

Палеонтологический журнал, 2022, № 6, стр. 92-104
Ископаемый каменный огарь (Tadorna petrina) и широконоска (Spatula praeclypeata sp. nov.) – древнейшие раннеплейстоценовые утиные (Aves: Anatidae) Крыма
Н. В. Зеленков *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: nzelen@paleo.ru
Поступила в редакцию 16.05.2022
После доработки 25.05.2022
Принята к публикации 26.05.2022
- EDN: CKTMCG
- DOI: 10.31857/S0031031X22060137
Аннотация
По материалам из пещеры Таврида (центральная часть Крымского полуострова; 1.9–1.5 млн л.) описаны древнейшие для Северного Причерноморья раннеплейстоценовые остатки гусеобразных птиц. Фрагментарный скелет пеганковой птицы (Tadornini) отнесен к ископаемому каменному огарю Tadorna petrina Kurochkin, 1985, первоначально описанному из верхнего плиоцена Забайкалья и морфологически схожему с современными огарями T. ferruginea. Это древнейшая находка этой филогенетической линии в Европе, проливающая свет на палеобиогеографическое распространение и эволюцию огарей в позднем кайнозое. Tadorna petrina рассматривается как наиболее вероятный стволовой представитель клады, включающей современные виды T. ferruginea, T. cana, T. tadornoides и T. variegata. Spatula praeclypeata sp. nov. из пещеры Таврида представляет собой древнейшую находку уток-широконосок в палеонтологической летописи и рассматривается как стволовой таксон по отношению к современным S. smithii, S. rhynchotis и S. clypeata. Фаунистическая ассоциация огаря и широконоски характерна для многих средне- и позднеплейстоценовых авифаун Европы; крымские находки свидетельствуют в пользу ее более древнего и, вероятно, восточного происхождения.
Раннеплейстоценовый (2.58–0.77 млн л.) этап эволюции авифаун Палеарктики, один из важнейших для понимания истории формирования современного таксономического разнообразия птиц этого обширного региона, остается одним из наименее изученных (Зеленков, 2013, 2014). Во всей Евразии местонахождения этого временного интервала с остатками птиц крайне редки, а известные авифауны представлены обычно небольшим числом таксонов. В Северном Причерноморье раннеплейстоценовые орнитокомплексы известны из нескольких местонахождений южной части Украины (например, Ногайск, Каиры, Тихоновка 2), датируемых преимущественно второй половиной калабрия (1.2–0.77 млн л. MQR7-8; Горобец, 2018; см. Тесаков, 2004). Более древние (позднегелазийские–раннекалабрийские) сообщества птиц, имеющие существенное значение для реконструкции эволюции позднекайнозойских авифаун региона, представлены только таксономически бедными и очень слабо изученными комплексами из местонахождений Крыжановка (окрестности Одессы) и Тарханкут (Северо-Западный Крым; Воинственский, 1967; Горобец, 2018; Зеленков, Горобец, 2020). В связи с этим большое значение имеет открытие в 2018 г. богатого местонахождения раннеплейстоценовых позвоночных в пещере Таврида в Центральном Крыму (Лопатин, 2019; Лопатин и др., 2019). Фауна пещеры Таврида включает в себя заметное разнообразие птиц, представляющее на сегодняшний день практически единственный источник сведений об экологическом и эволюционном облике авифауны Северного Причерноморья в калабрии. По крупным млекопитающим фауна пещеры была датирована интервалом 1.9–1.5 млн л., при этом существенное сходство с фауной местонахождения Дманиси в Грузии указывает на вероятный возраст около 1.8 млн л. (Лопатин и др., 2019). Ранее из пещеры Таврида был описан гигантский страус Pachystruthio dmanisensis; также было отмечено присутствие в местонахождении еще нескольких таксонов птиц (Zelenkov et al., 2019). В этой статье приведено описание собранных в 2020–2021 гг. материалов по утиным (Anseriformes: Anatidae), представляющих пеганковых (Tadornini) и речных уток (Anatini). Примечательна находка фрагментарного скелета ископаемого каменного огаря (Tadorna petrina Kurochkin, 1985) – это древнейшее указание на присутствие группы огарей в раннем плейстоцене Европы. Утка из Тавриды представлена ископаемой широконоской (Spatula praeclypeata sp. nov.) – это самая древняя находка данной группы специализированных утиных в палеонтологической летописи.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД ANSERIFORMES
СЕМЕЙСТВО ANATIDAE LEACH, 1820
Род Tadorna Boie, 1822
Tadorna petrina Kurochkin, 1985
Tadorna petrina: Курочкин, 1985, с. 94, табл. XV , фиг. 1, 2; Зеленков, Курочкин, 2015, c. 168.
Рис. 1.
Элементы крыла Tadorna petrina Kurochkin, 1985 в сравнении с локтевыми костями современных представителей рода Tadorna (локтевые кости масштабированы по размеру проксимальной суставной поверхности для иллюстрации пропорциональной укороченности кости у ископаемого вида): а, г, ж, з – Tadorna petrina Kurochkin, 1985, нижний плейстоцен пещеры Таврида (Крым): а, г – экз. ПИН, № 5644/221, правая локтевая кость; ж, з – экз. ПИН, № 5644/223, дистальный фрагмент левой лучевой кости; б, д – Tadorna ferruginea (Pallas, 1764), современный; в, е – Tadorna tadorna (Linnaeus, 1758), современный; а–в, з – вид с вентральной стороны; г–е – вид с краниальной стороны; ж – вид с дорсальной стороны. Обозначения: ol – олекранон; tc – tuberculum carpale. Длина масштабной линейки – 1 см.
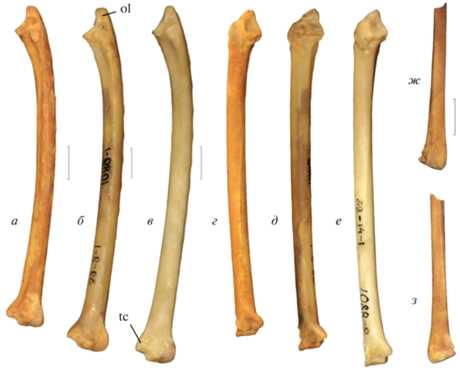
Рис. 2.
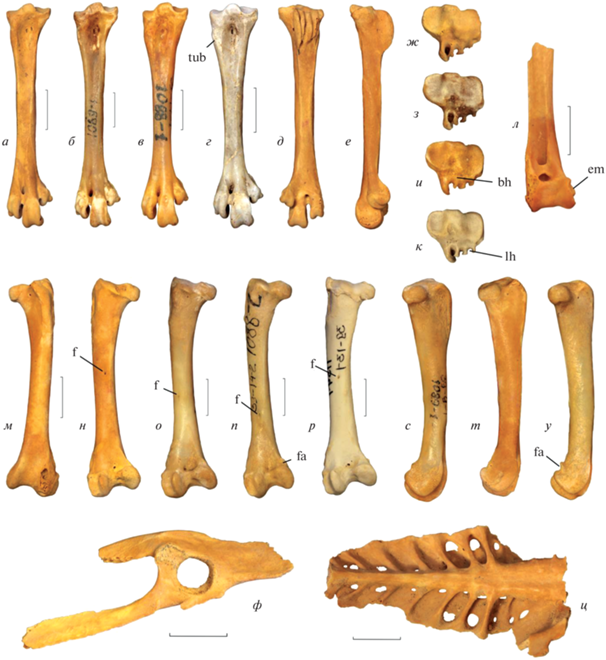
Tadorna petrina Kurochkin, 1985 в сравнении с современными и ископаемыми представителями рода Tadorna: а, д, е, ж, л, м, н, т, ф, ц – Tadorna petrina Kurochkin, 1985, нижний плейстоцен пещеры Таврида (Крым): а, д, е, ж – экз. ПИН, № 5644/227, правый тарсометатарсус; л – экз. ПИН, № 5644/226, дистальный фрагмент левого тибиотарсуса; м, н, т – экз. ПИН, № 5644/224, левая бедренная кость; ф – экз. ПИН, № 5644/209, фрагмент левого таза; ц – экз. ПИН, № 5644/230, фрагмент синсакрума; б, з, о, с – Tadorna ferruginea (Pallas, 1764), современный: б, з – тарсометатарсус; о, с – бедренная кость; в, и, п, у – Tadorna tadorna (Linnaeus, 1758), современный: в, и – тарсометатарсус; п, у – бедренная кость; г, к – Tadorna tadorna (Linnaeus, 1758), экз. ПИН, 2613/53, тарсометатарсус; местонахождение Черноусов лог, Республика Хакассия, Россия; средний голоцен; р – Tadorna tadornoides (Jardine et Selby, 1828), современный, бедренная кость; а–г – вид с дорсальной стороны; д – вид с плантарной стороны; е, с–у – вид с медиальной стороны; ж–к – вид с проксимальной стороны; л, м – вид с краниальной стороны; н–р – вид с каудальной стороны; ф – вид с латеральной стороны. Обозначения: bh – основание гипотарсуса; em – epicondylus medialis; f – питательное отверстие; fa – площадка в дистальной части crista supracondylaris medialis; lh – латеральный желоб гипотарсуса; tub – утолщение в проксимо-латеральной части стержня тарсометатарсуса. Длина масштабной линейки – 1 см.
Голотип – ПИН, № 2975/2, фрагментарная грудина; Россия, республика Бурятия, местонахождение Береговая; верхний плиоцен, чикойская свита.
Описание (рис. 1, а, г, ж, з; 2, а, д, е, ж, л, м, н, т, ф, ц). В грудине spina interna приподнятая, с хорошо выраженной выемкой посередине; между боковым краем этой spina и началом labrum internum sternae (Livezey, Zusi, 2006) имеется достаточно широкая борозда; incisurae costales глубокие, processus articularis sternocostalis между ними высокие, в целом ряд реберных вырезок длинный (Курочкин, 1985, с изменениями). Локтевая кость несколько укорочена; olecranon короткий и утолщенный при виде с вентральной стороны; tuberculum carpale резко выдается медиально относительно стержня и имеет закругленную вершину. В бедренной кости crista supracondylaris medialis не формирует выраженной площадки непосредственно проксимальнее condylus medialis (в его медиальном профиле отсутствует выраженная вырезка); impressiones ansae m. iliofibularis хорошо выражены, проксимальный отпечаток выпуклый; питательное отверстие на каудальной поверхности стержня сдвинуто заметно проксимально (располагается проксимальнее середины стержня). В тарсометатарсусе гипотарсус имеет низкое основание, в результате чего каналы гипотарсуса приближены к плантарному краю cotylae; дно латерального желоба гипотарсуса сдвинуто плантарно относительно дна центрального желоба гипотарсуса; сам гипотарсус сдвинут латерально; выраженное утолщение латерального края кости вблизи проксимального эпифиза отсутствует. Размеры – с мелких представителей современного T. ferruginea.
Размеры в мм. Локтевая кость (экз. ПИН, № 5644/221): наибольшая длина – 97.7; наименьшая дорсовентральная ширина стержня – 5.4; дорсовентральная ширина проксимального эпифиза – 10.0; диагональная ширина проксимального эпифиза (от вершины olecranon до дорсального угла cotyla dorsalis) – 12.5; диагональная ширина дистального эпифиза (от вершины tuberculum carpale до вершины condylus dorsalis) – 11.8. Лучевая кость: ширина дистального эпифиза – 7.8; максимальная высота дистального эпифиза – 4.0. Бедренная кость: максимальная длина – 52.8; максимальная ширина проксимального эпифиза – 12.9; минимальная ширина стержня – 5.1; максимальная ширина дистального эпифиза – 12.8. Тибиотарсус: минимальная ширина стержня – 4.9; высота condylus medialis вместе с медиальным каудальным гребем – 10.5; краниокаудальная высота incisura intercondylaris – 6.6. Тарсометатарсус: максимальная длина – 56.7 (экз. ПИН, № 5644/227), 56.6 (экз. ПИН, № 5644/228); ширина проксимального эпифиза – 11.3 (оба экз.); высота проксимального эпифиза до вершины crista medialis hypotarsi – 11.0 (экз. ПИН, № 5644/227), 10.8 (экз. ПИН, № 5644/228); минимальная ширина стержня – 4.8 (экз. ПИН, № 5644/227), 4.7 (экз. ПИН, № 5644/228); максимальная ширина дистального эпифиза – 12.2 (экз. ПИН, № 5644/227), 12.1 (экз. ПИН, № 5644/228); высота trochlea metatarsi III – 7.2 (оба экз.). Размеры голотипа – см. Курочкин (1985).
Сравнение. Грудина T. petrina (см. Курочкин, 1985, табл. XV , фиг. 1, 2) от таковой изученных современных видов рода (T. radjah, T. tadorna, T. ferruginea, T. tadornoides, T. variegata) отличается хорошо выраженной бороздкой, располагающейся вентролатеральнее spina interna, глубокими incisurae costales (их глубина приблизительно равна их ширине), высокими processus articularis sternocostalis и, в целом, удлиненным реберным рядом. У современных видов incisurae costales либо совсем невогнутые, либо очень мелкие, весь реберный ряд короче, spina interna с неглубокой вырезкой посередине, а борозда, расположенная латеральнее этой spina, не так четко выражена. Как было замечено ранее (Курочкин, 1985), эта борозда лучше выражена у T. ferruginea и T. tadornoides. С T. ferruginea и T. tadornoides грудина T. petrina также сходна морфологией центральной (медиальной) части labrum externum sternae при виде с вентральной стороны: у огарей она не резко выступает краниально относительно латеральной части губы (резко – у T. tadorna).
Локтевая кость несколько укорочена по сравнению с таковой у современных T. tadorna и T. ferruginea при сохранении общих пропорций проксимального эпифиза и стержня (рис. 1). Olecranon при виде с вентральной стороны несколько короче, чем у современных представителей рода. Tuberculum carpale резко выдается медиально относительно стержня, как у T. ferruginea, но в отличие от T. tadorna, у которых этот бугорок полого переходит на стержень; его вершина более округлая, чем у современных видов.
Бедренная кость отличается от таковой T. tadorna отсутствием площадки (рис. 2, fa) в дистальной части crista supracondylaris medialis, которая хорошо развита у T. tadorna и при виде с медиальной стороны формирует вырезку. У T. ferruginea и других представителей рода эта площадка выражена менее явно. Impressiones ansae m. iliofibularis выпуклые и хорошо выраженные, в отличие от T. tadorna, где они неявные или практически не выступают относительно уровня стержня. Питательное отверстие (рис. 2, f), располагающееся у огаря из Тавриды в проксимальной половине стержня, у T. tadorna располагается заметно дистальнее. У T. ferruginea оно располагается примерно в центре стержня, т.е. проксимальнее, чем у T. tadorna, но заметно дистальнее, чем у T. petrina. Сходное расположение питательного отверстия обнаружено у T. tadornoides, который также проявляет сходство с ископаемым видом в строении crista supracondylaris medialis и в выраженности impressiones ansae m. iliofibularis.
В дистальном тибиотарсусе характерно наличие хорошо выраженного epicondulys medialis (рис. 2, em), как у T. ferruginea, но в отличие от T. tadorna. Pons supratendineus ориентирован субперпендикулярно длинной оси кости – в большей степени, как у T. ferruginea, в то время как у T. tadorna этот мостик ориентирован более косо.
В строении тарсометатарсуса отличается от T. tadorna более низким основанием гипотарсуса, особенно в его медиальной части (рис. 2, bh). В результате этого каналы гипотарсуса в значительной степени приближены к плантарному краю cotylae, как у T. ferruginea. Кроме того, дно латерального желоба гипотарсуса (рис. 2, lh) у T. tadorna располагается примерно на одном уровне с дном центрального желоба, в то время как T. ferruginea и T. petrina дно латерального гребня выраженно смещено дорсальнее, т.е. располагается глубже в теле гипотарсуса. У T. petrina и T. ferruginea гипотарсус смещен латерально, в то время как у T. tadorna он занимает большую часть ширины проксимального эпифиза. Для подавляющего большинства экз. T. tadorna также характерно наличие крупного утолщения в проксимо-латеральной части стержня (рис. 2, tub). Различий между видами Tadorna в строении дистального эпифиза тарсометатарсуса не выявлено.
Замечания. Фрагментарный скелет пеганковой птицы из пещеры Таврида морфологически сходен с современными огарем T. ferruginea и надежно отличается от пеганок T. tadorna (см. выше). Устойчивость выявленных отличий в строении тарсометатарсуса подтверждается сходством между использованными для сравнения современными и среднеголоценовыми T. tadorna (рис. 2). При этом находка из Тавриды находится на нижнем пределе размерной изменчивости T. ferruginea (Woelfle, 1967). Так, большинство исследованных и опубликованных экземпляров T. ferruginea заметно крупнее по размерам (например, Woelfle, 1967), однако один экз. в коллекции лаборатории исторической экологии Института проблем экологии и эволюции имени А.Н. Се́верцова РАН (ИПЭЭ РАН) (экз. № 1487) по длине тарсометатарсуса идентичен таковому из пещеры Таврида. Минимальная длина плюсны современных представителей вида (54–57 мм у наименьших экз. обоих полов; Тугаринов, 1941) также соответствует длине тарсометатарсуса у ископаемого экземпляра.
Огарь из пещеры Таврида может быть надежно отличен от сходных по размеру мелких представителей Anserinae (например, мелких представителей рода Branta), которые отличаются более удлиненным тарсометатарсусом с более расставленными trochleae metatarsorum, расширенным в своем основании trochlea metatarsi III при виде с дорсальной стороны, заметно более низкими гребнями гипотарсуса, сильно вытянутым проксимодистально crista medialis hypotarsi, а также наличием явной вогнутости в проксимолатеральной поверхности кости на уровне fossa infracotylaris. В строении локтевой кости Tadornini могут быть уверенно определены на основании ориентации tuberculum carpale относительно проксимального эпифиза: у Tadornini вершина отростка ориентирована краниально, в то время как у сходных по размерам Mergini он ориентирован заметно более вентрально и более робустный. У филогенетически более продвинутых Anatinae (таких как Cairina) tuberculum carpale заметно более мелкий.
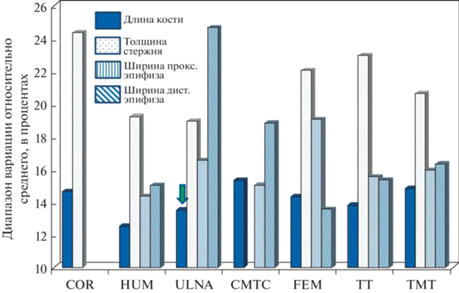
Ископаемый огарь из Тавриды характеризуется укороченной локтевой костью при сходных с современными T. ferruginea размерах сочленовных поверхностей и толщины стержня кости (рис. 2). Соотношение “длина кости/минимальная толщина стержня” у ископаемого крымского огаря меньше, чем у всех исследованных современных Tadorna (с учетом данных из работы: Poland, 2018), а значение коэффициента “длина кости/косой диаметр проксимального эпифиза”, наоборот, превышает таковое у современных представителей рода (рис. 3). По нашим данным, длина плечевой и локтевой костей – параметры, подверженные наименьшей индивидуальной изменчивости у уток (рис. 4; см. Zelenkov, 2019), поэтому наблюдаемое отклонение в пропорциях указывает на явную морфологическую специфику раннеплейстоценового крымского огаря и, с учетом его геологического возраста и отличий в строении бедренной кости, свидетельствует также о его видовой специфичности. Согласно молекулярным оценкам (Fulton et al., 2012; Sun et al., 2017), дивергенция современного T. ferruginea и сестринского южноафриканского T. cana датируется средним плейстоценом (около 600 тыс. л.). Таким образом, более древние раннеплейстоценовые популяции евроазиатских огарей заслуживают отнесения к отдельному ископаемому виду.
Рис. 3.
Соотношение пропорций локтевой кости у ископаемого Tadorna petrina (красный квадрат), современных T. ferruginea (желтый треугольник) и современных T. tadorna (голубой круг; данные из работы: Poland, 2018).
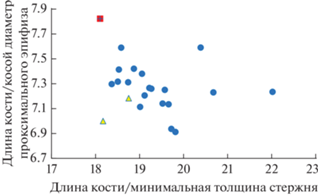
Рис. 4.
Индивидуальная изменчивость основных параметров посткраниального скелета Anatidae на примере европейских речных уток (Anas s.l.). По оси абсцисс: элементы скелета. По оси ординат: сумма максимальных показателей отклонения в большую и меньшую стороны относительно среднего для каждого вида, в процентах (приведен средний показатель для всех изученных видов). Обозначения: CMTC – карпометакарпус; COR – коракоид; FEM – бедренная кость; HUM – плечевая кость; TMT – тарсометатарсус; TT – тибиотарсус; ULNA – локтевая кость. Стрелкой обозначена невысокая относительная изменчивость общей длины локтевой кости (см. текст).
Находки огарей в плиоцене–раннем плейстоцене единичны и ограничены ископаемым T. petrina из позднего плиоцена местонахождения Береговая (верхи зоны MN 16b; около 2.6 млн л.; Ербаева и др., 2005; Erbajeva, Alexeeva, 2013) в Забайкалье (Курочкин, 1985), а также формами, отнесенными к современному виду T. ferruginea, из раннего плейстоцена Италии (~1.4 млн л.; Bedetti, Pavia, 2013) и Турции (~0.9–1.0 млн л.; Louchart et al., 1998). Здесь принимается конспецифичность T. petrina и раннеплейстоценового крымского огаря на основании следующих соображений. Во-первых, продолжительность существования видов птиц, как следует из многочисленных молекулярных оценок, подкрепленных палеонтологической летописью, оценивается от нескольких сотен тысяч до 1.5–2 млн лет и даже более (обзор см.: Зеленков, 2014; Паластрова, 2022). Таким образом, разница в 700–900 тыс. л. между забайкальской и крымской находками не противоречит их возможной конспецифичности. Во-вторых, поздневиллафранкское время, к которому относится фауна пещеры Таврида, характеризуется в Юго-Восточной Европе иссушением климата и значительным распространением ксерофитных ландшафтов (Naidina, Richard, 2016), что сопровождалось расселением в регион Северного Причерноморья азиатских фаунистических элементов, в том числе птиц (Лопатин и др., 2019; Zelenkov et al., 2019). Ряд найденных в Тавриде таксонов млекопитающих имел широкое распространение в Евразии в раннем плейстоцене (Лопатин, 2019). В-третьих, чикойский фаунистический комплекс, откуда описан T. petrina, имеет общие виды неворобьиных птиц – обитателей открытых ландшафтов (например, Perdicini и Coturnicini из курообразных) – с плио-плейстоценовыми местонахождениями Восточной Европы (Зеленков, Курочкин, 2009, 2015). В-четвертых, современный огарь T. ferruginea имеет обширный ареал, простирающийся от Марокко до Восточной Монголии (а на зимовке – вплоть по побережья Тихого океана в Восточном Китае). Нет сомнений, что широкое распространение могли иметь и плио-плейстоценовые предшественники современных огарей. В дополнение к этому, общее остеологическое однообразие представителей рода Tadorna едва ли позволит различать дополнительные виды в рамках эволюции группы на границе плиоцена и плейстоцена (представители рода, остеологически сходные с современными, регистрируются уже в конце раннего–среднем миоцене; Зеленков, 2019).
При первоописании T. petrina не было обосновано отнесение этого вида к трибе Tadornini и роду Tadorna, в частности (Курочкин, 1985). При этом по размерам голотип T. petrina соответствует не только представителям рода Tadorna, но также некрупным Branta и самым мелким современным представителям рода Anser (например, A. erythropus). Переизучение голотипа этого вида подтвердило его таксономическую принадлежность. Отнесение данной формы к Tadornini основано на присутствии хорошо выраженной spina interna с выемкой в ее центральной части. У других утиных spina interna совсем или практически не выражена. Полное отсутствие пневматизации вдоль краниального края внутренней (кардиальной) поверхности грудины надежно отличает T. petrina от представителей рода Branta. У Anser пневматизация отсутствует, как и у T. petrina, но при этом spina interna имеет вид бугорка без вырезки в центре, а реберный ряд при виде с дорсальной стороны более расширен в своей средней части. Наличие борозды между spina interna и началом labrum internum sternae позволяет относить экземпляр к роду Tadorna, поскольку только у этого рода проявляется этот признак. Глубокие краниальные incisurae costales характерны также для Alopochen, но у этого рода упомянутая борозда не выражена, а реберный ряд короткий. Также удается подтвердить (см. выше) самостоятельный видовой статус T. petrina и отнесение его к группе огарей (T. ferruginea, T. cana, T. tadornoides и T. variegata).
Кажущаяся морфологическая дистинктивность грудины T. petrina по сравнению с другими известными элементами скелета, очевидно, связана с выраженной структурной изменчивостью грудной кости по сравнению с длинными костями конечностей. С другой стороны, отличные пропорции локтевой кости T. petrina (по материалам из пещеры Таврида), указывающие на несколько специфичное строение летательного аппарата у плио-плейстоценовых огарей, могут объяснять и несколько иное строение грудины как основы для крепления летательной мускулатуры.
Материал. Фрагментарный расчлененный скелет, в составе: экз. ПИН, №№ 5644/221, полная правая локтевая кость; 5644/222, проксимальный фрагмент левой локтевой кости; 5644/223, дистальный фрагмент правой лучевой кости; 5644/212, краниальный фрагмент синсакрума с подвздошными костями; 5644/229, фрагментарный правый таз; 5644/230, фрагментарный синсакрум; 5644/224, полная левая бедренная кость; 5644/225, проксимальный фрагмент левого тибиотарсуса; 5644/226, дистальный фрагмент правого тибиотарсуса; 5644/227, полный правый тарсометатарсус; 5644/228, полный левый тарсометатарсус; 5644/213, базальная фаланга третьего пальца стопы – пещера Таврида; окрестности пос. Зуя, Белогорский район, Крым; костеносный слой BL (см. Оксиненко, Лавров, 2021), поздний виллафранк, калабрий, нижний плейстоцен.
Род Spatula Boie, 1822
Spatula praeclypeata Zelenkov, sp. nov.
Название вида – от clypeata – современное видовое название обыкновенных широконосок.
Голотип – ПИН, № 5644/220, правый коракоид; местонахождение пещера Таврида; Крым, Белогорский р-н, окрестности пос. Зуя, пещера Таврида; нижний плейстоцен, калабрий, поздний виллафранк, костеносный слой BL (см. Оксиненко, Лавров, 2021).
Описание (рис. 5, в, е–и). Cotyla scapularis расположен в латеральной части стержня и маленький, его диаметр не превышает половину ширины стержня при виде с дорсальной стороны; crista acrocoracoidea ориентирован примерно на 45 градусов относительно длинной оси кости, короткий (заметно короче facies articularis humeralis) и тонкий; sulcus m. supracoracoidei с хорошо очерченным глубоким углублением в дорсальной части, над которой не нависает facies articularis clavicularis; вентральная часть sulcus m. supracoracoidei формирует утолщенный валик, сливающийся краниально с вентральными двумя третями facies articularis clavicularis; processus procoracoideus с выпуклым медиокаудальным краем; facies articularis clavicularis с рецессией в центральной части; стернальный край кости умеренно скошен в своей латеральной части (не перпендикулярен длинной оси кости). Пропорции стержня удлиненные.
Рис. 5.
Коракоид Spatula praeclypeata sp. nov. и избранных современных Anatidae: а, б, к – Spatula clypeata (Linnaeus, 1758), современный; в, е–и – Spatula praeclypeata sp. nov., голотип ПИН, № 5644/220; пещера Таврида, Крым; калабрий, нижний плейстоцен; г – Mareca penelope (Linnaeus, 1758), современный; д – Aythya fuligula (Linnaeus, 1758), современный; а–д – вид с дорсальной стороны; е – вид с вентральной стороны; ж – вид с латеральной стороны; з – вид с дорсомедиальной стороны; и, к – вид с краниовентральной стороны на processus acrocoracoideus. Обозначения: ca – crista acrocoracoidea; cs – cotyla scapularis; dep – углубление в дорсальной части sulcus m. supracoracoidei; fac – facies articularis clavicularis; ila – impressio lig. acrocoracohumeralis; pp – processus procoracoideus. Длина масштабной линейки – 1 см.
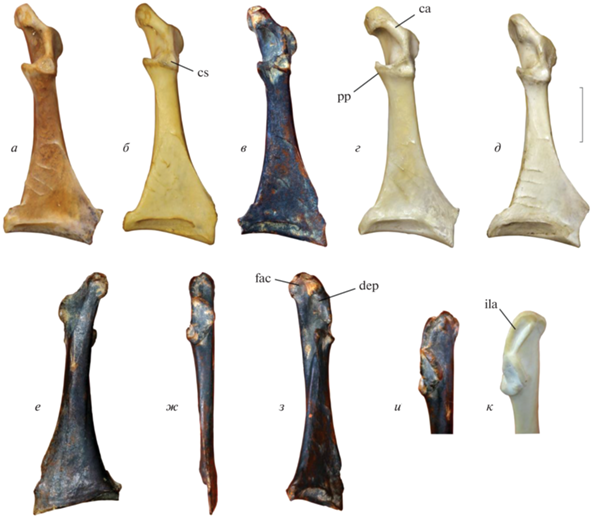
Размеры в мм. Медиальная длина – 42.6; длина от краниального края до каудального края cotyla scapularis – 14.9; максимальная дорсовентральная высота facies articularis humeralis – 5.2; минимальная ширина стержня – 4.7.
Сравнение. Отличается от S. clypeata, широко представленных в Евразии со среднего плейстоцена по современность, заметно меньшим и смещенным латерально cotyla scapularis, выпуклым медиокаудальным краем processus procoracoideus, а также отчетливо изогнутым impressio lig. acrocoracohumeralis. У S. clypeata cotyla scapularis заметно более крупный и при виде с дорсальной стороны занимает значительную часть стержня, impressio lig. acrocoracohumeralis у S. clypeata обычно прямое или несколько изогнутое, но без выраженного перегиба, характерного для S. praeclypeata.
Замечания. Полный правый коракоид из пещеры Таврида принадлежит средней по размерам утке, метрически сходной с современными видами Aythya fuligula, Aix galericulata, Spatula clypeata и Mareca penelope. У Aythya, в отличие от описываемого экземпляра, processus acrocoracoideus выровнен с продольной осью кости, а плоскость этого отростка наклонена относительно дорсальной плоскости кости (т.е. processus acrocoracoideus ориентирован более вертикально; см. Worthy, Lee, 2008: признак 45). Crista acrocoracoidea (Elzanowski et al., 2012) у Aythya более параллелен длинной оси кости, а angulus medialis заметно крупнее, чем у описываемого экземпляра. Для Mareca характерен умеренно вогнутый по всей своей площади sulcus m. supracoracoidei, несколько укороченный стержень, более медиально выдающийся facies articularis clavicularis и обычно медиально выдвинутый processus acrocoracoideus – все эти признаки не прослеживаются у голотипа ПИН, № 5644/220. Наиболее морфологически схожи по строению коракоида представители рода Aix и Spatula clypeata, однако у Aix facies articularis clavicularis всегда нависает над отчетливо выемчатым sulcus m. supracoracoidei, в то время как у Spatula выраженное углубление имеется только в дорсальной части sulcus m. supracoracoidei (рис. 5, dep), при этом оно не заходит краниально под facies articularis clavicularis, и в результате последняя не нависает над ней. У S. clypeata вентральнее вышеупомянутого углубления в sulcus m. supracoracoidei имеется расширение медиальной кромки стержня (валик), подстилающее значительную часть каудального края facies articularis clavicularis.
Несмотря на общее структурное сходство с современными S. clypeata, голотип ПИН, № 5644/220 демонстрирует выраженные отличия в строении cotyla scapularis и impressio lig. acrocoracohumeralis, которые, наряду с геологическим возрастом находки и представлениями о времени дивергенции современных видов, подтверждают отдельный видовой статус широконоски из пещеры Таврида. Согласно различным молекулярным оценкам (Fulton et al., 2012; Mitchell et al., 2014; Sun et al., 2017), расхождение современных голарктических широконосок S. clypeata и близких видов (австралийской широконоски S. rhynchotis и африканской широконоски S. smithii) датируется от 1.0 до 0.5 млн л., то есть самым концом раннего–второй половиной среднего плейстоцена. Таким образом, более древние раннеплейстоценовые представители этой филогенетической линии не могут быть помещены в современный вид S. clypeata. Включение стволовых представителей обсуждаемой клады в один из современных видов противоречит филогенетической концепции вида и, что немаловажно, будет служить ошибочной калибровочной точкой в эволюционных исследованиях, базирующихся на данные палеонтологии.
Материал. Только голотип.
ОБСУЖДЕНИЕ
Обнаруженные в пещере Таврида остатки утиных имеют важное палеобиогеографическое значение, поскольку для широконосок (клада, включающая голарктический вид Spatula clypeata) это древнейшая находка в палеонтологической летописи глобально, а для огарей из группы T. ferruginea – древнейшая находка в Европе. Более древние позднемиоценовые находки “Anas” clypeata из Венгрии и Украины (Соболев, 2004; Kessler, 2009) были недавно отнесены к ископаемому виду Anas kurochkini Zelenkov et Panteleyev, 2015, метрически сходному с широконоской, но морфологически – отчасти с современными A. platyrhynchos (Zelenkov, Panteleyev, 2015) и, таким образом, представляющему отдельную филогенетическую линию речных уток. Отличия A. kurochkini от современных речных уток и широконосок, в частности, отмеченные в первоописании (Zelenkov, Panteleyev, 2015), могут также быть дополнены укороченным стержнем. Форма, близкая к A. kurochkini, также была указана из позднего миоцена местонахождения Шкодова гора под Одессой (Ковальчук и др., 2017).
Согласно молекулярным оценкам, группа специализированных фильтраторов широконосок, включающая, помимо S. clypeata, также южноамериканскую широконоску S. platalea, австралийскую широконоску S. rhynchotis и капскую широконоску S. smithii, дивергировала только в раннем плейстоцене, около 2.6 млн л. н. (Fulton et al., 2012), 2.2 млн л. н. (Sun et al., 2017) или даже позднее, около 1.7 млн л. н. (Mitchell et al., 2014). В палеонтологической летописи широконоски появляются в калабрии Европы. Древнейшие ранее известные находки были указаны из местонахождений Пьетрафитта (~1.6 млн л.; Masini, Sala, 2011) и Пирро Норд (“Пирро-дэль-Эрба”; ~1.35 млн л.; Masini, Sala, 2011) в Италии (Tyrberg, 2008; Bedetti, Pavia, 2013), Хрустальной пещеры Беременд в Венгрии (первоначально называвшегося “Беременд 16”; Janossy, 1992), датируемой 1.5–1.2 млн л. (Pazonyi et al., 2019), местонахождения Бетфия-9 в Румынии, датируемого около 1.2 млн л. (Kessler, 2013; см. Terzea, 1996), а также несколько более древнего (1.3 млн л.) местонахождения Бетфия-2 (Tyrberg, 2008; Kessler, 2013). Древнейшая находка широконоски в Западной Европе относится к местонахождению Хуескар-1 (Huescar-1) на юге Испании, датируемому 0.9 млн л. (Sanchez Marco, 1989; см. Gibert et al., 2007). Примерно этому возрасту (0.9–0.7 млн л.; Тесаков, 2004) соответствует и единственная ранее известная раннеплейстоценовая находка вида в Причерноморье, происходящая из Морозовки-1 (“Черевичное-1”) в окрестностях Одессы (Воинственский, 1967). В центральной Турции широконоска известна из местонахождения Дурсунлу, также датируемого 1.0–0.9 млн л. (Louchart et al., 1998).
Южноамериканский вид S. platalea – наиболее базальная форма в кладе широконосок, поведенчески и морфологически близкая к внешней группе и, в частности, голубокрылым чиркам S. cyanoptera (Johnsgard, 2010); филогенетическое положение S. platalea в составе клады широконосок реконструируется с затруднениями (Nelson et al., 2017). При этом географическое распространение S. platalea и сестринской для широконосок группы чирков S. discors и S. cyanoptera уверенно указывает на то, что клада широконосок имеет американское (возможно – неотропическое) происхождение (Johnsgard, 2010). Spatula praeclypeata представляет собой древнейшую находку группы в палеонтологической летописи и, по всей видимости, относится к первой волне расселения широконосок в Старом Свете. Реликтом раннечетвертичных старосветских широконосок может оказаться морфологически примитивная (см. Livezey, 1991) и поведенчески наиболее близкая к S. platalea (Johnsgard, 2010) южноафриканская широконоска S. smithii, часто реконструируемая как таксон, сестринский к кладе S. rhynchotis + S. clypeata (Fulton et al., 2012; Mitchell et al., 2014).
Было показано, что широконоски, обладающие высокоспециализированным фильтрационным челюстным аппаратом (Kooloos et al., 1989), более эффективно фильтруют мелкие пищевые частицы (диаметром до 1.5 мм) только в присутствии непотребляемого детрита (Gurd, 2007). В том случае, если детрит отсутствует, эффективность работы их челюстного аппарата оказывается такой же, как у менее специализированной кряквы (там же). Таким образом, пищевая специализация широконосок, заключающаяся в способности крайне эффективно извлекать мелкие пищевые частицы из детритной смеси, указывает на становление группы в условиях водоемов, обедненных макро-органикой. Предки этих птиц, по-видимому, были вынуждены находить пищу в условиях, где фильтрация обычным (плезиоморфным) типом челюстного аппарата оказывалась неэффективной. В связи с этим появление широконосок в палеонтологической летописи в раннем плейстоцене позволяет предполагать, что их эволюционное формирование могло быть связано с нарастающей аридизацией климата и падением уровня Мирового океана в гелазии (Miller et al., 2020). Примечательно, что широконоски становятся очень обычными в палеонтологической летописи по всей Европе в среднем плейстоцене (Tyrberg, 1998, 2008) – в условиях общего похолодания климата их способность к эффективному питанию в относительно олиготрофных водоемах могла оказаться существенной и в конечном счете определить их последующий эволюционный успех и широкое распространение в “холодных” фаунах позднего плейстоцена и современности.
Как и широконоска, огарь T. ferruginea также обычен в среднем плейстоцене Европы (Tyrberg, 2008), но более древние раннеплейстоценовые находки этого или близких видов единичны и ограничиваются материалами из местонахождений Пирро Норд в Италии (~1.35 млн л.; Bedetti, Pavia, 2013) и Дурсунлу (1.0–0.9 млн л.) в Центральной Турции (Louchart et al., 1998). Tadorna sp. также указан из раннего плейстоцена Тибета (Stidham et al., 2015), но эта находка также может представлять и пеганку (T. tadorna) – более древнюю и более обычную в палеонтологической летописи, известную уже из пограничных плиоцен-плейстоценовых отложений Европы и Северной Африки (Mourer-Chauviré, 1993; Mourer-Chauviré, Geraads, 2010). Группа огарей (T. ferruginea и близкие виды), согласно молекулярным оценкам, дивергировала относительно других Tadornini на рубеже миоцена и плиоцена или в плиоцене, а расхождение старосветских видов T. ferruginea и T. cana оценивается концом раннего–средним плейстоценом (1.0 или 0.6 млн л.; Fulton et al., 2012; Sun et al., 2017). Таким образом, позднеплиоцен–раннеплейстоценовый T. petrina, скорее всего, является по меньшей мере предком современных T. ferruginea и T. cana. Однако если представления о времени дивергенции старосветских T. ferruginea и T. cana относительно австралийского T. tadornoides верны и составляют 1.8–1.4 млн л. (Fulton et al., 2012; Sun et al., 2017), то T. petrina, как наиболее древний известный представитель этой филогенетической линии, может также оказаться предковой формой и для более инклюзивной клады, включающей T. ferruginea, T. cana, T. tadornoides и новозеландского T. variegata. Лопатка из раннего–среднего плиоцена Австралии была предварительно отнесена к T. tadornoides (как единственному представителю рода в Австралии) на основании исключительно размеров (Worthy, 2008) – это единственное возможное указание на присутствие этой группы в Австралийском регионе в плиоцене. Однако не только видовая, но также и родовая принадлежность этого экземпляра едва ли может быть подтверждена, ввиду низкой диагностичности лопатки у утиных. Поскольку других находок Tadorna в позднем миоцене–раннем плейстоцене Австралии не известно (см. Vickers-Rich et al., 1991; Boles, 2006; Worthy, Nguyen, 2020), можно предположить, что ранняя эволюция обсуждаемой филогенетической линии Tadornini могла иметь место в Евразии, где похожие на современных представители трибы известны уже с конца раннего миоцена (Зеленков, 2019). Проникновение огарей, родственных T. ferruginea и населяющих исключительно открытые степные и луговые местообитания, из Центральной Азии в Австралию и Новую Зеландию может объясняться формированием коридора из открытых саванных ландшафтов в Юго-Восточной Азии (в частности, на островах Зондского архипелага) в раннем–среднем плейстоцене (Meijer et al., 2015; Louys, Roberts, 2020).
Совместное присутствие огаря T. ferruginea и широконоски Spatula clypeata характеризует многие среднеплейстоценовые местонахождения с территории Европы (Tyrberg, 1998, 2008), но для раннего плейстоцена эта ассоциация была выявлена только в несколько более молодом местонахождении Пирро Норд (“Пирро-дэль-Эрба”) в Центральной Италии, откуда реконструируется репрезентативная фауна птиц, включающая значительное число обитателей открытых луговых или степных ландшафтов (Bedetti, Pavia, 2013). Преобладание открытых саванноподобных ландшафтов также характерно и для окрестностей пещеры Таврида (Лопатин, 2019; Zelenkov et al., 2019), где обнаружены формы, близкие к вышеуказанным современным видам. При этом если широконоска в настоящее время населяет разнообразные местообитания в умеренном поясе Евразии (в том числе, степные), то огари в естественных условиях – обитатели исключительно аридных луговых и степных стаций. Более древнее появление обозначенной авифаунистической ассоциации в Юго-Восточной Европе свидетельствует о том, что она, очевидно, имеет восточное происхождение.
Список литературы
Воинственский М.А. Ископаемая орнитофауна Украины // Природная обстановка и фауны прошлого. Вып. 3. Киев: Наук. думка, 1967. С. 3–76.
Зеленков Н.В. Зоологические проблемы четвертичной палеорнитологии // Зоол. журн. 2013. Т. 92. № 9. С. 1077–1087.
Зеленков Н.В. Этапы формирования современного таксономического разнообразия птиц (по данным палеонтологии) // Зоол. журн. 2014. Т. 93. № 10. С. 1173–1185.
Зеленков Н.В. Эволюция утиных (Aves: Anatidae s.l.) Евразии в кайнозое // Журн. общ. биол. 2019. Т. 80. С. 323–333.
Зеленков Н.В., Курочкин Е.Н. Неогеновые фазановые (Aves: Phasianidae) Центральной Азии. 2. Роды Perdix, Plioperdix и Bantamyx // Палеонтол. журн. 2009. Вып. 3. С. 79–86.
Зеленков Н.В., Курочкин Е.Н. Класс Aves // Ископаемые рептилии и птицы. Часть 3 / Ред. Курочкин Е.Н., Лопатин А.В., Зеленков Н.В. М.: ГЕОС, 2015. С. 86–290.
Зеленков Н.В., Горобец Л.В. Ревизия Plioperdix (Aves: Phasianidae) из плио-плейстоцена Украины // Палеонтол. журн. 2020. № 5. С. 90–101.
Горобец Л. Птахи як iндикатори палеокекологiчних змiн екосистем пiвдня схiдноï Європи (на прикладi еоцен-голоценових авiфаун. Дисс… д.б.н. Киев: Київ. нац. унів. ім. Т. Шевченка, 2018. 406 с.
Ербаева М.А., Карасев В.В., Алексеева Н.В. Новые данные по стратиграфии плиоцен-плейстоценовых отложений Забайкалья // Геол. и геофиз. 2005. № 4. С. 414–423.
Ковальчук А.Н., Горобец Л.В., Зеленков Н.В. и др. Позвоночные из понтических отложений Шкодовой горы (Северо-Западное Причерноморье, верхний миоцен) // Палеонтол. журн. 2017. № 4. С. 78–92.
Курочкин Е.Н. Птицы Центральной Азии в плиоцене. М.: Наука, 1985. 120 с.
Лопатин А.В. Затерянный мир Тавриды: древнейшая ископаемая пещерная фауна в Крыму // Природа. 2019. Вып. 6. С. 53–61.
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Докл. Акад. Наук. 2019. Т. 485. С. 381–385.
Оксиненко П.В., Лавров А.В. История формирования пещеры Таврида – памятника фауны позвоночных позднего виллафранка и ее палеонтологическое значение // Вестн. Моск. ун-та. Сер. геогр. 2021. № 1. P. 27–42.
Паластрова Е.С. Птицы востока Южной Сибири на рубеже плиоцена и плейстоцена. Дис. … канд. б. н. М.: ПИН РАН, 2022. 228 с.
Соболев Д.В. Водоплавающие птицы позднего неогена Украины // Природничi науки на межi столiть. Матер. наук.-практ. конф. Нiжин, 2004. С. 83–84.
Тесаков А.С. Биостратиграфия среднего плиоцена–эоплейстоцена Восточной Европы (по мелким млекопитающим). М.: Наука, 2004. 247 с.
Тугаринов А.Я. Пластинчатоклювые. М.: Изд-во Акад. наук СССР, 1941. 382 с. (Фауна СССР. Птицы. Т. 1. Вып. 4).
Bedetti C., Pavia M. Early Pleistocene birds from Pirro Nord (Puglia, southern Italy) // Palaeontogr. Abt. A. 2013. V. 298. P. 31–53.
Boles W.E. The avian fossil record of Australia: an overview // Evolution and biogeography of australasian vertebrates / Eds. Merrick J.R., Archer M., Hickey G.M., Lee, M.S.Y. Sidney: Australian Sci. Publ., 2006. P. 387–411.
Elzanowski A., Bieńkowska-Wasiluk M., Chodyń R., Bogdanowicz W. Anatomy of the coracoid and diversity of the Procellariiformes (Aves) in the Oligocene of Europe // Palaeontology. 2012. V. 55. P. 1199–1221.
Erbajeva M., Alexeeva N. Late Cenozoic mammal faunas of the Baikalian Region: Composition, biochronology, dispersal, and correlation with Central Asia // Fossil mammals of Asia: Neogene biostratigraphy and chronology / Eds. Wang X., Flynn L.J., Fortelius M. N.-Y.: Columbia Univ. Press, 2013. P. 495–507.
Fulton T.L., Letts B., Shapiro B. Multiple losses of flight and recent speciation in steamer ducks // Proc. R. Soc. B. 2012. V. 279. № 1737. P. 2339–2346.
Gibert L., Scott G., Martin R., Gibert J. The Early to Middle Pleistocene boundary in the Baza Basin (Spain) // Quatern. Sci. Rev. 2007. V. 26. № 17. P. 2067–2089.
Gurd D.B. Predicting resource partitioning and community organization of filter-feeding dabbling ducks from functional morphology // Amer. Natur. 2007. V. 169. P. 334–343.
Jánossy D. Lower Pleistocene bird remains from Beremend (S-Hungary, loc. 15 and 16) // Aquila. 1992. V. 99. P. 9–25.
Johnsgard P.A. Ducks, geese, and swans of the World. Lincoln (Nebraska): Univ. Nebraska–Lincoln Libraries, 2010. 404 p.
Kessler E. New results with regard to the Neogene and Quaternary Avifauna of the Carpathian Basin, Part I // Földt. Közl. 2009. V. 139. P. 445–468.
Kessler J. A Kárpát-medence madárvilágának őslénytani kézikönyv. Könyvmühel: Miskolc, 2013. 506 p.
Kooloos J.G.M., Kraaijeveld A.R., Langenbach G.E.J., Zweers G.A. Comparative mechanics of filter feeding in Anas platyrhynchos, Anas clypeata and Aythya fuligula (Aves, Anseriformes) // Zoomorphology. 1989. V. 108. P. 269–290.
Livezey B.C. A phylogenetic analysis and classification of recent dabbling ducks (tribe Anatini) based on comparative morphology // Auk. 1991. V. 108. P. 471–507.
Livezey B.C., Zusi R.L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. I. // Bull. Carn. Mus. Natur. Hist. 2006. V. 37. P. 1–544.
Louchart A., Mourer-Chauviré C., Guleç E. et al. The avifauna of Dursunlu, Turkey, Lower Pleistocene: climate, environment and biogeography // C. R. Acad. Sci. Ser. Earth Planet. Sci. 1998. V. 327. № 5. P. 341–346.
Louys J., Roberts P. Environmental drivers of megafauna and hominin extinction in Southeast Asia // Nature. 2020. V. 586. № 7829. P. 402–406.
Masini F., Sala B. Considerations on an integrated biochronological scale of Italian Quaternary continental mammals // Il Quaternario. Ital. J. Quatern. Sci. 2011. V. 24. № 2. P. 193–198.
Meijer H.J.M., Kurniawan I., Setiabudi E. et al. Avian remains from the Early/Middle Pleistocene of the So’a Basin, central Flores, Indonesia, and their palaeoenvironmental significance // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2015. V. 440. P. 161–171.
Miller K.G., Browning J.V., Schmelz W.J. et al. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records // Sci. Adv. 2020. V. 6: eaaz1346.
Mitchell K.J., Wood J.R., Scofield R.P. et al. Ancient mitochondrial genome reveals unsuspected taxonomic affinity of the extinct Chatham duck (Pachyanas chathamica) and resolves divergence times for New Zealand and sub-Antarctic brown teals // Mol. Phylog. Evol. 2014. V. 70. P. 420–428.
Mourer-Chauviré C. The Pleistocene avifaunas of Europe // Archaeofauna. 1993. V. 2. P. 53–66.
Mourer-Chauviré C., Geraads D. The Upper Pliocene avifauna of Ahl al Oughlam, Morocco. Systematics and biogeography // Rec. Austral. Mus. 2010. V. 62. P. 157–184.
Naidina O.D., Richards K. Pollen evidence for Late Pliocene–Early Pleistocene vegetation and climate change in the North Caucasus, North-Western Caspian Region // Quatern. Intern. 2016. V. 409. P. 50–60.
Nelson J.T., Wilson R.E., McCracken K.G. et al. Divergence and gene flow in the globally distributed blue-winged ducks // J. Avian Biol. 2017. V. 48. P. 640–649.
Pazonyi P., Trembeczki M., Meszaros L., Szentesi Z. Preliminary report on the Early Pleistocene vertebrate sites of Beremend Crystal Cave (Beremend 16, South Hungary) and on their palaeoecological importance // Fragm. Palaeontol. Hungar. 2019. V. 36. P. 115–140.
Poland J.G. A methodological approach to the identification of duck and goose remains from archaeological sites with an application to Roman Britain. Unpubl. Ph.D diss. Sheffield: Univ. of Sheffield, 2018. 323 p.
Sánchez Marco A. Huéscar-1 (Granada, España): Avance de la lista de aves y consiguientes conjeturas sobre paleoambiente y paleogeografi’a // Geología y paleontología de la Cuenca de Guadix-Baza. Trabajos sobre Neógeno-Cuaternario. 1989. V. 11. P. 175–182.
Stidham T.A., Wang X., Li Q., Ni X.-J. A shelduck coracoid (Aves: Anseriformes: Tadorna) from the arid early Pleistocene of the Qinghai-Tibetan Plateau, China // Palaeontol. Electron. 2015. V. 18.2.24A: 1–10.
Sun Z., Pan T., Hu C. et al. Rapid and recent diversification patterns in Anseriformes birds: Inferred from molecular phylogeny and diversification analyses // PLoS One. 2017. V. 12: e0184529.
Terzea E. Biochronology of the Pleistocene deposits at Betfia (Bihor, Romania) // Acta Zool. Cracov. 1996. V. 39. № 1. P. 531–540.
Tyrberg T. Pleistocene birds of the Palearctic: a catalogue. Cambridge, Mass.: Nuttall Ornithological Club, 1998. 720 p.
Tyrberg T. Supplement to Pleistocene Birds of the Palearctic. http://web.telia.com/~u11502098/pleistocene.pdf. 2008.
Vickers-Rich P., Monaghan J.M., Baird R.F., Rich T.H. (Eds.) Vertebrate paleontology of Australia. Melbourne: Pioneer Design Studio, 1991. 1437 p.
Worthy T.H. Pliocene waterfowl (Aves: Anseriformes) from South Australia and a new genus and species // Emu. 2008. V. 108. P. 153–165.
Worthy T.H., Lee M.S.Y. Affinities of Miocene waterfowl (Anatidae: Manuherikia, Dunstanetta and Miotadorna) from the St Bathans Fauna, New Zealand // Palaeontology. 2008. V. 51. № 3. P. 677–708.
Worthy T.H., Nguyen J.M.T. An annotated checklist of the fossil birds of Australia // Trans. R. Soc. South Australia. 2020. V. 144. № 1. P. 66–108.
Zelenkov N.V. Variability in the postcranial skeleton of the European dabbling ducks (Aves, Anatidae): identifying nodes of strong and relaxed selection // J. Morphol. 2019. V. 280. Suppl. 1. S. 243.
Zelenkov N.V., Lavrov A.V., Startsev D.B. et al. A giant early Pleistocene bird from eastern Europe: unexpected component of terrestrial faunas at the time of early Homo arrival // J. Vertebr. Paleontol. 2019. V. 39: e1605521.
Zelenkov N.V., Panteleyev A.V. Three bird taxa (Aves: Anatidae, Phasianidae, Scolopacidae) from the Late Miocene of the Sea of Azov (Southwestern Russia) // Paläontol. Z. 2015. V. 89. № 3. P. 515–527.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал