (проба GER3-92ф) (Плотицын, Груздев, 2016) совместно с комплексами конодонтов, характеризующими зону praesulcata фаменского яруса. Состав комплекса конодонтов разреза на р. Сывъю указан выше. В разрезе на р. Изъяель новый вид входит в состав таксономически более богатого комплекса, где основу слагают Bispathodus stabilis M1, Neopolygnathus communis, Palmatolepis gracilis sigmoidalis, Protognathodus collinsoni, Pr. meischneri Ziegler, а также многочисленные экземпляры, демонстрирующие морфологический переход между Bispatodus stabilis M1 и Protognathodus meischneri. В подчиненном количестве присутствуют Apatognathus varians, Bispathodus aculeatus aculeatus, Bi. stabilis M1, Branmehla disparilis, Br. inornata, Br. werneri, Palmatolepis gracilis expansa, Pa. gracilis gracilis, Polygnathus delicatus, Po. parapetus Druce, Po. vogesi, Protognathodus kockeli (Bischoff), Pseudopolygnathus marburgensis trigonicus и Siphonodella praesulcata.
На Южном Урале Po. serriformis sp. nov. обнаружен в нижней части гумеровского горизонта в разрезе Сиказа (слой 6в). Описание разреза опубликовано О.В. Артюшковой с соавт. (2011). Новый вид встречен с комплексом конодонтов, характеризующим верхнефаменскую зону praesulcata: Bispathodus aculeatus aculeatus, Bi. stabilis, Neopolygnathus communis, Palmatolepis gracilis sigmoidalis, Pelekysgnathus sp., Polygnathus ex gr. inornatus, Protognathodus collinsoni, Pr. meischneri.
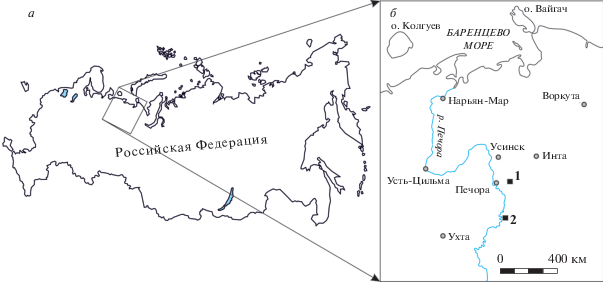
Схема расположения разрезов, из которых происходят голотипы новых видов Polygnathus tsygankoi sp. nov. и Polygnathus serriformis sp. nov., показана на рис. 1. Более подробная характеристика рассматриваемых разрезов, в которых найдены новые виды, опубликована ранее (Артюшкова и др., 2011; Плотицын и др., 2014; Плотицын, Груздев, 2016).
Рис. 1.
Схема расположения разрезов, из которых происходят голотипы новых видов Polygnathus tsygankoi sp. nov. и Po. serriformis sp. nov.: а – обзорная схема; б – детальная схема: 1 – разрез на р. Сывъю; 2 – разрез на р. Изъяель.
При описании новых видов рассматривались только Pa-элементы, так как корректная реконструкция аппаратов не представляется возможной по причине обедненности комплексов конодонтов рамиформными элементами. Надродовая систематика основана на классификации У. Свита (Sweet, 1988).
Коллекции конодонтов хранятся в музее им. А.А. Чернова в Ин-те геологии им. акад. Н.П. Юшкина Коми НЦ УрО РАН (Музей ИГ, Сыктывкар) под №№ 333/24, 333/26, 512/7, 512/9 и 512/12, а также на каф. палеонтологии геологического фак-та МГУ им. М.В. Ломоносова под № 359. Фотографирование конодонтовых элементов производилось на сканирующих электронных микроскопах VEGA3 TESCAN (Ин-т геологии Коми НЦ УрО РАН) и VEGA2 TESCAN (Палеонтологический ин-т им. А.А. Борисяка РАН).
Авторы выражают глубокую благодарность А.С. Алексееву, А.В. Журавлеву и Л.И. Кононовой за ценные советы и замечания, способствовавшие улучшению статьи; С.С. Шевчуку и А.С. Шуйскому за качественные фотографии конодонтов на сканирующем электронном микроскопе.
Исследование выполнено при частичной финансовой поддержке РФФИ в рамках научного проекта № 18-35-00330.
КЛАСС CONODONTA
ОТРЯД OZARKODINIDA
СЕМЕЙСТВО POLYGNATHIDAE BASSLER, 1925
Род Polygnathus Hinde, 1900
Polygnathus tsygankoi Plotitsyn et Gatovsky, sp. nov.
Табл. XI , фиг. 1–8 (см. вклейку)
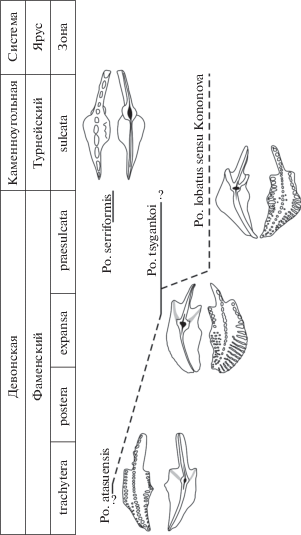
Рис. 2.
Стратиграфическое распространение новых видов Polygnathus tsygankoi sp. nov. и Po. serriformis sp. nov., а также предполагаемые филоморфогенетические связи Po. tsygankoi sp. nov.
Polygnathus aff. semicostatus Branson et Mehl, 1934: Плотицын и др., 2014, рис. 4 , фиг. 34 .
Название вида – в честь В.С. Цыганко.
Голотип – Музей ИГ, колл. № 512/12, правосторонний элемент; Приполярный Урал, среднее течение р. Сывъю (приблизительно в 6 км от устья р. Западная Сывъю вверх по течению реки), проба С-22; верхний девон, фаменский ярус, конодонтовая зона Late expansa (табл. XI , фиг. 1).
Описание. Карминипланатный Pa-элемент с массивной, асимметричной, коробчатой в передней и субтреугольной в задней части аркообразно изогнутой платформой. Передний конец в плане (вид сверху) прямой или округлый с угловатыми заостренными боковыми краями. Задний конец заострен. Боковые края платформы в передней части высоко подняты, с образованием глубоких адкаринальных трогов. Троги полностью выполаживаются к заднему языковидному плоскому концу платформы. Боковые края платформы в передней ее части ростроподобно сужаются и лопастевидно расширяются в средней части. Наиболее хорошо лопастеподобное расширение выражено на внешней стороне платформы правосторонних элементов; платформа правосторонних элементов шире, чем у левосторонних. У правосторонних элементов выше поднят внешний край платформы, а у левосторонних – внутренний. Орнаментация верхней стороны платформы представлена многочисленными длинными тонкими поперечными ребрами или рядами бугорков, собранных в поперечные ребра, которые почти достают осевого гребня, но не сливаются с ним. Осевой гребень дугообразно изогнут и достигает заднего конца платформы. Короткий и высокий передний свободный лист составляет 1/4–1/5 длины платформы. Базальная ямка узкая, щелевидная, с широкими флангами, располагается в передней трети платформы. От флангов базальной ямки в стороны угловатых передних концов платформы отходят валики. Фланги базальной ямки имеют оттянутые в сторону валиков угловатые края, что придает флангам субромбическую форму. Эти угловатые края, вероятно, намечают зачаточные короткие дополнительные кили. Киль невысокий, четкий. Его высота незначительно снижается позади базальной ямки.
Асимметричные по очертаниям Pa-элементы Po. tsygankoi sp. nov. на всех стадиях онтогенеза демонстрируют в аппарате зеркальную асимметрию, что дает основание отнести их классу III симметрии (по Lane, 1968).
Онтогенез. В процессе перехода к поздним возрастным стадиям Pa-элементы Po. tsygankoi sp. nov. приобретают более асимметричные очертания платформы. Благодаря появлению отчетливого рострообразного сужения передней части платформы, в срединной части в месте выполаживания боковых краев происходит формирование лопастевидного расширения.
Сравнение. Po. tsygankoi sp. nov. наиболее сходен с Po. inornatus Branson, 1934, который известен из верхов фамена–турне (зоны expansa–anchoralis). Новый вид отличается от Po. inornatus более асимметричной платформой, наличием “зачаточных” боковых килей и хорошо выраженным рострообразным сужением передней части платформы. Также Pa-элементы рассматриваемого вида сходны с таковыми Po. lacinatus Huddle, 1934, но отличаются более асимметричной и изогнутой в плане платформой, языковидным плоским задним концом платформы и “зачаточными” дополнительными килями на флангах базальной ямки. Po. lacinatus распространен в зонах praesulcata–anchoralis от самых верхов фамена до конца турне. На поздних возрастных стадиях очертания и орнаментация верхней стороны платформы нового вида сходны с представителями турнейской группы Po. lobatus Branson et Mehl, 1934, в основном благодаря наличию лопастевидного расширения средней части платформы. Однако для группы Po. lobatus не характерно наличие “зачаточных” дополнительных килей и языковидного плоского заднего конца платформы. Удлиненно-языковидная платформа с приподнятыми в передней части краями, которые выполаживаются назад, характеризует фаменские виды Po. semicostatus Branson et Mehl, 1934 (распространен в зонах crepida–expansa) и Po. obliquicostatus Ziegler, 1962 (распространен в зонах postera–expansa). Однако оба эти вида не характеризуются наличием “зачаточных” дополнительных килей. Кроме того, от Po. semicostatus Po. tsygankoi sp. nov. отличается осевым гребнем, достигающим заднего конца платформы, а также присутствием хорошо выраженного ростроподобного сужения и узкими флангами базальной ямки. От Po. obliquicostatus отличается отсутствием косой ребристости на внутренней стороне платформы, наличием флангов базальной ямки, отчетливо выраженного ростроподобного сужения и более грубой ребристой орнаментацией.
Наиболее вероятной предковой формой Po. tsygankoi sp. nov. является вид Po. atasuensis Kuzmin, 1992 (рис. 2) из среднефаменских отложений Атасуйского рудного района в Центральном Казахстане (Кузьмин, 1992). При общем сходстве очертаний платформы новый вид отличается от Po. atasuensis более деликатной и упорядоченной орнаментацией, представленной длинными поперечными ребрами, почти достигающими осевого гребня, и наличием зачаточных дополнительных килей.
Предполагается, что потомком Po. tsygankoi sp. nov. является Po. lobatus sensu Kononova (рис. 2) из пограничных фаменско-турнейских отложений (топкинская толща, абышевский горизонт) юга Западной Сибири (Бушмина, Кононова, 1981). При общем сходстве очертаний платформы и орнаментации верхней стороны ее у экземпляров, изображенных Л.С. Бушминой и Л.И. Кононовой (1981, табл. 19 , фиг. 1‑4 ), Po. tsygankoi sp. nov. демонстрирует присутствие валиков и небольших “зачаточных” дополнительных килей, незначительно выдающихся за пределы округлых флангов базальной ямки на всех стадиях онтогенеза, тогда как у Po. lobatus sensu Kononova от флангов базальной ямки в угловатые концы краев платформы отходят два полноценных дополнительных киля.
Замечания. Pa-элементы нового вида и его вероятного потомка, благодаря разности высоты краев внешней и внутренней сторон платформы, а также наличию зачатков или полноценных дополнительных килей, демонстрируют некоторое морфологическое сходство с представителями рода Mashkovia Aristov, Gagiev et Kononova (Аристов и др., 1983; Belka, 1998). Однако присутствие широких флангов базальной ямки и наличие поперечной ребристости, а также отсутствие в передней части платформы одного или двух краевых выдающихся продольных шиповатых гребней, характерных для представителей рода Mashkovia, позволяют отнести новый вид к роду Polygnathus.
Распространение. Верхний девон, фаменский ярус, конодонтовая зона Middle–Late expansa: зап. склон Приполярного Урала, разрез Сывъю; зап. склон Южного Урала, разрезы Сиказа, Зиган, Ряузяк, Усуйли, Мурадымово; вост. склон Южного Урала, разрез Дзержинка-Поповский и скв. Магнитогорская 2016.
Материал. 32 экз. хорошей сохранности: 12 экз. из разреза на р. Сывъю; 8 экз. из обр. С-22, 2 экз. из обр. С-23, 1 экз. из обр. С-28 и 1 экз. из обр. С-40 и 20 экз. из разреза Сиказа; 8 экз. из сл. 4, обр. 2; 4 экз. из сл. 5, обр. 5/2 и 8 экз. из сл. 6, обр. 6/12.
Polygnathus serriformis Plotitsyn et Gatovsky, sp. nov.
Табл. XI , фиг. 9–14
Название вида serriformis лат. – пиловидный.
Голотип – Музей ИГ, колл. № 512/7-153, левосторонний элемент; юж. часть гряды Чернышева, среднее течение р. Изъяель (левый приток р. Косью), обр. GER3-92ф; верхний девон, фаменский ярус, зона Late praesulcata (табл. XI , фиг. 9 ).
Описание. Карминипланатный Pa-элемент c маленькой, относительно тонкой, слабоорнаментированной, в незначительной степени асимметричной, каплевидной (на ранних стадиях роста) или коробчато-каплевидной, коробчатой (на поздних стадиях роста) платформой. При виде сбоку элемент коленчато изогнут с точкой перегиба, расположенной позади базальной ямки. Угол изгиба элемента (угол между передним и задним отростком) составляет 155°–160°. Передний конец платформы в плане округлый. Задний конец плавно заостряется и переходит в задний свободный лист. Боковой край внутренней стороны платформы незначительно поднят из-за наличия 2–3 зубцов. Внешняя сторона в большинстве случаев плоская и лишена каких-либо признаков орнаментации. Исключение составляют экземпляры, которые на поздних возрастных стадиях могут на внешней стороне нести 1–2 неотчетливых зубца, формирующих незначительное поднятие края платформы. Орнаментация на внутренней стороне платформы выражена лучше, чем на внешней стороне, как у левосторонних, так и у правосторонних элементов. В целом, за исключением зубцов на боковых краях платформы, остальная часть верхней стороны платформы гладкая. Осевой гребень имеет слабый дугообразный изгиб. Карина и задний свободный лист характеризуются близко расположенными друг к другу или сросшимися зубцами. В зависимости от стадии онтогенеза длина заднего свободного листа может колебаться от 3/4 до 1/2 длины платформы. Передний свободный лист имеет разреженную зубчатость. Под разреженной зубчатостью свободного листа авторами понимается, когда разъединенные зубцы не слиты друг с другом на протяжении всей своей длины, при этом промежуток между зубцами сопоставим с 1/2–1 ширины зубца. Длина переднего свободного листа превышает длину платформы в 2–3 раза. Задний свободный лист обычно составляет 1/3–1/2 длины платформы, изредка их размеры одинаковы. Узкая, щелевидная базальная ямка с широкими флангами располагается в передней части платформы. Киль четкий, непрерывный, его высота крайне незначительно снижается в месте перегиба платформы позади базальной ямки.
Асимметричные по очертаниям Pa-элементы нового вида на всех стадиях онтогенеза демонстрируют в аппарате зеркальную симметрию, что дает основание отнести их к классу II симметрии (по Lane, 1968).
Онтогенез. В онтогенезе Pa-элемент Po. serriformis sp. nov. претерпевает значительные преобразования. Если на самых ранних возрастных стадиях платформа полностью лишена какой-либо орнаментации и имеет каплевидную форму, то на более поздних стадиях платформа постепенно расширяется и приобретает угловатые очертания, при этом обосабливается задний свободный лист и появляется орнаментация в виде зубцов по краям платформы.
Сравнение. Морфологически с Po. serriformis sp. nov. наиболее сходны виды Po. zikmundovae Zhuravlev, 1991 и Po. restrictus Vorontsova, 1993, которые распространены в зонах praesulcata–duplicata и postera–praesulcata, соответственно. От этих видов Po. serriformis sp. nov. отличается более длинными по отношению к платформе передним и задним свободными листами, разреженной зубчатостью переднего листа, более округлой платформой и широкими флангами базальной ямки. У нового вида, в отличие от Po. restrictus, менее развита орнаментация верхней стороны платформы. Po. restrictus орнаментирован короткими ребрами, формирующими бугорки на всем протяжении обоих краев платформы. При этом, в отличие от Po. zikmundovae с гладкой платформой, лишенной скульптуры, орнаментация Po. serriformis sp. nov. развита лучше. Po. serriformis sp. nov. на поздних стадиях онтогенеза сходен с Po. brevilaminus Branson et Mehl, 1934, который встречается от верхов франского яруса до зоны marginifera фаменского яруса. От Po. brevilaminus новый вид отличается меньшей длиной заднего свободного листа, разреженной зубчатостью переднего свободного листа и расположением базальной ямки впереди платформы.
Замечания. Близкородственные виду Po. serriformis sp. nov. формы достоверно не установлены (рис. 2). Однако, учитывая слабую развитость платформы у Pa-элемента нового вида и ее незначительные по сравнению с осевым гребнем размеры, можно сделать предположение о его происхождении от таксонов с листовидными “спатогнатидными” Pa-элементами. Сходной разреженной зубчатостью Pa-элементов характеризуются некоторые представители семейства Priniodinidae Bassler (Dzik, 2006). Яркими примерами подобных морфологических инноваций, связанных с образованием платформы на листовидном элементе, являются обособление от представителей рода Mehlina вида Youngquistognathus angustidiscus (Youngquist, 1945) через переходную форму Y. praeangustidiscus Zhuravlev, 2015 (Журавлев, 2015) во фране или вида Polynodosus lauriformis (Dreesen et Dusar, 1974) через P. transitus Dzik, 2006 (Dzik, 2006) в фамене.
Благодаря небольшим размерам платформы и Pa-элемента в целом, Po. serriformis sp. nov. демонстрирует некоторое морфологическое сходство с позднефаменско-раннетурнейскими представителями рода Neopolygnathus со слабоскульптированной платформой [напр., Neopolygnathus communis (Branson et Mehl, 1934) и Ne. quadratus (Wang, 1989)]. От этих представителей рода Neopolygnathus новый вид отличается отсутствием депрессии позади базальной ямки и наличием не характерной для неополигнатид разреженной зубчатости переднего свободного листа.
Распространение. Верхний девон, фаменский ярус, зона praesulcata: Приполярного Урала, разрез Сывъю; юж. часть гряды Чернышева, разрез Изъяель и зап. склон Юж. Урала, разрез Сиказа.
Материал. 28 экз. различной степени сохранности: 2 экз. из разреза на р. Сывъю (обр. С-40), 16 экз. из разреза на р. Изъяель (обр. GER3-92ф) и 10 экз. из разреза Сиказа.
Список литературы
Аристов В.А., Гагиев М.Х., Кононова Л.И. Филоморфогенез и стратиграфическое значение рода Mashkovia gen. nov. (Conodontophorida) // Изв. АН СССР. Cер. геол. 1983. № 2. С. 72–83.
Артюшкова О.В., Маслов В.А., Пазухин В.Н. и др. Типовые разрезы девона и нижнего карбона на западном склоне Южного Урала: Путеводитель полевой экскурсии перед международной конференцией “Биостратиграфия, палеогеография и события в девоне и раннем карбоне”, Уфа, Стерлитамак, Россия, 20–25 июля 2011. Уфа, 2011. 92 с.
Бушмина Л.С., Кононова Л.И. Микрофауна и биостратиграфия пограничных слоев девона и карбона (юг Западной Сибири). М.: Наука, 1981. 124 с.
Журавлев А.В. Происхождение и эволюция рода Younquistognathus Myshkina et Zhuravlev (конодонты, поздний девон) // Вестн. Ин-та геол. Коми НЦ УрО РАН. 2015. № 1. С. 3–6.
Кузьмин А.В. Использование конодонтов для расчленения фаменских отложений Атасуйского района (Центральный Казахстан) // Изв. АН СССР. Сер. геол. 1992. № 8. С. 30–40.
Плотицын А.Н., Груздев Д.А. Граница девонской и каменноугольной систем на р. Изъяель (юг гряды Чернышева) // Геология, геоэкология и ресурсный потенциал Урала и сопредельных территорий: Матер. VI Всерос. молодеж. конф / Ред. Ардисламов Ф.Р., Сначев А.В. Уфа, 2016. С. 102–107.
Плотицын А.Н., Журавлев А.В., Соболев Д.Б. и др. Граница девона и карбона на западном склоне севера Урала и Приуралья // Тр. Палеонтол. об-ва. Т. I / Ред. Рожнов С.В. М.: ПИН РАН, 2018. С. 90–107.
Плотицын А.Н., Соболев Д.Б., Груздев Д.А. Новые данные о пограничных отложениях девона и карбона на Приполярном Урале (р. Сывъю) // Вестн. Ин-та геол. Коми НЦ УрО РАН. 2014. № 6. С. 13–20.
Belka Z. Taxonomy, phylogeny and biogeography of the late Famennian conodont genus Mashkovia // J. Micropalaeontol. 1998. V. 17. P. 119–124.
Corradini C., Spaletta C., Mossoni A. et al. Conodonts across the Devonian/Carboniferous boundary: a review and implication for the redefinition of the boundary and a proposal for an updated conodont zonation // Geol. Mag. 2016. V. 154. № 4. P. 1–15.
Dzik J. The Famennian “Golden Age” of conodonts and ammonoids in the Polish part of the Variscan Sea // Palaeontol. Pol. 2006. V. 63. P. 1–360.
Kaiser S.I., Becker R.T. The required revision of the Devonian-Carboniferous boundary // J. Stratigr. 2007. V. 31. Suppl. 1. P. 1.
Kaiser S.I., Corradini C. Should the Devonian-Carboniferous boundary be redefined? // Newsl. Subcom. Devonian Stratigr. 2008. V. 23. P. 55–56.
Lane H.R. Symmetry in conodont element-pairs // J. Paleontol. 1968. V. 42. № 5. P. 1258–1263.
Sweet W.C. The Conodonta. Morphology, Taxonomy, Paleoecology and Evolutionary History of Long-Extinct Animal Phylum. Oxford: Clarendon Press, 1988. 212 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал