

Палеонтологический журнал, 2022, № 6, стр. 80-91
К ревизии пермских лучеперых рыб Европейской России. Часть 2
А. С. Бакаев *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: alexandr.bakaev.1992@mail.ru
Поступила в редакцию 25.04.2022
После доработки 01.06.2022
Принята к публикации 01.06.2022
- EDN: OTXYEA
- DOI: 10.31857/S0031031X22060034
Аннотация
Проведена ревизия части оригиналов пермских лучеперых рыб Европейской России из коллекции Д.Н. Есина, которая была передана в Палеонтологический институт им. А.А. Борисяка РАН (ПИН РАН). Приведены описания и изображения таксонов, изменивших первоначальный (присвоенный при первоописании) систематический статус: Reticulolepis insolita (переименован в Strelnia insolita), Samarichthys nikolaevae (переименован в Acroctenolepis nikolaevae), Wardichthys inobilis (младший синоним Kargalichthys pritokensis), Amblypterina pectinata (младший синоним Isadia suchonensis) и Amblypterina grandicostata (младший синоним Isadia aristoviensis). При этом изображение голотипов Reticulolepis insolita и Samarichthys nikolaevae приводится впервые.
ВВЕДЕНИЕ
Среди палеоихтиологов, изучавших лучеперых рыб перми Европейской России, Дмитрий Николаевич Есин был одним из наиболее значимых специалистов (Бакаев, в печати). Из собственных сборов им был описан ряд новых таксонов пермских лучеперых рыб. Коллекция Есина после его смерти в 2003 г. была передана в ПИН РАН из Московского государственного университета им. М.В. Ломоносова (МГУ), на базе которого и была собрана. До последнего момента коллекция не подвергалась ревизии, а хранилась под устаревшими номерами МГУ.
Всего в коллекции Есина находились оригиналы 12 описанных им видов рыб: Wardichthys inobilis Esin, 1993 (Есин, 1993), Acropholis kamensis Esin, 1995, Kazanichthys golyushermensis Esin, 1995, Koinichthys ivachnenkoi Esin, 1995, Elonichthys contortus Esin, 1995 (Есин, 1995а), Acropholis silantievi Esin, 1996, Kazanichthys viatkensis Esin, 1996, Boreolepis tataricus Esin, 1996, Reticulolepis insolita Mashin et Esin, 1996, Acroctenolepis nikolaevae Esin, 1996, Amblypterina grandicostata Esin, 1996, A. pectinata Esin, 1996 (Есин, Машин, 1996). При осмотре переданной коллекции оригинальные экземпляры Koinichthys ivachnenkoi и Elonichthys contortus обнаружить не удалось. Среди оставшихся таксонов некоторые сохранили свой систематический статус (Acropholis kamensis, A. silantievi, Kazanichthys golyushermensis K. viatkensis, Boreolepis tataricus) (Бакаев, 2022), а другие изменили его (Wardichthys inobilis, Reticulolepis insolita, Acroctenolepis nikolaevae, Amblypterina grandicostata, A. pectinata). В настоящей статье мы рассмотрим те таксоны, у которых изменился систематический статус (см. ниже).
МАТЕРИАЛ И МЕТОДЫ
Изучаемый материал представлен как изолированными чешуями хорошей и удовлетворительной сохранности (Amblypterina grandicostata, A. pectinata, Reticulolepis insolita), так и фрагментами скелетов (Wardichthys inobilis и Acroctenolepis nikolaevae). Оригинал Reticulolepis insolita происходит из местонахождения Полдарса. Чешуи Amblypterina grandicostata происходят из местонахождения Пронькино, а чешуи A. pectinata – из местонахождения Кочевала-2. Оригиналы Wardichthys inobilis и Acroctenolepis nikolaevae были обнаружены в местонахождении Шихово-Чирки.
Описания изолированных чешуй подготовлены с использованием предложенной Есиным (1990) методики. Отдельные чешуи отсняты в кабинете приборной аналитики ПИН РАН на микроскопах TESCAN VEGA-II XMU и TESCAN VEGA-III XMU (без напыления). Общее фото голотипов Wardichthys inobilis и Acroctenolepis nikolaevae выполнено при помощи фотоаппарата Canon EOS 650D.
ГЕОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
Полдарса. Вологодская обл., Великоустюгский р-н, левый берег р. Сухона у с. Полдарса (Татарские слои…, 2001).
Костеносный уровень: мергели зеленовато-серые, светло-серые, отдельными прослоями сиреневые, микрозернистые, слабоглинистые, тонкослоистые, с ходами илоедов и остатками остракод (Татарские слои…, 2001).
Палеонтологический материал: рыбы: Strelnia insolita, Uranichthys pritokensis (в колл. автора); а также: Xenosynechodus sp., Platysomus biarmicus, Platysomus sp., Geryonichthys sp., Varialepis sp. (Татарские слои…, 2001; Есин, 1995б); тетраподы Suchonica vladimiri; двустворчатые моллюски (Татарские слои…, 2001).
Стратиграфический уровень: верхнепермский (татарский) отдел, северодвинский ярус, верхнесеверодвинский подъярус, путятинский горизонт, нижнепутятинский подгоризонт, полдарсская свита, устьполдарская пачка (Татарские слои…, 2001).
Кочевала-2. Вологодская обл., Нюксеницкий р-н, правый берег р. Сухона, 500 м ниже по течению от устья р. Кочевала, 200 м ниже по течению от линии электропередач (Есин, 1995б).
Костеносный уровень: слой 7. Песчаник розовато-серый, светло-коричневый плотный (Есин, 1995б).
Палеонтологический материал: рыбы: Toyemia tverdochlebovi, Geryonichthys longus, Varialepis cf. stanislavi; тетраподы Microphon exiguus (Ивахненко и др., 1997).
Стратиграфический уровень: верхнепермский (татарский) отдел, северодвинский ярус, верхнесеверодвинский подъярус, путятинский горизонт, полдарcская свита (Ивахненко и др., 1997).
Пронькино. Оренбургская обл., Сорочинский р-н, левый коренной склон долины р. Боровка, в 1 км от центральной площади с. Пронькино, примерно на половине высоты склона (Ефремов, Вьюшков, 1955).
Костеносный уровень: слой 31. Песчаник светло-серый или зеленоватый, реже красноватый, пестрый, неравномерно уплотненный, сильно глинистый, мелкозернистый, переслаивающийся с глинистым алевролитом. Мощность 0.5 м (Ефремов, Вьюшков, 1955).
Палеонтологический материал: рыбы: Gnathorhiza cf. otschevi Minich, Boreolepis tataricus, Toyemia blumentalis Minich, Isadia aristoviensis Minich, I. cf. suchonensis Minich, Isadia sp., Varialepis vitalii Minich, Acropholis cf. stensioei (Есин, 1995б; Tverdokhlebov et al., 2005); тетраподы: Dvinosaurus primus, Karpinskiosaurus ultimus, Chroniosuchus paradoxus, Kotlassia cf. prima, Leogorgon klimovensis, Pareiasaurida fam. indet., Peramodon sp. (Ивахненко и др., 1997); конхостраки Esteria lineata Lutk., Е. angulata Lutk., E. rotunda Lutk. и E. elongata Netsch. (Ефремов, Вьюшков, 1955).
Стратиграфический уровень: верхнепермский (татарский) отдел, вятский ярус, кутулукская свита (Голубев, Наумчева, 2019).
Шихово-Чирки. Кировская обл., Слободской р-н, правый берег р. Вятка, выше устья р. Чепца, ниже д. Чирки, 22 км от г. Киров. Костеносная зона расположена в северо-восточном углу карьера, около границы полей Шихово-Чирков и земель известкового завода. Подробное описание см. (Гоманьков и др., 1993).
Костеносный уровень: слой 4. Известняк светлый или серый, плотный или мягкий, переслаивающийся с мергелями серыми, иногда слоистыми. Мощность до 9 м (Ефремов, Вьюшков, 1955).
Палеонтологический материал: рыбы: Kazanichthys viatkensis, Samarichthys nikolaevae, Palaeoniscum kasanense, P. freiselebeni, Platysomus biarmicus, Elonichthys cf. contortus, Kargalichthys pritokensis (Есин, 1995б); тетраподы: Leptoropha talonophora, Melosaurus platyrhinus, Nyctiboetus kassini, Platyoposaurus watsoni; двустворчатые моллюски Palaeomutela vjatkensis, Prilukiella sp., P. lata; растения Phyllotheca sp., Catamites sp. (Голубев, 1995).
Рис. 1.
Географическое и стратиграфическое распространения описываемых рыб: а – карта европейской части России с используемыми местонахождениями: 1 – Кочевала-2, 2 – Полдарса, 3 – Пронькино, 4 – Шихово-Чирки; б – стратиграфическая шкала и распространение описанных таксонов, основанное на комплексе имеющихся литературных и авторских данных (не только по типовым местонахождениям). Сокращения: МСШ – международная стратиграфическая шкала; ОСШ – общая стратиграфическая шкала; РСШ – региональная стратиграфическая шкала.
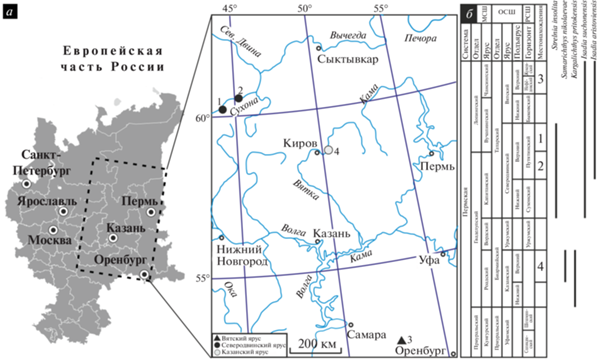
Стратиграфический уровень: среднепермский (биармийский) отдел, казанский ярус, верхнеказанский подъярус, верхнеуслонские слои (Стратотипический …, 2001).
НАДКЛАСС OSTEICHTHYES
КЛАСС ACTINOPTERYGII
ОТРЯД ELONICHTHYIFORMES
FAMILIA INC. SED.
Род Strelnia A. Minich, 2009
Strelnia insolita (Esin, 1996)
Reticulolepis insolita: Есин, Машин, 1996, с. 284, рис. 5.5 -10.
Голотип – ПИН, № 5842/1 [ранее – МГУ, № SU 32 PL 15/1 (Есин, Машин, 1996), на образце номер PL 9/1], чешуя участок В; Вологодская обл., Великоустюгский р-н, местонахождение Полдарса; верхнепермский (татарский) отдел, северодвинский ярус, верхнесеверодвинский подъярус, путятинский горизонт, нижнепутятинский подгоризонт, полдарсская свита, устьполдарская пачка.
Описание (по: Есин, Машин, 1996, с изменениями, для чешуй участка В) (рис. 2). Чешуи крупные. Сочленовный шип высокий, с широким основанием. Его средняя линия проходит чуть впереди переднего края свободного поля. Передний край прямой. Антеродорсальный угол сильно вытянут вверх и превышает сочленовный шип. Антеровентральный угол у чешуй из передней части тела срезан крутым углом. Постеровентральный угол также срезан, но под незначительным углом. Вентральный край чешуи очень выпуклый, приостренный. Дорсальный край сильновогнутый. Ямка глубокая и длинная, киль хорошо выражен. Свободное поле широкое, у передних чешуй составляет около половины длины чешуи; передний край его почти прямой. Окончания языковидных выступов округлые. Гребни широкие, округлые в сечении. Борозды, разделяющие гребни, в основном неширокие и гораздо у́же гребней, вытянуты по диагонали чешуи. В бороздах находятся многочисленные поры. Большинство гребней продолжаются до середины чешуи, где они анастомозируют или оканчиваются заостренными выступами. За ними берут начало гребни второго порядка. Некоторые гребни проходят до заднего края чешуи, где, наряду с гребнями второго порядка, оканчиваются лишь немного выходя за край костной пластинки и образуют длинные (пектинатные) зубцы. Наиболее короткие и ярко выраженные гребни наблюдаются в постеродорсальном и антеровентральном углах свободного поля.
Рис. 2.
Strelnia insolita (Esin, 1996): а, г – голотип ПИН, № 5842/1; б, д – экз. ПИН, № 5842/3; в, е – экз. ПИН, № 5842/2; ж, з – детали строения фиг. д, е при большем увеличении; Вологодская обл., Великоустюгский р-н, местонахождение Полдарса; верхнепермский (татарский) отдел, северодвинский ярус, верхнесеверодвинский подъярус, путятинский горизонт, нижнепутятинский подгоризонт, полдарсская свита, устьполдарская пачка. Длина масштабной линейки 5 мм для фиг. а–в, 1 мм для остальных.

Размеры. Длина голотипа (изолированной чешуи) – 0.6 см; высота – 0.6 см. Размеры всей рыбы реконструировать затруднительно.
Сравнение. От сходного вида S. certa A. Minich отличается менее ярко выраженной скульптурой: у S. certa гребни, образующие скульптуру свободного поля, более выпуклые, а разделяющие их борозды (на соответствующих участках чешуйного покрова) глубже, шире и многочисленнее, чем у S. insolita. Так, на чешуях участков В и С рассекающие переднюю часть свободного поля борозды у S. certa шире разделяемых ими гребней и проходят дальше, чем у S. insolita, а борозды, рассекающие заднюю часть, глубже и берут начало ближе к передней части чешуи. Соответственно, свободное поле в целом более изрезано, а скульптура четче и ярче выражена у S. certa.
Замечания. При первом описании (Есин, Машин, 1996) этот вид был отнесен к роду Reticulolepis из верхней перми (лопингский отдел) Западной Европы (Aldinger, 1937). Однако, несмотря на определенное сходство с Reticulolepis, чешуи R. insolita в большей степени похожи на Strelnia certa. Соответственно, S. insolita должен быть отнесен к роду Strelnia, а Strelnia и Reticulolepis являются близкими родственниками. Мы предполагаем, что Strelnia insolita менее специализированный и предковый для S. сerta вид, что выражено в меньшем развитии у него признаков орнамента чешуи. Это также подтверждается распространением S. certa в более молодых частях разреза р. Сухона с устьполдарсской по кичугскую пачки (Молостовская и др., 2001).
Распространение. Европейская часть России; верхнепермский (татарский) отдел, северодвинский ярус.
Материал. Кроме голотипа, также в колл. ПИН переданы образцы МГУ №№ SU 32 PL 9/2, 9/4, получившие номера ПИН №№ от 5842/2 и 5842/3 соответственно.
Род Samarichthys A. Minich, 1990
Samarichthys nikolaevae (Esin, 1996)
Acroctenolepis nikolaevae: Есин, Машин, 1996, с. 285, рис. 5.5 -11 а, б, табл. 5.5 -4, фиг. 7–10 .
Samarichthys nikolaevae: Миних, Миних, 2009, с. 71, табл. 7 , фиг. 6–9 .
Голотип – ПИН, № 5802/14 (ранее – МГУ № ШЧ 1/12), скелет рыбы (голова и грудные плавники не сохранились); Кировская обл., Слободской р-н, местонахождение Шихово-Чирки; среднепермский (биармийский) отдел, казанский ярус, верхнеказанский подъярус, верхнеуслонские слои.
Описание (по Есин, Машин, 1996, с изменениями, для чешуй участка В) (рис. 3). Чешуи почти прямоугольной или ромбической формы. Передний край прямой. Погруженное поле узкое, составляет от 1/6 до 1/4 длины чешуи. Нижний край плавно выгнут вниз. Антеровентральный угол слабо скошен; на чешуях передней части тела он заострен и направлен вниз, образуя нижнюю шпору. Шип крупный, высокий, широкий у основания и заостренный на конце. Направлен вертикально вверх. Ямка крупная, глубокая. Киль узкий, расположен впереди шипа. Передний верхний угол низкий, с заостренной вершиной.
Рис. 3.
Samarichthys nikolaevae (Esin, 1996): а – голотип ПИН, № 5802/14; б–к – то же, детали строения при большем увеличении; Кировская обл., Слободской р-н, местонахождение Шихово-Чирки; среднепермский (биармийский) отдел, казанский ярус, верхнеказанский подъярус, верхнеуслонские слои. Обозначения: Af – анальный плавник; Cf – хвостовой плавник; Df – спинной плавник; H – голова; Pecf – грудной плавник; Pelf – брюшной плавник. Длина масштабной линейки 5 мм для фиг. а, 1 мм для остальных.

Языковидные выступы имеют заостренные окончания. Гребни скульптуры высокие, треугольные в сечении, короткие в постеродорсальной части свободного поля и более длинные, параллельные нижнему краю чешуи в антеровентральной части свободного поля. Гребни несут выраженные гребешки второго порядка, заканчиваются заостренными зубцами, не выходящими за край чешуи. Гребни могут довольно сильно извиваться, соединяться и разделяться на отдельные ветви, однако в целом направлены диагонально. Задний край несет короткие зубцы, которыми оканчиваются доходящие до края гребни. Число их различно.
Размеры. Узнать длину тела голотипа затруднительно из-за неполной сохранности, реконструируемая длина – примерно 25 см.
Сравнение. S. nikolaevae очень похож на S. luxus (Esin, 1996) но отличается более уплощенными и широкими гребнями, а также прямыми, относительно широкими окончаниями языковидных выступов. Кроме того, четче выделяются зоны длинных продольных гребней в антеровентральной половине свободного поля, и коротких треугольных – в постеродорсальной.
Замечания. При первом описании (Есин, Машин, 1996) голотип был указан, но не изображен. Однако голотип был изображен в неопубликованной диссертации Есина (1995б, табл. XIII , фиг. 8 ), что позволило безошибочно установить его принадлежность. Несмотря на то, что изображение голотипа при его описании желательно, “Международный кодекс зоологической номенклатуры” (2004) не запрещает подобный тип опубликования, то есть Samarichthys nikolaevae является валидным таксоном с момента публикации первого описания (Есин, Машин, 1996).
Данный вид впервые был установлен Есиным, но отнесен к роду Acroctenolepis (Есин, Машин, 1996). А.В. и М.Г. Миних (2009) отнесли его к ранее описанному роду Samarichthys, но рассматривали S. nikolaevae как отдельный вид, несмотря на очень значительную степень сходства с S. luxus. Оба таксона надежно отличаются только при наличии репрезентативной выборки чешуй.
Распространение. Европейская часть России; среднепермский (биармийский) отдел, казанский ярус, верхнеказанский подъярус.
Материал. В переданной коллекции не удалось обнаружить чешуи МГУ №№ ШЧ 200/3-42, 1-39, 3-4, 3-32 (Есин, Машин, 1996, табл. 5.5 -4, фиг. 7–10 ), кроме тех, что содержатся на голотипе. Однако опубликованные изображения и чешуи голотипа позволяют безошибочно отнести некоторые чешуи из типового местонахождения Шихово-Чирки к этому виду.
ОТРЯД BOBASATRANIIFORMES
СЕМЕЙСТВО PLATYSOMIDAE YOUNG, 1866
Род Kargalichthys A. Minich, 1990
Kargalichthys pritokensis Minich, 1992
Kargalichthys pritokensis: Миних, 1992, с. 143, табл. II , фиг. 2; Миних, Миних, 2009, с. 162, табл. 46 , фиг. 4; табл. 50 , фиг. 3.
Рис. 4.
Kargalichthys pritokensis Minich, 1992: а – экз. ПИН, № 5802/16; б–з – то же, детали строения при большем увеличении; Кировская обл., Слободской р-н, местонахождение Шихово-Чирки; среднепермский (биармийский) отдел, казанский ярус, верхнеказанский подъярус, верхнеуслонские слои. Условные обозначения как на рис. 3. Длина масштабной линейки 5 мм для фиг. а, 1 мм для остальных.

Wardichthys inobilis: Есин, 1993, с. 131, рис. 1, г, д.
Голотип – СГУ, № 104-Б/2052, группа чешуй; Оренбургская обл., Александровский р-н, местонахождение Притокский; среднепермский (биармийский) отдел, казанский ярус, верхнеказанский подъярус, белебеевская свита.
Описание (по Есин, 1993, с изменениями) (рис. 4). Чешуи высокие, тонкие, прямоугольные. Свободное поле скульптировано мощными, высокими, короткими и длинными гребнями, отдельных зубцов мало. Гребни округло-треугольные (часто – с уплощенной вершиной) в сечении, широкие, толстые, часто соединяющиеся. Борозды между гребнями уже самих гребней, несут многочисленные поры. Скульптура на всех чешуях почти одинакова, а если незначительно изменяется, то не демонстрирует устойчивой закономерности. Края прямые. Антеродорсальный угол практически не выражен, антеровентральный угол часто сильно вытянут вниз. Оба задних угла прямые. Сочленовный шип треугольный, очень высокий и широкий, плоский, основание его часто почти равно ширине чешуи; часто несет вертикальные желобки. Ямка также очень широкая и длинная (обычно доходит примерно до половины высоты чешуи). Киль выражен слабо, в виде плоского вздутия впереди от ямки. Вдоль киля сзади идет вертикальный ряд пор, за которым расположены беспорядочно разбросанные немногочисленные поры. Погруженное поле занимает 1/2–1/3 длины чешуи. Передний край свободного поля прямой, окончания языковидных выступов прямые.
Размеры. Узнать длину тела затруднительно из-за значительной деформации. Реконструируемая длина экз. ПИН, № 5802/16 примерно равна 20 см.
Сравнение. От K. efremovi отличается скульптурой свободного поля: более широкими, мощными, часто соединяющимися гребнями, гораздо меньшим числом коротких зубцов.
Замечания. Есин (1993) описал Wardichthys inobilis как самостоятельный вид, но впоследствии (Миних, Миних, 2009) была продемонстрирована синонимичность этого вида ранее описанному Kargalichthys pritokensis Minich, 1992.
Распространение. Европейская часть России; среднепермский (биармийский) отдел, казанский ярус.
Материал. Экз. ПИН, № 5802/16 (ранее – МГУ № 263-2/12, голотип Wardichthys inobilis), фрагмент скелета (задняя часть туловища); Кировская обл., Слободской р-н, местонахождение Шихово-Чирки; среднепермский (биармийский) отдел, казанский ярус, верхнеказанский подъярус, верхнеуслонские слои.
ОТРЯД EURYNOTOIDIFORMES
СЕМЕЙСТВО EURYNOTOIDIDAE MINIKH ET A. MINIKH, 1990
Род Isadia A. Minich, 1990
Isadia suchonensis A. Minich, 1990
Табл. XIV , фиг. 1–6 (см. вклейку)
Isadia suchonensis: Миних, А. Миних, 1986, с. 18, рис. 4, in litt.; М. Миних, А. Миних, 1990, с. 94, табл. VI , фиг. 1–4; А. Миних, М. Миних, 2009, с. 195, рис. 28 , табл. 59 , фиг. 1–6 .
Amblypterina pectinata: Есин, Машин, 1996, с. 87, рис. 5.5 -12б, табл. 5.5 -II, фиг. 1–4.
Голотип – СГУ, № 104-Б/З-2а, б, передняя часть тела с головой; Вологодская обл., Великоустюгский р-н, местонахождение Мутовино; верхнепермский (татарский) отдел, северодвинский ярус, путятинский горизонт, полдарсская свита, кичугская пачка.
Описание (по Есин, Машин, 1996, с изменениями, для чешуй участка А). Чешуи тонкие, передний и задний края прямые. Нижний край сильновыпуклый, верхний сильновогнутый. Антеродорсальный угол высокий, оттянут немного вперед (на некоторых образцах) или прямой, конец заострен. Антеровентральный угол скошен, прямой или немного закругленный, имеет резкий перелом с передним краем. На наиболее крупных чешуях в этом месте есть нижняя шпора. Постеровентральный угол не резко скошен, закруглен, переход к заднему краю плавный. Постеродорсальный угол тупой, несколько приподнят. Сочленовный шип и ямка сильно развитые, высокие, расположены в середине длины чешуи. Сочленовный шип немного ниже антеродорсального угла. Киль плоский, сильно смещен вперед. Между килем и задним краем чешуи располагаются многочисленные, равномерно расположенные поры.
Погруженное поле широкое, занимает 1/2–1/3 длины чешуи (в редких случаях – около 1/4). На нем располагаются редкие поры. Передний край свободного поля прямой. Языковидные выступы расположены не на одной линии, имеют окончания различной формы (закругленные, прямые и острые), несут четко выраженные гребешки второго порядка. Разделяющие гребни борозды короткие (доходят не далее конца первой трети свободного поля), неправильной формы или узкие, в основном направлены горизонтально или с небольшим наклоном вниз. Гребни скульптуры короткие, сужаются кзади или не меняют свою ширину, достаточно многочисленные (до 20). Они могут быть весьма резкими, от треугольных до округлых в сечении. Иногда вдоль переднего края свободного поля присутствуют отдельные короткие, треугольные гребни, не соединяющиеся с остальной частью свободного поля. Задняя часть свободного поля несет два–три очень плоских, почти сливающихся с поверхностью вертикальных ряда зубцов; зубцы поcледнего ряда многочисленные (до 20), серратные. Несколько зубцов также расположено на задней, скошенной и закругленной части нижнего края. Число гребней и зубцов примерно совпадает и, вероятно, связано с возрастом индивида, которому принадлежала чешуя.
Зубы (изображения см. в: Pindakiewicz et al., 2020; Бакаев, 2020; Bakaev et al., in press) I. suchonensis грацильные, контрфорс мал, едва достигает основания коронарного расширения (угол между лингвальным краем и длинной осью зуба – около 10°–15°). Ближе к вершине коронка сильно уплощается и загибается лингвально сильнее, чем у других изученных морфотипов. Апикальная часть широкая, примерно в 2.5 раза шире шейки. Передний край зуба загнут лингвально, угол между остью рабочей поверхности и основанием составляет 70°–80°. Зубцы заостренные, несут характерные широкие, уплощенные режущие канты. Разделяющие их пространства широкие в верней части, но резко сужающиеся у основания зуба в тонкую щель. Акродиновый колпачок округлый в сечении, составляет не более половины высоты зубца. Рабочая поверхность, образуемая совокупностью зубцов, сильно рассечена и асимметрично выпуклая в латеральной проекции. Самым крупным является предпоследний зубец ряда. Следы прижизненного износа редки и слабо выражены. Площадки истирания выпуклые, а их края расположены как на лабиальной, так и на лингвальной стороне зубцов. Также некоторый износ наблюдается на режущих кантах.
Размеры. Длина рыбы, реконструируемая по голотипу СГУ № 104-Б/З-2 – примерно 15 см.
Сравнение. Данный вид отличается от всех видов Isadia: (зубы) удлиненно-конической (не уплощенной) формой зубцов и наличием хорошо развитых режущих кантов, начинающихся от оснований акродиновых колпачков и расширяющихся в базальном направлении; суженными до узких щелей промежутками между основаниями зубцов; образующими не более трети высоты зубца и имеющими округло-коническое окончания акродиновыми колпачками; сильно рассеченной рабочей поверхностью коронок челюстных зубов; (чешуи) тонкими чешуями, зубовидными гребнями скульптуры вдоль переднего края свободного поля, большей уплощенностью гребней свободного поля, лучшим развитием вертикальных рядов плоских, почти сливающихся с поверхностью зубцов. От I. aristoviensis вид по зубам дополнительно отличается более широкой коронкой, несущей большее количество зубцов. От I. arefievi вид по зубам отличается большим числом зубцов, большей грацильностью зубов, не развитым контрфорсом, сильно изогнутой рабочей поверхностью. От I. opokiensis вид по зубам дополнительно отличается большей грацильностью зубов, не развитым контрфорсом, сильно изогнутй рабочей поверхностью.
Распространение. Европейская часть России; верхнепермский (татарский) отдел, с основания северодвинского до кровли вятского яруса.
Материал. В колл. ПИН переданы образцы МГУ №№ МГУ № SU 32 KV 1s, KV 2s, KV 3s, KV 4s, KV 5s и KV 6s, получившие, соответственно, номера ПИН, №№ 5652/232, 230, 235, 239, 240, 231; Вологодская обл., Нюксеницкий р-н, местонахождение Кочевала-2; северодвинский ярус, верхнесеверодвинский подъярус, путятинский горизонт, полдарcская свита.
Isadia aristoviensis A. Minich, 1990
Табл. XIV , фиг. 7–12
Isadia aristoviensis: М. Миних, А. Миних, 1990, с. 96, табл. VII , фиг. 3–7 .
Amblypterina grandicostata: Есин, Машин, 1996, с. 87, рис. 5.5 -12а, табл. 5.5 -II, фиг. 5–7 .
Isadia suchonensis: А. Миних, М. Миних, 2009, с. 196, табл. 60 , фиг. 1–6 .
Голотип – СГУ, № 104-Б/Р-87, фрагмент тела рыбы со спинным и анальным плавниками; Оренбургская обл., Красногвардейский р-н, местонахождение Вязовка; верхнепермский (татарский) отдел, вятский ярус, кутулукская свита.
Описание (по Есин, Машин, 1996, с изменениями, для чешуй участка В). Высота чешуи участка В больше длины в 1.5–2 раза. Передний и задний края прямые; вентральный немного выпуклый, дорсальный немного вогнутый. Антеродорсальный угол на образцах не сохранился. Постеровентральный угол не резко скошен, закруглен, переход к заднему краю плавный. Сочленовный шип не сохранился, но, судя по строению ямки, был высоким. Погруженное поле широкое, занимает 1/2 длины чешуи. Передний край свободного поля прямой. Языковидные выступы имеют окончания различной формы (закругленные, острые), несут четко выраженные гребешки второго порядка. Разделяющие гребни борозды короткие, относительно широкие (но значительно у́же самих гребней), в основном направлены назад и вниз. Гребни скульптуры короткие, высокие, мощные, часто раздвоенные на переднем конце, округло-треугольные в сечении, представляют собой отдельные короткие, треугольные гребни, заостренные кончики которых нависают над поверхностью свободного поля. За первым рядом коротких гребней находится второй (иногда и третий). Последний ряд таких гребней образует до 10 серратных зубцов на заднем крае. Несколько зубцов также расположены на задней, скошенной и закругленной части нижнего края. Число гребней и зубцов примерно совпадает и, вероятно, связано с возрастом индивида, которому принадлежала чешуя.
Участок C. Чешуи толстые, их высота примерно равна или чуть больше длины. Передний и задний края прямые. Вентральный край слабовыпуклый, дорсальный – слабовогнутый. Антеродорсальный угол высокий, прямой, конец заострен. Антеровентральный угол скошен, немного закруглен, задненижний угол нерезко срезан или прямой. Сочленовный шип и ямка развитые, относительно высокие, расположены в середине длины. Сочленовный шип обычно достигает высоты переднего верхнего угла. Киль плоскоокруглый, немного смещен вперед. Погруженное поле занимает не более 1/3 длины чешуи. Передний край свободного поля прямой. Языковидные выступы расположены на одной линии, имеют окончания различной формы (закругленные и острые), несут гребешки второго порядка. Разделяющие гребни борозды короткие, узкие, глубокие, в основном направлены диагонально. Гребни скульптуры короткие, не меняют свою ширину, относительно малочисленны (до шести), часто раздваиваются на концах. Они могут быть округло-треугольными или округлыми в сечении. Задняя часть свободного поля гладкая.
Зубы тонкие (изображения см.: Pindakiewicz et al., 2020; Бакаев, 2020; Bakaev et al., in press). Контрфорс I. aristoviensis небольшой (угол между лингвальным краем и длинной осью зуба – около 15°–20°), но отчетливо выраженный, достигает основания коронарного расширения. Передний край зуба незначительно загнут лингвально, угол между остью рабочей поверхности и основанием составляет 80°–90°. Ближе к вершине коронка сильно уплощается и загибается лингвально. Апикальная часть примерно в 1.5 раза шире шейки. Зуб имеет обычно два или четыре широких лопатовидных зубца прямоугольной (при стирании колпачка) или округло-трапециевидной формы, с крупными акродиновыми колпачками, занимающими около половины высоты зубчиков. Зубцы отчетливо разделены и не смыкаются боковыми сторонами, сохраняя у оснований небольшой, но хорошо выраженный режущий кант. Рабочая кромка выпуклая. Крайние зубцы меньше средних.
Целый череп I. aristoviensis неизвестен, но, предположительно, верхнечелюстные зубы у данного вида имеют три-четыре зубца, в то время как у зубов нижней челюсти коронки бикуспидные. Площадки истирания плоские и, как правило, перпендикулярны продольной оси зуба. Почти все зубы несут выраженные следы износа в виде сколов и потертостей. Часто встречаются зубы со стертыми до основания акродиновыми колпачками, в результате чего формируется единая плоская поверхность. При такой степени износа стирание достигает не только дентиновых канальцев, но даже пульпарной полости.
Зубы на palatinum монокуспидные, тупые, конические, загнутые каудально. Акродиновый колпачок округлый, широкий, в сечении цилиндрический или немного сжатый в кранио-каудальном направлении. Под колпачком основание зуба заметно сужается, формируя перетяжку, а затем вновь образуя расширение. В некоторых случаях это расширение несет небольшие бугорки по бокам от акродинового колпачка.
Размеры. Длина рыбы, реконструируемая по голотипу СГУ, № 104-Б/Р-87 – примерно 40 см (Миних, Миних, 2009).
Сравнение. От всех видов рода Isadia этот вид отличается: (зубы) значительно более узкими коронками (апикальная часть не более чем в 1.5 раза шире шейки), акродиновым колпачком, составляющим не более половины высоты зубца, плоскими площадками износа на концах зубцов, ориентированными под прямым углом к продольной оси зубца; (чешуи) длинными, рельефными (округло-треугольными или треугольными в сечении), параллельными друг другу гребнями скульптуры, разделенными глубокими бороздами. По зубам дополнительно отличается: от I. opokiensis и I. arefievi – грацильностью зубов, слабо развитым контрфорсом; от I. opokiensis и I. suchonensis –меньшим числом зубцов; от I. arefievi – отсутствием увеличенного, шпателевидного зубца.
Распространение. Европейская часть России; верхнепермский (татарский) отдел, верхи северодвинского и вятский ярус.
Материал. В колл. ПИН переданы образцы МГУ №№ PR 203/2s-1, PR 203/2s-2, PR 203/2s-3, PR 203/2s-4, PR 203/2s-5 и PR 203/2s-6, получившие, соответственно, номера ПИН, №№ 5784/2, 11, 5, 8, 9, 12; Оренбургская обл., Сорочинский р-н, местонахождение Пронькино; верхнепермский (татарский) отдел, вятский ярус, кутулукская свита.
* * *
Автор благодарен Т.В. Кузнецовой и $\boxed{{\text{Е}}{\text{.К}}{\text{.}}\,\,{\text{Сычевской}}}$ за передачу сборов Д.Н. Есина, а также за ценную информацию. Данное исследование осуществлено при поддержке грантов РФФИ №№ 19-34-90040 и 21-54-10003. Работа выполнена за счет средств субсидии, выделенной Казанскому федеральному ун-ту для выполнения государственного задания № 671-2020-0049 в сфере научной деятельности.
Список литературы
Бакаев А.С. Новый морфотип зубов рыб отряда Eurynotoidiformes (Actinopterygii) из верхнепермских отложений Европейской России // Палеонтол. журн. 2020. № 2. С. 78–86.
Бакаев А.С. К ревизии пермских лучеперых рыб Европейской России. Часть 1 // Палеонтол. журн. 2022. № 5. С. 95–102.
Бакаев А.С. История изучения пермских костных рыб европейской части России // Учен. зап. Казан. ун-та. Сер. Естеств. науки. (в печати).
Голубев В.К. Новые виды Melosaurus (Amphibia, Labyrinthodontia) из казанских отложений Прикамья // Палеонтол. журн. 1995. № 3. С. 86–97.
Голубев В.К., Наумчева М.А. Новые данные о возрасте местонахождения пермских тетрапод Пронькино (Оренбургская область) // Палеострат-2019. Годичн. собр. (науч. конф.) секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва при РАН. М.: ПИН РАН, 2019. С. 23.
Гоманьков А.В., Голубев В.К., Есин Д.Н. и др. Палеонтологическая характеристика верхнепермских отложений местонахождения Шихово-Чирки (бассейн р. Вятка) // Бюлл. Региональной межведомственной стратиграфической комиссии по центру и югу Русской платформы. 1993. Вып. 2. С. 93–96.
Есин Д.Н. Чешуйный покров Amblypterina costata (Eichwald) и таксономия палеонисцид по изолированным чешуям // Палеонтол. журн. 1990. № 2. С. 89–96.
син Д.Н. Новые виды высокотелых актиноптеригий (Platysomidae) из верхней перми Восточно-Европейской платформы // Палеонтол. журн. 1993. № 3. С. 128–132
Есин Д.Н. Раннеказанские палеонисциды севера европейской части России и Прикамья // Палеонтол. журн. 1995а. № 2. С. 119–132.
Есин Д.Н. Позднепермские палеонисциды европейской части России. Дис. … канд. геол.-мин. наук. М., 1995б. 350 с.
Есин Д.Н., Машин В.Л. Ихтиолиты // Стратотипы и опорные разрезы верхней перми Поволжья и Прикамья / Ред. Есаулова Н.К., Лозовский В.Р. Казань: Экоцентр, 1996. С. 270–293.
Ефремов И.А., Вьюшков Б.П. Каталог местонахождений пермских и триасовых наземных позвоночных на территории СССР. М.: Изд-во АН СССР, 1955. 185 с.
Ивахненко М.Ф., Голубев В.К., Губин Ю.М. и др. Пермские и триасовые тетраподы Восточной Европы. М.: ГЕОС, 1997. 216 с. (Тр. Палеонтол. ин-та РАН. Т. 268).
Миних А.В., Миних М.Г. Новый отряд лучеперых из верхней перми Русской платформы. Саратов: Изд-во Сарат. ун-та, 1986. 23 с. (Деп. в ВИНИТИ 18.04.1986, № 2839-В86).
Миних А.В., Миних М.Г. Ихтиофауна перми Европейской России. Саратов: Изд. центр “Наука”, 2009. 244 с.
Миних М.Г. Новые высокотелые рыбы из верхнепермских отложений Восточно-Европейской платформы // Вопросы палеонтологии (Межвузовский научный сборник). СПб.: Изд-во СПб. ун-та, 1992. С. 137–146.
Миних М.Г., Миних А.В. Ревизия некоторых палеонисков и новые рыбы из верхней перми Восточно-Европейской платформы и возможности использования их в стратиграфии // Вопросы геологии Южного Урала и Нижнего Поволжья. Сб. науч. трудов / Ред. Алешечкин О.И. Саратов: Изд-во Сарат. ун-та, 1990. С. 84–104.
Международный кодекс зоологической номенклатуры. Изд. 4-е. М.: КМК, 2004. 223 с.
Молостовская И.И., Миних М.Г., Молостовский Э.А., Миних А.В. О выборе эталонного разреза татарских отложений стратотипической области // Проблемные вопросы региональной и местной стратиграфии фанерозоя Поволжья и Прикаспия. Матер. конф. Саратов: НВ НИИГГ, 2001. С. 38–39.
Стратотипический разрез татарского яруса на реке Вятке. М.: ГЕОС, 2001. 140 с. (Тр. Геол. ин-та РАН. Вып. 532).
Татарские слои реки Сухоны / Ред. Молостовский Э.А., Миних А.В. Саратов: Научн. книга, 2001. 204 с.
Aldinger H. Permisch Ganoidfische Oströnland // Medd. Grönland. 1937. V. 102. № 3. 392 p.
Bakaev A.S., Bulanov V.V., Kogan I. et al. Early ray-finned herbivores: the dental system of Eurynotoidiidae (Actinopterygii; Middle–Late Permian, European Russia) and its implications for palaeobiology and palaeoecology // Palaeontology. (in press).
Pindakiewicz M., Tałanda M., Sulej T. et al. New finds of teeth of the herbivorous actinopterygians from the latest Permian of East European Platform and feeding convergence among extinct and extant ray-finned fish // Acta Palaeontol. Pol. 2020. V. 65. № 1. P. 71–79.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал