

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 5, стр. 111-128
Биоиндикаторы условий формирования верхненеоплейстоценовых каргинских и сартанских отложений Колымской низменности и особенности их тафономии при многократном воздействии криогенных факторов
О. Г. Занина 1, Д. А. Лопатина 2, *
1 Институт физико-химических и биологических проблем почвоведения РАН
Московская обл., Пущино, Россия
2 Геологический институт РАН
Москва, Россия
* E-mail: dalopat@mail.ru
Поступила в редакцию 30.10.2021
После доработки 09.03.2022
Принята к публикации 23.03.2022
- EDN: IQBGSK
- DOI: 10.31857/S0869592X22050076
Аннотация
Приводятся результаты изучения отложений каргинского и сартанского возраста Колымской низменности, синхронных морским изотопным стадиям (МИС) 3 и 2, и микрофитофоссилий из них. Во время накопления каргинских отложений на рассматриваемой территории существовали разнообразные ландшафты – сообщества влажных и суховатых тундр, лиственничных редколесий и тундровых болот с включением местообитаний, занятых пионерной и степной растительностью. Отложения сартанского возраста формировались в более однообразных, холодных и сухих условиях. Результаты изучения микробиоморф свидетельствуют о преобладании в этот период на территории изучаемого региона тундровых травянистых, преимущественно злаковых ассоциаций в комплексе с наземным моховым покровом. Удовлетворительная сохранность микрофитофоссилий из этих толщ обусловлена спецификой криолитозоны с преобладанием низких температур в течение года и низкой микробиологической активностью. Присутствие спор и пыльцы с повреждениями физического типа (разрывами и трещинами) и корродированных форм фитолитов может быть связано с нахождением их в сезонно-талом слое при попеременном чередовании циклов промерзания и оттаивания.
ВВЕДЕНИЕ
Значительный объем отложений верхнего неоплейстоцена на территории Колымской низменности занимают криопедолиты – толщи алевритов, проработанные в ходе своего накопления синлитогенным почвообразованием, пронизанные мощными жилами льда и называемые ледовым комплексом или едомой (Каплина и др., 1978; Каплина, 2009; Томирдиаро, 1980). Специфика накопления отложений определила отсутствие дифференциации материала толщ и общую морфологическую монотонность (Губин, 1984). С.В. Губин (2002) показал, что криопедолиты каргинского (молотковского) возраста, соотносимые на основании полученных радиоуглеродных датировок с образованиями морской изотопной стадии (МИС) 3, и криопедолиты сартанского возраста, отвечающие МИС 2, близки по морфологическому строению – монотонной окраске, присутствию высокодисперсного растительного детрита и тонких корней трав, слоистости и криотекстурам. Анализ гранулометрического состава этих отложений также показал их сходство – абсолютное преобладание пыли с доминированием фракции 0.05–0.01 мм. Криопедолиты рассматриваемых возрастных интервалов обладают нейтральной или слабощелочной реакцией (рН 7–8.5), сходными значениями Сорг (0.8–1.8%) как по простиранию слоя, так и по его мощности, что, наряду с уникально высокими содержаниями подвижных форм фосфора (до 60 мг/100 г), указывает на значительное участие при их формировании живых организмов (Занина, 2006). Сходство перечисленных характеристик свидетельствует об определенной близости условий накопления данных толщ. Различия между ними заключаются в несколько большем общем количестве органического материала в сартанских образованиях и их повышенной грубогумусности, а также в более контрастных отличиях в содержании Сорг. между слоями этих отложений (Губин, 1994, 1998). Для каргинских отложений отмечены признаки трансформации органических остатков и гумификации, установлено присутствие оторфованного материала со значительным участием мхов, что свидетельствует о лучшей теплообеспеченности летнего периода времени их формирования и относительно высокой его влажности, чем в сартанское время (Губин, 2002). Каргинские толщи содержат ископаемые норы грызунов и сусликов (Губин и др., 2001, 2003а; Занина, 2005; Zanina et al., 2011; Лопатина, Занина, 2006), а также четыре профиля разновозрастных погребенных почв (Губин, Занина, 2013, 2014; Лопатина, Занина, 2020; Lopatina, Zanina, 2020). Нижняя погребенная почва (ПП) отнесена к раннекаргинскому педокомплексу (40 тыс. лет назад и более), а три расположенные выше погребенные почвы – к позднекаргинскому (I ПП – 37–35 тыс. лет назад, II ПП – 33–31 тыс. лет назад и III ПП около 28 тыс. лет назад). В сартанских отложениях исследуемого региона погребенные почвы не обнаружены, что, возможно, связано с усилением суровости климата и увеличения привноса минерального материала.
В верхненеоплейстоценовых отложениях Колымской низменности установлен значительный объем ископаемого растительного материала (остатки стволов, ветвей и стеблей, корней, а также листья, плоды, семена, споры и пыльца растений), определены способные к прорастанию семена высших растений, в частности смолевки (Яшина и др., 2002). Относительно быстрый переход остатков растений в погребенное состояние и последующее их пребывание в слое ежегодного летнего оттаивания при относительно низких положительных температурах, чему способствовали высокие запасы холода в вечномерзлых отложениях, обусловили хорошую сохранность исследуемого растительного материала. С.В. Губиным с соавторами (2003б) сделан вывод о том, что трансформация растительного материала существовала лишь в верхней части сезонно-талого слоя, в зоне с суточными и сезонными колебаниями температуры и, соответственно, с неоднократными фазовыми переходами воды, активным действием почвенных растворов и микробиоты. Органический материал находился в данной зоне лишь один-два месяца в году в течение первых десятков лет. Далее с увеличением глубины погребения и до перехода в вечномерзлое состояние летние температуры не превышали 0…+3°С, а это способствовало понижению активности микрофлоры и хорошей сохранности органического материала. Проводимый авторами статьи анализ органических (спор и пыльцы) и минеральных микрофоссилий (фитолитов, диатомовых водорослей, спикул губок) из едомных отложений Колымской низменности выявил среди них определенное количество поврежденных форм, что позволило выдвинуть предположение о том, что часть их разрушалась при многократных циклах промерзания-оттаивания, которые сопровождали переход осадочных отложений в многолетнемерзлые.
Целью настоящей работы является изучение с помощью методов микроскопии компонентов биогенной фракции образца – органической (пыльцы, спор, древесного и травянистого детрита) и биогенного кремнезема (фитолитов), как биоиндикаторов для выявления отличий между криопедолитами каргинского и сартанского времени на Колымской низменности; детализация реконструкций растительности и ландшафта рассматриваемых временных интервалов; анализ сохранности микрофитофоссилий; определение возможностей его использования при палеоэкологических исследованиях и выявление механизмов длительной криоконсервации растительных остатков.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
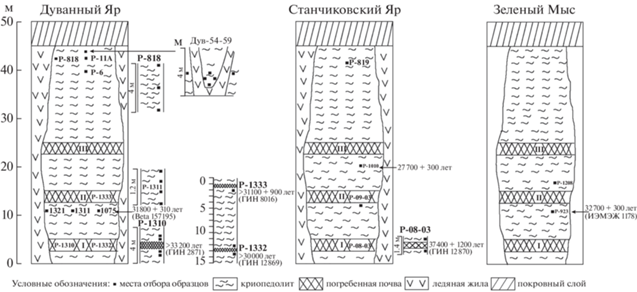
Материалом для настоящей работы послужила серия образцов, отобранных из криопедолитов каргинского и сартанского возраста и ископаемых нор в трех ключевых верхненеоплейстоценовых обнажениях Колымской низменности – Дуванный Яр, Станчиковский Яр и Зеленый Мыс (Шер, 1971; Каплина и др., 1978, 1980; Губин, 1984; Zanina et al., 2011; Murton et al., 2015; Лопатина, Занина, 2020; Lopatina, Zanina, 2020). Рассматриваемые разрезы расположены между 68° и 69° с.ш. в поясе современных предтундровых редколесий подзоны северной тайги (рис. 1).
Рис. 1.
Расположение изученных разрезов на Колымской низменности. 1 – Дуванный Яр, 2 – Станчиковский Яр, 3 – Зеленый Мыс.
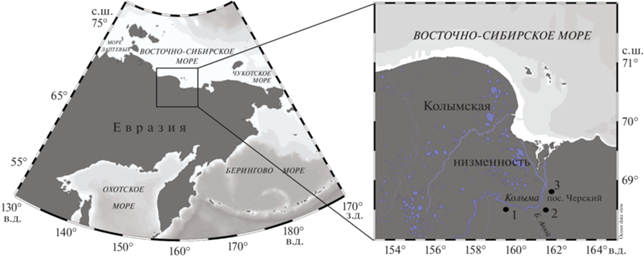
Для анализа каргинских отложений из разреза Дуванный Яр отобран материал криопедолитов, вмещающих профили I ПП (Р-1310 и Р-1332) и II ПП (Р-1333) позднекаргинского педокомплекса. Был изучен материал ископаемой норы грызуна (Р-1311) и криопедолита выше и ниже ее уровня. Из этого же слоя описаны норы 1321 и 1075. Из разреза Станчиковский Яр отобраны образцы криопедолитов, вмещающих профиль I ПП позднекаргинского педокомплекса (Р-08-03), а также материал из норы суслика 1010 и криопедолита, расположенного на одном уровне с ней. Из разреза Зеленый Мыс изучен материал из двух нор 1208 и 923. Полученные датировки погребенных почв и материала нор приведены на рис. 2. Изучение материала из ископаемых нор, на наш взгляд, здесь методически оправданно, так как ранее было показано, что днища нор как современных, так и ископаемых находились на границе сезонно-талого слоя и быстро переходили в многолетнемерзлое состояние (Занина, 2005). Это доказывают многочисленные находки остатков грызунов, в том числе их мягких тканей в ископаемых норах (Губин и др., 2003а; Faerman et al., 2017). В более ранних публикациях авторов частично приводится палинологическая характеристика отложений этих разрезов (Лопатина, Занина, 2006, 2020; Lopatina, Zanina, 2020). Сартанские отложения опробованы преимущественно на VI останце разреза Дуванный Яр. С интервалом 2 м отобрана серия образцов Р-818, взяты две отдельные пробы Р-6 и Р-11А. С целью выявления возможных отличий в химических и палеоботанических характеристиках материала в центре и по краям минерального блока в связи с его сложным генезисом при росте ледяных жил, отобрана серия проб Дув 54–59. В разрезе Станчиковский Яр взят образец Р-819 (рис. 2).
В настоящей работе проводился подсчет всех имеющихся в препарате микробиоморф, определение таксономического состава ископаемой флоры и количественная оценка поврежденных микрофоссилий. Благодаря массовой встречаемости и удовлетворительной сохранности палиноморф, спорово-пыльцевой анализ признается одним из ведущих палеоэкологических методов, применяемых при изучении верхненеоплейстоценовых отложений Колымской низменности (Каплина и др., 1978, 1980; Гитерман, 1985; Васильчук, 2003; Лопатина, Занина, 2006; Zanina et al., 2011; Murton et al., 2015). Однако таксономический состав ископаемой палинофлоры в данных отложениях однообразен, и при стратиграфическом расчленении рассматриваемых толщ и реконструкции растительности используются обычно количественные характеристики. Принимая во внимание высокую активность эоловых процессов при формировании отложений данного региона, это не всегда объективно. В спектрах в значительных количествах может содержаться заносная пыльца, которая затушевывает роль части таксонов в составе растительности, например пыльцы разнотравья, незначительное присутствие которой не отражает ее участие в составе растительности (Лопатина, Занина, 2016).
Фитолитный анализ позволяет определять не все произраставшие на территории растения, а лишь те из них, которые образуют устойчивые формы, обладающие характерной морфологией (хвойные, злаки, осоки, ряд двудольных трав, мхи). Определение таксонов до уровня семейства или рода здесь не всегда возможно, этот метод позволяет выделять комплексы, характеризующие определенный фитоценоз (Гольева, 2001). Для фитолитов не характерен занос, поскольку растительный опад более локализован по сравнению со спорами и пыльцой.
Спорово-пыльцевой и фитолитный методы анализа имеют свои области применения, их ограничения требуют внесения определенных поправок при интерпретации полученных результатов. Палинологический анализ позволяет дать характеристику региональной растительности; фитолитный анализ помогает выявить состав растительности локальных местообитаний и используется для подтверждения участия в ее составе ряда таксонов, определенных спорово-пыльцевым методом.
Обработку образцов проводили с использованием сепарационного метода В.П. Гричука без обработки мацерата ацетолизной смесью; мацераты для палинологического анализа в целях очистки от минеральных частиц дополнительно обрабатывались концентрированной плавиковой кислотой (Пыльцевой…, 1950; Палеопалинология, 1966). Подсчет палиноморф проводили до 200 зерен, фиксировалось количественное содержание палиноморф с повреждениями. Оценивалось также содержание так называемой недоразвитой пыльцы, на находки которой в спектрах из четвертичных отложений Северо-Востока Азии указывает А.К. Васильчук (2005, 2007; Васильчук Ю.К., Васильчук А.К., 2018). Это пыльца с гладкой, тонкой, иногда с тонкосетчатой экзиной, обычно с тремя неясно выраженными бороздами, без каких-либо еще четких морфологических признаков, предположительно относящаяся к разнотравью. Поскольку ее систематическое положение неясно (можно лишь предположить, что близкие по морфологическим признакам недоразвитые пыльцевые зерна могут, в частности, продуцировать представители сем. Ranunculaceae, Lamiaceae и Rosaceae), ее количество не включалось в расчетную сумму, велся подсчет ее содержания на 200 экземпляров спор и пыльцы. Количественное содержание фитолитов определялось путем их подсчета в пяти вертикальных рядах стекла 24 × 24 мм. Для фитолитов, имеющих четкие формы, рассчитывалось процентное содержание каждой формы; неопределимые и корродированные фитолиты подсчитывались отдельно, для них рассчитывался процент отношения к общей сумме неповрежденных форм. Снимки микрофитофоссилий выполнены на электронном микроскопе Tescan Vega 3 LSU в Центре коллективного пользования Института физико-химических и биологических проблем почвоведения РАН (Пущино).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Палинологический анализ изученных отложений показал, что в образцах из каргинских криопедолитов преобладает пыльца трав и кустарничков, преимущественно Poaceae и Cyperaceae. Второстепенные таксоны трав и кустарничков различной экологической приуроченности (Ericaceae, Caryophyllaceae, Asteraceae, Ranunculaceae, Fabaceae, Valeriana и др.) отмечены единично. Содержание пыльцы деревьев и кустарников (Larix, Pinus s/g Haploxylon, Betula sect. Nanae, Duschekia) обычно составляет 10–12%. При этом определен ряд спектров с повышенным содержанием пыльцы этой группы (30% и выше) – 1310 АС 1 и 1333АС 1 из разреза Дуванный Яр, 08-03 АС 2 из разреза Станчиковский Яр, 1208 и 923 из материала проб из нор разреза Зеленый Мыс. Споры занимают в спектрах подчиненное положение. Спектры из материала нор в целом сходны с таковыми, изученными из криопедолитов, отличаясь большим содержанием пыльцы злаков (более половины от всех палиноморф; пробы 1075, 1010) и гвоздичных (пробы 1075, 1311), что обусловлено использованием этих растений млекопитающими (рис. 3, табл. I ). Недоразвитая пыльца в спектрах проб из отложений, соответствующих МИС 3, единична или отсутствует, лишь в четырех пробах ее содержание составляет 20–30 экземпляров на 200 подсчитанных палиноморф. В количестве от 1 до 4 экз. на 200 палиноморф установлена переотложенная пыльца Pinus, Picea, Tsuga, Podocarpus, Juglans, Myrica, Ulmus и споры Cyathea.
Рис. 3.
Процентное содержание спор и пыльцы в каргинских отложениях разрезов Дуванный Яр, Станчиковский Яр и Зеленый Мыс. Условные обозначения см. на рис. 2.

Таблица I . Споры, пыльца, остатки диатомовых водорослей, спикулы губок из каргинских и сартанских отложений Колымской низменности. 1, 2 – Lycopodium; 3–5 – Pinus s/g Haploxylon; 6 – Cyperaceae; 7, 8 – Poaceae; 9 – недоразвитая пыльца; 10 – Ranunculaceae; 11 – Sphagnum; 12, 15, 16, 17 – панцири диатомовых водорослей; 13, 14, 18 – спикулы губок.
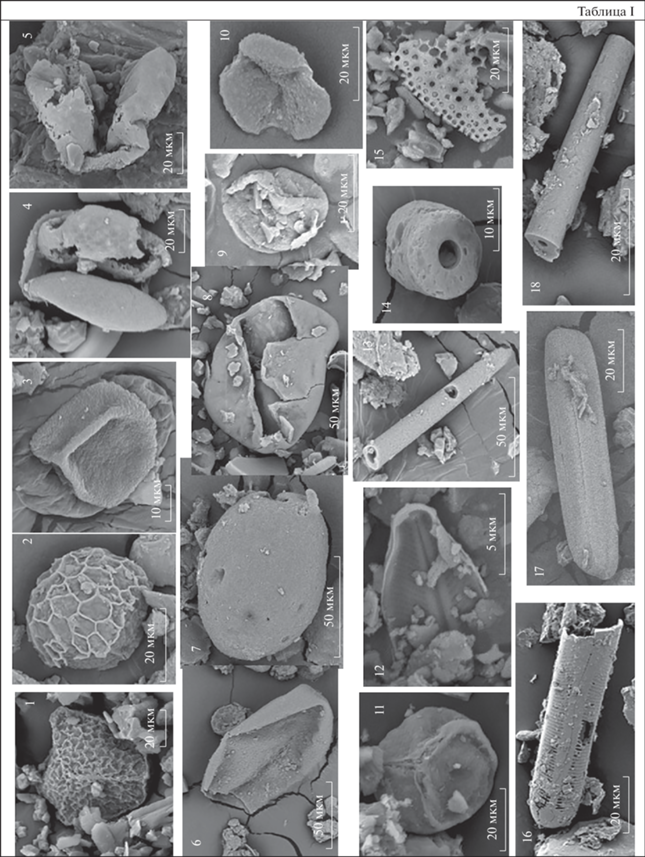
Таблица II. Формы фитолитов из каргинских и сартанских отложений Колымской низменности. 1, 6 – удлиненная гладкая форма; 2 – удлиненная шиповатая форма, 3, 5 – килевидная форма с шипом; 4 – килевидная форма с широким основанием; 7, 13–15 – трапециевидные лопастные формы; 8–10 – округлые формы; 11 – удлиненная волнистая форма; 12 – ребристая блочная форма; 16–20 – корродированные формы фитолитов.

В спектрах из сартанских криопедолитов рассматриваемых разрезов содержание пыльцы деревьев и кустарников обычно не превышает 10%. Около половины от общего количества всех палиноморф составляет пыльца Poaceae, Cyperaceae (приблизительно 10%). Таксономическое разнообразие и количество пыльцы разнотравья снижается по сравнению с каргинскими криопедолитами. Содержание спор около 20%, преобладает Selaginella rupestris (L.) Spring. (рис. 4, табл. I ). Недоразвитая пыльца отмечена во всех спектрах, ее среднее содержание составляет 40–50 зерен на 200 палиноморф. Переотложенные палиноморфы (Pinus, Picea, Tsuga, Ulmus, Carya, Osmunda) единичны (1–7 экз. на 200 палиноморф).
Рис. 4.
Процентное содержание спор и пыльцы в сартанских отложениях разрезов Дуванный Яр и Станчиковский Яр. Условные обозначения см. на рис. 2.
Результаты палинологического анализа отложений из рассматриваемых разрезов приведены в публикациях Р.Е. Гитерман (1985) и J. Murton et al. (2015). Р.Е. Гитерман на основании изучения спор и пыльцы из каргинских отложений Колымской низменности, в том числе разрезов Дуванный Яр и Станчиковский Яр, сделан вывод о чередовании теплых и холодных фаз. Спектры теплых интервалов характеризуются заметным содержанием (около 40%) пыльцы деревьев и кустарников (лиственницы, кедрового стланика, древовидной и кустарниковой берез, ольховника); в группе пыльцы трав и кустарничков, составляющей приблизительно половину от общего количества, преобладают Poaceae, Cyperaceae, разнотравье. В спектрах начала холодных фаз доминируют пыльцевые зерна различных травянистых растений, в спектрах конца этих фаз преобладает пыльца Poaceae, Artemisia, разнотравья, среди спор превалируют Selaginella rupestris. В спектрах из сартанских отложений доминируют Poaceae, Artemisia, Selaginella rupestris.
В работе J. Murton et al. (2015) указывается, что спорово-пыльцевые спектры из едомных отложений разреза Дуванный Яр характеризуются различным сочетанием пыльцы древесной и недревесной групп и трудно отличимы друг от друга. Так, в зоне D (5–26 м над урезом воды в реке (у. в. р.), 48–42 тыс. лет назад) отмечено чередование спектров с преобладанием Poaceae и разнотравья и спектров, в которых в значительном количестве (до 60%) определена пыльца деревьев и кустарников (Pinus s/g Haploxylon, Larix, Betula, Duschekia). Спектры из зоны С (26–33 м над у. в. р., 33–30 тыс. лет назад) характеризуют отложения, которые накапливались перед максимумом последнего оледенения. В них доминируют Poaceae, разнотравье и Selaginella rupestris, пыльцы деревьев и кустарников менее 10%, единично отмечен Larix. В спектрах зоны В (33–37 м над у. в. р., 25–17 тыс. лет назад), соответствующей максимуму последнего оледенения, содержание пыльцы деревьев и кустарников 20–60%, трав (Poaceae и разнотравье) 20–60%, Selaginella rupestris 10–40%. Отличием спектров этой зоны от спектров зон D и С является меньшее разнообразие таксонов и отсутствие в них лиственницы.
В результате проведенного палинологического анализа каргинских криопедолитов, нор из этих отложений и сартанских криопедолитов из разрезов Колымской низменности с учетом данных, полученных в результате более ранних исследований других авторов, установлено, что комплексы спор и пыльцы из этих отложений в целом мало отличаются друг от друга. Установить различия между ними достаточно сложно в силу приблизительно одинакового таксономического состава и колебаний содержания палиноморф. Так, в каргинских криопедолитах процентное соотношение пыльцы деревьев и кустарников 10–15%, при этом отмечены отдельные спектры с содержанием менее 10% и более 30%. В спектрах из отложений сартанского времени пыльцы деревьев и кустарников, как правило, менее 10%, но есть и до 16%. На низкое содержание пыльцы этой группы в спектрах из сартанских отложений указывает Р.Е. Гитерман (1985), тогда как в публикации J. Murton et al. (2015) отмечено, что спектры из толщ, датированных 25–17 тыс. лет назад, характеризуются высоким содержанием пыльцы этой группы, являющейся, по их мнению, заносной. Таким образом, количество пыльцы деревьев и кустарников не является критерием для подразделения этих толщ. Сходен и таксономический состав пыльцы этой группы в рассматриваемых отложениях. Но следует подчеркнуть, что в спектрах из сартанских отложений отсутствует пыльца лиственницы, единичные зерна которой определены в большинстве спектров из каргинских отложений. В спектрах из отложений, соответствующих МИС 3 и 2, преобладает пыльца трав и кустарничков с доминированием Poaceae. В спектрах из каргинских криопедолитов содоминантами в этой группе являются Cyperaceae, содержание пыльцы которых зачастую приблизительно одинаковое с Poaceae, а в некоторых пробах и выше. И это, на наш взгляд, является отличием спорово-пыльцевых спектров из этих отложений. Заметное количество пыльцы осоковых в спектрах из каргинских отложений определено Р.Е. Гитерман (1985). Кроме того, для большинства спектров их этих отложений характерно таксономическое разнообразие пыльцы разнотравья. В пробах из сартанских криопедолитов монодоминантом является семейство Poaceae, составляющее более половины от всей суммы палиноморф, тогда как содержание пыльцы Cyperaceae обычно менее 10%. Количество пыльцы разнотравья здесь несколько снижается. Состав спор в рассматриваемых отложениях одинаков, при этом их количество в сартанских отложениях выше за счет Selaginella rupestris (до 20%). Таким образом, по результатам палинологического анализа криопедолиты каргинского и сартанского возраста имеют следующие отличия: в спектрах из каргинских отложений обычна единичная пыльца Larix; они, как правило, имеют более разнообразный состав за счет трав, содоминантами Poaceae в этой группе являются Cyperaceae; в спектрах из сартанских больше спор за счет Selaginella rupestris.
Останавливаясь более подробно на анализе недоразвитой пыльцы в изученных спектрах, следует отметить следующее. Обычно это мелкие (в среднем 15 мкм) пыльцевые зерна с тремя нечетко выраженными бороздами, без каких-либо ясных морфологических признаков структуры экзины, обычно гладкой и тонкой. Предположительно это пыльца разнотравья, которая не успевала созревать из-за нарушения вегетации. Результаты исследования пыльцы нетипичного морфологического строения, без механических повреждений из ископаемых спектров приведены, в частности, в работах Е.Н. Анановой (1966), О.Ф. Дзюбы (2007), А.К. Васильчук (2005, 2007); Ю.К. Васильчук и А.К. Васильчук (2018), Т.А. Евстигнеевой (2017). По мнению этих исследователей, появление пыльцы подобного рода происходило из-за резких, экстремальных изменений природных условий, таких как пожары, повышенное ультрафиолетовое излучение, интенсивная вулканическая деятельность, понижение температуры, недостаточная влажность и пр. Е.Н. Ананова (1966) отмечает присутствие этой пыльцы в отложениях, связанных с оледенением; состав спектров, в которых присутствует пыльца подобного типа, обычно бедный. Она приводит результаты исследования современной пыльцы из незрелых соцветий: обычно она была смята, имела слабую или, напротив, густую окраску, неоформленные окончательно морфологические признаки и стратификацию экзины. По материалам А.К. Васильчук (2005), Ю.К. Васильчук и А.К. Васильчук (2018), в палиноспектрах из верхненеоплейстоценовых отложений Северо-Востока Азии недоразвитая пыльца может содержаться в количестве 60–80%. В современных поверхностных пробах она встречается редко, поскольку быстро разрушается и уничтожается микрофауной. Максимальное содержание недоразвитой пыльцы фиксируется в поверхностных спектрах из гипоарктической тундры, где растения продуцируют пыльцу в заметном количестве, но выраженные перепады температур в течение фазы мейоза оказывают негативное воздействие на ее формирование. Подобные нарушения в рассматриваемом регионе связаны с ранним и быстрым формированием снежного покрова, поздним таянием снега или его выпадением на цветущие растения, резким понижением температуры, заливом паводковыми водами растений в сезон вегетации и пр. Для того чтобы недоразвитая пыльца могла сохраниться, она почти сразу должна попасть в условия низких температур и замерзнуть (Васильчук, 2005, 2007). Авторами статьи единичные экземпляры недоразвитой пыльцы определены лишь в нескольких спектрах поверхностных проб Колымской низменности.
В спектрах проб из каргинских отложений Колымской низменности недоразвитая пыльца, как правило, определяется в незначительном количестве. В сартанских отложениях она присутствует обычно массово (40–50 экз. на 200 подсчитанных палиноморф). На наш взгляд, наличие в изученных спектрах недоразвитой пыльцы является важным показателем, свидетельствующим как о развитом травянистом покрове, так и о многократно повторяющихся неблагоприятных условиях в течение сезона вегетации и низких температурах поверхностного почвенного слоя, способствующих быстрому замерзанию палиноморф и переходу в многолетнемерзлое состояние. Разрушение недоразвитой пыльцы происходит в первую очередь, так как у нее изменены и биохимические свойства, от которых зависит прочность ее оболочки (Дзюба, 2007).
Установлено, что содержание поврежденных палинологических остатков в криопедолитах, соответствующих МИС 3, из изученных обнажений Колымской низменности составляет в среднем 8–9%, при этом зафиксированы пробы как с относительно высоким содержанием поврежденных палиноморф (14%), так и с низким (3%). Количество спор и пыльцы с повреждениями в образованиях, отвечающих МИС 2, ниже и колеблется от 1 до 5%. В изученных верхненеоплейстоценовых толщах преобладают споры и пыльца с разрушениями физического типа (разрывы и трещины), палиноморфы с повреждениями химико-биотического типа (истончение экзины, каверны), являющимися преимущественно результатом химического воздействия микробов, зафиксированы единично и спорадически. Вероятно, это обусловлено спецификой криолитозоны с преобладанием низких температур в течение года и подавленной микробиологической активностью. В пробах из каргинских образований среди поврежденной пыльцы преобладает пыльца Pinus s/g Haploxylon с разрывами и трещинами, в меньшем количестве отмечены представители семейства Poaceae. Пыльца Larix, Betula, Cyperaceae, Caryophyllaceae, споры Selaginella rupestris с повреждениями этого типа фиксируются единично и спорадически. В пробах из сартанских отложений в единичном количестве присутствует разорванная пыльца Pinus s/g Haploxylon и Poaceae, а также недоразвитая пыльца; пыльца Betula, Caryophyllaceae Asteraceae, споры Selaginella rupestris зафиксированы спорадически. Г.В. Мусиной и Р.С. Сахибгареевым (1983) при анализе сохранности палинологических остатков в отложениях установлено, что наиболее подвержены разрушению физического типа палиноморфы крупных размеров, а также пыльца голосеменных. Возможно, что этими двумя причинами обусловлено заметное содержание поврежденной пыльцы сосны в отложениях. Хотя не исключается, что часть ее является заносной, и она могла повредиться в результате транспортировки. Стабильное присутствие единичных разорванных зерен пыльцы злаков, возможно, объясняется тем, что это семейство является доминирующим в спектрах, поэтому в данном случае мы располагаем большим числом зерен для статистического анализа. Практически вся пыльца злаков и осоковых смята в складки.
Относительно большее количество в каргинских толщах пыльцы с разрывами и трещинами, возможно, обусловлено следующим. Для периода, соотносимого с МИС 3, характерно наличие сезонных повышений температуры, во время которых происходило протаивание ледяных жил или застаивание поверхностных вод на более плотных нижележащих толщах. Попадающая в осадок пыльца разбухала в условиях повышенного гидроморфизма, затем, при наступлении холодного периода, происходило ее подсушивание и вымораживание; при неоднократном повторении этих циклов на оболочках спор и пыльцы происходило образование разрывов и трещин. Температурный режим МИС 2 был более стабилен, попадавшая в осадок пыльца оказывалась в условиях низких температур, замерзала и соответственно лучше сохранялась.
Фитолитный анализ показал, что в материале, выделенном из каргинских криопедолитов, содержание фитолитов колеблется от первых десятков до нескольких сотен форм. Преобладают удлиненные гладкие формы, образующиеся в столбчатой паренхиме листьев и относимые к двудольным травам, хотя не исключается, что часть из них могла формироваться и в однодольных растениях. Их объединение в одну группу обусловлено тем, что данные формы отмечены практически во всех растениях гумидной зоны, но при этом для однодольных растений, в частности злаков, характерны и другие формы, тогда как для двудольных другие формы обычно не свойственны. Не исключается, что доминирование удлиненных гладких фитолитов связано с их строением – цельной цилиндрической формой, отсутствием перфораций и внутренней полости. Подобная конфигурация позволяет им лучше сохраняться в отложениях, меньше подвергаясь разрушительным воздействиям. Данные формы малоинформативны и не используются в качестве диагностических, но в нашем случае они подчеркивают значительную роль травянистых ассоциаций (лугового разнотравья и злаков) в составе растительности. Значительно содержание килевидных и разнообразных трапециевидных форм (трехлопастных, лопастных несимметричных, многолопастных симметричных, округлых), характерных для осок и злаков, произрастающих на влажных почвах. Это подтверждается и постоянным присутствием тканей мхов (сфагновых и политриховых). Необходимо отметить, что при этом специфические конические фитолиты осок обнаруживаются в каргинских отложениях крайне редко, вероятно разрушаясь при фоссилизации. Характерные для ксерофитной растительности шиповатые фитолиты содержатся в небольшом количестве, ребристые блочные частицы, характерные для рода Pinus, единичны (рис. 5, табл. II).
Рис. 5.
Содержание фитолитов в каргинских и сартанских отложениях разрезов Дуванный Яр и Станчиковский Яр. Условные обозначения см. на рис. 2.
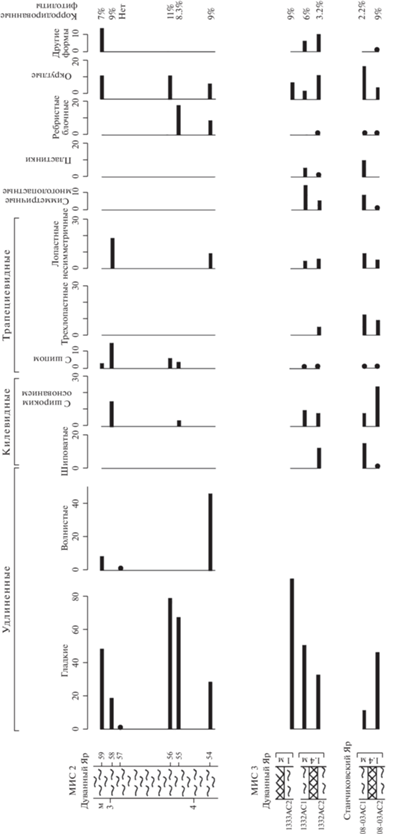
В криопедолитах сартанского времени содержание фитолитов незначительное (до 20 экз.) при низком разнообразии морфологических форм; в основном они мелкие (10–20 мкм). Низкое содержание фитолитов может быть связано со слабым накоплением аморфных форм кремнезема в органах растений и/или со слабой гумификацией растительного опада и доминированием процесса детритообразования при накоплении толщ. Большая часть экземпляров деформирована настолько, что у них невозможно диагностировать первоначальную форму. Это, вероятно, свидетельствует как о более изреженной и угнетенной растительности, так и об усилении континентальности климата в период формирования этих толщ. Как и в спектрах из каргинских отложений, здесь отмечается преобладание удлиненных гладких форм и стабильное присутствие остатков мхов, но значительно сокращается число форм, характерных для растений влажных местообитаний, обычных для спектров, соответствующих МИС 3. Так, килевидные формы здесь отсутствуют, трапециевидные формы представлены исключительно лопастными несимметричными формами и определены не во всех пробах. Установлены не отмеченные в материале из каргинских отложений удлиненные волнистые формы, характерные для злаков. В спектрах из двух образцов определены фитолиты хвойных (рис. 5, табл. II).
В изученных пробах зафиксированы корродированные и сильно изъязвленные формы фитолитов, в пробах из каргинских толщ их содержание составляет 2–9% от общей суммы, в пробах из сартанских отложений – 7–11%. В результате исследования выявлено несколько типов разрушения фитолитов: истончение мелких элементов формы (ножек, шипиков и пр.), корродирование массивного тела формы (появление каверн и сквозных перфораций), чешуйчатое повреждение (образование мелких сколов, плотно прилегающих друг к другу, создающих впечатление чешуи), сколы, появляющиеся в результате растрескивания (табл. II). Следует отметить, что повреждения выявлены также на других кремнистых микрофоссилиях – спикулах губок и панцирях диатомовых водорослей, часто встречаемых в исследованном материале. Спикулы губок округлой формы с полым центральным каналом содержатся в спектрах проб из образований, соотносимых с МИС 3 и МИС 2, в количестве 10–20 экз. (табл. I ). Приблизительно половина экземпляров обломана, зафиксирован ряд форм с чешуйчатым типом коррозии. Панцири диатомей определены только в спектрах проб из каргинских толщ и являются показателем условий повышенного гидроморфизма при формировании отложений. Количество поврежденных форм с истончением створок, особенно в районе ареол, трещинами и разрушением стенок составляет до 30%. Можно предположить, что в зоне распространения многолетнемерзлых пород разрушение кремнистых микрофитофоссилий было связано с нахождением их в сезонно-талом слое при попеременном чередовании циклов промерзания и оттаивания. В отложениях из нор корродированные фитолиты выявлены не были, что может быть связано с непродолжительным нахождением их в сезонно-талом слое и слабой деструкцией растительного материала в процессе фоссилизации. Погребенные позднеплейстоценовые почвы содержат 1–2% корродированных фитолитов, что, возможно, обусловлено как более мягкими, гумидными условиями их фоссилизации, так и общим большим количеством выявленных в них форм (Лопатина, Занина, 2020).
В пробах из рассматриваемых толщ проводился анализ растительного детрита размером 10–200 мкм. Его обычным компонентом в каргинских отложениях являются специфические формы с окаймленными порами, характерные для древесины лиственницы. В значительном количестве зафиксирован детрит мхов и остатки эпидермиса трав Cyperaceae, Poaceae и Ericaceae, различающихся по устьичным комплексам. Состав растительного детрита из сартанских криопедолитов более бедный, в его составе определены остатки злаков и мхов.
В публикациях Б.Ф. Хасанова (1999), С.В. Губина с соавторами (2001, 2003а) приведены результаты изучения крупномерных растительных остатков из нор сусликов рассматриваемых разрезов. Остатки древесной растительности (древесина Larix cajanderi Mayr.) обнаружены лишь в норах обнажения Зеленый Мыс, кустарники (обломки веточек и листьев ив) обычны в составе подстилки. На первом месте в комплексе карпологических остатков среди трав стоят психрофиты (растения, произрастающие на влажных и холодных почвах), преобладают лапчатка снежная Potentilla nivea L., змеевик живородящий Bistorta vivipara (L.) S.F. Gray, мятлик арктический Poa arctica R. Br., щавель арктический Rumex arcticus Trautv, различные виды осок Carex bonanzensis Britt., C. vesicata Meinsh., C. norvegica Retz. В значительном количестве представлены семена степных растений – мятликов оттянутого и кистевидного Poa attenuata Trin. и P. botryoides (Trin. ex Griseb.) Roshev, смолевки узколистной Silene stenophylla Ledeb., пионеров сухих местообитаний – крупки Draba, подорожника седоватого Plantago canescens Adams и пионеров влажных местообитаний – Ranunculus repens L. Эти виды встречаются в современных тундрах севера Якутии и позволяют диагностировать растительный покров участков, расположенных вблизи норы, как тундровый с участием нарушенных местообитаний, занятых пионерной и степной растительностью. Единственная ископаемая нора мелкого грызуна из сартанских отложений описана из разреза Дуванный Яр. Из нее определены обломки веточек дриады Dryas sp., брусники Vaccinum vitis-idaea L., полярной ивы Salix polaris Wahlenb, единичные семена пионеров сухих местообитаний Draba sp., Plantago canescens, Taraxacum lateritum Dahlst, степных растений – лука торчащего Allium strictum Schrad., Poa botryoides и психрофита Potentilla nivea, остатки мхов и лишайников. Видовой состав приведенных макростатков растений указывает на существование низкопродуктивного степного сообщества в окрестностях рассматриваемой норы на фоне развития тундровой растительности (Губин и др., 2003а).
Анализ спор, пыльцы, фитолитов и растительного детрита с учетом результатов изучения карпологического материала позволяет реконструировать для интервала, являющегося аналогом МИС 3, тундровые ландшафты с преобладанием травянистых группировок (злаковых, злаково-осоковых, злаково-разнотравных). Установленное по палинологическим данным преобладание злаков в растительном покрове подтверждается данными анализа фитолитов, растительного детрита, содержимого ископаемых нор, в которых остатки этих растений определены в большом количестве. Преобладание в комплексах карпологических остатков психрофитов, присутствиe панцирей диатомовых водорослей, а среди фитолитов форм, характерных для увлажненных ценозов, свидетельствует о широком распространении тундровых лугов на влажных почвах. Они могли развиваться в результате увеличения мощности и увлажнения сезонно-талого слоя из-за подтаивания повторно-жильных льдов при некотором потеплении климата. Присутствие пыльцы разнотравья в изученных спектрах, как правило, единично. Анализ палинологических остатков из субрецентных спектров Колымской низменности показал, что пыльца сем. трав и кустарничков в них не всегда отражает многообразие локальной растительности, а ее количество (в частности, сем. Onagraceae, Lamiaceae, Ranunculaceae Liliaceae и др.) – адекватное присутствие тех или иных семейств в ее составе. Это обусловлено невысокой пыльцевой продуктивностью этих растений в суровых климатических условиях и переходом их на вегетативное размножение (Лопатина, Занина, 2016). На основании этого можно предположить значительную роль разнообразных трав и кустарничков в составе растительности каргинского и сартанского времени. Провести экологическую интерпретацию пыльцы разнотравья в спектрах сложно, поскольку семейства гвоздичных, сложноцветных, бобовых, лютиковых, гречишных и др. представлены большим числом видов различной экологической приуроченности, однако их присутствие в целом соответствует данной реконструкции. Разнообразный состав фитолитов подчеркивает богатый состав растительности. На заболоченных участках произрастали осоки, пыльца которых является в спорово-пыльцевых спектрах субдоминатом, а находки их плодов обычны в норах. В то же время в результате подтаивания повторно-жильных льдов на рассматриваемой территории активизировались склоновые и эрозионные процессы, шло интенсивное образование осыпей, заселяемых пионерной растительностью – крупкой, подорожником, которая в процессе сукцессионных смен заменялась степной, злаково-разнотравными ассоциациями. Присутствие в комплексе фитолитов в небольшом количестве форм с шипами, характерных для ксерофитной, в том числе степной, флоры, подтверждает данный вывод.
Пыльца деревьев и кустарников составляет в рассматриваемых спектрах обычно 10–12%, в некоторых выше 30%. Можно предположить, что вариации, хотя и незначительные, в соотношении групп растительности, основных и второстепенных таксонов в спектрах проб из каргинских криопедолитов свидетельствуют о климатических флуктуациях в течение каргинского времени и периодическом увеличении или, наоборот, уменьшении роли редкостойных лиственничных лесов на территории, занятой преимущественно травянисто-кустарничковой тундровой растительностью. В комплексе макроостатков определены лишь Larix и Salix. Отсутствие растений этой группы не может быть объяснено минерализацией в результате погребения и почвообразования, поскольку в рассматриваемых толщах присутствует значительное количество остатков травянистой и моховой растительности в прижизненном положении, быстрее подвергающейся гумификации и минерализации по сравнению с древесиной. Палеопочвенные исследования рассматриваемых толщ не выявили признаков лесного почвообразования в слоях каргинских криопедолитов (Губин, Занина, 2013). Принимая во внимание очень плохую сохранность пыльцы лиственницы Каяндера (Larix cajanderi Mayr) и ее малую миграционную способность, единичную находку ее древесины в норе Зеленого Мыса, а также присутствие в растительном детрите остатков, характерных для ее древесины, можно предположить участие в составе реконструируемой растительности небольших островков лиственничных лесов. Присутствие древесины этого рода в норах арктического суслика, жителя открытых пространств, может указывать на существование в местах его обитания лишь лиственничных редколесий. Учитывая высокую продуктивность и миграционную способность сосны, обычно трудно оценить ее роль в растительном покрове. В субрецентных спектрах Колымской низменности значительная ее часть является заносной (Лопатина, Занина, 2016). В спектрах проб из каргинских и сартанских отложений среди экземпляров пыльцы Pinus s/g Haploxylon много разорванных форм, что, в частности, может быть последствием транспортировки и позволяет предположить ее заносное происхождение. Однако находки в исследуемых толщах фитолитов хвойных, имеющих обычно хорошую сохранность благодаря значительному количеству смолистых веществ в составе хвои, препятствующих их быстрому разрушению, свидетельствуют об участии кедрового стланика в составе растительности, возможно в качестве подлеска в лиственничниках или небольших зарослей в долинах.
Споры занимают в спектрах подчиненное положение, однако в норах в большом количестве обнаружены вегетативные побеги мхов, а в фитолитных спектрах стабильно присутствуют характерные для них формы, а также ткани, что позволяет сделать вывод о достаточно развитом моховом покрове на рассматриваемой территории.
Результаты анализа микрофитофоссилий свидетельствуют о развитии на рассматриваемой территории в течение интервала, синхронного МИС 2, тундровых растительных сообществ, преимущественно злаковых и злаково-разнотравных, более обедненных по сравнению с таковыми МИС 3. Наличие в заметном количестве в спектрах проб из сартанских отложений недоразвитой пыльцы, предположительно относящейся к разнотравью, позволяет реконструировать для этого времени развитие лугов и, возможно, неблагоприятные условия сезона вегетации, не способствующие полному формированию пыльцевых зерен. Значительное сокращение среди фитолитов числа форм, характерных для влажных ценозов, присутствие пыльцы и карпологических остатков сложноцветных, гвоздичных, бобовых, лапчатки, крупки свидетельствует о широком распространении сухих, хорошо дренируемых участков. Обилие в спектрах спор плаунка наскального указывает на наличие сухих каменистых осыпей на склонах.
ВЫВОДЫ
Изучение микрофитофоссилий, как биоиндикаторов растительных сообществ из каргинских и сартанских отложений Колымской низменности, позволило получить дополнительную информацию по генезису исследуемых толщ, более полно охарактеризовать состав растительности и ландшафта и детализировать реконструкции природной среды.
Состав палиноморф в пробах из каргинских и сартанских отложений практически сходный, преобладает пыльца трав и кустарничков с доминированием Poaceae. Их различия, которые следует принимать во внимание при стратиграфическом расчленении, следующие: более разнообразный состав трав, участие Cyperaceae в качестве содоминантов Poaceae, присутствие единичной пыльцы Larix в спектрах из каргинских отложений, а также большее количество спор за счет Selaginella rupestris и недоразвитой пыльцы в сартанских образованиях.
В комплексе фитолитов из каргинских толщ преобладают формы, характерные для полевого и лугового разнотравья и злаков, присутствует значительное количество форм, принадлежащих осокам и злакам, произрастающим на влажных почвах, фитолитов мхов, а также отмечено единичное содержание форм, характерных для ксерофитной флоры, и хвойных. Содержание фитолитов в сартанских криопедолитах ниже по сравнению с таковым в каргинских, их размеры мельче, разнообразие морфотипов ниже, что указывает на существование в этот период относительно изреженной и угнетенной растительности. Отмечается стабильное присутствие фитолитов двудольных трав и остатков мхов, но значительно сокращается количество и разнообразие форм, характерных для влаголюбивых растений, обычных для спектров из толщ, соответствующих МИС 3. С помощью фитолитного анализа подтверждена роль в составе растительности ряда растений, в частности мхов, вегетативные побеги которых многочисленны в норах, но при этом в палинологических спектрах их количество невелико, а также разнотравья, определяемого в спектрах в виде единичных зерен и недоразвитой пыльцы. Установлено участие в составе растительных группировок кедрового стланика, пыльца которого в спектрах может рассматриваться как заносная при отсутствии этого рода в комплексе микро- и макроостатков.
Знание особенностей морфологии современной пыльцы, сформировавшейся в условиях неблагоприятной экологической обстановки, специфики процессов, связанных с ее формированием в таких условиях, позволяет вывить в спектрах недоразвитую пыльцу, предположительно относящуюся к разнотравью. Ее массовое присутствие позволяет предположить развитый травянистый покров, преимущественно неблагоприятные условия сезона вегетации и низкие температуры поверхностного слоя почвы, способствующие ее криоконсервации. Присутствие пыльцы подобного рода в ископаемых спектрах может быть использовано в качестве дополнительного критерия при реконструкции состояния климата и ландшафтов прошлого.
Состав спор, пыльцы и фитолитов из каргинских слоев позволяет сделать вывод о том, что в течение данного интервала времени на рассматриваемой территории существовали разнообразные ландшафты – сообщества влажных и суховатых тундр, лиственничных редколесий и тундровых болот с включением местообитаний, занятых пионерной и степной растительностью. Формирование криопедолитов сартанского интервала проходило в более однообразных, холодных и сухих условиях на протяжении всей стадии их накопления. Данные анализа микробиоморф из этих отложений подтверждают факт активного участия, вплоть до полного доминирования, в этот период на территории изучаемого района травянистых, преимущественно злаковых ассоциаций в комплексе с наземным моховым покровом.
Анализ сохранности палиноморф из отложений, синхронных МИС 3 и МИС 2, Колымской низменности показал, что повреждения химико-биотического типа, обусловленные действием микробов, на них единичны. Возможно, это объясняется спецификой криолитозоны с преобладанием низких температур в течение года и низкой микробиологической активностью. Содержание палиноморф с повреждениями физического типа, к появлению которых, вероятнее всего, привели попеременное подсушивание и увлажнение, обычно не превышает в каргинских толщах 10%, в сартанских – 5%. Для периода накопления каргинских отложений характерно более выраженное проявление сезонного повышения температуры, во время которого происходило таяние ледяных жил и застаивание вод в понижениях рельефа. В условиях повышенной влажности попавшие в осадок палиноморфы разбухали, при наступлении холодного периода происходило их замерзание и подсушивание; в результате повторения этих циклов на оболочках спор и пыльцы образовывались разрывы и трещины. Условия сартанского времени были более стабильны, споры и пыльца при низких температурах лучше сохранялись. На это указывают также массовые находки в этих отложениях недоразвитой пыльцы. Полученные данные дополняют представления о характере преобразования растительного материала в процессе криогенеза и могут служить критерием при интерпретации результатов анализа микрофитофоссилий из плейстоценовых отложений криолитозоны.
Источники финансирования. Работа выполнена при финансовой поддержке РФФИ в рамках научного проекта № 20-05-00559 А, обработка образцов производилась по проекту № 19-29-05003 МК; полевые исследования обнажений Колымской низменности выполнены по программе госзадания ИФХиБПП РАН, анализ палинологического материала – по госзаданию ГИН РАН.
Список литературы
Ананова Е.Н. О недоразвитой пыльце в плейстоценовых отложениях // Комиссия по изучению четвертичного периода. 1966. № 32. С. 18–22.
Васильчук А.К. Отражение событий Хайнриха на датированных по радиоуглероду спорово-пыльцевых диаграммах повторно-жильных льдов и вмещающих их едомных отложений низовий р. Колымы // Криосфера Земли. 2003. Т. VII. № 4. С. 3–13.
Васильчук А.К. Особенности формирования палиноспектров в криолитозоне России. М.: Изд-во МГУ, 2005. 245 с.
Васильчук А.К. Палинология и хронология полигонально-жильных комплексов в криолитозоне России. М.: Изд-во МГУ, 2007. 488 с.
Васильчук Ю.К., Васильчук А.К. Зимние палеотемпературы воздуха в низовьях Колымы 30–12 тысяч лет назад по результатам изучения изотопного состава едомы Плахинского Яра // Криосфера Земли. 2018. Т. XXII. № 5. С. 3–19.
Гитерман Р.Е. История растительности Северо-Востока СССР в плиоцене и плейстоцене. М.: Наука, 1985. 95 с.
Гольева А.А. Фитолиты и их информационная роль при исследовании природных и археологических объектов. М.: Полтекс, 2001. 200 с.
Губин С.В. Палеопедологический анализ верхнеплейстоценовых (едомных) отложений обнажения Дуванный Яр // Бюлл. Комиссии по изучению четвертичного периода. 1984. № 53. С. 125–128.
Губин С.В. Позднеплейстоценовое почвообразование на Приморских низменностях на севере Якутии // Почвоведение. 1994. № 8. С. 5–14.
Губин С.В. Почвообразование сартанского криохрона в западном секторе Берингии // Почвоведение. 1998. № 5. С. 605–609.
Губин С.В. Педогенез – составная часть механизма формирования отложений позднеплейстоценового ледового комплекса // Криосфера Земли. 2002. Т. VI. № 3. С. 82–91.
Губин С.В., Занина О.Г. Изменение почвенного покрова в ходе формирования отложений ледового комплекса на Колымской низменности (часть 1) // Криосфера Земли. 2013. Т. XVII. № 4. С. 48–56.
Губин С.В., Занина О.Г. Изменение почвенного покрова в ходе формирования отложений ледового комплекса на Колымской низменности (часть 2) // Криосфера Земли. 2014. Т. XVIII. № 1. С. 77–82.
Губин С.В., Максимович С.В., Занина О.Г. Анализ состава семян растений из ископаемых нор сусликов лессово-ледовых отложений обнажения Зеленый Мыс как показатель местных условий обитания // Криосфера Земли. 2001. Т. V. № 2. С. 76–82.
Губин С.В., Занина О.Г., Максимович С.В., Кузьмина С.А., Зажигин В.С. Реконструкция условий формирования отложений ледового комплекса по результатам изучения позднеплейстоценовых нор грызунов // Криосфера Земли. 2003а. Т. VII. № 3. С. 13–22.
Губин С.В., Максимович С.В., Давыдов С.П., Гиличинский Д.А., Шатилович А.В., Спирина Е.В., Яшина С.Г. О возможности участия позднеплейстоценовой биоты в формировании биоразнообразия современной криолитозоны // Журн. общей биологии. 2003б. Т. 64. № 2. С. 160–165.
Дзюба О.Ф. Тератоморфные пыльцевые зерна в современных и палеопалинологических спектрах и некоторые проблемы палиностратиграфии // Нефтегазовая геология. Теория и практика. 2007. Т. 2. С. 1–21.
Евстигнеева Т.А. Морфологические аномалии ископаемой пыльцы сосен как индикатор климатических условий прошлого // Современные проблемы науки и образования. 2017. № 5. С. 1–8.
Занина О.Г. Ископаемые норы грызунов из мерзлых позднеплейстоценовых отложений Колымской низменности // Зоологический журн. 2005. Т. 84. № 6. С. 728–736.
Занина О.Г. Почвообразование и природные условия каргинского времени на Колымской низменности. Автореф. дисс. … канд. географ. наук. 2006.
Каплина Т.Н. Аласные комплексы Северной Якутии // Криосфера Земли. 2009. Т. ХIII. Вып. 4. С. 3–17.
Каплина Т.Н., Гитерман Р.Е., Лахтина О.В., Абрашов Б.А., Киселев С.В., Шер А.В. Дуванный Яр – опорный разрез позднеплейстоценовых отложений Колымской низменности // Бюлл. Комиссии по изучению четвертичного периода. 1978. № 48. С. 49–65.
Каплина Т.Н., Лахтина О.В., Рыбакова Н.О. История развития ландшафтов и мерзлых толщ Колымской низменности по радиоуглеродным, криолитологическим и палинологическим данным (на примере разреза Станчиковский яр на р. Мал. Анюй) // Геохронология четвертичного периода. М.: Наука, 1980. С. 243–258.
Лопатина Д.А., Занина О.Г. Палеоботанический анализ материала ископаемых нор сусликов и вмещающих их верхнеплейстоценовых отложений низовий р. Колымы // Стратиграфия. Геол. корреляция. 2006. Т. 14. № 5. С. 94–107.
Лопатина Д.А., Занина О.Г. Субрецентные спорово-пыльцевые спектры низовьев р. Колыма и их значение для реконструкции четвертичной палеогеографии региона // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 2. С. 103–112.
Лопатина Д.А., Занина О.Г. Условия формирования палеопочв времени МИС 3 Колымской низменности по данным палинологического и фитолитного методов // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 3. С. 134–147.
Мусина Г.В., Сахибгареев Р.С. Разрушение оболочек пыльцы и спор в процессе литогенеза // Стратиграфия и корреляция осадков методами палинологии. Свердловск: УНЦ АН СССР, 1983. С. 154–160.
Палеопалинология. Т. I. Ред. Покровская И.М. Л.: Недра, 1966. 352 с.
Пыльцевой анализ. Ред. Криштофович А.Н. М.: Госгеолиздат, 1950. 571 с.
Томирдиаро С.В. Лёссово-ледовая формация Восточной Сибири в позднем плейстоцене и голоцене. М.: Наука, 1980. 184 с.
Хасанов Б.Ф. Ботанический анализ кормовых запасов нор плейстоценового арктического суслика из Колымско-Индигирской низменности // Зоологический журн. 1999. Т. 78. № 2. С. 240–244.
Шер А.В. Млекопитающие и стратиграфия плейстоцена крайнего Северо-Востока СССР и Северной Америки. М.: Наука, 1971. 310 с.
Яшина С.Г., Губин С.В., Шабаева Э.В., Егорова Е.Ф., Максимович С.В. Жизнеспособность семян высших растений позднеплейстоценового возраста из вечномерзлых отложений, обнаруживаемая в культуре in vitro // Докл. АН. 2002. Т. 383. № 5. С. 714–717.
Faerman M., Bar-Gal Gila Kahila, Boaretto E., Boeskorov G., Dokuchaev N., Ermakov O., Golenishchev F., Gubin S., Mintz E., Simonov E., Surin V., Titov S., Zanina O., Formozov N. DNA analysis of a 30.000-year-old Urocitellus glacialis from northeastern Siberia reveals phylogenetic relationships between ancient and present-day arctic ground squirrels // Sci. Reports. 2017. V. 7. № 1. P. 1–10.
Lopatina D.A., Zanina O.G. Microbiomorphic analysis of paleosols from the Upper Quaternary deposits of the Kolyma Lowland // Paleontol. J. 2020. V. 54. № 8. P. 45–53.
Murton J., Goslar T., Edwards M., Bateman M., Danilov P., Savvinov G., Gubin S., Ghaleb B., Haile J., Kanevskiy M., Lozhkin A., Lupachev A., Murton D., Shur Yu., Tikhonov A., Vasil’chuk A., Vasil’chuk Yu., Wolfe S. Palaeoenvironmental interpretation of edoma silt (ice complex) deposition as cold-climate loess, Duvanny Yar (Northeast Siberia) // Permaphrost and Periglacial Processes. 2015. V. 26. P. 208–288.
Zanina O.G., Gubin S.V., Kuzmina S.A. Maximovich S.V., Lopatina D.A. Late-Pleistocene (MIS 3–2) palaeoenvironments as recorded by sediments, palaeosols, and ground-squirrel nests at Duvanny Yar, Kolyma lowland, northeast Siberia // Quaternary Sci. Rev. 2011. V. 30. P. 2107–2123.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция