

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 2, стр. 71-100
Кимеридж-волжские палинологические комплексы разреза Еганово (Московская область) и возможности их использования для биостратиграфии, корреляции и фациального анализа
Е. Б. Пещевицкая 1, *, А. В. Лидская 2, Ю. И. Ростовцева 3
1 Институт нефтегазовой геологии и геофизики СО РАН
Новосибирск, Россия
2 Геологический институт РАН
Москва, Россия
3 Московский государственный университет им. М.В. Ломоносова
Москва, Россия
* E-mail: PeschevickayaEB@ipgg.sbras.ru
Поступила в редакцию 27.08.2021
После доработки 26.10.2021
Принята к публикации 07.11.2021
- EDN: DWNWMG
- DOI: 10.31857/S0869592X22020053
Аннотация
Приведены результаты изучения палиноморф морского и наземного генезиса в нижнем кимеридже, средне- и верхневолжском подъярусах в разрезе Еганово (Московская обл.). Разрез хорошо охарактеризован аммонитами, что обеспечивает биостратиграфический контроль установленных палиностратонов. Слои со спорами и пыльцой установлены для волжского интервала, слои с диноцистами – для всего разреза. Определены таксоны, которые позволяют сопоставлять синхронные последовательности диноцист европейской части России, Сибири и Западной Европы. По спорам и пыльце возможна корреляция с одновозрастными отложениями севера Урала, Западной Европы и Австралии. Фациальный анализ двух палинологических групп позволил восстановить особенности морских альгофлор и их взаимосвязь с трансгрессивно-регрессивной динамикой палеобассейна, климатическими колебаниями и изменением прибрежных ландшафтов. Сравнительный анализ латерального распределения одновозрастных диноцистовых ассоциаций из разреза Еганово и других бореальных районов показал существенное сходство альгофлор европейской России, Североморского региона и Болгарии в кимеридже, но к концу волжского века усилилась латеральная дифференциация альгофлор. В то же время разнообразие диноцистовых ассоциаций в европейской части России оставалось высоким. Сделано предположение, что это было обусловлено благоприятными температурными и трофическими условиями, которые восстанавливаются с привлечением результатов фациального анализа наземных палиноморф. Особенности таксономического состава последних характерны для Евро-Синийской палеогеографической области с субтропическим климатом. На фоне постепенного похолодания во второй половине волжского века восстанавливаются небольшие климатические максимумы, которые прослеживаются также в Западной Европе, Сибири и на севере Урала. Период аридизации, характерный для терминальной юры Западной Европы, не выявлен. Достаточно влажные условия привели к формированию в европейской части России и на прилегающих с востока территориях Урала и окраин Западной Сибири своеобразных палинофлор, отличающихся обилием глейхениевых папоротников.
ВВЕДЕНИЕ
Палинологические исследования верхней юры на территории европейской части России успешно ведутся с середины прошлого столетия. За это время накопился обширный материал, который не весь опубликован, но активно использовался при составлении нескольких поколений региональных стратиграфических схем юрских отложений Восточно-Европейской платформы (Митта и др., 2012 и др.). Спорово-пыльцевыми данными хорошо охарактеризован волжский интервал на территории Московской и Костромской областей, Среднего Поволжья и других районов. В то же время данных по кимериджу очень немного, поскольку в палинологических спектрах здесь доминирует микрофитопланктон, а наземные палиноморфы встречаются в основном единично (Шрамкова, 1970; Орлова, Цыбин, 1972; Добруцкая, 1973; Болховитина, 1973; Селькова и др., 2011 и др.). В этих работах охарактеризован качественный и количественный состав палинокомплексов отдельных ярусов, их частей или аммонитовых зон, приведено сравнение с комплексами сопредельных территорий. Однако в большинстве случаев отсутствуют детальные привязки к подразделениям, установленным по другим группам биофоссилий. Биостратиграфические подразделения по палинологическим данным не устанавливались. В результате не были четко определены биостратиграфические рубежи, на которых происходили наиболее важные изменения в составе наземных палиноморф, их положение относительно стандартной региональной шкалы и палинологические критерии, что позволяло бы самостоятельно расчленять разрезы по спорам и пыльце и проводить корреляцию с палиностратонами других территорий.
Вместе с тем биостратиграфические возможности диноцист были использованы в полной мере: по материалам из Среднего и Верхнего Поволжья и Печорской синеклизы для бат-рязанского интервала установлена непрерывная последовательность диноцистовых зон для европейской части России, выявлены биостратиграфические маркеры и коррелятивные таксоны для некоторых уровней (Riding et al., 1999). Определенные уточнения и дополнения внесены в кимеридж-волжскую часть диноцистовой шкалы по материалам из разрезов Городищи и Кашпир (Harding et al., 2011; Пещевицкая, 2021).
В основу настоящего исследования было положено совместное изучение последовательностей кимеридж-волжских палиноморф морского и наземного генезиса по материалам из разреза Еганово (Московская обл.), хорошо датированного по аммонитам, с целью уточнения и/или дополнения диноцистовой шкалы, выделения самостоятельных палиностратонов по спорам и пыльце, а также оценки корреляционной возможности палиностратиграфических рубежей для сопоставления как местных, так и удаленных разрезов. Палинологическое изучение разреза проведено впервые. Также ожидается, что совместный биофациальный анализ двух палинологических групп позволит одновременно реконструировать условия в морском палеобассейне и обстановки на прилегающих участках суши, проследить их сопряженную динамику во времени и взаимное влияние.
МАТЕРИАЛЫ И МЕТОДЫ
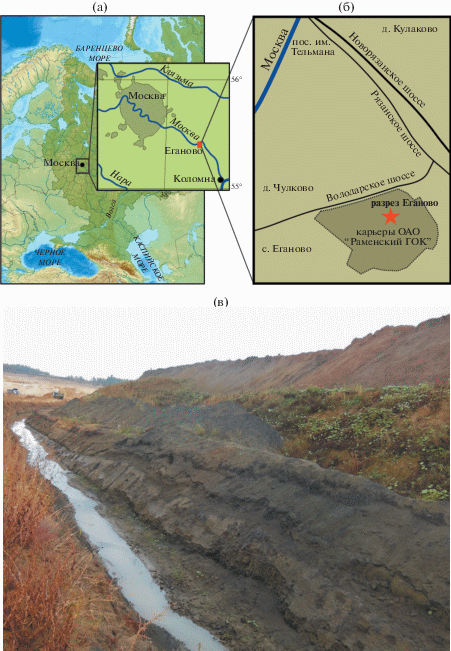
Изученный разрез расположен в Раменском районе Московской области недалеко от д. Еганово (рис. 1). Он вскрыт дренажными канавами карьера ОАО “Раменский ГОК”, который ведет разработку стекольных песков (Ширшов, 2005). Разрез представлен глинами, алевритами и песками кимериджа и волжского яруса. Подробное описание и детальное расчленение разреза по аммонитам приведены в работе М.А. Рогова (2017). В разрезе установлены аммонитовая зона (а-зона) bauhini в нижнем кимеридже и последовательность зон panderi–catenulatum в волжской части, а также входящие в состав этих зон биогоризонты. Таким образом, палинологический материал имеет точные привязки, и уровни его отбора детально выверены по ортостратиграфической группе.
Рис. 1.
Разрез Еганово: (а) – общее местоположение разреза; (б) – схема расположения карьеров ОАО “Раменский ГОК”; (в) – фотография разреза Еганово.
Образцы на палинологический анализ отобраны в 2017–2018 гг. на полевой экскурсии, приуроченной к VII Всероссийскому совещанию “Юрская система России: проблемы стратиграфии и палеогеографии”, и во время экспедиционных работ под руководством М.А. Рогова (ГИН РАН, Москва). Лабораторная подготовка образцов включала последовательно обработку горячим 10%-ным раствором пирофосфата натрия, отмучивание глинистой фракции и центрифугирование в тяжелой жидкости (CdI + KI, у.в. 2.25) для выделения органической фракции. Для части проб применялась обработка плавиковой и соляной кислотами, а также ацетолиз. Количественный подсчет палиноморф в образце проводился минимум до 200 экз. сначала совместно для морских и наземных форм, чтобы установить их соотношение, затем в той и другой группе отдельно. Биостратиграфический анализ и выделение слоев с палинофлорой, как биостратонов комплексного обоснования, базировались, в первую очередь, на появлении/исчезновении стратиграфически важных таксонов, которые прослеживаются на обширных территориях и интервалы распространения которых подтверждены фаунистическими датировками. Динамика количества и разнообразия были использованы в качестве дополнительных критериев. Для биофациального и палеогеографического анализа микрофитопланктона и наземных палиноморф были привлечены литературные данные и существующие концепции по этим вопросам (Вахрамеев, 1988; Leereveld, 1995; Jansonius, McGregor, 1996; Abbink, 1998; Van Konijnenburg-Van Cittert, 2002 и др.). Для оценки степени сходства ассоциаций диноцист на севере Евразии применен коэффициент Жаккара (Jaccard, 1901).
РАСЧЛЕНЕНИЕ РАЗРЕЗА ПО ДИНОЦИСТАМ
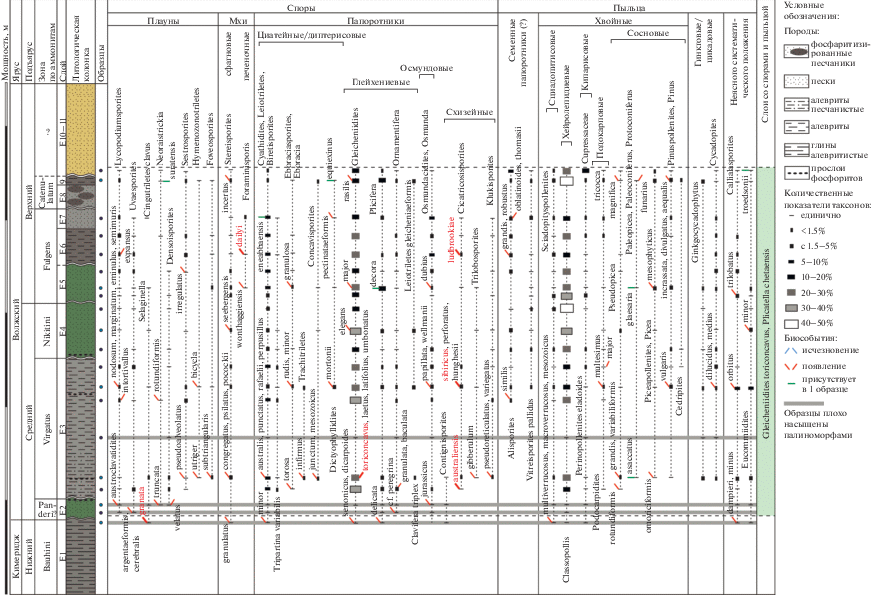
Морские палиноморфы представлены разнообразными диноцистами, акритархами, празинофитами и выстилками микрофораминифер. Иногда присутствуют зеленые водоросли озерного генезиса (Botryococcus). В разрезе установлено два биостратона в ранге слоев с диноцистами. Распределение диноцист в разрезе приведено на рис. 2.

Слои с Cribroperidinium? longicorne, Systematophora? daveyi
Характерный комплекс. Доминируют проксиматные (16%) и хоратные (18%) диноцисты плохой сохранности, Systematophora spp. (14%), Impletosphaeridium varispinosum (Sarjeant) Islam (16%). Часто встречаются (7%) диноцисты рода Cribroperidinium (в том числе C.? longicorne (Downie) Lentin et Williams, C.? ehrenbergii (Gitmez) Helenes, C. angulosum (Gitmez) Poulsen). Немногочисленны представители группы Pilosidinium (роды Pilosidinium, Sentusidinium, Barbatacysta; 4%). Определены диноцисты родов Dingodinium, Evansia, Gonyaulacysta, Rhynchodiniopsis, Leptodinium, Scriniodinium и др. Также определены акритархи Solisphaeridium stimuluferum (Deflandre) Pocock (1%) и празинофиты родов Leiosphaeridia (2.5%), Pterospermella (1.5%). Стратиграфически важные виды перечислены ниже.
Стратиграфически важные черты. Присутствуют диноцисты, которые появляются на севере Западной Европы с верхнего оксфорда (Occisucysta balios Gitmez – а-зона rosenkranzi, Egmontodinium polyplacophorum Gitmez et Sarjeant – а-зона serratum), а также таксоны, появление которых на севере Западной Европы отмечается в нижнем кимеридже: C. angulosum, C.? ehrenbergii (а-зона baylei) и C.? longicorne, Systematophora? daveyi Riding et Thomas (а-зона cymodoce) (Sarjeant, 1979; Thomas, Cox, 1988; Powell, 1992; Poulsen, Riding, 2003).
Границы. Нижняя граница не определена, условно проводится по нижнему уровню опробования, верхняя граница совмещается с литологической границей между слоями Е1 и Е2.
Стратиграфическое положение. Верхняя часть слоя Е1, а-зона bauhini, нижний кимеридж.
Слои с Gochteodinia villosa, Isthmocystis distincta
Характерный комплекс. Доминируют проксиматные диноцисты плохой сохранности (17–29%), Cribroperidinium (2.5–43%). Иногда обильны представители группы Escharisphaeridia (1–5, до 17%). Достаточно многочисленны проксимохоратные диноцисты плохой сохранности (2–14%), группы Pilosidinium (3.5–11%) и Circulodinium–Tenua (1–10%), иногда Dingodinium (до 5%), Apteodinium (до 7%), Stiphrosphaeridium (до 5.5%), Perisseiasphaeridium (до 5%), Senoniasphaera jurassica (Gitmez et Sarjeant) Lentin et Williams (до 7%), Impletosphaeridium (до 5%), Cassiculosphaeridia (до 15%). Постоянно в небольшом количестве встречаются диноцисты родов Pareodinia, Tubotuberella, Leptodinium, Rhynchodiniopsis, Meiourogonyaulax, Scriniodinium и др. Стратиграфически важные виды перечислены ниже.
Стратиграфически важные черты. Вид Cribroperidinium hansenii Poulsen отмечается с низов портланда (а-зона albani) Дании (Poulsen, 1996) и нижней части средневолжского подъяруса (а-зона panderi) на Русской плите (Пещевицкая, 2021). Наиболее древние находки рода Stiphrosphaeridium зафиксированы на Русской плите в разрезе Городищи (верхняя часть а-зоны panderi) (Пещевицкая, 2021). Виды Isthmocystis distincta Duxbury и Gochteodinia villosa (Vozzhennikova) Norris появляются в середине портланда (а-зона kerberus) на севере Западной Европы (Powell, 1992; Poulsen, Riding, 2003). Последний вид также присутствует со средневолжского подъяруса в разрезах на Русской плите (а-зона nikitini) (Harding et al., 2011; Пещевицкая, 2021) и севере Сибири (верхи фораминиферовой зоны JF55, что соответствует нижней части а-зоны okensis) (Nikitenko et al., 2018). Achomosphaera neptuni (Eisenack) Davey et Williams обнаружен в переходных средне-верхневолжских слоях в Дании (Heilmann-Clausen, 1987), хотя не исключено засорение, поскольку диноцисты изучены из образцов шлама. Наиболее древние находки вида обнаружены в средневолжском подъярусе разреза Городищи (верхняя часть а-зоны panderi) (Пещевицкая, 2021). Ранее в том же разрезе данный вид был установлен начиная с а-зоны virgatus (Harding et al., 2011). Spiniferites ramosus (Ehrenberg) Mantell появляется в переходных средне-верхневолжских слоях Дании (Heilmann-Clausen, 1987) и в верхней части средневолжского подъяруса (а-зона nikitini) на Русской плите (Пещевицкая, 2021). На севере Урала он обнаружен в верхневолжских отложениях (Лебедева, Никитенко, 1998). С берриаса (а-зона jacobi) вид присутствует во Франции (Monteil, 1992). Представители этого рода (вид S. alatus Duxbury) обнаружены также в верхнем титоне Болгарии (Dodekova, 1994). Наиболее древние находки Cassiculosphaeridia reticulata Davey зафиксированы в верхах портланда (а-зона lamplughi) в Нидерландах (Abbink et al., 2001a), в верхневолжском подъярусе Дании (Heilmann-Clausen, 1987) и Сибири (Nikitenko et al., 2018). Batioladinium “varigranosum” (Duxbury) Davey обнаружен в верхах портланда в Североморском регионе (а-зона lamplughi) (Herngreen et al., 2000) и на о. Ньюфаундленд (Van Helden, 1986), а также в верхневолжском подъярусе на Баренцевоморском шельфе (Smelror et al., 1998) и в Сибири (Nikitenko et al., 2018). Виды Cribroperidinium undoryense Smith et Harding и C. magnificum Smith et Harding являются характерными таксонами средне- и верхневолжского подъярусов на Русской плите, первый из них появляется в а-зоне virgatus, второй – в а-зоне panderi (Smith, Harding, 2004; Harding et al., 2011).
Границы. Нижняя граница совмещена с подошвой слоя Е2, верхняя – не определена, условно проведена по верхнему уровню опробования.
Стратиграфическое положение. Слои Е2–Е9, основание слоя Е10, а-зоны panderi–catenulatum.
РАСЧЛЕНЕНИЕ РАЗРЕЗА ПО СПОРАМ И ПЫЛЬЦЕ
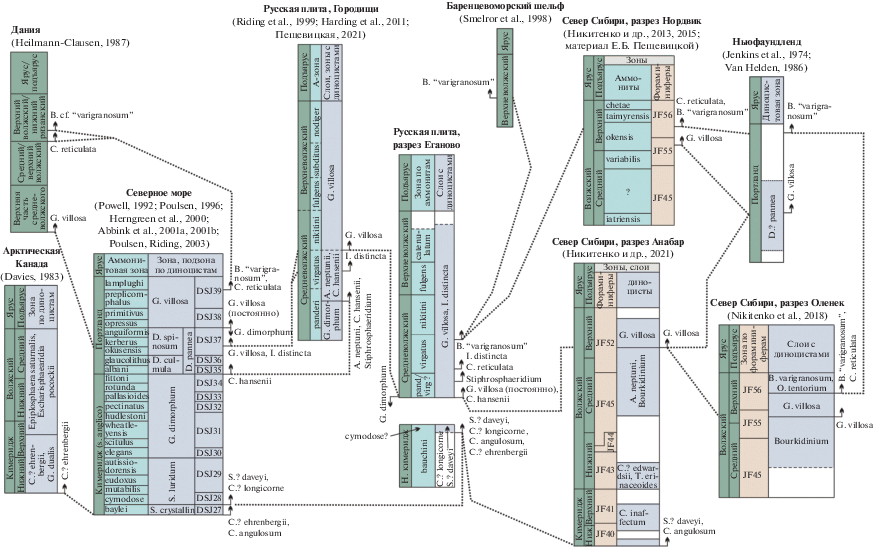
В разрезе Еганово представительные комплексы спор и пыльцы изучены только из волжской части, в нижнем кимеридже наземные палиноморфы встречались редко (рис. 3). В волжском ярусе выявлены слои с Gleicheniidites toriconcavus, Plicatella chetaensis, которые ранее были установлены на Русской плите в разрезе Городищи в средне- и верхневолжском подъярусах (средняя часть а-зоны panderi–catenulatum) (Пещевицкая, 2021).
Слои с Gleicheniidites toriconcavus, Plicatella chetaensis
Характерный комплекс. Среди спор доминируют глейхениевые папоротники родов Gleicheniidites (15–32%) и Plicifera (1–7%). Достаточно многочисленны циатейные папоротники группы Cyathidites–Leiotriletes (1–8.5%), иногда схизейные (до 6%). Споры осмундовых папоротников встречаются в небольшом количестве (1–3%). Разнообразны сфагновые мхи Stereisporites (1–3%). Плауны представлены родами Lycopodiumsporites, Selaginella, Uvaesporites, Neoraistrickia, Densoisporites и др. (рис. 3). В пыльцевой части комплекса наиболее многочисленны и разнообразны хвойные, представленные сосновыми, кипарисовыми, сциадопитисовыми, подокарповыми и обильными хейролипидиевыми (Classopollis, 17–48%). Постоянно присутствует пыльца гинкговых/цикадовых (0.5–3%). Стратиграфически важные виды, присутствующие в комплексе, перечислены ниже.
Стратиграфически важные черты. В небольшом количестве присутствуют ребристые и бугорчатые споры схизейных папоротников родов Trilobosporites и Cicatricosisporites, что характерно для портланда на территории северных областей Западной Европы (Jansonius, McGregor, 1996 и др.). Эти морфотипы также отмечаются в волжском ярусе на севере Урала и в Сибири, но встречаются там значительно реже (Ильина, 1985; Дзюба и др., 2018; Nikitenko et al., 2018 и др.). В разрезе Еганово определен вид Cicatricosisporites sibiricus (Kara-Mursa) Chlonova, который появляется в средневолжском подъярусе (середина а-зоны panderi) на Русской плите в разрезе Городищи (Пещевицкая, 2021) и в верхневолжском подъярусе на севере Урала (Дзюба и др., 2018). Cicatricosisporites australiensis (Cookson) Potonie является ключевым видом для верхнего титона Австралии, где по его появлению установлена одноименная зона, охватывающая также берриас и низы валанжина (Sajjadi, Playford, 2002). В основании этой зоны появляется Foraminisporis dailyi (Cookson et Dettmann) Dettmann, который определен и в разрезе Еганово. Важной чертой изученного палинокомплекса является присутствие вида Gleicheniidites toriconcavus Krutzsch, наиболее древние находки которого отмечаются с верхов верхней юры в Германии (Döring, 1965) и в средней части волжского яруса (середина а-зоны panderi) на Русской плите (Добруцкая, 1973; Пещевицкая, 2021).
Границы. Нижняя граница совмещена с подошвой слоя Е2, верхняя – не определена, условно проведена по верхнему уровню опробования.
Стратиграфическое положение. Слои Е2–Е9, основание слоя Е10, а-зоны panderi–catenulatum.
КОРРЕЛЯЦИОННЫЕ ВОЗМОЖНОСТИ ДИНОЦИСТОВЫХ БИОСТРАТОНОВ
В изученном разрезе определены представительные комплексы спор и пыльцы наземных растений и микрофитопланктона, что позволило провести биостратиграфическое расчленение по двум палинологическим группам. Благодаря детальным фаунистическим датировкам (Рогов, 2017), установленные палиностратоны имеют привязку к аммонитовым зонам. Надежное возрастное обоснование по палинологическим данным базируется на биостратиграфическом анализе спор и пыльцы наземных растений и диноцист, проведенном авторами с использованием литературных данных.
Данные по диноцистам. Среди диноцист в нижней части разреза присутствуют стратиграфически важные виды Cribroperidinium? ehrenbergii и C. angulosum, которые позволяют проводить корреляцию вмещающих их отложений с нижним кимериджем Североморского региона, Арктической Канады и Сибири (рис. 4). Это хорошо согласуется с аммонитовыми датировками, которые показывают наличие в разрезе Еганово нижнекимериджской а-зоны bauhini (Рогов, 2017). В то же время в комплексе присутствуют C.? longicorne и Systematophora? daveyi, которые появляются на севере Западной Европы в а-зоне cymodoce (Powell, 1992; Poulsen, Riding, 2003). Это позволяет предположить, что в верхней части кимериджского интервала в разрезе Еганово могут быть представлены и более высокие стратиграфические горизонты.
Ранее в разрезах Костромской области и Среднего Поволжья для нижнего кимериджа и нижней части верхнего кимериджа (зоны baylei–eudoxus) была установлена диноцистовая зона Gonyaulacysta jurassica subsp. jurassica (Riding et al., 1999). Однако в кимеридже разреза Еганово отсутствуют виды, маркирующие ее нижнюю границу: Scriniodinium anceps (Raynaud) Jan du Chene et al., Glossodinium dimorphum Ioannides et al., Perisseiasphaeridium pannosum Davey et Williams, а также значительно отличается качественный и количественный состав комплексов диноцист, что осложняет сопоставление диноцистовых последовательностей. В этой связи в кимеридже разреза Еганово выделены слои с Cribroperidinium? longicorne, Systematophora? daveyi.
В волжской части разреза имеются некоторые расхождения в биостратиграфической интерпретации палеонтологического материала по аммонитам и диноцистам. По данным М.А. Рогова (2017), находки аммонитов позволяют восстановить в нижней части этого интервала полную зональную последовательность средневолжского подъяруса. В то же время особенности таксономического состава комплексов диноцист более характерны для верхов средневолжского подъяруса и верхневолжского подъяруса. В нижней части волжского интервала, в слоях 2 и 3, вместе с Cribroperidinium hansenii, Stiphrosphaeridium dictyophorum и S. antophorum, которые являются ключевыми таксонами для низов портланда и средней части средневолжского подъяруса, определены Isthmocystis distincta и Gochteodinia villosa, которые ранее не были обнаружены ниже а-зоны nikitini на Русской плите (Harding et al., 2011; Пещевицкая, 2021), а-зоны okensis на севере Сибири (Nikitenko et al., 2018) и а-зоны kerberus на севере Западной Европы (Powell, 1992; Poulsen, Riding, 2003). Интересно также отметить присутствие проксимохоратных диноцист Dinophyceae gen. indet. sp. A (табл. I ). Наличие апикального археопиля и коронообразной септы (иногда двух), которые расположены антапикально и/или в нижней части гипоцисты, сближает эти формы с родами Stephanelytron (Sarjeant, 1961) и Lagenadinium (Piel, 1985). Такие же морфотипы наблюдались в средне- и верхневолжском подъярусах (а-зоны nikitini, fulgens) в разрезе Городищи (данные Е.Б. Пещевицкой).
Таблица I . Палиноморфы из нижнего кимериджа и верхней части волжского яруса разреза Еганово. Длина масштабной линейки 20 мкм. 1 – Senoniasphaera jurassica (Gitmez et Sarjeant) Lentin et Williams, сл. Е2; 2 – Circulodinium distinctum (Deflandre et Cookson) Jansonius, сл. Е3; 3 – Mendicodinium groenlandicum (Pocock et Sarjeant) Davey, сл. Е2; 4 – Perisseiasphaeridium insolutum Davey, сл. Е3; 5 – Glossodinium dimorphum Ioannides et al., сл. Е2; 6 – Cribroperidinium? longicorne (Downie) Lentin et Williams, сл. Е1; 7 – Cribroperidinium hansenii Poulsen, сл. Е10; 8 – Surculosphaeridium sp. I Davey 1982, сл. Е8; 9, 10 – Gochteodinia villosa (Vozzhennikova) Norris: 9 – сл. Е2, 10 – сл. Е4; 11 – Batioladinium matyjae Poulsen, сл. Е5; 12 – Tubotuberella apatela (Cookson et Eisenack) Ioannides et al., сл. Е5; 13 – Oligosphaeridium sp., сл. Е10; 14 – Cingutriletes clavus (Balme) Dettmann, сл. Е7; 15 – Egmontodinium toryna (Cookson et Eisenack) Davey, сл. Е4; 16 – Cassiculosphaeridia reticulata Davey, сл. Е3; 17 – Scriniodinium crystallinum (Deflandre) Klement, сл. Е2; 18 – Kaiwaradinium sp., сл. Е10; 19, 20, 25 – Dinophyceae gen. indet. sp. A: 19 – сл. Е4; 20 – сл. Е3; 25 – сл. Е6; 21 – Uvaesporites cerebralis Tralau, сл. Е2; 22 – Gleicheniidites toriconcavus Krutzsch, сл. Е3; 23 – Hystrichodinium pulchrum Deflandre, сл. Е4; 24 – Dichadogonyaulax culmula Neale et Sarjeant, сл. Е6; 26 – Podocarpidites multesimus (Bolkhovitina) Pocock, сл. Е6; 27 – Sestrosporites pseudoalveolatus (Couper) Dettmann, сл. Е3.

Вид G. villosa на севере Западной Европы является важным биостратиграфическим и корреляционным маркером. Одноименная зона (=Pareodinia dasyforma, сейчас младший синоним G. villosa) была установлена в разрезах Англии по появлению и постоянному присутствию вида-индекса и охватывала верхнюю часть портланда (с а-зоны opressus) и почти полностью рязанский ярус (до а-зоны albidum) (Woollam, Riding, 1983; Powell, 1992 и др.). Примерно в том же объеме (основание верхневолжского подъяруса–нижняя часть рязанского яруса) эта диноцистовая зона была установлена на Русской плите (Riding et al., 1999). В дальнейшем редкие находки G. villosa были обнаружены в более низких стратиграфических горизонтах (а-зона kerberus на севере Западной Европы и а-зона nikitini на Русской плите), что позволило пересмотреть положение основания диноцистовой зоны в последнем регионе (Poulsen, Riding, 2003; Harding et al., 2011; Пещевицкая, 2021). На севере Западной Европы нижняя граница зоны осталась на прежнем рубеже, но в дополнение к постоянным находкам вида-индекса была обоснована исчезновением Dichadogonyaulax? pannea (Norris) Sarjeant, Glossodinium dimorphum Ioannides et al. и Dingodinium tuberosum Cookson et Eisenack (Poulsen, Riding, 2003). Dichadogonyaulax? pannea в разрезе Еганово отсутствует, а D. tuberosum обнаружен только в кимеридже. Однако G. dimorphum встречен в слое Е2 на уровне а-зоны panderi. С этого же уровня присутствует вид G. villosa, который встречается и выше, причем постоянно и не единично (1–1.5%) (рис. 2). Выше в изученном разрезе определены Cassiculosphaeridia reticulata и Batioladinium “varigranosum”, характерные для верхов портланда на севере Западной Европы (Нидерланды, Дания) и верхневолжского подъяруса на Баренцевоморском шельфе и в Сибири (Heilmann-Clausen, 1987; Smelror et al., 1998; Nikitenko et al., 2018 и др.). Таким образом, наблюдается определенная последовательность появления ключевых таксонов диноцист, маркирующих стратиграфические рубежи (рис. 4).
Полученные палинологические данные в большей степени согласуются с представлениями С.А. Ширшова (2005) о стратиграфическом диапазоне волжской части разреза в Раменском районе: глауконитовые пески с фосфоритами в начале волжской части разреза рассматривались как средневолжские, а вышележащий глинисто-алевритистый слой как переходный средне-верхневолжский. Расхождения с данными М.А. Рогова (2017) могут быть связаны с различной биостратиграфической интерпретацией палеонтологического материала по аммонитам разными специалистами как в разрезах Русской плиты, так и в других регионах. Находки аммонитов зоны panderi из средневолжских глауконитовых песков с уплотненным прослоем фосфоритов в кровле описывались из Московской синеклизы и прежде (Герасимов и др., 1995 и др.). Однако, по мнению А.Г. Олферьева (2012), это является результатом переотложения фосфоритовых конкреций с аммонитами из более древних отложений, в то время как пески, содержащие эти конкреции, могут относиться к более высоким горизонтам, не древнее зоны virgatus. Такая ситуация возможна и в разрезе Еганово. С другой стороны, находки в нижней части разреза Еганово видов диноцист, характерных на севере Западной Европы и в Сибири для более высоких стратиграфических горизонтов, могут быть связаны с более ранним возникновением этих видов на территории европейской части России и их последующим расселением в соседние регионы.
Таким образом, в разрезе Еганово хорошо опознаются характерные признаки зоны Gochteodinia villosa. Однако если учитывать точку зрения М.А. Рогова, то обнаруживается нетипично раннее появление зонального вида на уровне а-зоны panderi. Это существенно ниже, чем на севере Западной Европы (а-зона kerberus; Poulsen, Riding, 2003) и в Среднем Поволжье (а-зона nikitini; Harding et al., 2011; Пещевицкая, 2021). В результате, как рабочий вариант, в разрезе Еганово установлены слои с Gochteodinia villosa, Isthmocystis distincta (рис. 2).
Данные по спорам и пыльце. Представительные комплексы спор и пыльцы наземных растений изучены в разрезе Еганово только из волжской части. В нижнем кимеридже наземные палиноморфы встречаются редко (рис. 3). Отсутствуют ребристые и бугорчатые споры схизейных папоротников, которые рассматриваются как стратиграфически важные признаки спорово-пыльцевых комплексов верхнего оксфорда и кимериджа на севере Западной Европы и в Сибири (Jansonius, McGregor, 1996; Никитенко и др., 2015 и др.). Отметим, что данный интервал на европейской территории России не был охарактеризован этими морфотипами и ранее (Шрамкова, 1970; Добруцкая, 1973 и др.). Это может быть обусловлено как аридным климатом (Вахрамеев, 1988), не благоприятным для произрастания папоротников, так и слабой насыщенностью образцов наземными палиноморфами на фоне доминирующего микрофитопланктона (Добруцкая, 1973; Селькова и др., 2011 и др.).
Однако в разрезе Еганово интересно отметить присутствие Selaginella granata Bolchovitina. На севере Средней Сибири его наиболее древние находки отмечаются с середины нижнего кимериджа (средняя часть а-зоны kitchini), что позволяет проводить корреляцию с изученным разрезом. Это также подтверждает возможность присутствия в кровле слоя Е1 в разрезе Еганово уровня, соответствующего а-зоне cymodoce севера Западной Европы. Вид S. granata также отмечен в кимеридж-нижневолжских спорово-пыльцевых комплексах в центральных и северных районах Западной Сибири (Глушко, Пуртова, 1980). На территории Русской плиты слои с S. granata установлены в верхах кимериджа и в нижней части волжского яруса в разрезе Городищи (Пещевицкая, 2021). Тем не менее на юго-западе Украины и в Средней Азии редкие находки этого вида зафиксированы в бате (Яновская, 1973; Юрские…, 1981). Возможно, более раннее появление плаунов вида S. granata здесь связано с палеофлористической зональностью: в средней юре сибирские территории входили в состав одноименной палеофлористической области с теплоумеренным климатом, а Украина и Средняя Азия входили в состав Евро-Синийской палеофлористической области с более жарким субтропическим климатом (Вахрамеев, 1988). Расселение вида на север в поздней юре могло быть связано с общим климатическим потеплением (Вахрамеев, 1988) и возникновением в Сибири благоприятных условий для его произрастания, в то время как более южные районы были подвержены аридизации и мало подходили для развития плаунов. Территория, где находится разрез Еганово, в юре располагалась на севере Евро-Синийской палеофлористической области, и вид S. granata мог появиться здесь позже, чем в ее южных районах. Однако до сих пор его присутствие в спорово-пыльцевых комплексах средней и поздней юры как в центральных, так и в южных областях Русской платформы до настоящего времени не было установлено (Шрамкова, 1970; Добруцкая, 1973; Ростовцева, 2014 и др.). Таким образом, значение этого вида для палиностратиграфии верхней части юры здесь, возможно, будет уточнено с появлением новых данных.
В волжской части разреза Еганово по спорам и пыльце наземных растений выявлены слои с Gleicheniidites toriconcavus, Plicatella chetaensis, которые были установлены на Русской плите по материалам из разреза Городищи в верхней части волжского яруса (средняя часть а-зоны panderi–catenulatum; Пещевицкая, 2021). В разрезе Городищи была хорошо палинологически охарактеризована нижняя часть слоев, а в верхней части наземные палиноморфы встречались редко. Получить детальную характеристику верхней части палиностратона позволяют материалы из разреза Еганово (рис. 3).
С точки зрения палиностратиграфии здесь важно присутствие вида Gleicheniidites toriconcavus, наиболее древние находки которого отмечаются в верхах верхней юры в Германии (Döring, 1965), а также в средневолжском подъярусе в Костромском Заволжье (Добруцкая, 1973) и разрезе Городищи (Пещевицкая, 2021). Характерно присутствие в небольшом количестве ребристых и бугорчатых спор схизейных папоротников родов Trilobosporites и Cicatricosispotites, что также отмечается для портланда Западной Европы (Jansonius, McGregor, 1996 и др.) и верхнего титона Крыма (Доротяк и др., 2009). Эти морфотипы наблюдаются и в волжском ярусе на севере Урала и в Сибири, но встречаются там значительно реже (Ильина, 1985; Дзюба и др., 2018; Nikitenko et al., 2018 и др.). На территории Средней Азии схизейные встречаются единично, в титонских комплексах спор и пыльцы здесь резко доминирует пыльца Classopollis (95–99%) (Споры…, 1971).
Важно отметить присутствие в разрезе Еганово видов Cicatricosisporites sibiricus, C. ludbrookiae и C. australiensis (Cookson) Potonie (рис. 5). Первый их них появляется в средневолжском подъярусе (середина а-зоны panderi) на Русской плите в разрезе Городищи (Пещевицкая, 2021) и в верхневолжском подъярусе на севере Урала (Дзюба и др., 2018). Второй, наряду с морфологически близким видом C. abacus Burger, является стратиграфическим маркером верхов кимериджа (sensu anglico) и портланда на севере Западной Европы. В разрезах Англии, Нидерландов и шельфа Северного моря эти виды появляются в интервале а-зоны albani и постоянно встречаются в портланде (Burger, 1966; Hunt, 1985; Abbink, 1998; Herngreen et al., 2000). В Австралии C. ludbrookiae появляется в верхнем титоне (Sajjadi, Playford, 2002). Здесь же отмечаются наиболее древние находки C. australiensis. Этот вид является ключевым для верхнего титона Австралии. По его появлению установлена одноименная зона, охватывающая также берриас и низы валанжина (Sajjadi, Playford, 2002). Отметим, что в основании этой зоны появляется Foraminisporis dailyi, который определен в разрезе Еганово на близком стратиграфическом уровне. Споры, близкие по морфологии к C. australiensis, определены также в верхневолжском подъярусе на севере Урала (Дзюба и др., 2018). Таким образом, перечисленные выше виды можно рассматривать как характерные для верхней части волжского яруса Европейской России и севера Урала и стратиграфических аналогов на севере Западной Европы и в Австралии. Несмотря на то, что их появление в конкретных разрезах может наблюдаться на разных уровнях, нижний диапазон стратиграфического распространения ограничен для большинства видов а-зонами panderi/albani, для C. australiensis – а-зоной virgatus, для Foraminisporis dailyi – а-зоной fulgens (рис. 5). Отметим, что на севере Сибири эти виды появляются позднее, в бореальном берриасе/рязанском ярусе и/или раннем валанжине (Пещевицкая, 2010 и др.), что может быть обусловлено флористической зональностью.
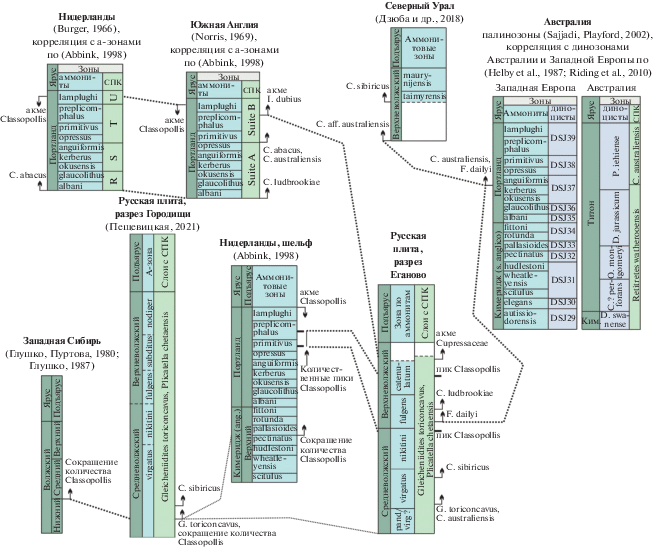
Рис. 5.
Корреляция биостратиграфических маркирующих уровней по спорам и пыльце. СПК – спорово-пыльцевой комплекс.
Характерной чертой слоев с Gleicheniidites toriconcavus, Plicatella chetaensis является также уменьшение количества пыльцы Classopollis (с 40–60 до 12–34%) и увеличение процентного содержания спор глейхениевых папоротников (с 17–38 до 23–50%) по сравнению с нижележащими горизонтами, что было прослежено в разрезе Городищи (Пещевицкая, 2021). Сходное соотношение этих палиноморф наблюдается и в средневолжском подъярусе в разрезе Еганово: Classopollis – 18–38%, Gleicheniaceae – 22–35%. При сопоставлении с более южными районами четкой корреляции не наблюдается. На территории Днепрово-Донецкой впадины количество Classopollis сокращается в зоне virgatus (до 18%, по сравнению с 50% и более в зоне panderi) (Воронова, Яновская, 1973). Однако, по материалам Г.В. Шрамковой (1970), в Воронежской антеклизе эта пыльца обильна в обеих зонах (panderi – 50–90%, virgatus – 83–85%).
В разрезе Еганово количество Classopollis увеличивается в верхней части слоев с Gleicheniidites toriconcavus, Plicatella chetaensis (до 48%). Сходные закономерности подмечены Н.А. Добруцкой (1969) в Костромском Заволжье в разрезах, охарактеризованных аммонитами: в средней части волжского яруса количество этой пыльцы составляло 15–30%, а в верхневолжском подъярусе – 40–70%. Повышенное содержание пыльцы Classopollis обнаружено в верхневолжском подъярусе и в других разрезах в Московской области (Кунцево и Мильково, а-зоны fulgens, catenulatum; данные А.В. Лидской), а также в Рязанской области (а-зона catenulatum) (Федорова, Грязева, 1984). Материалы по разрезу Еганово показывают, что увеличение содержания пыльцы Classopollis происходит не плавно, а скачкообразно: пики отмечаются в верхах а-зоны nikitini–низах fulgens и в верхах а-зоны catenulatum.
Обилие пыльцы Classopollis (50–80%) характерно и для портланда Западной Европы (Burger, 1966; Norris, 1969 и др.). По О. Аббинку (Abbink, 1998), этот род является главным компонентом группы теплолюбивых растений побережья. Количественные показатели этой группы несколько уменьшаются, начиная с а-зоны pallasioides (с 50–60 до 40%), но интересно отметить максимумы в а-зоне primitivus и средней части preplicomphalus (Abbink, 1998). Согласно корреляционной модели М.А. Рогова (2019), это хорошо сопоставляется с сокращением обилия Classopollis в разрезе Городищи в слоях с Gleicheniidites toriconcavus, Plicatella chetaensis и количественными пиками Classopollis в их верхней части в разрезе Еганово (рис. 5). Также хорошо коррелируется увеличение количества Cupressaceae и морфологически близкого типа, определенного Дж. Норрисом (Norris, 1969) как Inaperturopollenites dubius (Potonie et Venitz) Thomson et Pflug, в верхах волжского яруса и портланда (рис. 5).
В Западной Сибири большое количество пыльцы Classopollis характерно для нижневолжского подъяруса южных и центральных областей, где оно может достигать 70–80% (Глушко, Пуртова, 1980; Шейко, 1980). В средневолжском подъярусе процентное содержание Classopollis здесь также сокращается (Глушко, Пуртова, 1980; Глушко, 1987). На севере Средней Сибири в волжском ярусе закономерных изменений в количестве пыльцы Classopollis не установлено, поскольку здесь оно очень мало и редко превышает 2% (Павлов, 1970; Григорьева, 1981; Nikitenko et al., 2018 и др.). В волжских отложениях Вилюйской синеклизы эта пыльца встречается единично (Фрадкина, 1967).
Таким образом, при наличии биостратиграфических маркеров (палинологических и по другим группам биофоссилий), которые позволяют уверенно опознавать среднюю и верхнюю части волжского яруса, количественная динамика пыльцы Classopollis может служить хорошим инструментом для детальной корреляции как местных, так и удаленных разрезов (рис. 5).
АНАЛИЗ ПАЛИНОЛОГИЧЕСКИХ АССОЦИАЦИЙ В ЗАВИСИМОСТИ ОТ ФАЦИАЛЬНЫХ, КЛИМАТИЧЕСКИХ И ПАЛЕОГЕОГРАФИЧЕСКИХ ОСОБЕННОСТЕЙ
Биофациальный анализ палинологического материала подтверждает, что в Московской области в раннем кимеридже и второй половине волжского века существовали нормально-морские условия. Об этом свидетельствует большое количество микрофитопланктона, среди которого стабильно доминируют диноцисты (рис. 6). Во многих образцах присутствуют немногочисленные пресноводные водоросли Botryococcus, которые, видимо, поступали в морские осадки с суши вместе с другими палиноморфами наземного генезиса.
Рис. 6.
Количественное соотношение некоторых экологических групп морских и наземных палиноморф в разрезе Еганово и фациальные особенности. Среди теплолюбивых компонентов в составе группы “другие” рассматриваются Dictyophyllidites, Matonisporites, Tripartina, Plicifera, Klukisporites, Cupressaceae.
Наиболее глубоководные фации характерны для нижнего кимериджа, где в составе диноцист значительный процент принадлежит хоратным формам (с длинными радиальными выростами) и представителям подсемейства Leptodinioideae. Обилие этих форм часто рассматривается как показатель глубины (Wilpshaar, Leereveld, 1994; Leereveld, 1995; Nikitenko et al., 2018 и др.). В небольшом количестве присутствуют диноцисты с выростами, дистально соединенными трабекулами (Adnatosphaeridium, 5.5%). Такие морфотипы характерны для внешней части неритовой зоны и более глубоких областей (Pross, Brinkhuis, 2005; Sluijsa et al., 2005). Однако большое количество Systematophora (14%) и Impletosphaeridium (16%), которые широко развиты и в мелководных зонах (Wilpshaar, Leereveld, 1994; Пещевицкая, 2021 и др.), не позволяет предполагать здесь существование значительных глубин.
В волжском веке возрастает обилие диноцист, которые являются показателями мелководных и прибрежных обстановок (рис. 6). Среди них наиболее многочисленны представители групп Escharisphaeridia и Pilosidinium, способные переносить нестабильные обстановки (Jarvis et al., 1988; Никитенко и др., 2015 и др.). Частая смена условий, очевидно, была связана как с трансгрессивно-регрессивными процессами, так и с изменением гидродинамики, о чем также свидетельствуют перерывы в осадконакоплении и разнообразие литологических разностей в разрезе. Несмотря на осложненность разреза перерывами, общий регрессивный тренд с небольшими флуктуациями хорошо прослеживается вверх по разрезу по увеличению доли спор и пыльцы наземных растений и сокращению диноцист среди микрофитопланктона. Однако динамика разнообразия диноцист имеет обратную тенденцию: количество родов увеличивается вверх по разрезу, что в данном случае, видимо, связано не с трансгрессивно-регрессивными событиями, а с иными факторами.
Состав спорово-пыльцевых ассоциаций свидетельствует о теплом субтропическом климате и довольно влажных условиях. Многочисленной становится группа теплолюбивых таксонов, среди которых наиболее обильна пыльца Classopollis, достигающая максимального развития (до 80–100%) в областях с жарким и аридным климатом (Alvin, 1982; Ильина, 1985; Вахрамеев, 1988 и др.). Другие теплолюбивые компоненты в разрезе Еганово менее многочисленны (рис. 6). Периоды незначительного потепления восстанавливаются в конце средневолжского–начале поздневолжского времени и в фазу catenulatum. Тем не менее следует отметить, что по сравнению с кимериджем и началом волжского века на Русской плите отмечается постепенное похолодание, что в спорово-пыльцевых ассоциациях выражено в заметном сокращении содержания Classopollis (см. выше).
Предполагают, что хвойные растения семейства хейролепидиевых, которые продуцировали пыльцу Classopollis, были адаптированы к широкому спектру обстановок: могли образовывать редколесье на склонах и возвышенностях, заросли кустарников на побережьях, участвовать в мангровой растительности (Alvin, 1982; Ильина, 1985; Вахрамеев, 1988; Пещевицкая, 2021 и др.). Многие из хейролепидиевых хорошо переносили засуху. Однако материалы по разрезу Еганово показывают, что на территории Московской области во второй половине волжского века засушливые периоды если и существовали, то не были продолжительными, так как наряду с пыльцой Classopollis во всех образцах в большом количестве содержатся споры глейхениевых, которые предпочитают умеренно влажные условия. Среди папоротников они наиболее адаптированы к сокращению влаги и на открытых пространствах могут образовывать монодоминантные заросли типа саванн или прерий (Жизнь…, 1978; Van Konijnenburg-Van Cittert, 2002 и др.). Возможно, такой тип растительности был широко развит в фазу virgatus, о чем свидетельствует стабильно высокий процент спор глейхениевых (29–37%). Тем не менее эти папоротники могли также участвовать в растительности болотистых низин (Болховитина, 1968; Бугдаева и др., 2006). Такие местообитания, которые были также благоприятны для плаунов и циатейных/диптерисовых папоротников, могли образовываться вблизи морского палеобассейна или пресноводных водоемов. Существование последних подтверждается присутствием в составе микрофитопланктона зеленых водорослей Botryococcus, которые сегодня являются типичными обитателями озер и болот (Jansonius, McGregor, 1996 и др.).
Обилие глейхениевых папоротников может быть также связано с их способностью легко осваивать новые территории, которые могли появляться в результате колебаний уровня моря. Современные глейхениевые папоротники благодаря быстрому росту хорошо укореняются даже на голой земле, сильно прогреваемой солнцем в тропиках и субтропиках (Жизнь…, 1978).
Некоторое сокращение площади травянистых болот можно предположить в начале фазы nikitini и в фазы fulgens–catenulatum по увеличению роли древесной растительности, что в спорово-пыльцевых спектрах отражается в повышении процентного содержания мешковой пыльцы хвойных (рис. 6). Интересно отметить, что в это время также возрастает разнообразие диноцист. Возможно, это связано с бόльшим количеством минеральных питательных веществ, поступавших с менее заболоченной суши. Современные исследования показывают, что воды с их пониженным содержанием часто образуются вблизи болотистых областей благодаря активному выеданию питательных веществ различными организмами (Жизнь…, 1978). Наибольшего разнообразия диноцисты достигают в периоды, когда увеличение количества питательных веществ сопровождается потеплением (рис. 6). Таким образом, разнообразие диноцист в данном случае в большой степени контролируется количеством питательных веществ и температурным фактором. Это объясняет, почему динамика разнообразия диноцист в разрезе Еганово не совпадает с трансгрессивно-регрессивными флуктуациями.
Отметим, что увеличение разнообразия диноцист в фазы nikitini и fulgens также наблюдалось в мелководных фациях в разрезе Городищи, где это дополнительно сопровождалось большим разнообразием хоратных форм, что считается нетипичным для подобных обстановок. Для объяснения этих фактов было предложено рассмотреть более теплые условия как результат лучшего прогрева воды на мелководье (Пещевицкая, 2021). Данные по разрезу Еганово показывают, что более благоприятные температурные условия в фазы nikitini и fulgens на Русской плите могли быть связаны с климатическим потеплением. Интересно отметить, что анализ изотопов кислорода, углерода и состава глинистых минералов также подтверждает, что небольшое потепление на Русской плите начинается с фазы nikitini и становится более выраженным в поздневолжское время (Ruffell et al., 2002; Gröcke et al., 2003). На этот же временной интервал падают количественные максимумы теплолюбивых компонентов в спорово-пыльцевых ассоциациях на севере Западной Европы (Burger, 1966; Abbink, 1998; Herngreen et al., 2000 и др.). Небольшие температурные пики наблюдаются в поздневолжское время на территории Восточной Сибири и на севере Урала (Nikitenko et al., 2018; Дзюба и др., 2018).
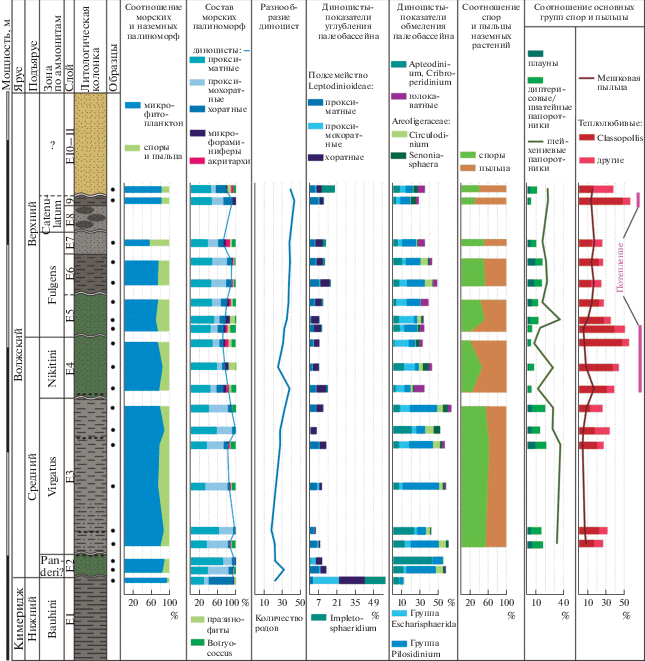
Таким образом, анализ динамики таксономического разнообразия диноцист в разрезе Еганово показывает, что в кимеридже оно значительно ниже, чем в волжском ярусе. Низкое родовое разнообразие также характерно для кимериджа Гренландии и севера Норвегии (10–15 родов) (рис. 7). Обеднение родового состава в Баренцевоморском регионе отмечается для кимериджских сообществ фораминифер, что связывают с более глубоководными обстановками, которые менее продуктивны, чем средняя часть шельфа (Никитенко, 2009). Однако небольшое разнообразие диноцист также наблюдается на юге Западной Европы, в Печорском бассейне и Западной Сибири (20–23 рода), хотя палеообстановки в этих районах значительно различаются (Riding et al., 1999; Ilyina et al., 2005; Borges et al., 2011). В Болгарии, Североморском регионе, на севере Урала и Восточной Сибири оно значительно выше (более 30 родов) (рис. 7). В данном случае нельзя сказать, что наблюдается однозначная зависимость разнообразия диноцист от палеогеографического фактора или глубины палеобассейна (рис. 8, 9). Вероятно, как и в районе Еганово, разнообразие диноцистовых ассоциаций является результатом действия различных факторов и во многом контролируется поступлением питательных веществ с континента, которое может быть ограничено в результате климатических условий, как на юге Западной Европы, где восстанавливаются аридные условия (Hallam, 1984; Abbink, 1998 и др.), или в результате меньшего влияния речного стока, как в центральных областях Западносибирского палеобассейна (Ilyina et al., 2005). Ассоциация кимериджских диноцист из разреза Еганово показывает значительное сходство с одновозрастными ассоциациями из большинства бореальных и суббореальных районов (особенно из Североморского региона), а также из Болгарии, что может свидетельствовать о хороших связях этих акваторий через Датско-Польский и Каспийский проливы (рис. 8, 9).
Рис. 7.
Анализ родового разнообразия диноцист в различных районах на севере Евразии. Условные обозначения: 1 – кимеридж; 2 – средневолжский подъярус, 3 – верхневолжский подъярус и их возрастные аналоги. Номера районов (в скобках) соответствуют таковым на рис. 8а.
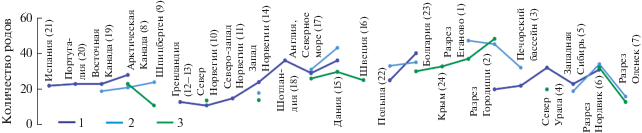
Рис. 8.
Находки кимеридж-волжских палиноморф: диноцист (а) и спор и пыльцы (б). Палеогеографические реконструкции по (Атлас…, 1968; Захаров, 1981; Abbink et al., 2001b; Никитенко, 2009; Rogov et al., 2009; Конторович и др., 2013; Scotese, 2016 и др.); положение палеополюса по (Butler, 2004; Никитенко, 2009 и др.). Положение суши показано для кимериджа, зеленым пунктиром – для волжского века. Рис. 8а. Проливы: У – Уральский, П – Печорский, К – Каспийский, Д – Датско-Польский, Н – Норвежско-Гренландский; районы: 1 – Еганово; 2 – Городищи; 3 – Печорский бассейн; 4 – север Урала; 5 – Западная Сибирь; 6 – разрез Нордвик; 7 – разрез Оленек; 8 – Арктическая Канада; 9 – Шпицберген; 10 – север Норвегии; 11 – северо-запад Норвегии; 12, 13 – Гренландия; 14 – запад Норвегии; 15 – Дания; 16 – Швеция; 17 – Англия, Северное море; 18 – Шотландия; 19 – Восточная Канада; 20 – Португалия; 21 – Испания; 22 – Польша; 23 – Болгария; 24 – Крым. Рис. 8б. Условные обозначения: 1 – граница Сибирской и Евро-Синийской палеогеографических областей по макрофлоре (Вахрамеев, 1988); 2 – граница этих областей в Сибири по палинологическим данным (К – для кимериджа, В – для волжского яруса); 3 – граница распространения северосибирских палинофлор (К – для кимериджа, В – для волжского яруса). Районы: 1 – Португалия; 2 – Испания; 3 – Шотландия; 4 – Северное море; 5 – Нидерланды; 6 – Германия; 7 – Швеция; 8 – Польша; 9 – междуречье Прут–Днестр; 10 – Крым; 11 – междуречье Днепр–Донец; 12 – Воронежская обл.; 13 – Кавказ; 14 – разрез Еганово; 15 – Костромская обл.; 16 – Коми; 17 – разрез Городищи; 18 – Оренбургская обл.; 19 – юг и юго-восток Прикаспийской впадины; 20 – Мангышлак; 21 – Устюрт; 22 – Кызылкум, Бухарская обл.; 23 – Гиссарский хребет; 24 – Павлодарская обл.; 25 – юг Западной Сибири; 26 – юго-запад Западной Сибири; 27 – Курганская обл.; 28 – восточный склон Урала, запад Западной Сибири; 29 – средняя часть Западной Сибири; 30 – Северный Урал, северо-запад Западной Сибири; 31 – Пурпейско-Сургутский район; 32 – Надым-Тазовский район; 33 – Туруханский район; 34 – Усть-Енисейский район; 35 – Норильский район; 36 – р. Хета; 37 – разрез Нордвик; 38 – разрез Анабар; 39 – разрез Оленек; 40 – Жиганский район; 41 – Вилюйская синеклиза; 42 – Иркутская обл.; 43 – Забайкалье; 44 – Буреинская впадина; 45 – Арктическая Канада.
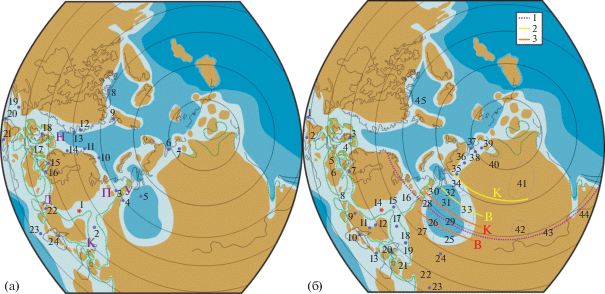
Рис. 9.
Диаграмма сходства ассоциаций диноцист в разрезе Еганово и различных районах на севере Евразии (по коэффициенту Жаккара). Условные обозначения: 1 – кимеридж; 2 – средневолжский подъярус, 3 – верхневолжский подъярус и их возрастные аналоги. Номера районов (в скобках) соответствуют таковым на рис. 8а. Данные по (Bujak, Williams, 1977; Davies, 1983; Van Erve et al., 1988; Van der Zwan, 1990; Wierzbowski, Aarhus, 1990; Powel, 1992; Dodekova, 1994; Poulsen, 1996; Riding et al., 1999; Wierzbowski et al., 2002; Piasecki et al., 2004; Ilyina et al., 2005; Borges et al., 2011; Lindström, Erlström, 2011; Берриас…, 2012; Никитенко и др., 2015; Dalseg et al., 2016; Nikitenko et al., 2018; Lebedeva et al., 2019; Пещевицкая, 2021 и др.).
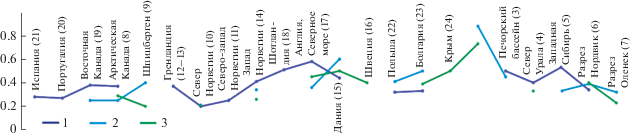
В волжском веке разнообразие диноцист на территории Русской плиты значительно возрастает (рис. 7). В остальных регионах наблюдаются менее значимые изменения, в основном в сторону сокращения разнообразия, что более выражено на севере Урала и, видимо, связано с более мелководными условиями по сравнению с кимериджем (Лебедева, Никитенко, 1998). Латеральные различия в разнообразии диноцистовых ассоциаций становятся более яркими. Сходство родового состава диноцист из разреза Еганово с составом диноцист в большинстве районов к концу волжского века постепенно сокращается, исключение составляют Дания, Болгария и, в меньшей степени, север Восточной Сибири (рис. 9).
Таксономический состав морской фауны (аммониты, двустворки, белемниты, фораминиферы) также показывает, что поздневолжские сообщества в целом менее разнообразны и более дифференцированы по регионам, что хорошо проявлено как в Сибири, так и на Русской плите (Захаров и др., 1994; Никитенко, 2009; Рогов, 2012; Дзюба, 2018 и др.). Это связано с обмелением и сокращением проливов, соединяющих как Арктические территории с южной Палеоатлантикой и Русским морем, так и Русское море с западными и южными акваториями (рис. 8). На территории Русской плиты сокращение возможностей для расселения морской фауны приводит в поздневолжское время к резкому обеднению таксономического состава в сообществах аммонитов, которые представлены 2–3 родами (Mitta, Sha, 2009; Рогов, 2012). Тем не менее обмеление и периодическое закрытие Датско-Польского, Печорского и Каспийского проливов не произвели столь существенного влияния на ассоциации диноцист Русского моря. Как уже было отмечено, их разнообразие даже увеличилось, что связано с температурным эффектом и благоприятными трофическими условиями. Коэффициент таксономического сходства с другими районами уменьшается незначительно, а в некоторых случаях даже возрастает (рис. 9). Вероятно, крайне мелководные условия в проливах, которые препятствовали миграции морской фауны, не были лимитирующим фактором для распространения диноцист. Возможно, это связано с жизненным циклом динофлагеллат, включающим стадию цист, что помогало этим организмам переживать неблагоприятные периоды и преодолевать экологические барьеры, которые для расселения морской фауны оказались препятствием и привели в поздневолжское время к существенному сокращению связей между фаунистическими сообществами разных регионов и, в конечном счете, к таксономическому обеднению. Следует, однако, отметить, что некоторые параметры местных палеообстановок иногда компенсировали отрицательное влияние палеогеографического фактора и для фауны. Так, повышение температуры в конце волжского века, которое способствовало образованию богатых ассоциаций диноцист в Русском море (разрезы Еганово, Городищи), оказалось также благоприятным для развития белемнитов на севере Урала, сообщества которых здесь по разнообразию сравнимы с кимериджскими (Дзюба, 2018).
Состав спорово-пыльцевых ассоциаций из разреза Еганово с обилием пыльцы Classopollis показывает, что этот район в поздней юре находился в пределах Евро-Синийской палеогеографической области с субтропическим жарким и аридным климатом, которая была выделена В.А. Вахрамеевым (1988) на основе палеоботанических данных. Для нее характерно большое количество беннеттитов, хейролепидиевых, семенных папоротников и араукариевых. Это хорошо согласуется с палинологическими данными (табл. 1, рис. 8): в южных областях Западной и Восточной Европы, на Кавказе, в Крыму, Средней Азии, на юге Урала и Сибири в спорово-пыльцевых ассоциациях кимериджа отмечается обилие пыльцы хейролепидиевых (Classopollis) и мешковой пыльцы (Disaccites), которую могли продуцировать как хвойные растения, так и семенные папоротники. Для Западной Европы характерна пыльца Callialasporites, которую некоторые исследователи относят к араукариевым (Abbink, 1998 и др.). Более влажные прибрежные и островные обстановки Шотландии, Португалии и севера Североморского региона отличаются обилием спор циатейных и глейхениевых папоротников и плаунов (табл. 1). В позднем кимеридже (sensu anglico) и портланде доминирование пыльцы Classopollis во многих районах Западной Европы становится особенно резким. Предполагается, что это было вызвано усилением аридизации, возможно, вследствие регрессии морского палеобассейна (Abbink, 1998; Herngreen et al., 2000 и др.). Жаркий и аридный климат наиболее ярко проявился в Средней Азии, что подтверждается составом палеофлор и литологическими особенностями (Вахрамеев, 1988 и др.). В спорово-пыльцевых комплексах резко доминирует пыльца Classopollis (табл. 1, 2).
Таблица 1.
Количественное соотношение основных компонентов кимериджских спорово-пыльцевых комплексов на севере Евразии
| Район | Резко преобладают, >60% | Преобладают, >40% | Обильны, 20–40% | Много, 10–20% | Значительно, 5–10% | Мало, 2–5% |
|---|---|---|---|---|---|---|
| Шотландия (3) | Cyatheaceae, Gleicheniaceae, Disaccites, Callialasporites, Sciadopityaceae, Classopollis | |||||
| Северное море (4) | Lycopodiaceae 30–50% | Cyatheaceae, Osmundaceae: 25–35% | Antulsporites (Bryophytes) | Ginkgoaceae | Disaccites | |
| Нидерланды (5) | Cyatheaceae, Classopollis, Disaccites, Callialasporites, иногда Peromonolites | иногда: Sciadopityaceae, Cyatheaceae, Callialasporites | иногда Gleicheniaceae | |||
| Португалия (1) | Disaccites, Callialasporites | Classopollis, Perinopollenites | ||||
| Испания (2) | Classopollis, Spheripollenites | |||||
| Междуречье Прут–Днестр (9) | Classopollis до 100% | |||||
| Междуречье Днепр–Донец (11) | Classopollis до 100% | |||||
| Разрез Городищи (17) | Classopollis 40–60% | Gleicheniaceae | ||||
| Разрез Еганово (14) | Микрофито-планктон 93% | Споры, пыльца единично | ||||
| Костромская обл. (15), Коми (16) | Микрофитоп-ланктон >90% | Споры, пыльца единично | ||||
| Мангышлак (20) | Classopollis 62–95% | Cyatheaceae | Disaccites, Osmundaceae, Gleicheniaceae | |||
| Павлодарская обл. (24) | Classopollis 54–96% | Disaccites, Cyatheaceae | Osmundaceae | |||
| Курганская обл. (27), ЮЗ Западной Сибири (26) | Classopollis | Disaccites | Cyatheaceae, Osmundaceae, Ginkgoaceae | |||
| Юг Западной Сибири (25), юг ее средней части (29) | Classopollis 70–90% | Disaccites, Cyatheaceae, Osmundaceae, Ginkgoaceae | ||||
| Север средней части Западной Сибири (29) | Classopollis 40–50% | Disaccites, Cyatheaceae | Osmundaceae, Ginkgoaceae | |||
| Восточный склон Урала, запад Западной Сибири (28) | Classopollis, Disaccites | Cyatheaceae, Sciadopityaceae, Gleicheniaceae | Osmundaceae, Ginkgoaceae, Lycopodiaceae | |||
| Пурпейско-Сургутский р-н (31) | Classopollis, Disaccites | Cyatheacaea, Ginkgoaceae | Osmundaceae, Lycopodiaceae | |||
| Северный Урал, СЗ Западной Сибири (30) | Cyatheaceae, Disaccites, Classopollis, Gleicheniaceae | Sciadopityaceae | Ginkgoaceae, Lycopodiaceae | |||
| Надым-Тазовский р-н (32) | Disaccites | Cyatheceae 20–35%, Classopollis 11–35% | Ginkgoaceae, Lycopodiaceae | |||
| Туруханский (33) и Усть-Енисейский (34) р-ны | Cyatheaceae, Disaccites | Classopollis | Ginkgoaceae, Lycopodiaceae, Gleicheniaceae | |||
| Норильский р-н (35) | Cyatheaceae, Disaccites | Ginkgoaceae | Classopollis, Osmundaceae | |||
| Разрез Нордвик (37) | Cyatheaceae 30–40% | Disaccites до 15% | Osmundaceae | Gleicheniaceae | ||
| Разрез Анабар (38) | Disaccites 67–75% | Osmundaceae, Cyatheaceae | Ginkgoaceae, Lycopodiaceae | |||
| Жиганский р-н (40) | Disaccites | Cyatheaceae, Osmundaceae, Ginkgoaceae | Lycopodiaceae, Sphagnaceae | |||
| Вилюйская синеклиза (42) | Cyatheaceae, Disaccites | Osmundaceae | Lycopodiaceae, Sphagnaceae, Ginkgoaceae | |||
| Иркутская обл. (43), Забайкалье (44) |
Classopollis, Disaccites | Cyatheaceae, Lycopodiaceae, Gleicheniaceae, Sphagnaceae, Ginkgoaceae | ||||
| Арктическая Канада (45) | Disaccites, Cyatheaceae | Classopollis | Lycopodiaceae, Sphagnaceae, Ginkgoaceae | |||
Примечание. Номера районов (в скобках) соответствуют таковым на рис. 8б. Данные взяты из (Фрадкина, 1967; Шрамкова, 1970; Котова, 1971; Пономаренко и др., 1971; Споры…, 1971; Стратиграфо-палеонтологическая…, 1972; Воронова, Яновская, 1973; Добруцкая, 1973; Шейко, 1980; Григорьева, 1981; Глушко, 1987; Смирнова, 1983; Van Erve et al., 1988; Van Konijnenburg-Van Cittert, Van der Burgh 1996; Herngreen et al., 2000; Скобло и др., 2001; Borges et al., 2011; Lindström, Erlström, 2011; Galloway et al., 2013; Маркевич, Бугдаева, 2014; McArthur et al., 2016; Дзюба и др., 2018 и др.).
Обилие пыльцы Classopollis и спор глейхениевых характерно для европейской части России. Хотя морские кимериджские осадки здесь редко содержат представительные комплексы спор и пыльцы (что наблюдается и в разрезе Еганово), доминирование Classopollis прослежено в разрезе Городищи. Однако ее количество здесь меньше (40–60%), чем в Западной Европе, на Украине и юге России (55–100%). Это, возможно, связано с более северным положением данного района в пределах Евро-Синийской области и более влажными условиями, что подтверждается обилием спор глейхениевых папоротников (20–40%) (табл. 1). Эта особенность (отчетливое доминирование пыльцы Classopollis и спор глейхениевых) хорошо прослеживается в волжском веке по материалам из разреза Еганово, а также по данным из Среднего Поволжья и Костромской области (табл. 2). Палинологические показатели более влажного климата хорошо согласуются с повышенным содержанием углерода, которое обычно связывают с гумидификацией, в волжских отложениях в разрезах Среднего Поволжья (Ruffell et al., 2002 и др.). Отметим, что количество Classopollis в волжских палинокомплексах непостоянно и варьирует в бόльших пределах по сравнению с кимериджем, что связано с климатическими колебаниями и изменением температурных условий. В настоящей статье это продемонстрировано на примере разреза Еганово. Однако обилие спор папоротников показывает, что, в отличие от Западной Европы, усиления аридизации здесь не было.
Таблица 2.
Количественное соотношение основных компонентов спорово-пыльцевых комплексов для волжского века и его возрастных аналогов на севере Евразии
| Район | Резко преобладают, >60% | Преобладают, 40–60% | Обильны, 20–40% | Много, 10–20% | Значительно, 5–10% | Мало, 2–5% | |
|---|---|---|---|---|---|---|---|
| Северное море (4) | Lycopodiaceae 50–60% | Cyatheaceae, Antulsporites (Bryophytes) | Disaccites | ||||
| Нидерланды (5) | Classopollis 50–95% | Disaccites, иногда Peromonolites | Callialasporites, Spheripollenites, иногда Cyatheaceae | Eucommiidites, Cupressaceae | |||
| Германия (6) | Classopollis 40–80% | Disaccites, Sciadopityaceae, Cupressaceae, иногда Peromonolites | Gleicheniaceae, Schizaeaceae, Cyatheaceae | ||||
| Польша (8) | Classopollis 50–80% | Cupressaceae 10–25%, Disaccites 10–12% | Gleicheniaceae (редко 15%), Cyatheaceae | ||||
| Швеция (7) | Disaccites | Cyatheaceae, Gleicheniaceae, Sciadopityaceae | Classopollis | ||||
| Междуречье Прут–Днестр (9) | Classopollis 70–80% | Cyatheaceae | Gleicheniaceae, Schizaeaceae, Disaccites | ||||
| Воронежская обл. (12) | Classopollis 50–95% | Gleicheniaceae, порой Cupressaceae | Ginkgoaceae | Disaccites, Cyatheaceae | |||
| Междуречье Днепр–Донец (11) | Classopollis более 50% (зона panderi) | Gleicheniaceae, Classopollis до 18% (зона virgatus) | Cyatheaceae, Disaccites | ||||
| Разрез Еганово (14) | Редко Classopollis 40–48% | Gleicheniaceae 22–35%, Classopollis 18–38% | Cyatheaceae, Disaccites, иногда Eucommiidites | ||||
| Разрез Городищи (17) | Редко Gleicheniaceae 46–50% | Gleicheniaceae 23–38, Classopollis 20–34% | Иногда Classopollis 12–20% | Cyatheaceae | Disaccites | ||
| Костромская обл. (15) | Редко Classopollis 40–70% | Classopollis 15–30%, Gleicheniaceae 18–20% | Podozamitaceae | Ginkgoaceae | Disaccites, Cyatheaceae | ||
| Оренбургская обл. (18) | Classopollis 50-60% | Podozamitaceae | Gleicheniaceae, Cyatheaceae | Schizaeaceae | |||
| Крым (10) | Classopollis 70–98% | Marratisporites, Schizaeaceae, Disaccites, Cyatheaceae, Cupressaceae, Ginkgoaceae, Eucommiidites, Callialasporites | |||||
| Северный Кавказ (13) | Classopollis 52–90% | Disaccites, Ginkgoaceae, Inaperturopollenites, Callialasporites | |||||
| Ю-ЮВ Прикаспийской впадины (19) | Classopollis 42–60% | Disaccites | Cyatheaceae, Cupressaceae, Disaccites, Ginkgoaceae, Inaperturopollenites, Callialasporites | ||||
| Мангышлак (20) | Classopollis 70–75% | Disaccites, Cupressaceae, Gleicheniaceae, Cyatheaceae | |||||
| Устюрт (21) | Classopollis 94–100% | Disaccites, Gleicheniacea, Cyatheaceae | |||||
| Кызылкум, Бухарская обл. (22) | Classopollis 72–76% | Disaccites, Ginkgoaceae, Inaperturopollenites, Gleicheniaceae, Cyatheaceae | |||||
| Гиссарский хр. (23) | Classopollis 98–99% | ||||||
| Павлодарская обл. (24) | Classopollis 54–96% | Disaccites, Cyatheaceae | |||||
| Курганская обл. (27), Ю и ЮЗ Западной Сибири (26 и 25) | Classopollis | Disaccites | Cyatheaceae, Ginkgoaceae | ||||
| Средняя часть Западной Сибири (29) | Classopollis (редко на Ю до 45%), Cyatheaceae, Disaccites | Gleicheniaceae | Osmundaceae, Ginkgoaceae | ||||
| Восточный склон Урала, запад Западной Сибири (28) | Classopollis, Disaccites, Cyatheaceae, Sciadopityaceae | Gleicheniaceae, Lycopodiaceae, Osmundaceae, Ginkgoaceae | |||||
| Пурпейско- Сургутский р-н (31) | Disaccites | Cyatheacaea, Ginkgoaceae, Classopollis | Gleicheniaceae, Osmundaceae | ||||
| Северный Урал, СЗ Западной Сибири (30) | Редко до 50% Disaccites, Gleicheniaceae | Gleicheniaceae, Disaccites или Cyatheaceae | Classopollis, Lycopodiaceae, Osmundaceae, Ginkgoaceae, Sciadopityaceae | ||||
| Надым-Тазовский р-н (32) |
Disaccites, Cyatheaceae | Classopollis, Gleicheniaceae (редко до 16%) | Lycopodiaceae, Osmundaceae, Ginkgoaceae | ||||
| Усть-Енисейский (34) и Туруханский (33) р-ны | Disaccites | Cyatheaceae | Osmundaceae, Ginkgoaceae, Classopollis (на Ю до 20%), Gleicheniaceae (редко до 16%) | ||||
| р. Хета (36) | Disaccites до 35% | Cyatheaceae до 25% | Lycopodiaceae | Osmundaceae | |||
| Разрез Нордвик (37) | Disaccites 30–60% | Cyatheaceae | Osmundaceae | Lycopodiaceae, Sphagnaceae | |||
| Разрез Анабар (38) | Disaccites 43–74% | Cyatheaceae 13–26%, Osmundaceae 7–16% | Ginkgoaceae, Lycopodiaceae | ||||
| Разрез Оленек (39) | Disaccites (40–70%) | Cyatheaceae | Osmundaceae, Ginkgoaceae, Sphagnaceae | ||||
| Жиганский р-н (40) | Disaccites, Cyatheaceae | Osmundaceae, Ginkgoaceae | Gleicheniaceae | ||||
| Вилюйская синеклиза (41) | Disaccites до 66%, Cyatheaceae до 46% | Osmundaceae (иногда меньше) | Ginkgoaceae, Gleicheniaceae | ||||
| Иркутская обл. (42), Забайкалье (43) |
Disaccites | Cyatheaceae, Lycopodiaceae, Gleicheniaceae, Sphagnaceae, Ginkgoaceae, Classopollis | |||||
| Буреинская впадина (44) | Disaccites до 56%, Cyatheaceae до 79% | Lycopodiaceae, Osmundaceae, Ginkgoaceae, Sphagnaceae | Classopollis | ||||
| Арктическая Канада (45) | Disaccites, Cyatheaceae | Classopollis | Lycopodiaceae, Sphagnaceae, Ginkgoaceae | ||||
Примечание. Номера районов (в скобках) соответствуют таковым на рис. 8б. Данные взяты из (Фрадкина, 1967; Шрамкова, 1970; Котова, 1971; Пономаренко и др., 1971; Споры…, 1971; Стратиграфо-палеонтологическая…, 1972; Воронова, Яновская, 1973; Добруцкая, 1973; Шейко, 1980; Григорьева, 1981; Глушко, 1987; Смирнова, 1983; Van Erve et al., 1988; Van Konijnenburg-Van Cittert, Van der Burgh 1996; Herngreen et al., 2000; Скобло и др., 2001; Borges et al., 2011; Lindström, Erlström, 2011; Galloway et al., 2013; Маркевич, Бугдаева, 2014; McArthur et al., 2016; Дзюба и др., 2018 и др.).
Обилие пыльцы Classopollis и спор глейхениевых характерно и для других территорий, расположенных на севере Евро-Синийской области (Среднего и Северного Урала, северо-запада Западной Сибири) и рассматриваемых некоторыми исследователями как экотонные (Ровнина, 1972; Вахрамеев, 1988). В кимеридже здесь сохраняется доминирование пыльцы Classopollis, хотя ее количество постепенно сокращается в северном направлении. Это позволило Л.В. Ровниной (1972) выделить Уральскую провинцию в составе Евро-Синийской палеогеографической области. Характерной особенностью этих палинофлор также является обилие пыльцы сциадопитисовых (табл. 1, 2). Разнообразие макроостатков Sciadopitys отмечается для макрофлор севера Норвегии, которые В.А. Вахрамеев (1988 и др.) также рассматривает как экотонные между Евро-Синийской и Сибирской палеогеографическими областями, но относит к последней. На востоке Евразийского континента экотонные палеофлоры обнаружены в Забайкалье и Приморье (Вахрамеев, 1988 и др.). В спорово-пыльцевых спектрах в Иркутской области и Забайкалье доминирует мешковая пыльца хвойных и Classopollis (Скобло и др., 2001; Башурова, 2005 и др.). В Западной Сибири граница палеофлористических областей в кимеридже проходит примерно в районе широтного течения р. Обь (Шейко, 1980; Вахрамеев, 1988 и др.): к югу резко доминирует пыльца Classopollis, к северу к доминантам добавляются циатейные, глейхениевые (на западе) и мешковая пыльца хвойных (табл. 1, рис. 8). К северосибирским палинофлорам близки палинофлоры Арктической Канады (табл. 1, рис. 8).
В волжском веке граница палеофлористических областей, видимо, смещается несколько к северу. Резкое доминирование Classopollis сохраняется в европейской части России, а на территории Западной Сибири – только в южных районах (табл. 2, рис. 8). Уральская провинция рассматривается уже в составе Сибирской палеогеографической области (Ровнина, 1972). Вероятно, это связано с началом климатического тренда похолодания, которое тем не менее не было устойчивым, что подтверждают материалы из разреза Еганово: в волжских палиноспектрах наблюдается значительное сокращение количества Classopollis по сравнению с кимериджем (разрез Городищи), но оно происходило не плавно, а скачкообразно: отмечаются количественные пики, отражающие периоды потепления (рис. 6). Как уже отмечалось, количественные флуктуации Classopollis для этого стратиграфического интервала также фиксируются на территории Западной Европы, севера Урала, Западной Сибири (Глушко, Пуртова, 1980; Abbink, 1998; Дзюба, 2018 и др.) и могут служить дополнительными корреляционными маркерами.
По палинологическим данным похолодание в волжском веке прослеживается и для северных областей Сибирской палеофлористической области. В кимеридже большое количество пыльцы Classopollis (до 22%, редко больше) сохраняется на севере Западной Сибири вплоть до Усть-Енисейского района (табл. 1, рис. 8). На северо-востоке этого региона, в Лено-Анабарском районе и Вилюйской синеклизе, преобладает мешковая пыльца хвойных и споры циатейных папоротников, количество Classopollis мало (0–5%), но увеличивается доля спор осмундовых (5–20%) (табл. 1, рис. 8). В волжском веке северосибирские палинофлоры распространялись в Усть-Енисейский, Туруханский и Ныдинский районы на территории Западной Сибири, в Забайкалье и Приморье (табл. 2, рис. 8).
ВЫВОДЫ
Впервые изучены комплексы спор, пыльцы и диноцист в разрезе Еганово (Московская обл.). Биостратиграфический анализ палинологического материала позволил провести расчленение кимеридж-волжского интервала по морским и наземным палиноморфам. Значительное количество стратиграфически значимых таксонов обеспечивает надежное палинологическое обоснование выделенных биостратонов и подтверждает присутствие в разрезе нижнего кимериджа и верхней части волжского яруса, установленных ранее по аммонитам. Хорошие корреляционные возможности слоев с диноцистами наблюдаются как для кимериджа, так и для волжского яруса, позволяя сопоставлять разрезы европейской части России, Западной Европы, Баренцевоморского шельфа и Сибири. По причине слабой насыщенности наземными палиноморфами нижней, кимериджской, части разреза Еганово провести полноценный биостратиграфический анализ по этой группе здесь не удалось. Однако выше по разрезу были определены обильные и разнообразные спорово-пыльцевые спектры и выявлены слои с Gleicheniidites toriconcavus, Plicatella chetaensis, ранее установленные в верхней части волжского яруса в разрезе Городищи. Материалы из разреза Еганово позволили детально охарактеризовать верхнюю часть этого палиностратона, которая в разрезе Городищи была изучена недостаточно. Благодаря характерному палинокомплексу и ключевым таксонам, этот палиностратон хорошо опознается на территории европейской части России. Ряд видов позволяет проводить корреляцию с одновозрастными отложениями севера Урала, Западной Европы и Австралии. Показано также, что на севере Евразии дополнительным инструментом для детальной корреляции в верхней части волжского яруса может служить количественная динамика пыльцы Classopollis, которая отражает климатические колебания.
Результаты биофациального анализа микрофитопланктона свидетельствуют о том, что на территории Московской области в кимеридже и волжском веке существовали нормально-морские условия. Более глубоководные фации характерны для раннего кимериджа. В волжском веке палеобассейн мелеет, вверх по разрезу наблюдается регрессивный тренд с постоянными флуктуациями и частой сменой параметров среды. В ассоциациях микрофитопланктона обильны диноцисты, способные переносить нестабильные обстановки. Наибольшего разнообразия диноцисты достигают в фазу catenulatum. Предполагается, что это связано с благоприятными трофическими условиями и кратковременным климатическим потеплением, что реконструируется на основе таксономической и количественной динамики в ассоциациях наземных палиноморф. Период незначительного потепления восстанавливается также в конце средневолжского времени. Сравнительный анализ спорово-пыльцевых ассоциаций из северных областей Евразии показал, что небольшие квазисинхронные климатические максимумы и минимумы также прослеживаются в Сибири, на севере Западной Европы и Урала.
В целом спорово-пыльцевые ассоциации позволяют говорить о теплом субтропическом климате и довольно влажных условиях. По особенностям таксономического состава эти ассоциации характерны для Евро-Синийской палеогеографической области, выделенной В.А. Вахрамеевым по палеоботаническим данным. Количественная динамика основных компонентов палинокомплексов в конце юрского периода, которая прослежена на территории Северной Евразии по материалам авторов и литературным данным, указывает на более жаркие условия в кимеридже и постепенное похолодание в волжском веке. Спорово-пыльцевые ассоциации с обилием теплолюбивых компонентов были распространены в кимеридже на обширной территории в южных областях Евразии. На севере Евро-Синийская палеогеографическая область захватывала южные области Западной и Восточной Сибири. В волжском веке границы распространения евро-синийских спорово-пыльцевых ассоциаций с обилием пыльцы Classopollis и северосибирских с доминированием мешковой пыльцы хвойных и повышенным количеством осмундовых смещаются в более низкие широты. Территория европейской части России остается в пределах Евро-Синийской палеогеографической области, но в палинологических спектрах значительно сокращается доля пыльцы Classopollis, среди доминантов добавляются споры глейхениевых, что хорошо демонстрируют материалы из разреза Еганово. На севере Западной Европы для временного интервала, синхронного волжскому веку, восстанавливается период аридизации климата, однако в европейской части России он не проявляется. На это указывают как палинологические материалы, так и опубликованные изотопные данные. Довольно влажные условия способствовали развитию на этой территории, на севере Урала и северо-западе Западной Сибири специфической растительности с обилием глейхениевых папоротников, которые могли легко осваивать новые участки суши, возникавшие в результате регрессий, и, возможно, образовывали монодоминантные заросли подобные тем, которые сегодня существуют в саваннах или прериях.
Морские альгофлоры Московской области, изученные из разреза Еганово, проявляют значительное сходство с альгофлорами большинства бореальных и суббореальных районов. В кимеридже морская альгофлора изучаемого района имела наибольшее сходство с альгофлорой Североморского региона и Болгарии, что может свидетельствовать о хороших связях между этими акваториями через Датско-Польский и Каспийский проливы. К концу волжского века сходство диноцистовых ассоциаций постепенно сокращается, что, вероятно, связано с постепенным обмелением и сокращением проливов, соединяющих Русское море с соседними палеобассейнами. Однако анализ динамики родового разнообразия диноцист в разрезе Еганово показывает, что в волжском веке оно значительно выше, чем в кимеридже. Однозначной зависимости разнообразия диноцист от палеогеографического фактора или глубины палеобассейна не установлено. Предполагается, что разнообразие диноцистовых ассоциаций является результатом сочетания различных факторов и во многом контролируется поступлением питательных веществ с континента, а также температурными условиями. В разрезе Еганово это продемонстрировано на основе совместного анализа морских и наземных палиноморф. В результате благоприятного сочетания различных факторов периодическое закрытие Датско-Польского, Печорского и Каспийского проливов в поздневолжское время не произвели на ассоциации диноцист Русского моря столь существенного влияния, как на фаунистические сообщества.
Благодарности. Авторы выражают благодарность докт. геол.-мин. наук. М.А. Рогову (ГИН, Москва) за помощь во время полевых работ, а также ценные консультации. Авторы также благодарны рецензентам Н.В. Ильиной, О.В. Шурековой, А.Г. Федяевскому, В.В. Аркадьеву за внимательное отношение к статье, важные замечания и научную дискуссию.
Источники финансирования. Работа выполнена при поддержке гранта РФФИ 20-05-00076, А.В. Лидской исследование выполнено в рамках гос. задания ГИН РАН.
Список литературы
Атлас литолого-палеографических карт СССР. Гл. ред. Виноградов А.П. М.: ГУГК, 1968. Т. 3. 77 л.
Башурова Н.Ф. Обоснование возраста континентальных мезозойских образований нерчинской серии Восточного Забайкалья // Палинология: теория и практика. Материалы XI Всероссийской палинологической конференции. М.: ПИН РАН, 2005. С. 23–24.
Берриас Горного Крыма. СПб.: Лема, 2012. 472 с.
Болховитина Н.А. Споры глейхениевых папоротников и их стратиграфическое значение. М.: Наука, 1968. 136 с.
Болховитина Н.А. Сравнительная характеристика палинологических комплексов поздней юры Русской платформы и Англии // Палинология мезофита. Труды III Международной палинологической конференции. Ред. Хлонова А.Ф. М.: Наука, 1973. С. 92–97.
Бугдаева Е.В., Волынец Е.Б., Голозубов В.В., Маркевич В.С., Амельченко Г.Л. Флора и геологические события середины мелового периода (Алчанский бассейн, Приморье). Владивосток: Дальнаука, 2006. 204 с.
Вахрамеев В.А. Юрские и меловые флоры и климаты Земли. М.: Наука, 1988. 215 с.
Воронова М.А., Яновская Г.Г. Сопоставление спорово-пыльцевых комплексов пограничных юрско-меловых образований Днепрово-Донецкой впадины и междуречья Прут–Днестр // Палинология мезофита. Тр. III Международной палинологической конференции. М.: Наука, 1973. С. 101–104.
Герасимов П.А., Митта В.В., Кочанова М.Д. Ископаемые волжского яруса Центральной России. М.: ВНИГНИ, 1995. 115 с.
Глушко Н.К. Палинологическая характеристика верхнеюрских отложений Сидоровского нефтегазоносного района // Биостратиграфия мезозоя Западной Сибири. Сб. научных трудов. Тюмень: ЗапСибНИГНИ, 1987. С. 50–54.
Глушко Н.К., Пуртова С.И. Сопоставление спорово-пыльцевых комплексов из осадков баженовской и тутлеймской свит // Палинологические исследования мезозоя и кайнозоя Западной Сибири. Тюмень: ЗапСибНИГНИ, 1980. С. 30–36.
Григорьева К.Н. Палинологическая характеристика верхнеюрских отложений р. Хеты (Енисей-Хатангский прогиб) // Стратиграфия и палеонтология Сибири. Сб. научных трудов СНИИГМиС. 1981. Вып. 287. С. 62–65.
Дзюба О.С. Бореальные белемниты (Megateuthididae, Cylindroteuthididae) юры и нижнего мела: систематика, биоразнообразие, зональные шкалы, био- и хемостратиграфические маркеры межрегиональной корреляции: Дисс. … докт. геол.-мин. наук. Новосибирск: ИНГГ, 2018. 321 с.
Дзюба О.С., Пещевицкая Е.Б., Урман О.С., Шурыгин Б.Н., Алифиров А.С., Игольников А.Е., Косенко И.Н. Разрез Маурынья как ключевой для приграничных юрско-меловых отложений мелководно-морского генезиса в Западной Сибири // Геология и геофизика. 2018. Т. 59. № 7. С. 1072–1102.
Добруцкая Н.А. Палинологическая характеристика верхнеюрских отложений центральной части Московской синеклизы // Изв. АН СССР. Сер. геол. 1969. № 6. С. 108–115.
Добруцкая Н.А. Спорово-пыльцевые комплексы юрских и меловых отложений северных районов Русской платформы и их значение для стратиграфии и палеогеографии // Палинология мезофита. Тр. III Международной палинологической конференции. М.: Наука, 1973. С. 108–112.
Доротяк Ю.Б., Матвеев А.В., Шевчук Е.А. Xарактеристика пограничных отложений юры и мела в Горном Крыму (фораминиферы, известковый наннопланктон, диноцисты, палинокомплексы) // Збірник наукових праць Інституту геологічних наук НАН України. Викопна фауна і флора України: Палеоекологічний та стратиграфічний аспекти. Київ: НАН України, 2009. С. 108–117.
Жизнь растений. М.: Просвещение, 1978. Т. 1. 487 с.; т. 4. 656 с.
Захаров В.А. Бухииды и биостратиграфия бореальной верхней юры и неокома. М.: Наука, 1981. 271 с.
Захаров В.А., Бейзель А.Л., Богомолов Ю.И., Константинов А.Г., Курушин Н.И., Меледина С.В., Никитенко Б.Л., Шурыгин Б.Н. Этапность и периодичность в эволюции морских экосистем бореального мезозоя // Экосистемные перестройки и эволюция биосферы. М.: Недра, 1994. С. 138–151.
Ильина В.И. Палинология юры Сибири. М.: Наука, 1985. 237 с.
Конторович А.Э., Конторович В.А., Рыжкова С.В., Шурыгин Б.Н., Вакуленко Л.Г., Гайдебурова Е.А., Данилова В.П., Казаненков В.А., Ким Н.С., Костырева Е.А., Москвин В.И., Ян П.А. Палеогеография Западно-Сибирского осадочного бассейна в юрском периоде // Геология и геофизика. 2013. Т. 54. № 8. С. 972–1012.
Котова Л.И. Спорово-пыльцевые комплексы средне-верхнеюрских отложений Мангышлака // Палинология Казахстана. Алма-Ата: Министерство геологии СССР, 1971. С. 34–43.
Лебедева Н.К., Никитенко Б.Л. Микрофитопланктон и микрофораминиферы опорного разреза нижнего мела Приполярного Зауралья (Западная Сибирь) // Геология и геофизика. 1998. Т. 38. № 6. С. 799–821.
Маркевич В.С., Бугдаева Е.В. Позднеюрско-раннемеловые растения-углеобразователи Буреинского бассейна (российский Дальний Восток) // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 3. С. 14–30.
Митта В.В., Вукс В.Я., Глинских Л.А., Дзюба О.С., Захаров В.А., Кириков В.П., Костылева В.В., Маленкина С.Ю., Никитенко Б.Л., Пещевицкая Е.Б., Рогов М.А., Ростовцева Ю.И., Сельцер В.Б., Тесакова Е.М. Унифицированная региональная стратиграфическая схема юрских отложений Восточно-Европейской платформы. Объяснительная записка. М.: ВНИГНИ, 2012. 64 с. Прил. на 14 листах.
Никитенко Б.Л. Стратиграфия, палеобиогеография и биофации юры Сибири по микрофауне (фораминиферы и остракоды). Новосибирск: Параллель, 2009. 680 с.
Никитенко Б.Л., Шурыгин Б.Н., Князев В.Г., Меледина С.В., Дзюба О.С., Лебедева Н.К., Пещевицкая Е.Б., Глинских Л.А., Горячева А.А., Хафаева С.Н. Стратиграфия юры и мела Анабарского района (Арктическая Сибирь, побережье моря Лаптевых) и бореальный зональный стандарт // Геология и геофизика. 2013. Т. 54. № 8. С. 1047–1082.
Никитенко Б.Л., Князев В.Г., Пещевицкая Е.Б., Глинских Л.А. Верхняя юра побережья моря Лаптевых: межрегиональные корреляции и палеообстановки // Геология и геофизика. 2015. Т. 56. № 8. С. 1496–1519.
Никитенко Б.Л., Девятов В.П., Пещевицкая Е.Б., Попов А.Ю., Фурсенко Е.А., Хафаева С.Н. Стратиграфия, литология и геохимия прибрежных и мелководно-морских разрезов верхов средней юры–низов мела р. Анабар (Арктическая Сибирь) // Геология и геофизика (в печати).
Олферьев А.Г. Стратиграфические подразделения юрских отложений Подмосковья // Бюлл. МОИП. Отд. геол. 2012. Т. 87. Вып. 4. С. 32–55.
Орлова Е.Д., Цыбин Ю.И. Средне- и верхнеюрские отложения левобережной части бассейна среднего течения р. Бузулук // Вопросы геологии Южного Урала и Поволжья. Вып. 8. Часть 1. Саратов: Изд-во Саратовского ун-та, 1972. С. 93–102.
Павлов В.В. Палинологическое обоснование границы верхнеюрских и нижнемеловых отложений на мысе Урдюк-Хая (п-ов Пакса, Анабарский залив // Ученые записки НИИГА. Палеонтология и биостратиграфия. 1970. Вып. 29. С. 32–35.
Пещевицкая Е.Б. Диноцисты и палиностратиграфия нижнего мела Сибири. Новосибирск: Гео, 2010. 230 с.
Пещевицкая Е.Б. Палиностратиграфия и палеообстановки в разрезе Городищи (Среднее Поволжье, кимеридж–готерив) // Геология и геофизика. 2021. Т. 62. № 7. С. 939–966.
Пономаренко З.К., Кальменева А.Х., Нестерова Е.В. О возрасте красноцветных отложений Прииртышского района // Палинология Казахстана. Алма-Ата: Министерство геологии СССР, 1971. С. 39–43.
Ровнина Л.В. Стратиграфическое расчленение континентальных отложений триаса и юры запада Западно-Сибирской низменности. М.: Наука, 1972. 77 с.
Рогов М.А. Широтный градиент таксономического разнообразия аммонитов в Северном полушарии в кимериджском и волжском веках // Палеонтол. журн. 2012. № 2. С. 40–48.
Рогов М.А. Аммониты и инфразональная стратиграфия кимериджского и волжского ярусов юга Московской синеклизы // Юрская система России: проблемы стратиграфии и палеогеографии. Научные материалы Седьмого Всероссийского совещания. Ред. Захаров В.А., Рогов М.А., Щепетова Е.В. М.: ГИН РАН, 2017. Вып. 615. С. 60–71.
Рогов М.А. Аммониты и инфразональная стратиграфия кимериджского и волжского ярусов Панбореальной надобласти. Дисс. … докт. геол.-мин. наук. Т. 1 (основная часть). М.: ГИН, 2019. 877 с.
Ростовцева Ю.И. Палинокомплексы из среднеюрских отложений центра европейской части России. Дисс. … канд. геол.-мин. наук. М.: МГУ, 2014. 201 с.
Селькова Л.А., Ветошкина О.С., Лыюров С.В. Результаты изучения разрезов юрских отложений в окрестностях села Визинги (Республика Коми) // Вест. Ин-та геологии Коми НЦ УрО РАН. 2011. № 9. С. 7–13.
Скобло В.М., Люмина Н.А., Руднев А.Ф., Лузина И.В. Континентальный верхний мезозой Прибайкалья и Забайкалья (стратиграфия, условия осадконакопления, корреляция). Новосибирск: Изд-во СО РАН, 2001. С. 332 с.
Смирнова С.Б. Палинокомплексы из пограничных юрско-меловых отложений Крыма, Кавказа и Восточного Прикаспия // Стратиграфия и корреляция осадков методами палинологии. Материалы IV Всесоюзной палинологической конференции. Свердловск: Уральский научный центр, 1983. С. 134–137.
Споры и пыльца юры и раннего мела Средней Азии. М.: Недра, 1971. 176 с.
Стратиграфо-палеонтологическая основа детальной корреляции нефтегазоносных отложений Западно-Сибирской низменности. Тюмень: ЗапСибНИГНИ, 1972. 227 с.
Федорова В.А., Грязева А.С. Палиностратиграфия пограничных отложений юры–мела в разрезах р. Оки // Пограничные ярусы юрской и меловой системы. М.: ИГиГ СО АН СССР, 1984. С. 150–161.
Фрадкина А.Ф. Спорово-пыльцевые комплексы мезозоя Западной Якутии. Л.: Недра, 1967. 124 с.
Шейко Л.Н. Палинокомплексы из васюганской, георгиевской и баженовской свит юго-восточной части Западной Сибири // Палинологические исследования мезозоя и кайнозоя Западной Сибири. Тюмень: ЗапСибНИГНИ, 1980. С. 27–29.
Ширшов С.А. Стекольные пески Чулковского месторождения (Московская область) // Вест. Воронежского ун-та. Геология. 2005. № 1. С. 177–193.
Шрамкова Г.В. Спорово-пыльцевые комплексы юры и нижнего мела Воронежской антеклизы и их стратиграфическое значение. Воронеж: ВГУ, 1970. 103 с.
Юрские континентальные отложения востока Средней Азии (датировка, расчленение, корреляция). М.: Наука, 1981. 188 с.
Яновская Г.Г. Спорово-пыльцевые комплексы юрских отложений междуречья Прут–Днестр и их значение для стратиграфии. Автореф. дис. … канд. геол.-мин. наук. Киев: Ин-т геологии, 1973. 24 с.
Abbink O.A. Palynological investigations in the Jurassic of the North Sea region. Utrecht: LPP Foundation, 1998. 192 p.
Abbink O.A., Callomon J.H., Riding J.B., Williams P.D.B., Wolfard A. Biostratigraphy of Jurassic–Cretaceous boundary strata in the Terschelling Basin, The Netherlands // Proc. Yorkshire Geol. So. 2001a. V. 53. № 4. P. 275–302.
Abbink O., Targarona J., Brinkhuis H., Visscher H. Late Jurassic to earliest Cretaceous palaeoclimatic evolution of the southern North Sea // Glob. Planet. Change. 2001b. V. 30. P. 231–256.
Alvin K.L. Cheirolepidiaceae: biology, structure and palaeoecology // Rev. Palaeobot. Palynol. 1982. V. 37. 71–98.
Borges M.E.N., Riding J.B., Fernandes P., Pereira Z. The Jurassic (Pliensbachian to Kimmeridgian) palynology of the Algarve Basin and the Carrapateira outlier, southern Portugal // Rev. Palaeobot. Palynol. 2011. V. 163. P. 190–204.
Bujak J. P., Williams G. L. Jurassic palynostratigraphy of offshore eastern Canada // Stratigraphic micropalaeontology of Atlantic Basin and Borderlands. Ed. Swain E.W. Amsterdam: Elsevier, 1977. P. 321–339.
Burger D. Palynology of uppermost Jurassic and lowermost Cretaceous strata in the Eastern Netherlands // Leidse Geologische Mededelingen. 1966. V. 35. P. 209–276.
Butler R.F. Paleomagnetism: Magnetic Domains to Geologic Terranes. Portland: Univ. of Portland, 2004. 238 p.
Dalseg T.S., Nakrem H.A., Smelror M. Dinoflagellate cyst biostratigraphy, palynofacies, depositional environment and sequence stratigraphy of the Agardhfjellet Formation (Upper Jurassic–Lower Cretaceous) in central Spitsbergen (Arctic Norway) // Norw. J. Geol. 2016. V. 96. № 2. P. 1–12.
Davies E.H. The dinoflagellate oppel-zonation of the Jurassic–Lower Cretaceous sequence in the Sverdrup Basin, Arctic Canada // Geol. Surv. Can. Bull. 1983. № 359. P. 1–59.
Dodekova L. Dinoflagellate cysts from the Bathonian–Tithonian (Jurassic) of North Bulgaria. III. Taxonomy of Oxfordian and Kimmeridgian dinoflagellate cysts // Geol. Balcan. 1992. V. 22. № 3. P. 33–69.
Dodekova L. Dinoflagellate cysts from the Bathonian–Tithonian (Jurassic) of North Bulgaria. III. Tithonian dinoflagellate cysts // Geol. Balcan. 1994. V. 24. № 5. P. 11–46.
Döring H. Die sporenpalaontologische Gliederung des Wealden in Westmecklenburg (Struktur Werle) // Geologie. 1965. V. 47. P. 1–119.
Galloway J.M., Sweet A.R., Swindles G.T., Dewing K., Hadlari T., Embry A.F., Sanei H. Middle Jurassic to Lower Cretaceous paleoclimate of Sverdrup Basin, Canadian Arctic Archipelago inferred from the palynostratigraphy // Mar. Petrol. Geol. 2013. V. 44. P. 240–255.
Gröcke D.R., Price G.D., Ruffell A.H., Mutterlose J., Baraboshkin E. Isotopic evidence for Late Jurassic–Early Cretaceous climate change // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003. V. 202. P. 97–118.
Hallam A. Continental humid and arid zones during the Jurassic and Cretaceous // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1984. V. 47. P. 19–223.
Harding I.C., Smith G.A., Riding J.B., Wimbledon W.A.P. Inter-regional correlation of Jurassic/Cretaceous boundary strata based on the Tithonian–Valanginian dinoflagellate cyst biostratigraphy of the Volga Basin, western Russia // Rev. Palaeobot. Palynol. 2011. V. 167. P. 82–116.
Heilmann-Clausen C. Lower Cretaceous dinoflagellate biostratigraphy in the Danish Central Trough // Denm. Geol. Unders. 1987. V. 17. P. 1–89.
Helby R., Morgan R., Partridge A.D. A palynological zonation of the Australian Mesozoic // Studies in Australian Mesozoic palynology. Ed. Jell P.A. Sydney: AAP, 1987. P. 1–95.
Herngreen G.F.W., Kerstholt S.J., Munsterman D.K. Callovian-Ryazanian (Upper Jurassic) palynostratigraphy of the Central North Sea Graben and Vlieland Basin, the Netherlands // Mededel. Nederl. Instit. Toegepaste Geowetenschappen. 2000. V. 63. P. 1–97.
Hunt C.O. Miospores from the Portland Stone Formation and the lower part of the Purbeck Formation (Upper Jurassic/Lower Cretaceous) from Dorset, England // Pollen et Spores. 1985. V. 27. № 3. P. 419–451.
Ilyina V.I., Nikitenko B.L., Glinskikh L.A. Foraminifera and dinoflagellate cyst zonation and stratigraphy of the Callovian to Volgian reference section in the Tyumenskaya superdeep well (West Siberia, Russia) // Recent developments in applied biostratigraphy. Micropalaeontol. Soc. Spec. Publ. 2005. P. 109–144.
Jaccard P. Distribution de la flore alpine dans le Bassin des Dranses et dans quelques regions voisines // Bull. Soc. Vaudoise sci. Natur. 1901. V. 37. Bd. 140. P. 241–272.
Jansonius J., McGregor D.C. (Eds.) Palynology: Principles and applications. Salt Lake City: Publishers Press, 1996. V. 1. 462 p.; V. 2. 910 p.
Jarvis I., Carson G.A., Cooper M.K.E., Hart M.B., Leary P.N., Tocher B.A., Horne D., Rosenfeld A. Microfossil assemblages and the Cenomanian–Turonian (Late Cretaceous) oceanic anoxic event // Cretaceous Res. 1988. V. 9. P. 3–103.
Jenkins W.A.M., Ascoli P., Gradstein F.M., Williams G.L. Stratigraphy of the Acomo IOE A-l Puffin B-90 well. Grand Banks of Newfoundland // Geol. Surv. Can. Bull. 1974. № 74–61. P. 1–12.
Lebedeva N.K., Nikitenko B.L., Colpaert C. Dinoflagellate cysts and Foraminifera of the Upper Jurassic Lopsiya River sections, Nether-Polar Urals, NW Western Siberia (Russia) // Rev. Micropaleontol. 2019. V. 64. https://doi.org/10.1016/j.revmic.2019.07.001
Leereveld H. Dinoflagellate cysts from the Lower Cretaceous Rio Argos succession (SE Spain). Utrecht: LPP Foundation, 1995. 175 p.
Lindström S., Erlström M. The Jurassic–Cretaceous transition of the Fårarp-1 core, southern Sweden: sedimentological and phytological indications of climate change // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011. V. 308. P. 445–475.
McArthur A.D., Jolley D.W., Hartley A.J., Archer S.G., Lawre H.M. Palaeoecology of syn-rift topography: a Late Jurassic footwall island on the Josephine Ridge, Central Graben, North Sea // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016. V. 459. P. 63–75.
Mitta V., Sha J.G. Late Jurassic ammonite evolution and paleoenvironment of the Russian Platform // Sci. China. Ser. D, Earth Sci. 2009. V. 52. № 12. P. 2024–2028.
Monteil E. Kystes de dinoflagelles index (Tithonien–Valanginien) du sud-est de la France. Proposition d’une nouvelle zonation palynologique // Rev. Paleobiologie. 1992. № 11. P. 299–306.
Nikitenko B.L., Pestchevitskaya E.B., Khafaeva S.N. High-resolution stratigraphy and palaeoenvironments of the Volgian–Valanginian in the Olenek section (Anabar-Lena region, Arctic Eastern Siberia) // Rev. Micropalaeontol. 2018. V. 61. P. 271–312.
Norris G. Miospores from the Purbeck Beds and marine Upper Jurassic of Southern England // Palaeontology. 1969. V. 12 № 4. P. 574–620.
Piasecki S., Larsen M., Therkelsen J., Vosgerau H. Jurassic dinoflagellate cyst stratigraphy of Hold with Hope, North-East Greenland // Geol. Surv. Denm. Greenl. 2004. Bull. 5. P. 73–88.
Piel K.M. Lagenadinium, a new dinocyst from British middle-late Callovian rocks, a possible evolutionary intermediate between Stephanelytron and Gardodinium // Rev. Palaeobot. Palynol. 1985. V. 45. P. 107–119.
Poulsen N.E. Dinoflagellate cysts from marine Jurassic deposits of Denmark and Poland // AASP Contrib. Ser. 1996. V. 31. P. 1–227.
Poulsen N.E., Riding J.B. The Jurassic dinoflagellate cyst zonation of Subboreal Northwest Europe // Geol. Surv. Denm. Greenl. 2003. Bull. 1. P. 115–144.
Powell A.J. (Ed.) A stratigraphic index of dinoflagellate cysts. London: Chapman and Hall, 1992. 290 p.
Pross J., Brinkhuis H. Organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene; a synopsis of concepts // Paläontologische Zeitschrift. 2005. V. 79. Bull. 1. P. 53–59.
Riding J.B., Federova V.A., Ilyina V.I. Jurassic and lowermost Cretaceous dinoflagellate cyst biostratigraphy of the Russian Platform and northern Siberia, Russia // AAPS Contribution ser. 1999. V. 36. 184 p.
Riding J.B., Mantle D.J., Backhouse J. A review of the chronostratigraphical ages of Middle Triassic to Late Jurassic dinoflagellate cyst biozones of the North West Shelf of Australia // Rev. Palaeobot. Palynol. 2010. V. 162. P. 543–575.
Rogov M., Zakharov V., Kiselev D. Molluscan immigrations via biogeographical ecotone of the Middle Russian Sea during the Jurassic // Volumina Jurassica. 2009. V. VI. P. 143–162.
Ruffell A.H., Price G.D., Mutterlos J., Kessels K., Baraboshkin E., Gröcke D.R. Palaeoenvironmental sensitivity of clay minerals, stable isotopes and calcareous nannofossils: evidence for palaeoclimatic change during the Late Jurassic–Early Cretaceous, Volga Basin, SE Russia // Geol. J. 2002. V. 37. P. 17–33.
Sajjadi F., Playford G. Systematic and stratigraphic palynology of Late Jurassic–earliest Cretaceous strata of the Eromanga Basin, Queensland, Australia, Part two // Palaeontographica. Ab. B. 2002. V. 261. P. 99–165.
Sarjeant W.A.S. Microplankton from the Kellaways Rock and Oxford Clay of Yorkshire // Palaeontology. 1961. V. 4. № 1. P. 90–118.
Sarjeant W.A.S. Middle and Upper Jurassic dinoflagellate cyst // AASP Contrib. Ser. 1979. V. 2. № 5. P. 133–156.
Scotese C.R. PALEOMAP PaleoAtlas for GPlates and the PaleoData Plotter Program. PALEOMAP Project. Technical Report. 2016. https://doi.org/10.13140/RG.2.2.34367.00166
Sluijsa A., Pross J., Brinkhuis H. From greenhouse to icehouse; organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene // Earth-Sci. Rev. 2005. V. 68. P. 281–315.
Smelror M., Mork A., Monteil E., Rutledge D., Leereveld H. The Klippfisk Formation: a new lithostratigraphic unit of Lower Cretaceous platform carbonates on the western Barents Shelf // Polar Res. New Ser. 1998. V. 17. Bull. 2. P. 181–202.
Smith G.A., Harding I.C. New dinoflagellate cyst species from Upper Jurassic to Lower Cretaceous sediments of the Volgian lectostratotype sections at Gorodische and Kashpir, Volga Basin, Russia // Rev. Palaeobot. Palynol. 2004. V. 128. P. 355–379.
Thomas J.E., Cox B.M. The Oxfordian–Kimmeridgian Stage boundary (Upper Jurassic) – Dinoflagellate cyst assemblages from the Harome borehole, North Yorkshire // England Rev. Palaeobot. Palynol. 1988. V. 56. № 3. P. 313–326.
Van der Zwan C.J. Palynostratigraphy and palynofacies reconstruction of the Upper Jurassic to lowermost Cretaceous of the Draugen Field, Offshore Mid Norway // Rev. Palaeobot. Palynol. 1990. V. 62. P. 157–186.
Van Erve A.W., Besems R.E., Love C.F. A palynological investigation of some Lower Kimmeridgian deposits from Spain // J. Micropalaeontol. 1988. V. 7. Iss. 2. P. 17–232.
Van Helden B.G.T. Dinoflagellate cysts at the Jurassic–Cretaceous boundary, offshore Newfoundland, Canada // Palynology. 1986. V. 10. P. 181–199.
Van Konijnenburg-Van Cittert J.H.A. Ecology of some Late Triassic to Early Cretaceous ferns in Eurasia // Rev. Palaeobot. Palynol. 2002. V. 119. P. 113–124.
Van Konijnenburg-Van Cittert J.H.A., Van der Burgh J. Review of the Kimmeridgian flora of Sutherland, Scotland, with reference to the ecology and in situ pollen and spores // Proc. Geol. Assoc. 1996. V. 107. P. 97–105.
Wierzbowski A., Aarhus N. Ammonite and dinoflagellate cyst succession of an Upper Oxfordian–Kimmeridgian black shale core from the Nordkapp Basin, southern Barents Sea // Newsl. Stratigr. 1990. V. 22. № 1. P. 7–19.
Wierzbowski A., Smelror M., Merk A. Ammonites and dinoflagellate cysts in the Upper Oxfordian and Kimmeridgian of the northeastern Norwegian Sea (Nordland VII offshore area): biostratigraphical and biogeographical significance // N. Jb. Geol. Paläont. Abh. 2002. V. 226. Bull. 2. P. 145–164.
Wilpshaar M., Leereveld H. Palaeoenvironmental changes in the Early Cretaceous Vocontian Basin (SE France) reflected by dinoflagellate cysts // Rev. Palaeobot. Palynol. 1994. V. 84. P. 121–128.
Woollam R., Riding J.B. Dinoflagellate cyst zonation of the English Jurassic // Inst. Geol. Sci. Rep. 1983. V. 83. P. 1–41.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция