

Палеонтологический журнал, 2022, № 6, стр. 74-79
Изменения состава эоблаттидовых насекомых (Insecta: Eoblattida) у границы перми и триаса
Д. С. Аристов *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: danil_aristov@mail.ru
Поступила в редакцию 14.02.2022
После доработки 21.03.2022
Принята к публикации 21.03.2022
- EDN: QQGBYK
- DOI: 10.31857/S0031031X22060022
Аннотация
Рассмотрены изменения семейственного состава эоблаттидовых насекомых (Insecta: Eoblattida) у границы перми и триаса. Из нижнетриасового местонахождения Петропавловка (Оренбургская обл. России) описан Baharellinus orenburgensis sp. nov. (Blattogryllidae). Семейство Gorochoviidae, известное из местонахождения Джайлоучо (средний или верхний триас Кыргызстана), перенесено из отряда Reculida в Eoblattida. Для рода Batkentak из того же местонахождения выделено семейство Batkentakidae fam. nov. Показано отсутствие существенных изменений в семейственном составе эоблаттидовых на границе палеозоя и мезозоя.
В настоящей работе рассматриваются изменения семейственного состава отряда Eoblattida c северодвинского века поздней перми по ладинский–карнийский века среднего–позднего триаса. Неточность верхней границы рассматриваемого интервала объясняется соответствующей датировкой местонахождения Джайлоучо (одно из обнажений мадыгенской свиты) в Кыргызстане (Shcherbakov, 2008; Voigt et al., 2017). Последовательность и возраст большинства местонахождений приведены по ранее опубликованным нами данным (Расницын и др., 2013). Местонахождение Недуброво (вохминская свита Вологодской обл. России) отнесено к вятскому ярусу верхней перми (Лозовский и др., 2016). Кедровские слои мальцевской свиты (местонахождение Бабий Камень в Кемеровской обл. России) отнесены к вятскому ярусу верхней перми (Yan et al., 2020). Одновозрастные (Могучева, Круговых, 2009) с мальцевской свитой отложения лебедевского горизонта (местонахождение Анакит, Красноярский край России) также рассматриваются как верхнепермские (Садовников, 2016). В соответствии с другой точкой зрения отложения Бабьего Камня имеют оленекский возраст (Davydov et al., 2021).
Эоблаттидовые насекомые известны с позднего карбона по современность. Обзор позднепермских эоблаттидовых дан по опубликованным данным (Aristov, 2015 и более поздние работы, ссылки на которые даны ниже) и по материалам, описанным в настоящей работе. Обзор триасовых эоблаттидовых дан по опубликованным данным (Стороженко, 1998 и более поздние работы, ссылки на которые даны ниже) и по оригинальным данным автора.
Из северодвинских отложений верхней перми на сегодняшний день известны представители 10‑ти семейств эоблаттидовых (табл. 1). К ним относятся Cacurgidae, Atactophlebiidae, Permotermopsidae, Protophasmatidae, Megakhosaridae, Mesorthopteridae, Blattogryllidae, Bardapteridae и Soyanopteridae из местонахождения Исады (Россия, Вологодская обл.). Из близковозрастных местонахождений Ново-Александровка и Кульчумово (Россия, Оренбургская обл.) описаны Blattogryllidae (Аристов, 2021) и Mesorthopteridae. Род Alekhosara, описанный как Megakhosaridae из Ново-Александровки (Аристов, 2008), относится к отряду Orthoptera (А.В. Горохов, личн. сообщ.). Из местонахождения Суриёкова I (Россия, Кемеровская обл.) известны Megakhosaridae. Из учапиньского (в табл. 1 отнесен к северодвинскому ярусу: Щербаков, 2008) местонахождения Бор Тологой (Южно-Гобийский аймак, Монголия) описаны Ideliidae.
Таблица 1.
Стратиграфическое распространение семейств отряда Eoblattida у границы перми и триаса. Сокращения: sdv, vt, ol, an, l/c – северодвинский, вятский, оленекский, анизийский, ладинский или карнийский яруса, соответственно. Местонахождение “Исады и др.” включает в себя, кроме Исад, местонахождения Ново-Александровка, Кульчумово и Суриёкова I
| раньше | Пермь | Триас | позже | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P3 | T1 | T2 | T2/3 | ||||||||||
| sdv | vt | ol | an | l/c | |||||||||
| Бор Тологой | Исады и др. | Моой Ривер | Караунгир | Балымотиха | Недуброво | Бабий Камень | Анакит | Петропавловка | Вогезы, Порт де Эстелленс | Джайлоучо | |||
| Cacurgidae | + | – | + | ||||||||||
| Atactophlebiidae | + | – | + | ||||||||||
| Permotermopsidae | + | – | + | ||||||||||
| Protophasmatidae | + | – | + | ||||||||||
| Bardapteridae | + | – | + | ||||||||||
| Soyanopteridae | + | – | + | ||||||||||
| Ideliidae | + | + | – | – | – | – | – | – | – | – | – | + | + |
| Megakhosaridae | + | – | + | + | + | + | – | + | + | – | + | + | |
| Mesorthopteridae | + | – | + | – | + | – | – | – | + | – | – | + | + |
| Blattogryllidae | + | – | + | – | – | – | + | + | + | + | + | + | + |
| Babalidae | + | ||||||||||||
| Gorochoviidae | + | ||||||||||||
| Batkentakidae | + | ||||||||||||
Из вятских отложений верхней перми из эоблаттидовых описаны представители семейства Megakhosaridae (местонахождение Балымотиха во Владимирской обл. России) и Blattogryllidae (местонахождение Недуброво в Вологодской обл.). Наиболее крупным местонахождением вятских эоблаттидовых является Бабий Камень (Кемеровская обл. России). Из этого местонахождения описаны представители Megakhosaridae, Blattogryllidae и эндемичные Babalidae (Аристов, 2020). Из близковозрастного местонахождения Анакит (Красноярский край России) описаны Megakhosaridae, Blattogryllidae (Aristov, 2015) и Mesorthopteridae (Аристов, 2021).
Из чансиньского (в табл. 1 отнесено к вятскому ярусу: Щербаков, 2008) местонахождения Моой Ривер в провинции Квазулу-Наталь, ЮАР, описаны Megakhosaridae. Из чансиньского местонахождения Караунгир в Казахстане описаны Megakhosaridae и Mesorthopteridae.
Индские эоблаттидовые на сегодняшний день неизвестны. Из оленекского местонахождения Петропавловка (Оренбургская обл. России) ниже описан новый вид Blattogryllidae – Baharellinus orenburgensis sp. nov.
Из среднетриасового (нижний анизий: Matamales-Andreu et al., 2021) местонахождения Порт де Эстелленс (Испания, о. Майорка) описаны Blattogryllidae (Aristov, Zessin, 2009). Из анизийского местонахождения Вогезы (группа местонахождений в Эльзасе и Лотарингии, Франция) описаны представители Megakhosaridae и Blattogryllidae (Аристов и др., 2011). Род Palmesorthopteron из этого местонахождения, описанный в семействе Mesorthopteridae (Аристов и др., 2011), был перенесен в семейство эмбий Alexarasniidae (Shcherbakov, 2015).
Наиболее богатым местонахождением мезозойских эоблаттидовых является триасовое местонахождение Джайлоучо (одно из обнажений мадыгенской свиты: Shcherbakov, 2008). Мадыгенская свита относится к ладинскому или карнийскому ярусу (Shcherbakov, 2008; Voigt et al., 2017). Из этого местонахождения были описаны представители семейств Mesorthopteridae, Ideliidae, Megakhosaridae, Blattogryllidae и Daldubidae (Aristov, 2015). Ниже к эоблаттидовым отнесено семейство Gorochoviidae (ранее – отряд Reculida) из Джайлоучо. Род Batkentak, описанный в семействе Daldubidae из того же местонахождения (Aristov, 2015), выделен ниже в новое монотипное семейство Batkentakidae.
Таким образом, наиболее существенные изменения семейственного состава эоблаттидовых насекомых в окрестностях границы перми и триаса происходят на границе северодвинского и вятского ярусов поздней перми (табл. 1). Из северодвинских семейств в вятский век и мезозой переходят только Megakhosaridae, Mesorthopteridae, Blattogryllidae и Ideliidae. Почти две трети (60%) семейств (Cacurgidae, Atactophlebiidae, Bardapteridae, Permotermopsidae, Protophasmatidae и Soyanopteridae) исчезают, что является самым крупным снижением разнообразия за всю историю отряда. Вымирание это некомпенсированное, в более молодых отложениях появления регистрируются в вятском веке верхней перми (Babalidae) и среднем или верхнем триасе (Gorochoviidae и Batkentakidae). При этом Gorochoviidae представлены тремя родами с восемью видами. Семейства Babalidae и Batkentakidae монотипные, известны только по голотипам. После этих семейств появляется только современное семейство Grylloblattidae, известное из Азии и Северной Америки (Стороженко, 1998).
На самой же границе перми и триаса существенных изменений фауны не происходит – все вятские семейства (Ideliidae не найдены в вятских отложениях, но описаны из северодвинских и триасовых), за исключением эндемичных Babalidae, переходят в мезозой.
Автор признателен А.П. Расницыну и А.Г. Пономаренко за замечания по рукописи. Работа поддержана грантом РНФ № 21-14-00284.
ОТРЯД EOBLATTIDA
СЕМЕЙСТВО BLATTOGRYLLIDAE RASNITSYN, 1976
Род Baharellinus Storozhenko, 1992
Baharellinus orenburgensis Aristov, sp. nov.
Табл. XIII , фиг. 1, 2 (см. вклейку)
Рис. 1.
Еоблаттидовые из триаса Евразии: а – Baharellinus orenburgensis sp. nov., голотип ПИН, № 5640/268, переднее крыло; Россия, Оренбургская обл., Петропавловка; нижний триас, оленекский ярус, верхнеоленекский подъярус, петропавловская свита; б – Gorochovia minuta Storozhenko, 1994, паратип ПИН, № 2069/1847; переднее крыло; в – Batkentak intactus Aristov, 2015, голотип ПИН, № 2785/2179, переднее и заднее крылья; Кыргызстан, Ошская обл., Джайлоучо; средний или верхний триас, ладинский или карнийский ярус, мадыгенская свита. Длина масштабной линейки 2 мм.
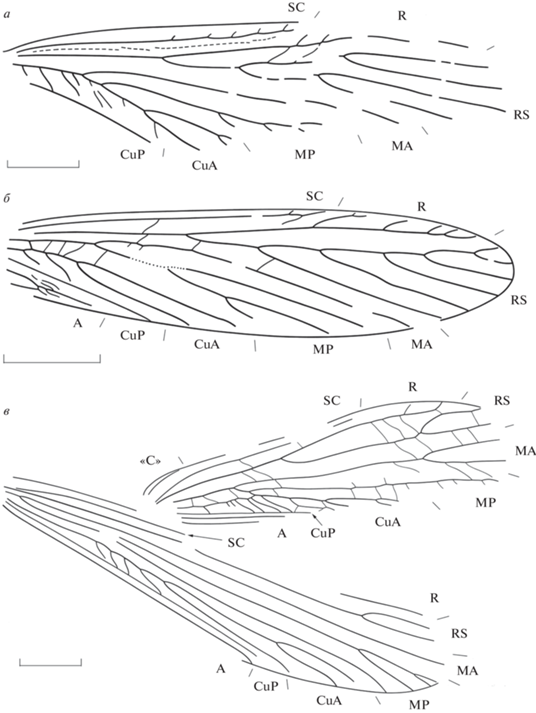
Название вида – от Оренбургской области.
Голотип – ПИН, № 5640/268, прямой и обратный отпечатки переднего крыла; Россия, Оренбургская обл., Сакмарский р-н, местонахождение Петропавловка; нижний триас, оленекский ярус, верхнеоленекский подъярус, петропавловская свита (Щербаков и др., 2019).
Описание (рис. 1, а). Средних размеров насекомые. Передний край переднего крыла выпуклый. Костальное поле резко сужено в основании, его ширина на уровне основания RS немного меньше ширины субкостального. SC с тонкими передними ветвями, заканчивается сразу за серединой крыла. В субкостальном поле проходит продольная складка. R с тонкими передними ветвями. RS начинается в конце базальной трети крыла, начинает ветвиться перед его серединой, дихотомический, с четырьмя ветвями. Основание М полностью слито с CuA, МА простая или с коротким развилком, МР гребенчатая назад, с четырьмя или более ветвями. Анастомоз М + CuA с простыми, S-образно изогнутыми и дихотомирующими задними ветвями. CuA начинает ветвиться на некотором расстоянии от основания МР, с тремя ветвями. CuP слегка изогнута вперед. Поперечные жилки простые, S-образно изогнутые.
Размеры в мм: предполагаемая длина крыла около 16.
Сравнение. Новый вид наиболее сходен с B. arctous Aristov, 2020 из местонахождения Бабий Камень (Россия, Кемеровская обл.; верхняя пермь, вятский ярус), от которого отличается простой МА и ветвящейся МР. У B. arctous МА ветвится, МР простая (Аристов, 2020).
Материал. Голотип.
СЕМЕЙСТВО GOROCHOVIIDAE STOROZHENKO, 1994
Замечания. Семейство Gorochoviidae было описано из местонахождения Джайлоучо (средний или верхний триас Кыргызстана) в составе отряда Grylloblattida (Storozhenko, 1994). При ревизии этого отряда Gorochoviidae были отнесены к отряду Reculida (Aristov, 2015). Однако переизучение типового материала показало наличие у гороховиид ряда признаков, не характерных для рекулидовых. Для этого семейства характерен многоветвистый (с шестью–восьмью, а чаще всего с семью, окончаниями), гребенчатый назад RS. Для паратипа Gorochovia minuta (ПИН, № 2785/2123а) был изображен трехветвистый RS (Storozhenko, 1994, рис. 6 ). Переизучение этого отпечатка показало, что вершина крыла не сохранилась, и у целого крыла ветвей RS должно было быть больше. Для рекулидовых многоветвистый, гребенчатый назад RS не характерен (Стороженко, 1998; семейства, относимые к Reculida, см.: Aristov, 2015), но обычен для некоторых эоблаттидовых (напр., Megakhosaridae: Стороженко, 1998).
Кроме того, для Gorochoviidae характерны М5 и задние ветви CuA в интеркубитальном поле (рис. 1, б; табл. XIII , фиг. 3 ). Эти жилки на некоторых экземплярах могут быть редуцированы почти до толщины поперечных жилок. Однако выраженность этих признаков даже у некоторых экземпляров позволяет предполагать, что такое строение М5 и CuA является исходным для Gorochoviidae. Среди рекулидовых М5 известна только у представителей семейства Geinitziidae; задние ветви CuA, характерные для многих эоблаттидовых, у представителей Reculida не известны (Стороженко, 1998; Aristov, 2015). Эти признаки позволяют перенести семейство Gorochoviidae из отряда Reculida в Eoblattida.
Среди эоблаттидовых гороховииды наиболее сходны с семейством Megakhosaridae из перми и триаса Евразии и Африки. От этого семейства отличаются узким (уже субкостального) костальным полем и разделением CuA на CuA1 и CuA2. У мегахозарид костальное поле шире субкостального поля или равно ему по ширине, CuA не разделена на CuA1 и CuA2 (Aristov, 2015).
СЕМЕЙСТВО BATKENTAKIDAE ARISTOV, FAM. NOV.
Типовой род – Batkentak Aristov, 2015.
Диагноз. Переднее крыло с небольшим прекостальным полем. Костальное поле у основания RS уже субкостального. R короткий, дистальная ветвь RS выходит на передний край крыла. М разделяется на МА и МР в базальной четверти крыла, М5 отходит от МР. Ствол М до отхождения от него М5, основание МР, М5 и ствол CuA после впадения в него М5 мощнее, чем основания МА и МР и вершина ствола CuA до впадения в М5. CuA c частыми S-образными задними ветвями в интеркубитальном поле, заканчивающимися на CuP. CuA гребенчатая назад, начинает ветвиться в своей дистальной трети.
В заднем крыле основание костального поля расширено. RS начинается сразу за базальной третью крыла. М разделяется на МА и МР в основании крыла. CuA выпуклая, с задними ветвями в интеркубитальном поле, начинает ветвиться у своей середины.
Состав. Типовой род из среднего или верхнего триаса Кыргызстана.
Сравнение. Новое семейство наиболее сходно с Daldubidae из карбона России, от которого, как и от остальных семейств эоблаттидовых, отличается наличием прекостального поля (Aristov, 2015). Мощный ствол М + М5 + CuA характерен для карбоновых представителей Cacurgidae (у пермских эоблаттидовых этот признак не известен), от которых Batkentakidae отличаются ранним разделением М на МА и МР. У какургид М начинает ветвиться у середины крыла (Аристов, 2012).
Род Batkentak Aristov, 2015
Batkentak: Aristov, 2015, с. 37.
Типовой вид – B. intactus Aristov, 2015.
Диагноз. Тот же, что и для семейства.
Видовой состав. Типовой вид.
Batkentak intactus Aristov, 2015
Табл. XIII , фиг. 4
Batkentak intactus: Aristov, 2015, с. 37, рис. 24–28 .
Голотип – ПИН, № 2785/2179, прямой и обратный отпечатки деформированных неполного переднего и целого заднего крыльев; Кыргызстан, Ошская обл., Баткенский р-н, урочище Мадыген, местонахождение Джайлоучо; средний или верхний триас, ладинский или карнийский ярус, мадыгенская свита.
Oписание (рис. 1, в). Передний край переднего крыла слабовогнутый перед серединой крыла, далее выпуклый. Костальное поле у основания RS немного уже субкостального, SC заканчивается перед дистальной третью крыла. RS начинается перед серединой крыла, начинает ветвиться в его дистальной трети, с двумя ветвями. МА не слита с RS, разделяется на две ветви в дистальной трети крыла, МР простая. CuA до впадения М5 тонкая, с изломами в местах впадения поперечных жилок, задние ветви частые, S‑образно изогнутые. CuA с тремя или более короткими основными ветвями. CuP прямая, А1 простая, А2 простая или с несколькими ветвями. Поперечные жилки простые.
В заднем крыле базальная часть костального поля, после расширения в основании, равнa по ширине субкостальному. RS с двумя короткими ветвями, МА простая, слабо S-образно изогнута, МР начинает ветвиться в базальной трети крыла, с четырьмя окончаниями. Ствол CuA до разделения на ветви изогнут S-образно, задние ветви редкие, CuA с тремя ветвями. CuP прямая, заканчивается за серединой крыла. А1 простая, прямая.
Размеры в мм: предполагаемая длина переднего крыла около 14, заднего – 12.
Материал. Голотип.
Список литературы
Аристов Д.С. Новые представители семейства Megakhosaridae (Insecta: Grylloblattida) из перми России // Палеонтол. журн. 2008. № 3. С. 45–49.
Аристов Д.С. Состав и распространение семейства Cacurgidae (Insecta; Grylloblattida) // Палеонтол. журн. 2012. № 3. С. 29–36.
Аристов Д.С. Новые гриллоновые насекомые (Insecta: Gryllones) из местонахождения Бабий Камень (верхняя пермь России). Часть 1. Отряд Eoblattida // Палеонтол. журн. 2020. № 1. С. 44–49.
Аристов Д.С. Новые эоблаттидовые насекомые (Insecta: Eoblattida) из верхней перми России // Палеонтол. журн. 2021. № 1. С. 77–79.
Аристов Д.С., Гровожель-Стамм Л., Маршал-Папье Ф. Новые гриллоблаттидовые насекомые (Insecta: Grylloblattida) из вольциевого песчаника Вогез (средний триас Франции) // Палеонтол. журн. 2011. № 2. С. 39–45.
Лозовский В.Р., Балабанов Ю.П., Карасев Е.В. и др. Терминальная пермь Европейской России: вязниковский горизонт и недубровская пачка и граница перми и триаса // Стратигр. Геол. корреляция. 2016. Т. 24. № 4. С. 38–54.
Могучева Н.К., Круговых В.В. Новые данные к стратиграфической схеме триасовых отложений Тунгусской синеклизы и Кузнецкого бассейна // Стратигр. Геол. корреляция. 2009. Т. 17. № 5. С. 60–68.
Расницын А.П., Аристов Д.С., Расницын Д.А. Насекомые у рубежа перми и раннего триаса (уржумский–оленекский века) и проблема пермотриасового кризиса биоразнообразия // Журн. общ. биол. 2013. Т. 74. № 1. С. 43–65.
Садовников Г.Н. Трапповый вулканизм Сибири и “пермо-триасовое вымирание” // Геол. и разведка. 2016. № 2. С. 8–14.
Стороженко С.Ю. Систематика, филогения и эволюция гриллоблаттидовых насекомых (Insecta: Grylloblattida). Владивосток: Дальнаука, 1998. 207 с.
Щербаков Д.Е. О пермских и триасовых энтомофаунах в связи с биогеографией и пермо-триасовым кризисом // Палеонтол. журн. 2008. № 1. С. 15–32.
Щербаков Д.Е., Башкуев А.С., Василенко Д.В. и др. Новое местонахождение раннетриасовых насекомых – Петропавловка // Палеострат–2019. Годичн. собр. (научн. конф.) секции палеонтол. МОИП и Моск. отд. Палеонтол. об-ва при РАН. М.: ПИН РАН, 2019. С. 68–69.
Aristov D.S. Classification of the order Eoblattida (Insecta: Blattidea) with description of new taxa // Far East. Entomol. 2015. № 301. P. 1–56.
Aristov D., Zessin W. Mallorcagryllus hispanicus n. gen. et sp. – eine neue Grylloblattide (Insecta: Grylloblattida: Blattogryllidae) aus dem Buntsandstein der Insel Mallorca, Spanien // Virgo, Mitt. Entomol. Ver. Mecklenburg. 2009. V. 12. № 1. P. 30–34.
Davydov V.I., Karasev E.V., Nurgalieva N.G. et al. Climate and biotic evolution during the Permian-Triassic transition in the temperate Northern Hemisphere, Kuznetsk Basin, Siberia, Russia // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2021. V. 573. Art. 110432. https://doi.org/10.1016/j.palaeo.2021.110432
Matamales-Andreu R., Peñalver E., Mujal E. et al. Early–Middle Triassic fluvial ecosystems of Mallorca (Balearic Islands): Biotic communities and environmental evolution in the equatorial western peri-Tethys // Earth-Sci. Rev. 2021. V. 222. Art. 103783. https://doi.org/10.1016/j.earscirev.2021.103783
Shcherbakov D.E. Madygen, Triassic Lagerstätte number one, before and after Sharov // Alavesia. 2008. № 2. P. 113–124.
Shcherbakov D.E. Permian and Triassic ancestors of webspinners (Embioidea) // Russ. Entomol. J. 2015. V. 24. № 3. P. 187–200. https://doi.org/10.15298/rusentj.24.3.01
Storozhenko S. New Triassic grylloblattids from Kirghizia (Insecta, Grylloblattida) // Spixiana. 1994. V. 17. № 1. P. 27–35.
Voigt S., Buchwitz M., Fischer J. et al. Triassic life in an inland lake basin of the warm-temperate biome – the Madygen Lagerstätte (Southwest Kyrgyzstan, Central Asia) // Terrestrial conservation Lagerstätten: Windows into the evolution of life on land / Eds. Fraser N.C., Sues H.-D. Edinburgh: Dunedin Acad. Press, 2017. P. 65–104.
Yan E.V., Beutel R.G., Lawrence J.F. et al. Archaeomalthus (Coleoptera, Archostemata) a ‘ghost adult’ of Micromalthidae from Upper Permian deposits of Siberia? // Hist. Biol. 2020. V. 32. № 8. P. 1019–1027. https://doi.org/10.1080/08912963.2018.1561672
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал