

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 2, стр. 60-70
О биостратиграфии нижнего–среднего келловея Саратовского Поволжья по микрофауне
Л. А. Глинских 1, *, Е. М. Тесакова 2, 3, В. Б. Сельцер 4
1 Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
2 Геологический факультет Московского государственного университета им. М.В. Ломоносова
Москва, Россия
3 Геологический институт РАН
Москва, Россия
4 Саратовский национальный исследовательский государственный университет
им. Н.Г. Чернышевского
Саратов, Россия
* E-mail: glor@mail.ru
Поступила в редакцию 27.05.2021
После доработки 22.08.2021
Принята к публикации 25.10.2021
- EDN: OJMVNH
- DOI: 10.31857/S0869592X22020028
Аннотация
Приведены результаты микропалеонтологического анализа разреза ТЭЦ-5 (г. Саратов), который является одним из важнейших для стратиграфии нижнего келловея–низов среднего келловея в Поволжье. Установлены систематический состав и распределение в разрезе фораминифер и остракод. Уточнен объем биостратиграфических подразделений по микрофауне, а также проведено их сопоставление со шкалой по аммонитам. В разрезе установлены две фораминиферовые зоны Восточно-Европейской платформы: Haplophragmoides infracalloviensis–Guttulina tatarensis и Lenticulina cultratiformis–Lenticulina pseudocrassa. Обосновано, что стратиграфический объем зоны Lenticulina cultratiformis–Lenticulina pseudocrassa не ограничен средним келловеем, как считалось ранее, а включает также верхнюю часть нижнего келловея (в объеме аммонитовых зон Koenigi (верхняя часть) и Calloviense). По результатам анализа распределения остракод установлено два биостратона: зона Acantocythere milanovskyi–Procytheridea cinicinnusa, сопоставляемая с нижнекелловейской аммонитовой зоной Subpatruus (с нижней частью биогоризонта Ch. saratovensis), и слои с Praeschuleridea wartae–Pleurocythere kurskensis, охватывающие верхи аммонитовой зоны Subpatruus с верхней частью биогоризонта Ch. saratovensis, зоны Koenigi и Calloviense нижнего келловея и нижнюю часть зоны Jason среднего келловея. Приведены изображения характерных форм.
ВВЕДЕНИЕ
Микрофауна келловея Поволжья изучается с 1930-х гг. (Мятлюк, 1939, 1961; Даин, 1948, 1961; Быкова, 1948; Любимова, 1955; Хабарова, 1959, 1961; Тесакова, Сельцер, 2013; Устинова, Тесакова, 2015 и др.). Так, Л.Г. Даин (1948) наметила схему расчленения юрских отложений, в том числе келловея, Саратовской области по фораминиферам, выделила слои с характерными комплексами, которые позднее составили основу зонального расчленения келловея по фораминиферам не только Ульяновско-Саратовского прогиба (Биостратиграфия…, 1982), но и в целом центральных и южных районов Восточно-Европейской платформы (ВЕП) (Практическое…, 1991; Унифицированная…, 1993, 2012). Как известно, в лектостратотипе фораминиферовых зон келловея – в Малиновом овраге у с. Хлебновка Саратовской области – границы фораминиферовых биостратонов совмещены с границами подъярусов и аммонитовых зон (Биостратиграфия…, 1982; Практическое…, 1991). Так, фораминиферовая зона Lenticulina cultratiformis–Lenticulina pseudocrassa, нижняя граница которой устанавливается по появлению зонального комплекса, а верхняя – по исчезновению видов-индексов, рассматривается в объеме среднего келловея. Однако в более поздней публикации по фораминиферам Малинового оврага, в которой авторы использовали новые данные по аммонитам (Репин, Рашван, 1996), нижняя граница зоны Lenticulina cultratiformis–Lenticulina pseudocrassa проводится не на границе нижнего и среднего келловея, а внутри нижнего келловея (Никифорова и др., 2001). Сведения о появлении в нижнем келловее Щигровского свода Воронежской антеклизы зонального комплекса биостратона Lenticulina cultratiformis–Lenticulina pseudocrassa приводились А.Г. Олферьевым с коллегами (1992). К сожалению, эти важные данные не были учтены при уточнении зонального расчленения по фораминиферам в стратиграфических схемах 1993 и 2012 гг. (Унифицированная…, 1993, 2012), где отражены устаревшие представления об объеме зоны L. cultratiformis–L. pseudocrassa. Это, в свою очередь, создает значительные трудности при возрастной интерпретации фораминиферовых комплексов из пограничного интервала нижнего–среднего келловея. Особенно остро эта проблема стоит при работе с керном, где находки аммонитов редки или отсутствуют. Например, в фатежской свите, вскрытой скважинами в Малоархангельском (Орловская обл.) и Фатежском (Курская обл.) районах, совместно с раннекелловейскими остракодами (Tesakova, 2013) был обнаружен комплекс фораминифер (включая виды-индексы), характерный для “среднекелловейской” зоны L. cultratiformis–L. pseudocrassa (Устинова, 2017). Стратиграфический конфликт, вызванный устаревшими представлениями об объеме зоны в (Унифицированная…, 2012), не позволил М.А. Устиновой выделить эту зону в изученных скважинах, а для расчленения отложений этих скважин использовались комплексы фораминифер, предложенные в публикации А.Г. Олферьева с коллегами (1992), и на этом основании отложения отнесли к нижнему келловею.
Таким образом, назрела необходимость уточнения зонального расчленения по микрофауне, а именно изменения стратиграфического объема и положения нижней границы зоны по фораминиферам Lenticulina cultratiformis–Lenticulina pseudocrassa и ее сопоставления с остракодовыми биостратонами.
ФАКТИЧЕСКИЙ МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
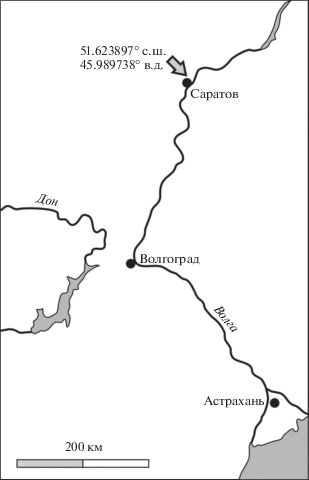
Материалом для микропалеонтологического исследования послужила коллекция фораминифер и остракод из 24 образцов, отобранных авторами из келловейских отложений в карьере вблизи ТЭЦ-5 (51.623897° с.ш., 45.989738° в.д.) на северной окраине г. Саратова (рис. 1) во время полевых сезонов: Е.М. Тесаковой, В.Б. Сельцером – в 2011 г., Л.А. Глинских, В.Б. Сельцером в составе полевого отряда под руководством В.В. Митта (ПИН РАН, г. Москва) – в 2012 г. Разрез нижнего–среднего келловея, вскрытый в карманообразной выемке у дороги, ведущей на территорию ТЭЦ-5, и надежно охарактеризованный аммонитами, белемнитами, двустворчатыми моллюсками, гастроподами и другой фауной, изучается В.Б. Сельцером с 1997 г., им же было сделано литологическое описание и биостратиграфическое расчленение по аммонитам (Попов и др., 2004). Позднее, в 2012–2013 гг., это обнажение было переизучено Д.Б. Гуляевым и А.П. Ипполитовым, которые предложили более детальное расчленение по аммонитам (Гуляев, Ипполитов, 2013; Gulyaev, Ippolitov, 2021), использующееся в настоящей работе.
Рис. 1.
Схема расположения изученного разреза келловея ТЭЦ-5 на северной окраине г. Саратова, по (Гуляев, Ипполитов, 2013), с изменениями.
Образцы для микрофаунистического анализа были обработаны по стандартной методике (Основы…, 1959). Пробы весом 200 г замачивали в воде на 1–2 недели, а затем дезинтегрировали простым кипячением в течение нескольких часов. После этого осадок отмывали в проточной воде через сито диаметром ячеи 56 мкм и сушили при комнатной температуре. После технической обработки образцов микрофауну отбирали из сухого осадка и изучали под стереомикроскопами Stemi 2000-С (Carl Zeiss) и Advance ICD (Bresser). Из 24 образцов фораминиферы обнаружены в 22, остракоды – в 7. Частота встречаемости каждого вида фораминифер и остракод определялась по семибалльной шкале: очень редко (1–2 экземпляра), редко (3–5 экземпляров), часто (5–10 экземпляров), очень часто (11–15 экземпляров), много (первые десятки экземпляров), очень много (многие десятки экземпляров), изобилие (сотни экземпляров) (Опорный…, 1969). Фотографирование микрофауны выполнялось с помощью сканирующих электронных микроскопов (СЭМ) в Институте геологии и минералогии им. В.С. Соболева (LEO-1430VP), в Институте нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН (Carl Zeiss EVO 10) и в кабинете приборной аналитики Палеонтологического института им. А.А. Борисяка РАН (TESCAN VEGA-II XMU).
Коллекции фораминифер № ИНГГ ТЭЦ-5 и остракод № ИНГГ ТЭЦ-5-О хранятся в Лаборатории микропалеонтологии ИНГГ СО РАН (г. Новосибирск); коллекция остракод № МГУ ТЭЦ-5 – на кафедре региональной геологии и истории Земли МГУ (г. Москва).
РЕЗУЛЬТАТЫ МИКРОПАЛЕОНТОЛОГИЧЕСКОГО АНАЛИЗА
Биостратиграфия по фораминиферам
Рис. 2.
Распределение фораминифер в нижнем келловее–низах среднего келловея разреза ТЭЦ-5 на северной окраине г. Саратова. Литологическая колонка и расчленение по аммонитам по (Gulyaev, Ippolitov, 2021), с изменениями. G – подзона Gowerianum.
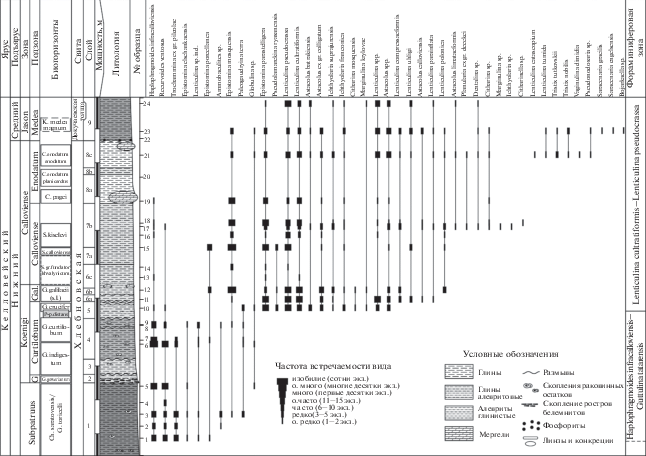
В изученном разрезе нижнего–среднего келловея определено 44 таксона. Фораминиферы распределены по разрезу неравномерно. Наряду с многочисленными и разнообразными в таксономическом отношении комплексами в большей части проб (рис. 2), встречаются уровни, на которых микрофауна не найдена (обр. 20, 22, рис. 2) или характеризуется обедненными комплексами, представленными только агглютинированными формами (обр. 4, рис. 2). Степень сохранности раковин фораминифер различная: от очень хорошей до неудовлетворительной, когда встречаются ядра секреционно-известковых фораминифер, у которых раковины полностью разрушены или сохранились лишь отдельные фрагменты (например, Lenticulina sp. ind., рис. 2), что обусловлено, по-видимому, степенью выветрелости вмещающих пород. По результатам анализа стратиграфического распределения фораминифер в разрезе ТЭЦ-5 установлены две последовательные фораминиферовые зоны: Haplophragmoides infracalloviensis–Guttulina tatarensis и Lenticulina cultratiformis–Lenticulina pseudocrassa (рис. 2). В нижней части разреза (слои 1–4, обр. 1–9) определен комплекс фораминифер, представленный в основном агглютинированными формами, в котором преобладают виды: Haplophragmoides infracalloviensis Dain (табл. I , фиг. 1), Recurvoides ventosus (Chabarova) (табл. I , фиг. 2), Trochammina ex gr. pileolae Startseva (табл. I , фиг. 4). Встречены единичные представители Ammobaculites sp. (табл. I , фиг. 3), Paleogaudryina terra (E. Bykova), Epistomina elschankaensis Mjatliuk, E. porcellanea Brueckmann, E. mosquensis Uhlig (табл. I , фиг. 19 ), Globulina sp., а также ядра раковин рода Lenticulina. Таксономический состав комплекса указывает на фораминиферовую зону Haplophragmoides infracalloviensis–Guttulina tatarensis (верхний бат–нижний келловей) (Унифицированная…, 2012). Выше по разрезу (слои 5–9, обр. 10–24) наблюдается резкая смена комплекса: фораминиферы представлены в основном известковыми формами, за исключением малочисленных представителей H. infracalloviensis, R. ventosus и P. terra, встречающихся в нижней части зоны, причем численно преобладают лентикулины. В комплекс входят виды: Epistomina parastelligera (Hofker), E. mosquensis, E. porcellanea, Pseudolamarckina rjasanensis (Uhlig) (табл. I , фиг. 18 ), Lenticulina pseudocrassa (Mjatliuk) (табл. I , фиг. 12, 13 ), L. cultratiformis (Mjatliuk) (табл. I , фиг. 9–11 ), L. polonica (Wisniowski) (табл. I , фиг. 14 ), L. compressaeformis (Paalzov), L. uhligi (Wisniowski), L. parainflata Grigelis, L. spp., Astacolus batrakiensis (Mjatliuk) (табл. I , фиг. 15 ), A. ex gr. colligatum (Brueckmann), A. calloviensis (Mjatliuk), A. limataeformis (Mitjanina), A. spp., Planularia ex gr. deekei (Wisniowski), Ichthyolaria suprajurensis (Mjatliuk) (табл. I , фиг. 6 ), I. franconica (Guembel) (табл. I , фиг. 7 ), I. sp. (табл. I , фиг. 8 ), Marginulina krylovae Mjatliuk, M. sp., Citharina mosquensis (Uhlig), C. sp. (табл. I , фиг. 17 ), Сitharinella sp., Dentalina sp., Globulina sp. В верхней части разреза (слои 8с–9) обнаружены представители видов: Lenticulina tumida (Mjatliuk), L. catascopium (Mitjanina), Tristix tutkovskii Kaptarenko, T. nobilis Kaptarenko (табл. I , фиг. 5 ), Vaginulina dimidia Grigelis, Saracenaria gracilis Kosyreva in Chabarova, S. engelsensis Kosyreva in Chabarova (табл. I , фиг. 16 ), Pseudonodosaria sp., Bojarkaella sp., характерных для зоны Lenticulina cultratiformis–Lenticulina pseudocrassa среднего келловея (Практическое…, 1991; Унифицированная…, 1993, 2012). Однако в данном разрезе (рис. 2) комплекс фораминифер зоны Lenticulina cultratiformis–Lenticulina pseudocrassa встречается совместно с аммонитами нижнего и среднего келловея (Попов и др., 2004; Гуляев, Ипполитов, 2013; Gulyaev, Ippolitov, 2021). Таким образом, фораминиферовая зона охватывает интервал изученного разреза, отвечающий аммонитовым зонам Koenigi (биогоризонты Crucifer и Galilaeii (s.l.)) и Calloviense нижнего келловея, а также зоне Jason среднего келловея (рис. 2). Полученные данные показывают, что нижняя граница фораминиферовой зоны Lenticulina cultratiformis–Lenticulina pseudocrassa удревняется, а объем ее не ограничен средним келловеем (зонами Jason, Соronatum), как считалось ранее (Биостратиграфия…, 1982; Практическое…, 1991; Унифицированная…, 1993, 2012). Полный объем вышеуказанной фораминиферовой зоны необходимо рассматривать в рамках аммонитовых зон Koenigi (верхняя часть) и Calloviense нижнего келловея, а также зон Jason и Соronatum среднего келловея.
Рис. 3.
Распределение остракод в нижнем келловее–низах среднего келловея разреза ТЭЦ-5 на северной окраине г. Саратова. Литологическая колонка и расчленение по аммонитам по (Gulyaev, Ippolitov, 2021), с изменениями. Условные обозначения см. рис. 2.

Рис. 4.
Схема сопоставления биостратонов по фораминиферам и остракодам для пограничного интервала нижнего–среднего келловея Восточно-Европейской платформы.
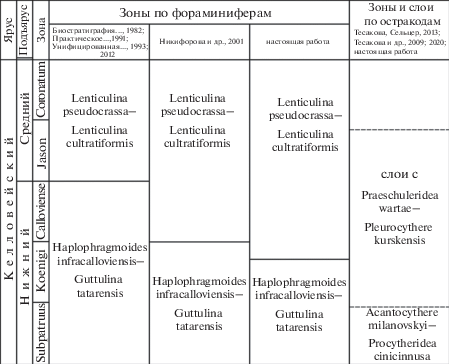
Таблица I . Фораминиферы из нижнего и среднего келловея разреза ТЭЦ-5 Саратовской обл. Фиг. 1, 2 – из зоны Koenigi, подзоны Curtilobum, биогоризонта G. indigestum, обр. 6; фиг. 3 – из зоны Koenigi, подзоны Curtilobum, биогоризонта G. curtilobum, обр. 8; фиг. 4 – из зоны Subpatruus, биогоризонта Ch. saratovensis, обр. 1; фиг. 5, 16, 17 – из зоны Jason, подзоны Medea, обр. 23; фиг. 6, 7, 9, 10, 11, 12, 13, 14, 19 – из зоны Koenigi, подзоны Galilaeii, биогоризонта G. galilaeii, обр. 12; фиг. 8, 15 – из зоны Calloviense, подзоны Calloviense, биогоризонта S. kiselevi, обр. 17; фиг. 18 – из зоны Koenigi, подзоны Curtilobum, биогоризонта G. crucifer, обр. 10. Длина масштабной линейки 100 мкм. 1 – Haplophragmoides infracalloviensis Dain, экз. ИНГГ ТЭЦ-5-01; 2 – Recurvoides ventosus (Chabarova), экз. ИНГГ ТЭЦ-5-02; 3 – Ammobaculites sp., экз. ИНГГ ТЭЦ-5-15; 4 – Trochammina ex gr. pileolae Startseva, экз. ИНГГ ТЭЦ-5-19; 5 – Tristix nobilis Kaptarenko, экз. ИНГГ ТЭЦ-5-03; 6 – Ichthyolaria suprajurensis (Mjatliuk), экз. ИНГГ ТЭЦ-5-04; 7 – Ichthyolaria franconica (Guembel), экз. ИНГГ ТЭЦ-5-16; 8 – Ichthyolaria sp., экз. ИНГГ ТЭЦ-5-05; 9–11 – Lenticulina cultratiformis (Mjatliuk): 9 – экз. ИНГГ ТЭЦ-5-06, вид сбоку, 10 – экз. ИНГГ ТЭЦ-5-07, вид сбоку, 11 – экз. ИНГГ ТЭЦ-5-08, вид со стороны устьевой поверхности; 12, 13 – Lenticulina pseudocrassa (Mjatliuk): 12 – экз. ИНГГ ТЭЦ-5-09, вид сбоку, 13 – экз. ИНГГ ТЭЦ-5-10, вид со стороны устьевой поверхности; 14 – Lenticulina polonica (Wisniowski), экз. ИНГГ ТЭЦ-5-17; 15 – Astacolus batrakiensis (Mjatliuk), экз. ИНГГ ТЭЦ-5-11; 16 – Saracenaria engelsensis Kosyreva in Chabarova, экз. ИНГГ ТЭЦ-5-14; 17 – Citharina sp., экз. ИНГГ ТЭЦ-5-18; 18 – Pseudolamarckina rjasanensis (Uhlig), экз. ИНГГ ТЭЦ-5-12; 19 – Epistomina mosquensis Uhlig, экз. ИНГГ ТЭЦ-5-13.
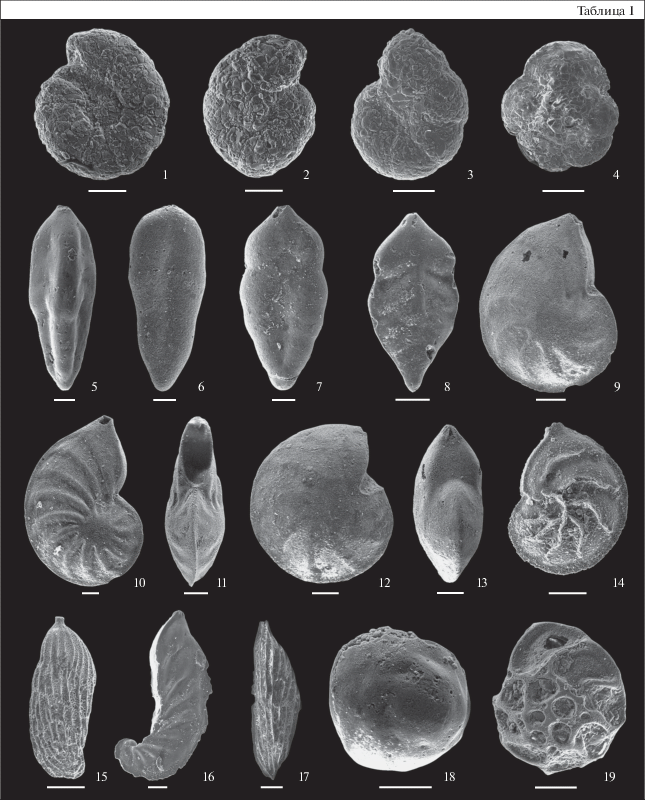
Таблица II. Остракоды из нижнего и среднего келловея разреза ТЭЦ-5 Саратовской обл. Фиг. 1, 4 – из зоны Subpatruus, биогоризонта Ch. saratovensis, обр. 4; фиг. 2, 3, 8, 9, 12, 15 – из зоны Calloviense, подзоны Enodatum, биогоризонта C. enodatum enodatum, обр. 21; фиг. 5, 7, 16, 17 – из зоны Koenigi, подзоны Galilaeii, биогоризонта G. galilaeii, обр. 11; фиг. 6 – из зоны Jason, подзоны Medea, обр. 24; фиг. 10, 11 – из зоны и подзоны Calloviense, биогоризонта S. kiselevi, обр. 15; фиг. 13 – из зоны Jason, подзоны Medea, обр. 24; фиг. 14 – из зоны Subpatruus, биогоризонта Ch. saratovensis, обр. 4. Длина масштабной линейки 100 мкм. 1, 4 – Procytheridea cinicinnusa (Mandelstam in Lyubimova), экз. МГУ ТЭЦ-5-06, ядро целой раковины самки: 1 – вид справа, 4 – вид со спинной стороны; 2, 3 – Praeschuleridea wartae Błaszyk: 2 – экз. ИНГГ ТЭЦ-5-О1, правая створка самки, 3 – экз. ИНГГ ТЭЦ-5-О3, правая створка самца; 5 – Camptocythere sp. A ssp. B sensu Tesakova, 2013, экз. МГУ ТЭЦ-5-27, левая створка ювенильной особи; 6 – Camptocythere (C.) sp. 1, экз. МГУ ТЭЦ-5-17, левая створка ювенильной особи; 7 – Glabellacythere nuda Wienholz, экз. МГУ ТЭЦ-5-29, левая створка ювенильной особи; 8 – Neurocythere plena (Triebel), экз. ИНГГ ТЭЦ-5-О12, левая створка самки; 9 – Neurocythere franconica (Triebel), экз. ИНГГ ТЭЦ-5-О10, правая створка самки; 10, 11 – Lophocythere sp. B: 10 – экз. МГУ ТЭЦ-5-37, правая створка самца; 11 – экз. МГУ ТЭЦ-5-11, левая створка самки; 12 – Lophocythere karpinskyi (Mandelstam in Lyubimova), экз. ИНГГ ТЭЦ-5-О9, правая створка ювенильной особи поздней стадии; 13 – Pleurocythere kurskensis Tesakova, экз. МГУ ТЭЦ-5-15, левая створка самки; 14 – Acantocythere milanovskyi (Lyubimova), экз. МГУ ТЭЦ-5-01, ядро целой раковины самки, вид слева; 15 – Pseudoperissocytheridea anastomosa Whatley, Ballent et Armitage, экз. ИНГГ ТЭЦ-5-О4, левая створка самки; 16 – Procytherura didictyon rossica Tesakova, экз. МГУ ТЭЦ-5-23, правая створка самки; 17 – Procytherura sp., экз. МГУ ТЭЦ-5-22, правая створка самки.
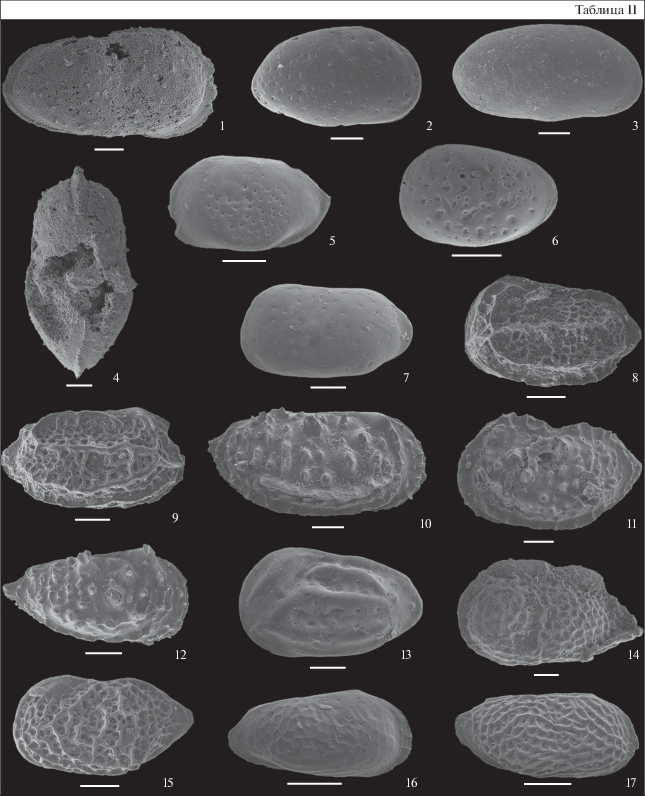
Биостратиграфия по остракодам
В изученном разрезе в общей сложности определено 14 таксонов, представленных, за исключением одного вида, единично и сохранившихся лишь на отдельных уровнях (рис. 3). Столь скудный материал объясняется сильной выветрелостью пород, доступных для изучения в разрезе ТЭЦ-5, и хрупкостью остракодовых створок, предрасположенных к растворению в породах при выветривании. Распределение ракушковых по разрезу позволило выделить четыре различных комплекса, природа которых связана с палеоэкологией, поскольку в ископаемое состояние (тем более в выветрелых породах) попадают массовые таксоны, т.е. виды-доминанты, по смене которых оценивается изменение палеоусловий.
Анализ распределения остракод по разрезу позволил установить два биостратона: остракодовую зону Acantocythere milanovskyi–Procytheridea cinicinnusa и слои с Praeschuleridea wartae–Pleurocythere kurskensis (рис. 3). В интервале разреза, датированном зоной Subpatruus (биогоризонтом Ch. saratovensis; сл. 1, обр. 4 и 5), определен комплекс остракод, состоящий только из видов-индексов. Редкие экземпляры Procytheridea cinicinnusa (Mandelstam in Lyubimova) представлены лишь ядрами целых раковин, а створки не сохранились (табл. II, фиг. 1, 4). Единственная раковина Acantocythere milanovskyi (Lyubimova) сильно деформирована (табл. II, фиг. 14 ), но хорошо узнается по характерной сетчатой скульптуре. Ранее уже было показано, что оба эти вида имеют узкое стратиграфическое распространение в интервале нижнекелловейской зоны Subpatruus в датированных по аммонитам разрезах Бартоломеевка Саратовской обл. (Тесакова, Сельцер, 2013) и Починки Нижегородской обл. (Тесакова и др., 2020), где они и доминировали на отдельных уровнях. При этом P. cinicinnusa появляется в самом низу зоны, в биогоризонте Surensis, а A. milanovskyi позже, в биогоризонте Subpatruus. По их совместному распространению предложено установить одноименную зону, отвечающую аммонитовым биогоризонтам Subpatruus–Saratovensis (Тесакова и др., 2020). Итак, часть разреза ТЭЦ-5, включающую первый комплекс, можно отнести к остракодовой зоне A. milanovskyi–P. cinicinnusa.
В интервале разреза, отвечающем верхней части зоны Koenigi, подзоне Galilaeii и низам одноименных зоны и подзоны Calloviense (биогоризонты G. galilaeii (сл. 6а, обр. 11)–S. calloviense (сл. 7а, обр. 15)), комплекс остракод представлен совершенно иными таксонами: Procytherura didictyon rossica Tesakova (табл. II, фиг. 16 ), P. sp. (табл. II, фиг. 17 ), Camptocythere sp. A ssp. B sensu Tesakova, 2013 (табл. II, фиг. 5 ), Glabellacythere nuda Wienholz (табл. II, фиг. 7 ), Lophocythere sp. B (табл. II, фиг. 10, 11 ) и Pleurocythere kurskensis Tesakova (табл. II, фиг. 13 ). Все перечисленные таксоны характерны для остракодовых слоев с Praeschuleridea wartae–Pleurocythere kurskensis, установленных по совместной встречаемости индексов в Курской области и соответствующих аммонитовой зоне Subpatruus и нижней части зоны Koenigi (подзоны Gowerianus и Curtilobus) (Tesakova, 2003; Тесакова и др., 2009). Следовательно, этот интервал разреза можно смело отнести к слоям с P. wartae–P. kurskensis, несмотря на то, что виды-индексы P. kurskensis и Praeschuleridea wartae Błaszyk (табл. II, фиг. 2, 3) встречены гораздо выше нижней границы биостратона. Более того, стратиграфическое распространение P. kurskensis охватывает интервал от верхов нижнекелловейской зоны Subpatruus до среднеюрской зоны Jason (Tesakova, 2003; Тесакова и др., 2009), а Pr. wartae появляется и вовсе в бате (Błaszyk, 1967). В верхней части разреза, в верхах зоны Calloviense, в подзоне Enodatum, и низах среднекелловейской зоны Jason, в подзоне Medea (биогоризонты C. enodatum enodatum и K. medea magnum; сл. 8с и нижняя часть сл. 9; обр. 21 и 23), совместно с транзитным P. kurskensis появляются и другие виды, характерные для слоев с Pr. wartae–P. kurskensis: Pr. wartae, Pseudoperissocytheridea anastomosa Whatley, Ballent et Armitage (табл. II, фиг. 15 ), Lophocythere karpinskyi (Mandelstam in Lyubimova) (табл. II, фиг. 12 ), Neurocythere plena (Triebel) (табл. II, фиг. 8 ) и N. franconica (Triebel) (табл. II, фиг. 9 ). Все перечисленные виды, кроме P. kurskensis, имеют широкое распространение в Западной, Центральной и Восточной Европе (Triebel, 1951; Lutze, 1960; Błaszyk, 1967; Whatley et al., 2001; Tesakova, 2003; Тесакова, Глинских, 2020 и др.), что говорит об их свободной миграции в пределах всей указанной территории. В самых верхах разреза, в низах зоны Jason в подзоне Medea (сл. 9, обр. 24), встречены: Lophocythere sp. B, P. kurskensis, C. sp. A ssp. B sensu Tesakova, 2013 и Camptocythere (C.) sp. 1 (табл. II, фиг. 6 ). Первые три являются фоновыми таксонами слоев с Pr. wartae–P. kurskensis и обосновывают отнесение данной части разреза к этому биостратиграфическому подразделению. Таким образом, слои с Pr. wartae–P. kurskensis охватывают интервал изученного разреза, отвечающий верхней части зоны Koenigi–низам зоны Jason (рис. 3).
Вопрос о проведении границы между зоной Acantocythere milanovskyi–Procytheridea cinicinnusa и слоями с P. wartae–P. kurskensis дискуссионный. Дело в том, что таксоны-индексы нижнего биостратона (зоны A. milanovskyi–P. cinicinnusa) наблюдались в разрезах Бартоломеевка и ТЭЦ-5 в нижней части биогоризонта Saratovensis, в то время как его верхняя часть там отсутствовала (Гуляев, Ипполитов, 2013, рис. 5 ; Гуляев, 2015, рис. 2; Gulyaev, Ippolitov, 2021, рис. 13 ), и в разрезе Починки во всех биогоризонтах зоны Subpatruus, включая нижнюю часть биогоризонта Saratovensis, при том что само присутствие последнего в этом разрезе неоднозначно (Гуляев, 2015; Тесакова и др., 2020). В том же биогоризонте, но в его верхней части (в то время как его нижняя часть отсутствовала) в разрезах Каневских дислокаций (Гуляев, Ипполитов, 2013, рис. 2; Тесакова и др., 2015, рис. 1; Gulyaev, Ippolitov, 2021, рис. 9 ) и Курской обл. (Тесакова и др., 2009, с. 26 и 29, рис. 2) были встречены другие виды-индексы – Acantocythere nikitini (Lyubimova) и P. kurskensis. Следует учитывать, что оба вида являются неотъемлемой частью слоев с Pr. wartae–P. kurskensis, также идентифицированных в разрезе ТЭЦ-5.
Подобное распределение во времени и пространстве видов-индексов остракод позволяет трактовать его двояко: либо считать их стратиграфическими маркерами нижней и верхней частей биогоризонта Saratovensis, либо экологическими маркерами двух водных масс, одна из которых – западная (Каневские и Курские разрезы) – постепенно смещалась с запада на восток (Саратовские и Нижегородские разрезы) в течение фазы Subpatruus. Для однозначной интерпретации необходимо изучение остракод из полного разреза биогоризонта Saratovensis. Пока у автора (Е.М. Тесаковой) нет подобного материала, но литературные данные о присутствии этих взаимоисключающих пар в одних и тех же разрезе как на Украине (Пяткова, Пермякова, 1978, с. 144), так и на Самарской Луке (разрез Репьевка; Любимова, 1955, с. 125–127, табл. 1, 2, 5 ) позволяют склоняться все же к стратиграфической последовательности и с осторожностью предполагать проведение границы между зоной A. milanovskyi–P. cinicinnusa и слоями с P. wartae–P. kurskensis внутри биогоризонта Saratovensis (рис. 4; граница показана пунктиром).
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования выявлен полный систематический состав комплексов фораминифер и остракод и их распределение в нижнем келловее и основании среднего келловея в разрезе ТЭЦ-5 на северной окраине г. Саратова. Полученные результаты микропалеонтологического анализа по фораминиферам и остракодам сопоставлены с данными по аммонитам (Репин, Рашван, 1996; Попов и др., 2004; Gulyaev, Ippolitov, 2021), а также с опубликованными данными по микрофауне (Олферьев и др., 1992; Никифорова и др., 2001; Устинова, 2017) и друг с другом (рис. 4). На основе анализа распространения фораминифер в изученном разрезе установлены две зоны: Haplophragmoides infracalloviensis–Guttulina tatarensis и Lenticulina cultratiformis–Lenticulina pseudocrassa; уточнен стратиграфический объем и положение нижней границы последней. Обосновано, что зона Lenticulina cultratiformis–Lenticulina pseudocrassa Восточно-Европейской платформы не ограничена только средним келловеем, как считалось ранее (Биостратиграфия…, 1982; Унифицированная…, 1993, 2012 и др.), а охватывает верхи нижнего келловея (в объеме аммонитовых зон Koenigi (верхняя часть) и Calloviense) и средний келловей (зоны Jason, Coronatum). И ее нижнюю границу следует проводить в верхней части зоны Koenigi. По распределению остракод в изученном разрезе выделено два биостратона: зона Acantocythere milanovskyi–Procytheridea cinicinnusa и слои с Praeschuleridea wartae–Pleurocythere kurskensis. Зона A. milanovskyi–P. cinicinnusa Восточно-Европейской платформы охватывает верхнюю половину нижнекелловейской зоны Subpatruus без терминальной части биогоризонта Saratovensis (Тесакова, Сельцер, 2013; Тесакова и др., 2020) и соответствует средней части фораминиферовой зоны H. infracalloviensis–G. tatarensis. Слои с Ps. wartae–P. kurskensis коррелируют с терминальной частью аммонитовой зоны Subpatruus (терминальной частью биогоризонта Saratovensis) и зонами Koenigi и Calloviense нижнего келловея, а также с нижней частью зоны Jason среднего келловея (Тесакова и др., 2009, 2015); они же отвечают следующим зонам по фораминиферам: верхней части H. infracalloviensis–G. tatarensis и нижней части L. cultratiformis–L. pseudocrassa.
Благодарности. Авторы глубоко признательны В.В. Митта (ПИН РАН, Москва) за организацию и проведение экспедиционных работ, Я.А. Шуруповой (МГУ, Москва) за отмывку образцов, отобранных из разреза Е.М. Тесаковой, Р.А. Ракитову (ПИН РАН, Москва) за неизменную помощь в работе на СЭМ и А.П. Ипполитову (ГИН РАН, Москва) за конструктивные замечания, улучшившие эту статью.
Источники финансирования. Работа выполнена в рамках тем госзадания №№ 0135-2019-0062 (ГИН РАН), АААА-А16-116033010096-8 (МГУ), при частичной поддержке РФФИ № 19-05-00130 и является вкладом в проект ФНИ № 0331-2019-0005.
Список литературы
Биостратиграфия верхнеюрских отложений СССР по фораминиферам. Вильнюс: Мокслас, 1982. 173 с.
Быкова Е.В. О значении ископаемых фораминифер для стратиграфии юрских отложений района Самарской Луки // Тр. ВНИГРИ. Нов. сер. 1948. Вып. 31. С. 83–108.
Гуляев Д.Б. Стратиграфия пограничных отложений бата и келловея Европейской России // Юрская система России: проблемы стратиграфии и палеогеографии. Шестое Всероссийское совещание. Научные материалы. Отв. ред. Захаров В.А. Махачкала: АЛЕФ, 2015. С. 94–101.
Гуляев Д.Б., Ипполитов А.П. Детальная биостратиграфия нижнего келловея района Каневских дислокаций (Черкасская обл., Украина) // Юрская система России: проблемы стратиграфии и палеогеографии. Пятое Всероссийское совещание, 23–27 сентября 2013 г., Тюмень. Научные материалы. Отв. ред. Захаров В.А. Екатеринбург: Издательский дом “ИздатНаукаСервис”, 2013. С. 65–72.
Даин Л.Г. Материалы к стратиграфии юрских отложений Саратовской области // Тр. ВНИГРИ. Нов. сер. 1948. Вып. 31. С. 49–81.
Даин Л.Г. Значение фораминифер для стратиграфии восточной полосы Русской платформы // Тр. ВНИГРИ. 1961. Вып. 29. Т. 3. С. 168–176.
Любимова П.С. Остракоды мезозойских отложений Среднего Поволжья и Общего Сырта // Тр. ВНИГРИ. Нов. сер. 1955. Вып. 84. С. 3–190.
Мятлюк Е.В. Фораминиферы верхнеюрских и нижнемеловых отложений Среднего Поволжья и Общего Сырта // Тр. НГРИ. Сер. А. 1939. Вып. 120. С. 1–76.
Мятлюк Е.В. Описание новых видов фораминифер верхнеюрских и меловых отложений Русской платформы // Тр. ВНИГРИ. 1961. Вып. 29. Т. 3. С. 142–164.
Никифорова Е.В., Колпенская Н.Н., Овчинникова Л.Л. Новые данные о составе микробиоты в отложениях келловейского яруса опорного разреза “Малиновый овраг” (Саратовское Поволжье) // Эволюция жизни на Земле. Материалы II Международного симпозиума, 12–15 ноября 2001 г., Томск. Отв. ред. Подобина В.М. Томск: Изд-во НТЛ, 2001. С. 291–293.
Олферьев А.Г., Меледина С.В., Азбель А.Я. Новые данные о стратиграфии келловея Щигровского свода Воронежской антеклизы // Геологическая история Арктики в мезозое и кайнозое. Материалы чтений памяти В.Н. Сакса. Кн. 1. СПб.: ВНИИОкеангеология, 1992. С. 49–57.
Опорный разрез верхнеюрских отложений р. Хеты (Хатангская впадина). Л.: Наука, 1969. 124 с.
Основы палеонтологии. Общая часть. Простейшие. М.: Изд-во АН СССР, 1959. 482 с.
Попов Е.В., Сельцер В.Б., Волков А.В. О находке зубов эласмобранхий (Chondrichtyes, Elasmobranchii) в нижнем келловее Саратова // Вопросы палеонтологии и стратиграфии верхнего палеозоя и мезозоя. Саратов: Научная книга, 2004. С. 133–154.
Практическое руководство по микрофауне СССР. Т. 5. Фораминиферы мезозоя. Ред. Азбель А.А., Григялис А.А. Л.: Недра, 1991. 375 с.
Пяткова Д.М., Пермякова М.Н. Фораминиферы и остракоды юры Украины. Киев: Наукова думка, 1978. 288 с.
Репин Ю.С., Рашван Н.Х. Келловейские аммониты Саратовского Поволжья и Мангышлака. СПб.: НПО “Мир и семья-95”, 1996. 256 с.
Тесакова Е.М., Глинских Л.А. Келловейские остракоды Центрального Дагестана: биостратиграфия, палеоэкология и палеогеография // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 4. С. 96–110. https://doi.org/10.31857/S0869592X20040092
Тесакова Е.М., Сельцер В.Б. Остракоды и аммониты нижнего келловея разреза Бартоломеевка (Саратовская обл.) // Бюлл. МОИП. Отд. геол. 2013. Т. 88. Вып. 2. С. 50–68.
Тесакова Е.М., Стреж А.С., Гуляев Д.Б. Новые остракоды из нижнего келловея Курской обл. // Палеонтол. журн. 2009. № 3. С. 25–36.
Тесакова Е.М., Ипполитов А.П., Гуляев Д.Б. Реконструкция по остракодам раннекелловейских событий района Каневских дислокаций (предварительные данные) // Юрская система России: проблемы стратиграфии и палеогеографии. Шестое Всероссийское совещание, 15–20 сентября 2015 г., Махачкала. Научные материалы. Отв. ред. Захаров В.А. Махачкала: АЛЕФ, 2015. С. 273–277.
Тесакова Е.М., Глинских Л.А., Федяевский А.Г., Мироненко А.А., Ипполитов А.П. Микрофоссилии из нижнекелловейской аммонитовой зоны Subpatruus Нижегородской области // Юрская система России: проблемы стратиграфии и палеогеографии. Материалы VIII Всероссийского совещания с международным участием. Онлайн-конференция, 7–10 сентября 2020 г. Отв. ред. Захаров В.А. Сыктывкар: ИГ Коми НЦ УрО РАН, 2020. С. 222–230.
Унифицированная стратиграфическая схема юрских отложений Русской платформы. СПб.: ВНИГРИ, 1993. 28 л., 71 с.
Унифицированная региональная стратиграфическая схема юрских отложений Восточно-Европейской платформы (14 листов). Объяснительная записка. М.: ПИН РАН–ВНИГНИ, 2012. 64 с.
Устинова М.А. Новые данные о фораминиферах келловея (средняя юра) пограничных районов Курской и Орловской областей // Бюлл. МОИП. Отд. геол. 2017. Т. 92. Вып. 5. С. 50–59.
Устинова М.А., Тесакова Е.М. Сравнительный анализ распространения раннекелловейских фораминифер и остракод из разреза Бартоломеевка (Саратовская область) // Вестник Моск. ун-та. Сер. 4. Геология. 2015. № 3. С. 73–84.
Хабарова Т.Н. Фораминиферы юрских отложений Саратовской области // Тр. ВНИГРИ. 1959. Вып. 137. С. 461–519.
Хабарова Т.Н. О микрофауне юрских отложений Саратовской области // Тр. ВНИГРИ. 1961. Вып. 29. Т. 3. С. 177–184.
Błaszyk J. Middle Jurassic ostracods of the Częstochowa Region (Poland) // Acta Palaeontol. Polon. 1967. V. 12. № l. P. 4–75.
Gulyaev D.B., Ippolitov A.P. Lower Callovian of Kanev Dislocations, Cherkassy Oblast, Ukraine: ammonites and stratigraphy // Stratigr. Geol. Correl. 2021. V. 29. № 7. P. 1–80.
Lutze G.F. Zur stratigraphie und paläontologie des Callovien und Oxfordien in Nordwest-Deutschland // Geol. Jahr. 1960. V. 77. P. 391–532.
Tesakova E.M. Callovian and Oxfordian ostracodes from the Central Region of the Russian Plate // Paleontol. J. 2003. V. 37. Suppl. 2. P. 107–227.
Tesakova E.M. Ostracode-based reconstruction of paleodepths in the Early Callovian of the Kursk Region, Central Russia // Paleontol. J. 2013. V. 47. № 10. P. 1–16.
Triebel E. Ostracoden aus dem hoheren Dogger Deutschlands // Abh. Senck. Naturf. Ges. 1951. V. 485. P. 87–101.
Whatley R.C., Ballent S., Armitage J. Callovian Ostracoda from the Oxford Clay of Southern England // Rev. Esp. Micropaleontol. 2001. V. 33. № 2. P. 135–162.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция