

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 5, стр. 99-110
Ильинский горизонт среднего миоцена Западной Камчатки, сообщества моллюсков и палеобиогеографические реконструкции
Ю. Б. Гладенков *
Геологический институт РАН
Москва, Россия
* E-mail: gladenkov@ginras.ru
Поступила в редакцию 27.11.2021
После доработки 23.01.2022
Принята к публикации 02.02.2022
- EDN: TMRMQP
- DOI: 10.31857/S0869592X22040032
Аннотация
Приводятся результаты изучения ильинского горизонта среднего миоцена Западной Камчатки и характерных для него моллюсков. Установлены его возрастные аналоги в Японии, на Сахалине и в Корякском нагорье. Выявлены особенности распространения сообществ моллюсков начала среднего миоцена на разных широтах шельфовых зон Северо-Западной Пацифики. Высказано предположение, что в начале среднего миоцена комплексы тепловодных (субтропических и низкобореальных) моллюсков мигрировали от широт Японии до Северной Камчатки. Это событие было связано, видимо, с “глобальным потеплением миоцена”, приведшим к определенным изменениям гидрорежима в океане, в частности к перемещению относительно теплых водных масс в северном направлении. В районах обрамления Северной Пацифики это событие последовало за тектоническими перестройками, которые повлияли на развитие морской трансгрессии, отразившейся в формировании осадочных формаций многих районов региона.
ВВЕДЕНИЕ
В послевоенные годы геологи СССР приступили к планомерному изучению широко развитых толщ кайнозоя Дальнего Востока, в частности Камчатско-Сахалинской области. Хотя эти образования начали исследовать еще в 1930-е годы, и уже тогда появились первые сведения о составе найденных в них моллюсках (Б.Ф. Дьяков, И.Б. Плешаков, И.П. Хоменко, В.С. Слодкевич, Л.В. Криштофович и др.), только в 1950-х годах здесь стали разрабатывать реальные стратиграфические схемы палеогена и неогена. В частности, в пределах Западной Камчатки были описаны опорные разрезы морских толщ, в которых были найдены последовательно сменяющиеся комплексы моллюсков, служившие тогда основой для корреляций свит.
В 1959 г. в г. Оха Северного Сахалина состоялось Первое межведомственное стратиграфическое совещание по Дальнему Востоку, на котором были впервые приняты региональные стратиграфические схемы палеогена и неогена с выделением в них свит и горизонтов. Огромную роль в их обосновании сыграли, в частности, специалисты по моллюскам Л.В. Криштофович, А.П. Ильина и другие. Хотя создание этих схем явилось важным шагом в изучении кайнозоя Дальнего Востока, многие вопросы стратиграфии оставались еще нерешенными. И поэтому стратиграфические схемы нуждались в уточнении как возраста стратонов, так и сопоставления свит разных районов. Широко развернутые геолого-съемочные и тематические работы в этот период позволили внести в схемы много нового. Результаты стратиграфических исследований были освещены в крупной сводке по геологии Камчатки середины 1960-х годов (Геология…, 1964).
В 1974 г. в г. Петропавловск-Камчатский состоялось Второе межведомственное стратиграфическое совещание по схемам кайнозоя Камчатки и Сахалина. К этому времени ряд стратиграфических проблем был освещен более полно, чем ранее, так как появились новые материалы не только по ископаемым моллюскам (например, Криштофович, 1964; Гладенков Ю., 1972), но и по фораминиферам, диатомеям, листовой флоре и пр. Кроме того, возникла возможность ознакомиться со многими материалами японских и американских специалистов – наших коллег по изучению кайнозоя Тихоокеанской области, что позволило внести много коррективов в корреляционные построения.
В 1970–1980-х гг. изучение кайнозойских разрезов региона, в том числе ископаемых комплексов, продвинулось далеко вперед. В печати появился ряд сводок по стратиграфии и палеонтологии кайнозоя разных регионов в виде атласов и монографий. В них, в частности, были приведены описания значительного числа видов ископаемой фауны и флоры (Жидкова и др., 1974; Серова, 1978; Атлас…, 1984; Гладенков Ю., Синельникова, 1990). В стратиграфических схемах кайнозоя 1994 г. (Решения…, 1998) эти данные были использованы для существенной коррекции и пополнения видовых списков комплексов, в частности моллюсков, характеризующих региоярусы (горизонты) Камчатки и Сахалина.
В пределах Западной Камчатки в палеоценовых горизонтах к началу XX века было определено несколько десятков видов моллюсков, что составило большой блок палеонтологического материала (геткилнинский горизонт – 78 видов, камчикский горизонт – 86 видов, ткаправаямский горизонт – 68 видов). Одновременно в верхнеэоценовых горизонтах было выявлено около 80 видов (аманинско-гакхинский горизонт). Если обратиться к миоцену, то здесь число определенных видов моллюсков в отдельных горизонтах тоже относительно высокое (кулувенский горизонт нижнего миоцена – 71 вид, ильинский горизонт низов среднего миоцена – 159, отдельные слои какертского горизонта – 117 и этолонского горизонта верхов среднего миоцена – до 135 видов). Естественно, что при изучении палеокомплексов горизонтов прежде всего обращалось внимание на особенности их видового состава, необходимые для определения их возрастного положения и понимания их фациальной приуроченности. Несмотря на определенную условность количественных характеристик палеокомплексов, они вместе с данными по видовому составу являются полезными как для их сравнения, так и палеобиогеографических реконструкций.
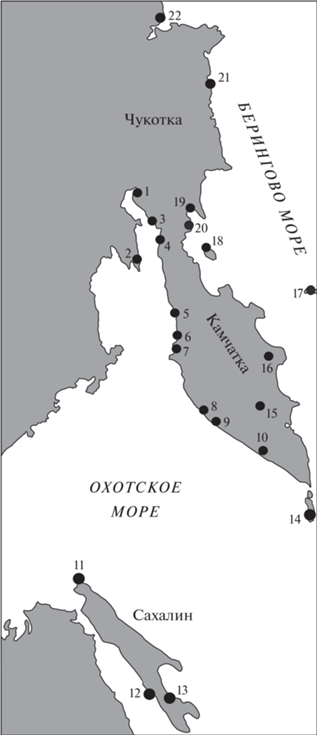
Уже давно внимание палеонтологов привлекал тот факт, что ильинский горизонт среднего миоцена по сравнению с другими горизонтами охарактеризован самым значительным числом видов – около 160. Надо иметь в виду, что это число отражает видовой состав комплекса не какого-либо отдельного разреза Западной Камчатки, а сумму видов, встреченных в нескольких ее разрезах. С севера на юг эти разрезы составляют три группы объектов, прослеживающихся вдоль восточного побережья Охотского моря на расстоянии до 1000 км (на рис. 1 отмечены номера разрезов, данные по которым используются далее). Первая из них расположена на севере Западно-Камчатской структурно-формационной зоны, на побережье Пенжинской губы (№ 1 – мыс Елистратова, мыс. Астрономический, № 2 – п-ов Тайгонос, № 3 – мыс Угольный – р. Рекинники, № 4 – мыс Большой – устье р. Пустая). Вторая группа разрезов приурочена к средней части той же зоны, к Тигильскому району (№ 5 – Точилинский опорный разрез между устьями р. Этолона и р. Тигиль, № 6 – бухта Майначская, № 7 – бухта Квачина). Третья, самая южная, группа протягивается от мыса Утхолокского практически до юга Камчатки (№ 8 – р. Хейсли, № 9 – р. Кавран, № 10 – Соболевская зона, р. Голыгина и пр.). Автор в течение ряда лет принимал участие в изучении разрезов и моллюсков ильинского горизонта из этих объектов (прежде всего второй и третьей группы), но особый вклад в эти исследования внесла В.Н. Синельникова (1985), с которой автор сотрудничал многие годы. В 1990 г. нами была выпущена книга, специально посвященная описанию моллюсков миоцена Западной Камчатки (Гладенков Ю., Синельникова, 1990). В это описание вошли сведения, в частности, и об ильинском комплексе. Однако некоторые геологические заключения по этому материалу сделаны не были, что заставляет сейчас вернуться к нему, тем более что в последние годы были изучены дополнительные палеонтологические данные по среднемиоценовым свитам (в частности, по разрезам Майначской и Квачинской бухт Западной Камчатки, а также других объектов Северо-Восточной Камчатки (Корфский район и Ильпинский п-ов) и Южного Сахалина (Холмский район (р. Арканзас–пос. Новоселово) и Макаровский район). Кроме того, для сравнения были привлечены материалы по среднему миоцену Корякского нагорья, а также Японии.
Рис. 1.
Расположение изученных разрезов нижней части среднего миоцена в Камчатско-Сахалинской области. 1 – мыс Астрономический, 2 – п-ов Тайгонос, 3 – мыс Угольный, 4 – мыс Большой, 5 – р. Тигиль (Точилинский опорный разрез), 6 – бухта Майнач, 7 – бухта Квачина, 8 – р. Хейсли, 9 – р. Кавран, 10 – р. Голыгина, 11 – п-ов Шмидта, 12 – Холмский р-он, 13 – Макаровский р-он, 14 – Курильские о-ва, 15 – Валагинский хребет, 16 – Кроноцкий п-ов, 17 – о. Беринга, 18 – о. Карагинский, 19 – Ильпинский п-ов, 20 – Корфский р-он, 21 – Хатырский р-он, 22 – Анадырский р-он.
ОСОБЕННОСТИ СТРОЕНИЯ СРЕДНЕМИОЦЕНОВОГО ИЛЬИНСКОГО ГОРИЗОНТА ЗАПАДНОЙ КАМЧАТКИ И СОСТАВА КОМПЛЕКСОВ МОЛЛЮСКОВ
Ниже дается характеристика ильинского горизонта среднего миоцена, который сформировался в шельфовой зоне Западно-Камчатского региона в период проявления ряда важных геологических событий (фаза тектогенеза конца раннего миоцена, последовавшая за ней масштабная морская трансгрессия и субглобальное потепление), что не могло не отразиться на составе и структуре свойственных горизонту ценозов моллюсков. Ильинский комплекс в целом относится к тепловодному типу, так как он содержит много паратропических родов и видов. Среди кайнозойских подразделений Западной Камчатки ильинский горизонт занимает маркирующую позицию нижнего члена кавранской серии (средний–верхний миоцен), которая сменяет в разрезе тигильскую и воямпольскую серии (палеоцен–нижний миоцен) и перекрывается с несогласием энемтенской свитой плиоцена.
Стратотипом ильинского горизонта является одноименная свита, выделенная в Точилинском разрезе Тигильского района Западной Камчатки (его название дано в 1959 г. на Охинском совещании вместо “горизонта базальных конгломератов” В.Ф. Дьякова). В этом разрезе горизонт залегает на кулувенской свите нижнего миоцена с размывом, но без видимого несогласия. В кулувенской свите (песчаники, мощность 270 м) отмечены характерные Periploma sakhalinensis (Slod.), Mytilus tihanovichi Mak., Thyasira disjuncta Gabb, Spisula equilateralis Kanno, Macoma simizuensis L. Krisht., Portlandia orientalis Glad. (в комплексе 38 видов).
Стратиграфически снизу вверх ильинскую свиту слагают (Атлас…, 1984) (рис. 2):
Рис. 2.
Стратотипический разрез ильинской свиты Западной Камчатки. 1 – конгломераты, гравелиты, 2 – песчаники, в том числе косослоистые, 3 – алевролиты, 4 – туфы, 5 – диатомиты, 6 – карбонатные конкреции, 7 – палеонтологические остатки, 8 – несогласное залегание, размывы.
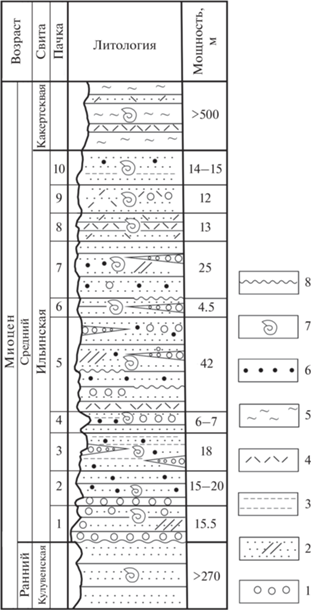
1. В основании (1 м) конгломераты среднегалечные, обычно с хорошо окатанной галькой разных пород (базальты, андезиты, песчаники, аргиллиты, опоки и др.), крепкие, местами переходящие в гравелиты. Они сменяются песчаниками с галькой. Породы грязно-серые, мусорные, переполненные раковинами моллюсков Mytilus ochotensis Slod., Modiolus wajampolkensis Slod., Panopea elongata Kanno, Pododesmus macroshisma (Deshayes) и др. Выше залегают гравелиты и песчаники голубовато-серые, местами косослоистые, грубозернистые, с кусочками древесины, с небольшими карбонатными конкрециями шаровой формы (до 5 см в диаметре), с остатками морских ежей и раковинами моллюсков Yoldia nabiliana Sim., Macoma sejugata (Yok.), Dosinia sp. и др. Мощность 15.5 м.
2. Песчаники грубозернистые, сизовато-серые, выбеленные, неяснослоистые, с тонкими прослоями алевролитов, в основании (1 м) скопление гальки. Вмещают карбонатные конкреции (до 10 см) с раковинами Periploma sachalinensis Slod., Polinices remonensis (Clark), Yoldia sp., Macoma sp. Мощность 15–20 м.
3. Песчаники и алевролиты темно-серые, массивные, крепкие, с линзами гравелитов и многочисленными мелкими шаровидными карбонатными конкрециями. Много раковин Thracia kavranensis Ilyina, Mya grewingki Mak., Periploma sakhalinensis Slod. и др. Мощность 18 м.
4. Те же породы с галечными прослоями, с остатками Mya elegans Eichwald, Turritella kadonasawaensis (Otuka). Мощность 6–7 м.
5. Песчаники темно-серые, среднезернистые, массивные, местами косослоистые. Линзы гравелитов и конгломератов. Небольшие размывы по слоистости. Внизу прослой туфа (0.5–0.6 м). Много рассеянных карбонатных конкреций – шаровых и караваеобразных. Среди остатков моллюсков Criptomya californica (Conr.), Macoma optiva (Yok.), Mya elegans. Мощность 42 м.
6. Песчаники светло-серые, мелкозернистые, плотные, с прослоями конгломератов, с остатками Mya cuneiformis Bohm, Glycymeris cf. gabbi Dall, Dosinia sp. Мощность 4 м.
7. Песчаники темно-серые, в основании с небольшим размывом, косослоистые, вверху с линзами гравелитов, с большим количеством караваеобразных конкреций (до 0.2–0.3 м). В них много раковин Crassatella vasta Devjatilova, Mytilus tihanovichii Mak., Mya elegans, Buccinum uvutschense Ilyina, Turritella kadonosawaensis, Velutina sp. Мощность 25 м.
8. Туфопесчаники сизо-серые, выбеленные, мелкозернистые, с прослоями (0.2 м) туфов андезито-дацитового состава. В верхней части мелкая галька и небольшие карбонатные конкреции неправильной формы. Много раковин моллюсков Spisula onnechiuria (Otuka), Chlamys kaneharai (Yok.), Mya cuneiformis, Macoma sejugata. Мощность 13 м.
9. Туфопесчаники голубовато-серые, мелкозернистые, кремнисто-глинистые, плотные, с раковинами Macoma cf. optiva, Mya sp. Мощность 12 м.
10. Туфопесчаники сизо-серые, алевритистые, разнозернистые, вверху шаровидные и караваеобразные конкреции, которые содержат раковины моллюсков Modiolus wajampolkensis Slod., Cyclocardia tokunagai (Yok.), Volutomitra sp. Мощность 14–15 м.
Общая мощность свиты около 150 м.
Выше согласно залегает какертская свита (до 500 м), которая включает в основном желтовато- и голубовато-серые выбеленные туфодиатомиты с прослоями кристалло-витрокластических андезито-дацитовых и дацитовых туфов. В самых нижних слоях свиты среди других обычно бореальных форм определены представители относительно тепловодных Arca, Anadara, Turritella и прочих. Но типичными формами для какертской свиты являются Mytilus cheisleveemensis Slod., M. kakertensis Sin., Yoldia thraciaeformis (Stor.), Macoma optiva, Chlamys cosibensis cosibensis (Yok.), Glycymeris iidensis Kanno, Buccinum sakhalinensis Dall и др.
Как отмечалось, ильинский горизонт трассируется практически вдоль всей Западной Камчатки с севера на юг: от северного побережья Пенжинской губы и далее на юг (до р. Кавран и Соболевской зоны), т.е. на несколько сотен километров. Залегание этого горизонта на подстилающих породах в одних случаях согласное, но с размывом, а в других случаях трансгрессивное и несогласное. Так, в северных разрезах Западной Камчатки ильинская свита залегает с размывом и резким угловым несогласием на мезозойских и палеогеновых образованиях. В основании обычно отмечаются брекчии и крупногалечные конгломераты (до 10 м) с галькой подстилающих пород – черных аргиллитов, песчаников и базальтов. Подобный же нижний контакт свиты наблюдается в районе бухты Квачина (средняя часть Охотоморского побережья Камчатки), где ильинская свита перекрывает меловые, палеогеновые и нижнемиоценовые толщи (Гладенков Ю. и др., 2018). Литологический состав горизонта в более южных разрезах повсеместно остается близким к вышеописанному: его слагают конгломераты, гравелиты и песчаники (часто косослоистые), то есть относительно мелководные образования. В базальной части горизонта валуны и галька представлены андезитами, порфиритами, песчаниками, кремнистыми сланцами, алевролитами и опоками мезозоя и палеогена. Песчаники обычно серые, зеленовато-серые, полимиктовые, разнозернистые, в верхней части туфогенные, часто плохо сортированные, косослоистые, при выветривании рыхлые, местами с карбонатными стяжениями. В них часто залегают линзы и пласты ракушняков. Мощность горизонта практически повсеместно составляет 140–150 м, и лишь в районе р. Кавран она оценивается в 170 м.
Отдельно следует сказать о комплексе моллюсков, типичных для этого горизонта. Как уже указывалось, он достаточно представительный (до 160 форм). Характерными для комплекса являются относительно мелководные (в основном верхне-сублиторальные) виды – Pododesmus macroshisma Deshayes, Modiolus wajampolkensis, Panomya elongata Kanno, Periploma sakhalinensis, Crassatella vasta, Arca watanabei Kanno, Dosinia tugaruana Nomura, Chlamys kaneharai, Macoma optiva, Anadara sp., Polinices ramonensis Clark, Turritella tokunagai Yok. и другие. Особенность этого биосообщества – присутствие представителей родов, характерных для относительно тепловодных бассейнов, что, вместе со значительным общим числом его видов, отличает данный комплекс от сообществ подстилающих и перекрывающих толщ.
Перечисленные формы в большом количестве, часто в скоплениях, встречаются во всех изученных разрезах, хотя в разных разрезах число обнаруженных видов варьирует. Так, в северных разрезах их количество достигает 130, в стратотипическом разрезе неогена Тигильского района их отмечается около 150, в южном (кавранском) разрезе – порядка 80. В последнем разрезе уменьшается, в частности, таксономическое разнообразие гастропод (отсутствуют Volutidae, мало Buссinidae). Это связано, видимо, с некоторым изменением фаций. В целом при появлении в горизонте, помимо конгломератов и гравелитов, песчано-глинистых пород количество видов обычно несколько увеличивается. Подобные изменения отмечены и в биоценозах верхней сублиторали современных морей. Из сравнения ильинского сообщества с современными комплексами (Кузнецов, 1963) следует, что его основу составляют моллюски неподвижных, малоподвижных и подвижных трофических групп.
Отметим, что стратиграфический возраст ильинского горизонта принят как нижняя часть среднего миоцена (Серова, 1978; Атлас…, 1984; Решения…, 1998; Гладенков А., 2007 и др.). Для этого использованы, в частности, данные по фораминиферам и диатомовым. Стратиграфическая позиция горизонта определена на уровне северотихоокеанских диатомовых зон Denticulopsis praelauta–Denticulopsis lauta, что соответствует интервалу 16.3–14.9 млн лет, то есть практически объему лангийского яруса низов среднего миоцена (Koizumi, 1977; Akiba, 1986). Эти “опорные” диатомовые зоны выявлены во многих упомянутых ниже разрезах Камчатки, Сахалина и Японии, как и в других районах Северной Пацифики. В Японии на о. Хонсю возрастные эквиваленты горизонта охарактеризованы также богатыми комплексами планктонных фораминиферовых зон № 8–9 шкалы У. Блоу того же возраста (Tsuchi, Ibaraki, 1981).
Ниже будут рассмотрены некоторые палеобиогеографические проблемы, связанные с распространением моллюсков в начале среднего миоцена в пределах шельфовой зоны Северо-Западной Пацифики. Для этого будут использованы литературные данные и материалы автора, который в прошлые годы смог изучить многие неогеновые разрезы как Камчатки, Корякского нагорья, Сахалина, так и Японии – Хоккайдо и Северного Хонсю.
КОРРЕЛЯЦИЯ ИЛЬИНСКОГО ГОРИЗОНТА С ВОЗРАСТНЫМИ АНАЛОГАМИ СОСЕДНИХ РЕГИОНОВ И РАСПРОСТРАНЕНИЕ СРЕДНЕМИОЦЕНОВЫХ КОМПЛЕКСОВ МОЛЛЮСКОВ В ШЕЛЬФОВЫХ ЗОНАХ СЕВЕРО-ЗАПАДНОЙ ПАЦИФИКИ
Первые попытки корреляции ильинского горизонта относятся еще к 1980-м годам прошлого века (Гладенков Ю., 1988). Сравнение ильинского горизонта по литологическому составу и палеонтологической характеристике с одновозрастными образованиями соседних районов уже тогда позволило наметить варианты их корреляции. Суммирование же последних данных приводит к новым и более обоснованным построениям. Прежде всего, было выявлено, что отложения низов среднего миоцена достаточно широко распространены в Охотоморском и Япономорском районах – на Сахалине и в Северной Японии, т.е. южнее Западной Камчатки. Комплексы моллюсков здесь обнаруживают как определенное сходство с рассмотренными ранее, так и отличия от них.
На Южном Сахалине (рис. 1; разрезы № 12, 13 – Холмский (с Александровским) и Макаровский районы, № 14 – Курильские о-ва) эти образования представлены аусинской свитой, которая залегает с размывом на подстилающих толщах (Жидкова и др., 1974). В ее песчанико-конгломератовых породах мощностью 50–150 м среди бореальных форм отмечены, в частности, тепловодные виды Dosinia tugaruana Nomura, Arca watanabei Kanno, A. amicula elongata Noda, Оstrea sp. и др. На Северном Сахалине (разрез № 11 – п-ов Шмидта) в пильской свите определены остатки Pitar, Оstrea (Гладенков Ю. и др., 2002).
Автору в свое время пришлось изучить ряд разрезов кайнозоя Японии, что позволило получить сравнительные данные, в частности, по среднему миоцену этого региона (Гладенков Ю., 1988). В кратком виде эти данные сводятся к следующему. На о. Хоккайдо, в районе Юбари находится формация Такиное низов среднего миоцена (конгломераты и грубозернистые выбеленные песчаники и алевролиты мощностью до 80 м), которая залегает на подстилающих толщах несогласно и с размывом в основании. В нижней части формации собраны раковины тепловодных родов моллюсков Dosinia, Anadara, Pitar, Crepidula, Turritella и проч. Обращает на себя внимание присутствие в комплексе редких типично тропических форм Vicariella, которые в более северных разрезах не встречены (Kanno, Ogawa, 1964; Ogasawara et al., 1982).
Южнее о. Хоккайдо, в северной части о. Хонсю (горы Китаками), отмечена формация Кадонозава с реперным комплексом моллюсков (с “фауной Кадонозава”). Она залегает с размывом и сложена в основном песчаниками с галькой и алевролитами мощностью около 100 м. Характерными видами для нее являются многочисленные относительно тепловодные Vicaria, Anadara, Ostrea и другие формы (Chinzei, 1978). В других районах Северного Хонсю (Сендай, Дзебан, Какегава, п‑ов Ога) в одновозрастных толщах (формации Тсунаки, Шиобара, Нисикуросава и другие), залегающих обычно с размывом, найдены сообщества конхилиофауны тропического и южнобореального типа с Vicaria, Vicariella, Dosinia, Anadara, Pitar, Turritella (Tsuchi, Ibaraki, 1981). Надо отметить, что в Северной Японии в ряде формаций отмечается теплолюбивая флора Дайдзима, которая маркирует “миоценовый климатический оптимум” (Tanai, 1967).
В южной части Японии в среднемиоценовых разрезах выявлены комплексы моллюсков типично тропического типа, которые южнее прослеживаются в шельфовых зонах Индонезии, Филиппин, Бирмы и Индии (Shuto, 1975). Корреляции неогеновых горизонтов Северной и Южной Японии посвящены уже многие работы (Ikebe, Tsuchi, 1984).
Вместе с тем возрастные аналоги ильинского горизонта встречаются и в других районах – на Восточной Камчатке и Корякском нагорье, которые омываются водами Тихого океана, с включением Берингова моря. В пределах восточной части Камчатки, на южном фланге Валагинского хребта (рис. 1, разрез № 15, р. Корниловская), выделяется, в частности, корниловский горизонт нижнего–среднего миоцена (Гладенков Ю., 2016). В его средней толще, сложенной конгломератами и песчаниками с галькой (100 м), отмечена группа тепловодных моллюсков, которые сходны с комплексом ильинской свиты: Dosinia sp., Modiolus wajampolkensis, Macoma optiva, Spisula onnechiuria и другие. В верхах толщи встречаются остатки теплолюбивых растений Fagus. В пределах Кроноцкого п-ова (разрез № 16, р. Ракитинская, р. Тюшевка и их притоки) выделяются несогласно и с размывом залегающие ракитинская свита и свита Горячих ключей (конгломераты, песчаники, ракушняки, спонголиты, мощностью до 50 м и более), которые вмещают моллюски Glycymeris, Crassatella, Securella, Pitar, Taras, Crenella и пр. Сходные остатки отмечены и в средней части горбушевской толщи того же района. Одновременно на Командорских о-вах (разрез № 17) в среднемиоценовых слоях толщи Северо-Западного мыса отмечены остатки тепловодного рода Pitar (Гладенков Ю., 1984).
На о. Карагинский Восточной Камчатки (разрез № 18) низы среднего миоцена (пестроцветная свита, нижняя часть) сложены гравелитами и песчаниками (100 м). Здесь тоже найдены остатки раковин тепловодных видов Securella chehalisensis Weaver, Neneridae, Columbaridae (Гладенков Ю. и др., 1992).
В целом на Восточной Камчатке подобные комплексы встречаются реже, чем на Западной Камчатке, в связи с тем, что верхнесублиторальные отложения в силу разных причин (в том числе тектонических и палеогеографических) представлены здесь не широко, а преобладающими являются флишоидные и относительно глубоководные фации, которые часто сопровождаются вулканическими образованиями. Тем не менее и здесь, как уже отмечено, встречены мелководные толщи со сравнительно тепловодными палеосообществами.
Помимо этого, достаточно представительные тепловодные комплексы среднего миоцена были найдены нами в пограничной зоне Северо-Восточной Камчатки и южного фланга Корякского нагорья, в районе пос. Корф – Ильпинского п-ова (Гладенков Ю. и др., 1987; Волобуева и др., 1994). Несколько севернее пос. Корф (разрез № 19) была выделена успенская свита, сложенная в основном песчаниками. В ее верхней части развиты толщи (75–100 м) гравелитов, алевролитов, песчаников с прослоями туфов и лигнитов. Они охарактеризованы сравнительно богатым сообществом тепловодных моллюсков (около 40 видов): Dosinia rumoensis Amano, D. tugaruana, D. kaneharai Yok., Yoldia nabiliana, Modiolus wajampolkensis., Thracia kavranensis Ilyina, Mya grewingki Fujie, Periploma cf. sakhalinensis., Anadara korfiensis Sin., Securella chehalisensis, Pitar gretschischkini, Macoma optiva, Ostrea makarovi L. Krisht., Mytilus ochotensis, Polinices ramonensis, Crepidula ungana Dall, Neverita jamesae Moore, Cancellaria oregonensis Conrad и другие. Вместе с тем севернее и западнее пос. Корф выделен “ежовый горизонт” (песчаники мощностью 100 м) с близкими сообществами моллюсков, а в северной части п-ова Ильпинский (№ 20) в песчано-конгломератовых породах также был отмечен сходный комплекс. Сообщество из района бухты Корфа отличается, во-первых, присутствием в нем значительного числа представителей родов, распространенных сейчас в субтропических и южнобореальных морях, а во-вторых, большим сходством данной ассоциации моллюсков с таковой ильинского горизонта Западной Камчатки. Последнее может свидетельствовать в пользу высказанного в прошлые годы предположения о прямой связи морских бассейнов среднего миоцена Западной Камчатки и северо-восточной части Камчатки.
Среднемиоценовые толщи отмечаются также северо-восточнее Камчатки – в пределах беринговоморских районов Корякского нагорья (Чукотка). Они описаны в Хатырском прогибе и Анадырской впадине. В Хатырском прогибе (рис. 1, разрез № 21), расположенном на западной окраине Берингова моря, эти отложения слагают ундал-уменскую свиту (конгломераты и песчаники, мощность несколько сотен метров). Она залегает с несогласием и охарактеризована бореальным комплексом моллюсков, но среди них не отмечено ни одного представителя субтропического-южнобореального типа (таких как Dosinia, Arca, Anadara и других родов, встреченных южнее, на Камчатке и Сахалине). Хотя здесь и отмечены редкие остатки фагусовых, появление которых на этих широтах было связано, видимо, с общим относительным потеплением (Гладенков Ю. и др., 1985). Что касается расположенной севернее Анадырской впадины (рис. 1, № 22), то в ней к среднему миоцену относится автаткульский горизонт (песчаники мощностью до 200–690 м). В нем тоже отсутствуют тепловодные виды моллюсков этого возраста (Агапитов, 1991).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Для более корректного проведения палеобиогеографических реконструкций представляется целесообразным кратко напомнить о некоторых особенностях распространения биотических сообществ, в том числе моллюсков, на современных шельфах переходной зоны Камчатско-Сахалино-Японского региона. Они подробно изложены в ряде публикаций, например в сводных работах А.И. Кузнецова (1963), О.А. Скарлато (1981), А.И. Кафанова и В.А. Кудряшова (2000) и других.
В конце прошлого века О.А. Скарлато (1981) на основании синтеза данных по двустворчатым моллюскам разных районов северо-западной части Тихого океана предложил оригинальную схему биогеографического районирования шельфа, относящегося к Тихоокеанской бореальной биогеографической области. Эту схему можно использовать для сравнения (рис. 3). Северная граница области практически совпадает с Беринговым проливом, южная – проведена в средней части о. Хонсю. В этой области были выделены две биогеографические подобласти – Берингийская высокобореальная и Северояпонская низкобореальная с границей на широте средней части Сахалина. В первой подобласти были намечены несколько провинций: Северокурильская, Охотоморская, Командорская (или округ), Восточночукотская, а также Анадырский округ. Во второй, Северояпонской, подобласти были выделены Северояпонская и Южносахалинская провинции. Между подобластями и провинциями существуют переходные зоны. Выделение этих биогеографических подразделений основано на анализе ареалов видов моллюсков. При этом учитываются три фоновых фактора, влияющих на ареалы, – широтная (климатическая) и вертикальная зональности моря, а также географическая провинциальность. Но в принципе температура является первичным фактором биогеографической дифференциации в море. В связи с этим обратим внимание на определение термопатии видов – способности организмов к обитанию в диапазоне температур, соответствующих экологическим требованиям вида. Термопатию наиболее четко определяют температуры, при которых происходит размножение видов. Эти “температуры размножения” коррелируются с зональными типами видовых ареалов. По О.А. Скарлато, согласно эмпирическим наблюдениям, среди двустворчатых моллюсков такие температуры составляют в среднем: у бореально-арктических видов 0–10°С, у широко распространенных бореальных 2–8°С, у низкобореальных 6–14°С, у субтропических и субтропическо-низкобореальных 14–20°С. При этом выявлено, что в субтропических и тропических широтах влияние температуры на распространение гидробионтов сравнительно ослаблено.
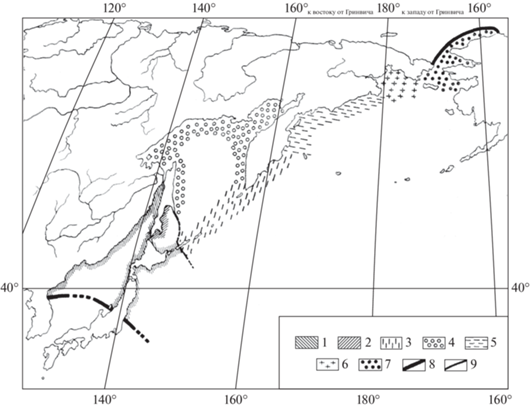
Рис. 3.
Биогеографическое районирование шельфа северо-западной части Тихого океана (Тихоокеанская бореальная биогеографическая область) по О.А. Скарлато (1981). 1, 2 – Северояпонская низкобореальная подобласть: 1 – Северояпонская провинция, 2 – Южносахалинская провинция; 3–7 – Берингийская высокобореальная подобласть: 3 – Северокурильская провинция, 4 – Охотоморская провинция, 5 – Командорская провинция (округ), 6 – Анадырский округ, 7 – Восточночукотская провинция; 8 – южная и северная границы Тихоокеанской бореальной биогеографической области, 9 – граница между Северояпонской низкобореальной и Берингийской высокобореальной подобластями.
С учетом этих данных можно попытаться расшифровать некоторые особенности палеобиогеографии шельфовой зоны Камчатско-Сахалино-Японского региона в начале среднего миоцена. Одна из таких особенностей – распространение относительно тепловодных моллюсков в дальневосточных морях прошлого, которое отличалось от современного. Принимая во внимание постоянно накапливающиеся стратиграфические и палеонтологические данные по разным временным срезам неогена, нельзя не прийти к выводу, что температурные характеристики водных масс дальневосточных морей прошлого менялись. В качестве показательного примера привлекает внимание ильинский горизонт с его фауной, позволяющий решить ряд палеобиогеографических вопросов. Поэтому не случайно О.А. Скарлато, крайне заинтересованный в расшифровке реальной биогеографии северотихоокеанского бассейна прошлого, неоднократно обсуждал эту проблему с палеонтологами и делился своими соображениями на этот счет.
Из анализа вышеприведенных сравнительных данных по разрезам осадочных толщ низов среднего миоцена следует, что относительно тепловодные роды моллюсков были распространены в шельфовой зоне значительной территории – от Японии до Северной Камчатки, в то время как в настоящее время они распространены в бассейнах тропических широт и частично низкобореальной подобласти, которая охватывает район от Средней Японии до Среднего Сахалина. И если в качестве “климатических реперов” использовать роды Dosinia, Arca, Anadara, Papyridea, Pitar и др., которые обычно составляют своеобразные “дозиниевые комплексы”, отмеченные в миоцене Камчатки, то не трудно видеть, что сейчас они встречаются в Северояпонской низкобореальной подобласти (по О.А. Скарлато), т.е. на широтах около 35°–40° с.ш. и южнее (в Индо-Вестпацифической тропической области) (рис. 4, 5).
Рис. 4.
Схема положения южной и северной границ современной (Q) и среднемиоценовой (N) низкобореальной биогеографической подобласти. 1 – границы (южная и северная) современной Северояпонской низкобореальной подобласти, 2 – предполагаемые границы (южная и северная) Пра-Берингийской высокобореальной подобласти начала среднего миоцена.
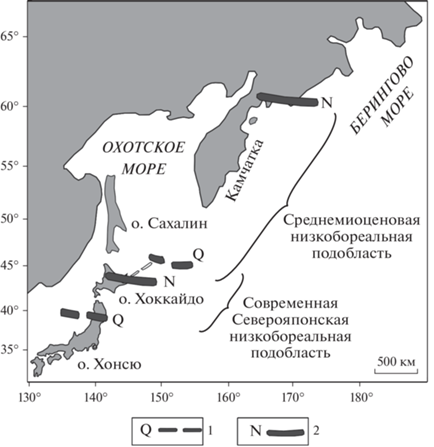
Рис. 5.
Схема миграции тепловодных комплексов моллюсков в бореальные широты шельфовой зоны Тихоокеанской бореальной биогеографической области в период “климатического оптимума” среднего миоцена и пространственный масштаб предполагаемых биотических миграций. 1–3 – типы комплексов: 1 – тропический с Vicaria-Vicariella, 2 – низкобореальный c Dosinia-Arca, 3 – высокобореальный; Q – современные, N – среднемиоценовые; ИВ – Индо-Вестпацифическая область.
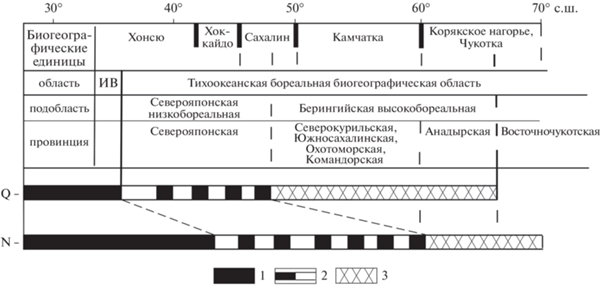
Естественно предположить, что в начале среднего миоцена произошли миграции тепловодных биокомплексов в дальневосточных морях к северу, что было связано, видимо, с глобальным потеплением (Haq et al., 1987; Zachos et al., 2001), приведшим к расширению тропической зоны в океане и смещению ее северной границы на север. По мнению японских специалистов (Ikebe, Tsuchi, 1984), это потепление соответствовало отрезку времени около 2 млн лет. В связи с этим встает ряд вопросов. Например, проявлялись ли в это время более мелкие климатические флуктуации (мы пока их не фиксируем на нашем палеонтологическом материале, хотя они отмечаются в четвертичном периоде)? Почему северная граница низкобореальной подобласти в начале среднего миоцена выглядит относительно резкой (где-то на широте Северной Камчатки, севернее которой – в Хатырском прогибе – тепловодная фауна, типа “комплекса с Dosinia”, отсутствует)? Связано ли видовое богатство сообществ Западной Камчатки с наличием значительного числа эндемичных форм, которые могли появиться в условиях полузамкнутого окраинного Охотоморского бассейна? Может быть, мы пока не умеем учитывать специфику развития биоты в условиях таких бассейнов, как и гидрологические особенности их водных масс (например, не можем оценить роль морских течений в расселении моллюсковых комплексов). К сожалению, очень трудно на ископаемом материале выявить с достоверностью и положение древних синператов (сгущения ареалов видов с локальными максимумами видового богатства), которые сейчас используются при проведении естественных границ фаунистических выделов. Возможно, эти и другие вопросы будут решены при осуществлении более детальных геологических, в частности палеонтологических, исследований. Но важно, что уже сейчас мы можем поставить вопросы, которые предстоит решать в будущем. В настоящее время при изучении шельфовых областей прошлого получено много новых данных о кайнозойских толщах в пределах обрамления современных морских бассейнов и их шельфовых зон, чему способствует развернувшееся бурение. С другой стороны, появилось много сейсмостратиграфических материалов (прежде всего по шельфу Сахалина и Японии), которые позволяют более объективно и более детально, чем раньше, выстраивать региональные и межрегиональные корреляции неогеновых горизонтов.
ВЫВОДЫ
Синтез изложенных материалов позволяет сделать определенные выводы в отношении как стратиграфических корреляций морского среднего миоцена Дальнего Востока, так и некоторых расшифровок геологических событий и палеогеографических обстановок в переходной зоне Северо-Тихоокеанской области. Среди них можно отметить следующие:
1. В Западно-Камчатской структурно-фациальной зоне в качестве одного из реперных подразделений неогена выделяется ильинский горизонт начала среднего миоцена, представленный конгломерато-песчаниковыми породами с характерным комплексом тепловодных моллюсков. Он прослеживается в пределах данного региона с севера на юг почти на 1000 км, маркируя мелководную зону древнего Пра-Охотского моря. Горизонт был образован во время значительной морской трансгрессии, в связи с чем его нижние слои залегают на подстилающих толщах с размывом и часто с несогласием.
2. Ильинский горизонт охарактеризован комплексом моллюсков, отличающимся от таковых из подстилающих и перекрывающих горизонтов наличием относительно большого числа родов и видов тепловодного типа (“комплекс с Dosinia”). Скорее всего, появление этих форм связано с миграциями тепловодных (субтропических и южно-бореальных) сообществ из низких широт в высокие (от Японии до Северной Камчатки), что было обусловлено, видимо, глобальным потеплением начала среднего миоцена (“миоценовый климатический оптимум”), которое привело к определенным изменениям гидрологических обстановок в Мировом океане, в частности к расширению площади водных масс тропического и низкобореального типа.
3. Возрастные аналоги ильинского горизонта Камчатки отмечаются в более южных районах – на Сахалине и в Японии, где относящиеся к ним свиты и формации тоже содержат тепловодные (от южнобореальных до типично тропических) комплексы конхилиофауны и тоже залегают с размывами (и часто с несогласием). Это может указывать на проявление в предшествующее время тектонических и палеогеографических перестроек в значительной части переходной зоны Северо-Западной Пацифики и Азиатского материка.
4. Ильинскому горизонту соответствуют также некоторые свиты Восточной Камчатки Беринговоморской области, в которых местами тоже отмечаются тепловодные моллюски. Самым северным районом, где нижние части среднего миоцена охарактеризованы тепловодными элементами морской фауны (сообщества с Dosinia), сходными с комплексами Западной Камчатки, являются участки древнего шельфа, расположенные в приграничных районах Северо-Восточной Камчатки и юга Корякского нагорья. В более северных районах Корякского нагорья теплолюбивых моллюсков этого возраста не найдено. Этот северокамчатский рубеж может приниматься, видимо, как граничная зона между высокобореальной и низкобореальной подобластями для начала среднемиоценового времени.
5. Из представленного материала следует, что в период потепления начала среднего миоцена тропический комплекс моллюсков Японии (с Vicaria-Vicariella) переместился к северу (от Южной Японии к Северной) на 400–500 км. В то же время субтропическо-южнобореальное сообщество (с Dosinia) передвинулось севернее (от Северной Японии–Южного Сахалина до Северной Камчатки) на 1500–2000 км. Другими словами, если пользоваться схемой биогеографического районирования О.А. Скарлато (1981), в первом случае южная граница Северояпонской низкобореальной подобласти сместилась приблизительно от 37° до 43°–45° с.ш., а южная граница Берингийской высокобореальной подобласти с 44°–45° до 60° с.ш. Это может означать, что по сравнению с тропической зоной “прирост” (или увеличение площади) низкобореальной подобласти в среднемиоценовое время был более значительным (рис. 4, 5). Подобная особенность (или даже закономерность), возможно, проявлялась и в другие периоды потеплений палеогена и неогена.
6. Сказанное выше позволяет сделать одно общее методическое замечание. Выявление следов масштабных миграций древних биосообществ в ряде случаев может помочь в решении проблемы пространственного сонахождения древних биокомплексов разного палеогеографического типа, которое трактуется иногда без должного обоснования как результат сочленения (или причленения) разных экзотических террейнов. К сожалению, в некоторых работах это увлечение террейнами часто уводило (да и сейчас подчас уводит) от решений реальных проблем геологического строения некоторых дальневосточных районов.
В заключение следует заметить, что в кайнозое Северо-Тихоокеанской области миграции тепловодных биокомплексов от южных широт к северным проходили и на других временных уровнях. Это касается, прежде всего, раннего эоцена, конца среднего миоцена и начала плиоцена, о чем уже сообщалось в литературе (Жидкова и др., 1974; Гладенков Ю., 1988, 2021 и др.). Не исключено, что приведенные выше материалы будут полезны при расшифровке особенностей палеогеографических событий и в другие периоды фанерозоя.
Кайнозойские образования Камчатско-Сахалинского региона, относящегося к переходной зоне Северо-Тихоокеанской области, нуждаются сейчас в более масштабных и более детальных, чем ранее, исследованиях. Без этого получить реальные расшифровки палеобиогеографических событий весьма трудно.
Благодарности. Я хотел бы с благодарностью вспомнить коллег по изучению кайнозойских моллюсков дальневосточных бассейнов – гидробиологов О.А. Скарлато, А.Н. Голикова, А.П. Кузнецова и биостратиграфов В.И. Волобуеву, В.О. Савицкого, Л.С. Жидкову и В.Н. Синельникову, с которыми мне пришлось в прошлом обсуждать поднятые в данной заметке биогеографические проблемы. Все они были замечательными специалистами. Я также хотел бы поблагодарить С.В. Попова, С.И. Бордунова и В.С. Вишневскую за советы по улучшению текста статьи.
Источники финансирования. Работа выполнена по теме государственного задания ГИН РАН и при поддержке гранта РФФИ № 19-05-00361.
Список литературы
Агапитов Д.И. Новые данные о стратиграфии неогеновых и палеогеновых отложений Анадырской впадины // Геология и стратиграфия кайнозойских отложений Северо-Западной Пацифики. Владивосток: ДВО АН CCCH, 1991. С. 70–95.
Атлас фауны и флоры палеогеновых отложений Дальнего Востока. Точилинский опорный разрез. Отв. ред. Меннер В.В. М.: Наука, 1984. 335 с. (Труды ГИН АН СССР. Вып. 385).
Волобуева В.И., Гладенков Ю.Б., Беньямовский В.Н., Витухин Д.И., Минюк П.С., Музылев Н.Г., Олейник А.Э., Синельникова В.Н., Соколова З.Ш., Титова Л.В., Фрегатова Н.А., Ширая О.А. Опорный разрез морского палеогена севера Дальнего Востока (п-ов Ильпинский). Ч. 1. Стратиграфия. Магадан: СВКНИИ ДВО РАН, 1994. 64 с.
Геология СССР. Т. 31. Камчатская область, Курильские и Командорские острова. Ч. 1. М.: Недра, 1964. 729 с.
Гладенков А.Ю. Детальная стратиграфия и морские экосистемы позднего кайнозоя севера Тихоокеанского региона (по диатомеям). М.: ГЕОС, 2007. 296 с. (Труды ГИН РАН. Вып. 571).
Гладенков Ю.Б. Неоген Камчатки. М. Наука, 1972. 251 с. (Труды ГИН АН СССР. Вып. 214).
Гладенков Ю.Б. Комплексы моллюсков из третичных толщ Командорских островов // Докл. АН СССР. 1984. Т. 274. № 3. С. 678–680.
Гладенков Ю.Б. Стратиграфия морского неогена северной части Тихоокеанского пояса (анализ стратиграфических схем дальневосточных районов СССР, Северной Америки и Японии). М.: Наука, 1988. 212 с. (Труды ГИН АН СССР. Вып. 428).
Гладенков Ю.Б. Опорные разрезы палеогена и неогена Восточной Камчатки // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 1. С. 64–80.
Гладенков Ю.Б. Морской палеоген Корякского нагорья, Северо-Восточная Азия: стратиграфия, моллюски, корреляция и геологические события // Стратиграфия. Геол. корреляция. 2021. Т. 29. № 3. С. 72–92.
Гладенков Ю.Б., Синельникова В.Н. Моллюски и климатические оптимумы миоцена Камчатки. М.: Наука, 1990. 174 с. (Труды ГИН АН СССР. Вып. 453).
Гладенков Ю.Б., Волобуева В.И., Митрофанова Л.И. Третичные толщи Нижнехатырского прогиба (Корякское нагорье) // Изв. АН СССР. Сер. геол. 1985. № 3. С. 77–93.
Гладенков Ю.Б., Братцева Г.М., Синельникова В.Н. Морской кайнозой залива Корфа Восточной Камчатки // Очерки по геологии Северо-Западного сектора Тихоокеанского тектонического пояса. М.: Наука, 1987. С. 5–73.
Гладенков Ю.Б., Баринов К.Б., Басилян А.Э., Бордунов С.И., Братцева Г.М., Зырянов Е.В., Кураленко Н.П., Витухин Д.И., Орешкина Т.В., Ганзей С.С., Кияшко С.И., Трубихин В.М. Детальное расчленение неогена Камчатки. М.: Наука, 1992. 208 с. (Труды ГИН РАН. Вып. 478).
Гладенков Ю.Б., Баженова О.К., Гречин В.И., Маргулис Л.С., Сальников Б.А. Кайнозой Сахалина и его нефтегазоносность. М.: ГЕОС, 2002. 225 с.
Гладенков Ю.Б., Гладенков А.Ю., Бордунов С.И., Чехович В.Д., Александрова Г.Н., Водовозов В.Ю., Гептнер А.Р., Дмитриева Т.В., Покровский Б.Г., Трубихин В.М., Фрегатова Н.А. Опорный разрез кайнозоя Западно-Камчатской структурно-формационной зоны. М.: ГЕОКАРТ, ГЕОС, 2018. 202 с.
Жидкова Л.С., Мишаков Г.С., Неверова Т.И., Попова Л.А., Сальников Б.А., Сальникова Н.Б., Шереметьева Г.Н. Биофациальные особенности мезокайнозойских бассейнов Сахалина и Курильских островов. Новосибирск: Наука, 1974. 251 с.
Кафанов А.И., Кудряшов В.А. Морская биогеография. М.: Наука, 2000. 176 с.
Криштофович Л.В. Моллюски третичных отложений Сахалина. Л.: Недра, 1964. 343 с. (Труды ВНИГРИ. Вып. 232).
Кузнецов А.П. Фауна донных беспозвоночных прикамчатских вод Тихого океана и северных Курильских островов. М.: Изд-во АН СССР, 1963. 271 с.
Решения рабочих межведомственных региональных стратиграфических совещаний по палеогену и неогену восточных районов России – Камчатки, Корякского нагорья, Сахалина и Курильских островов. Объяснительная записка к стратиграфическим схемам. М.: ГЕОС, 1998. 147 с.
Серова М.Я. Стратиграфия и фораминиферы неогена Камчатки. М.: Наука, 1978. 142 с. (Труды ГИН АН СССР. Вып. 323).
Синельникова В.Н. Стратиграфия и палеонтологическая характеристика миоцена Северо-Западной Камчатки // Корреляция кайнозойских толщ Дальнего Востока. Ч. 1. М.: ГИН АН СССР, 1985. С. 56–83.
Скарлато О.А. Двустворчатые моллюски умеренных широт западной части Тихого океана. Л.: Наука, 1981. 480 с. (Определители по фауне СССР, издаваемые Зоологическим ин-том АН СССР. № 126).
Akiba F. Middle Miocene to Quaternary diatom biostratigraphy in the Nankai Trough and Japan Trench, and modified lower Miocene through Quaternary diatom zones for the middle-to-high latitudes of the North Pacific // Init. Rep. DSDP. Washington: U.S. Govt. Printing Office, 1986. V. 87. P. 393–481.
Chinzei K. Neogene molluscan faunas in the Japanese Islands: an ecological and zoogeographic synthesis // Veliger. 1978. V. 21. P. 155–170.
Haq B.U., Hardenbol J., Vail P.R. Chronology of fluctuating sea levels since the Triassic // Science. 1987. V. 235. P. 1156–1167.
Ikebe N., Tsuchi R. Synopsis of Japanese Neogene stages // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1984. V. 46. P. 1–10.
Kanno S., Ogawa H. Molluscan fauna from the Momijiyama and Takinoue Districts, Hokkaido, Japan // Sci. Rept. Tokyo Kyoiku Daigaku. Sect. C. 1964. V. 8. P. 109–294.
Koizumi I. Diatom biostratigraphy in the North Pacific region // Proc. First Int. Congress on Pacific Neogene Stratigraphy. Eds. Saito T., Ujiie H. Tokyo: Kaiyo Shuppan Co. Ltd., 1977. P. 235–251.
Ogasawara K., Kotaka T., Masuda K., Noda Y. Outlines of the Neogene molluscan faunas from Hokkaido // Problems of the Neogene Biostratigraphy of Hokkaido. Ed. Tanai T. Sapporo: Hokkaido Univ., 1982. P. 3–13.
Shuto T. Preliminary correlation of the Neogene molluscan faunas in southeast Asia // Geology and Palaeontology of Southeast Asia. V. 15. Tokyo: University of Tokyo Press, 1975. P. 289–301.
Tanai T. Tertiary floral changes of Japan // Jubil. Publ. Commem., Prof. Sasa’s 60th birthday. Sapporo: Hokkaido Univ., 1967. P. 317–334.
Tsuchi R., Ibaraki M. Stratigraphic outline of selected Neogene sequences; Kakegawa area // Neogene of Japan: Its biostratigraphy and chronology. IGCP-114 National Working Group of Japan. Ed. Tsuchi R. Shizuoka, Japan, 1981. P. 123–138.
Zachos J., Pagain M., Sloan L., Thomas E., Billups K. Trends, rhythms, and aberrations in global climate 65 Ma to present // Science. 2001. V. 292. P. 686–691.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция