

Палеонтологический журнал, 2022, № 6, стр. 58-73
Позднебайосские и раннебатские остракоды Русской плиты. Часть IV. Филолиния Glyptocythere aspera (Khabarova) → G. bathonica sp. nov.
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
b Геологический институт РАН
119017 Москва, Россия
* E-mail: ostracon@rambler.ru
Поступила в редакцию 31.03.2022
После доработки 05.07.2022
Принята к публикации 11.07.2022
- EDN: OXVCKQ
- DOI: 10.31857/S0031031X22060125
Аннотация
Проведена ревизия вида Glyptocythere aspera (Khabarova, 1955), важного для стратиграфии верхнего байоса – нижнего бата Русской плиты (РП), Западного Казахстана и Узбекистана. Выполнено его расширенное и актуализированное переописание и выделен неотип. Вид G. bathonica из нижнего бата РП и Западного Казахстана описан как новый. Реконструирована филолиния G. tuberodentina Brand et Malz in Brand et Fahrion (bj2, аммонитовая зона Parkinsoni) → G. aspera (bj2-bt1, аммонитовые зоны Michalskii и нижняя часть Besnosovi) → G. bathonica sp. nov. (bt1, верхняя часть аммонитовой зоны Besnosovi), которая развивалась на РП с опережающей эволюцией у самцов (путем педоморфоза). Видовые признаки (контур и линейные параметры раковины, замок и скульптура) изменялись у самок и самцов с разной скоростью, у самцов с ретардацией, что привело к сближению облика раковин полов у G. bathonica sp. nov. Т.е., эволюция филолинии сопровождалась ослаблением полового диморфизма. Вид G. tuberodentina появился и развивался в Северной Германии в позднем байосе (фаза Parkinsoni) и в это же время мигрировал через Польское море в акватории РП. Северогерманская зона распространения вида G. tuberodentina (bj2, зона Parkinsoni, верхняя часть подзоны Truellei и подзона Bomfordi) впервые прослежена в Польше и Днепровско-Донецкой впадине (ДДВ). В ДДВ объем зоны G. tuberodentina сокращен за счет выделения в зоне Michalskii новой остракодовой филозоны G. aspera. На базе реконструированной филолинии установлены новые филозоны: G. tuberodentina (bj2, зона Parkinsoni) для ДДВ; G. aspera (bj2-bt1, зоны Michalskii и Besnosovi с биогоризонтами O. mojarowskii, O. sciticum) для ДДВ, Центральной России, Поволжья и Западного Казахстана; и G. bathonica (bt1, зона Besnosovi, биогоризонты O. issae–A. excentricum) для Центральной России, Поволжья и Западного Казахстана.
ВВЕДЕНИЕ
Настоящая работа является продолжением систематического изучения остракод из верхнего байоса и нижнего бата Русской плиты (РП) по материалам из разрезов Нижнего Поволжья и центральных районов России, результаты которого частично опубликованы автором ранее (Тесакова, 2022а, б, в). Были представлены результаты ревизии, а также уточненные и расширенные описания стратиграфически значимых родов Plumhoffia Brand, 1990, Pseudohutsonia Wienholz, 1967, Procytherura Whatley, 1970 (семейство Cytheruridae) и Aaleniella Plumhoff, 1963 (семейство Eucytheridae). Для трех видов, установленных Т.Н. Хабаровой (1955) – Plumhoffia tricostata (Khabarova, 1955) Pseudohutsonia clivosa (Khabarova, 1955) и Procytheridea? bajociensis (Khabarova, 1955) – выделены неотипы, проведена ревизия и выполнены актуализированные переописания. Восемь других видов: Camptocythere (Anabarocythere) triangula, Aaleniella franzi, A. volganica, A.? ovoidea, Procytherura iyae, Acrocythere sokurensis, Nanacythere octum и Trachycythere peculiaris описаны как новые. Для четырех таксонов, оставленных в открытой номенклатуре (Nanacythere sp. 1, N. sp. 2, Ljubimovella sp. 1 и Gen. et sp. 8), приведены сведения о материале и распространении по изученным разрезам. Перечисленные виды (новые и виды Хабаровой) являются индексами комплексов, образующих стратиграфическую последовательность в отложениях терминального байоса и нижнего бата (аммонитовые зоны Michalskii и Besnosovi) РП; нижние границы этих комплексов определяются по первому появлению видов-индексов. Скорее всего, эти комплексы имеют палеоэкологическую природу, но при прослеживании на большой площади могут приобрести стратиграфическое значение и статус слоев с фауной или зон. Кроме того, в статье Е.М. Тесаковой (2022в) был опубликован вид Camptocythere (A.) muricata Gerke et Lev из верхнего бата–келловея севера Сибири, Тимано-Печорской провинции и Баренцевоморского шельфа. Публикация описания, составленного О.М. Лев (НИИГА), но не увидевшего свет, расширенного и дополненного Тесаковой, обеспечило валидность вида и устранило недостатки в стратиграфической литературе, где C. (A.) muricata фигурировал как nom. nud. Вероятное родство между этим видом и C. (A.) triangula позволяет рассматривать их индексами соответствующих филозон в камптоцитеровой шкале по эволюции подрода C. (Anabarocythere) Nikitenko, 1994 (Тесакова, 2022в).
В настоящей статье продолжается монографическое описание остракод рода Glyptocythere Brand еt Malz, 1962 из сводного Сокурского разреза (Саратовская обл.), скважины Обвал (Пензенская обл.) и разреза Плетневского карьера (Пензенская обл.) (рис. 1). Глиптоцитеры – весьма разнообразные и обильные остракоды со сравнительно крупной, хорошо орнаментированной раковиной, широко расселившиеся в морях Европы в байосском и батском веках. Приуроченность рода именно к этим двум ярусам наблюдалась во всех разрезах суббореальной и бореально-атлантической палеогеографических провинций Западной, Центральной и Восточной Европы (ссылки на литературу см. ниже). И только в более тепловодных шельфовых морях северной окраины Тетис (в Узбекистане) они дожили до раннего келловея (Масумов, 1973). В Северной Германии этот род успешно использовался в биостратиграфии (Brand, Malz, 1966; Brand, Mönnig, 2009). Совместное нахождение глиптоцитер в ассоциации с заведомо морскими фаунами, в том числе стеногалинными (аммониты, белемниты и др.), позволяет считать их нормально-морскими шельфовыми видами (Brand, Malz, 1962). Однако, некоторые представители смогли расширить свою галинную валентность в пределах 5–35‰ (Wakefield, 1995).
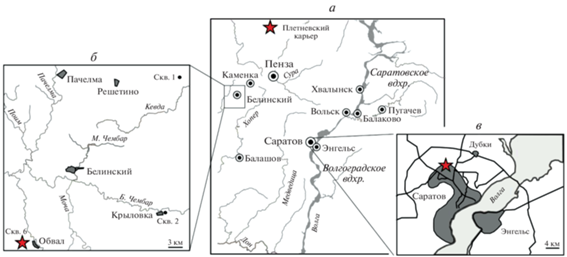
Рис. 1.
Разрезы, из которых изучены остракоды, отмечены на карте-схеме звездочками: а – Плетневский карьер в Иссинском р-не Пензенской обл., б – скважина Обвал в Тамалинском р-не Пензенской обл., в – Сокурский карьер и скважина на северной окраине г. Саратов.
Свободная миграция среднеюрских остракод в морях Западной Европы подчеркивается высокой общностью комплексов глиптоцитер (наряду с местной спецификой) Англии (Sylvester-Bradley, 1948; Bate, 1965, 1967, 1969, 2009; Sheppard, 1981 и др.), Франции (Sheppard, 1981; Dépêche, 1984, 1985 и др.), Германии (Brand, Fahrion, 1962; Brand, Malz, 1962, 1966; Lutze, 1966; Brand, 1990; Franz et al., 2009, 2014; Luppold, 2012; Dietze et al., 2017, 2018 и др.) и Швейцарии (Tesakova, 2017). Эти комплексы заметно отличаются от комплексов глиптоцитер из Польши (Błaszyk, 1967; Bielecka et al., 1988 и др.); еще бóльшие различия наблюдаются с украинскими представителями (Пермякова, 1970, 1975; Пяткова, Пермякова, 1978); а общих видов со среднерусскими таксонами (Хабарова, 1955; Преображенская, 1966; Шурупова и др., 2016) и Узбекистаном (Масумов, 1973) нет совсем. Виды глиптоцитер, общие между Западной Европой и Польшей – G. tuberodentina Brand et Malz in Brand et Fahrion, 1962, G. interrete Brand et Malz, 1966, G. similis Brand et Malz, 1966, G. regulariformis Brand et Malz in Brand et Fahriоn, 1962, G. medisulcata Blaszyk, 1972, G. perpolita magna Blaszyk, 1967, G. posterocostata Blaszyk, 1972 (Bielecka, Styk, 1969; Bielecka et al., 1988; Tesakova et al., 2008); общие с Украиной – G. concentrica Brand et Malz, 1966, G. tuberodentina, G. regulariformis (Пяткова, Пермякова, 1978).
Четыре представителя этого рода из верхнебайосской жирновской толщи Волгоградской обл. (G. aspera, G. strigatus, G. praerimosa и G. tuberosa) были описаны Т.Н. Хабаровой (1955) как новые (к сожалению, недостаточно подробно и без надлежащего сравнения) и ошибочно отнесены к родам Protocythere Triebel, 1938 и Palaeocytheridea Mandelstam, 1947. Их остатки известны из верхнего байоса и нижнего бата Саратовской и Волгоградской областей, а также Заволжья и Западного Казахстана (Хабарова, 1955, 1961; Салтыков и др., 2008). Самый обильный и распространенный таксон, G. aspera (Khabarova, 1955), был отмечен (кроме упомянутой территории) в синхронных отложениях Курской (Преображенская, 1966) и Белгородской (Колпенская, 2002) областей и Узбекистана (Масумов, 1973), что позволило считать его одной из руководящих форм для верхнего байоса – нижнего бата РП (Преображенская, 1964, 1966; Колпенская, 2002).
Корректное использование в биостратиграфии видов Хабаровой, в т.ч. глиптоцитер, требовало выделения для них неотипов (место хранения их голотипов в настоящий момент неизвестно) и актуализиции описаний (Тесакова, 2022а), что и стало основной задачей настоящей работы. Сначала автор изучила на сканирующем микроскопе стратиграфически важный таксон G. aspera, в результате чего был существенно расширен комплекс видовых признаков и установлены особенности, различные у позднебайосских и раннебатских представителей. Последние выделены в новый вид G. bathonica sp. nov. (см. ниже).
Другая задача этой статьи была связана с прояснением стратиграфического значения описанных видов и возможности их использования в биостратиграфии. Анализ развития в онтогенезе изученных видов (и известных из литературы) таких признаков, как форма раковины, замок и скульптура, причем раздельно для самок и самцов, выявил гетерохронии (см. ниже), которые доказывают родство видов G. tuberodentina (bj2, аммонитовая зона Parkinsoni) → G. aspera (bj2-bt1, аммонитовые зоны Michalskii и нижняя часть Besnosovi) → G. bathonica sp. nov. (bt1, верхняя часть аммонитовой зоны Besnosovi). В С.-З. Европе остракодовая зона выделена по распространению таксона G. tuberodentina (Brand, Mönnig, 2009), которую можно рассматривать и как филозону тоже. А в пределах РП нами предложены две новые филозоны по эволюции G. aspera и G. batho-nica sp. nov. (см. ниже).
Для научных учреждений, упомянутых в тексте, приняты следующие аббревиатуры: ВГУ (Воронежский государственный ун-т), ГИН РАН (Геологический ин-т РАН, Москва), ИГ РАН (Ин-т географии РАН, Москва), ИНГГ СО РАН (Ин-т нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН, Новосибирск), МГУ (Московский государственный ун-т им. М.В. Ломоносова), ПИН РАН (Палеонтологический ин-т им. А.А. Борисяка РАН, Москва), СНИГУ (Саратовский национальный исследовательский государственный ун-т им. Н.Г. Чернышевского), ЦНИЛ (Центральная научно-исследовательская лаборатория) объединения “Саратовнефть” (ныне ПАО “Саратовнефтегаз”; см. Тесакова, 2022а), НИИГА (Научно-исследовательский ин-т геологии Арктики; впоследствии преобразован во Всесоюзный научно-исследовательский ин-т геологии и минеральных ресурсов Мирового океана – ВНИИОкеангеология, СПб.), LGRB (Государственное управление геологии, сырья и горных работ, г. Фрайбург-им-Брайсгау, Германия).
МАТЕРИАЛ И МЕТОДЫ
О происхождении материала и методах его изучения из Сокурского сводного разреза (разрезы скважины, пробуренной в Сокурском карьере, и открытой части этого карьера; северная окраина г. Саратов) и разреза скважины Обвал (Тамалинский р-н, Пензенская обл.) (рис. 1, б, в) подробно написано ранее (Тесакова, 2022а).
Остракоды из разреза Плетневского карьера, расположенного на правом берегу р. Исса напротив с. Каменный Брод и д. Плетневка Иссинского р-на Пензенской обл. (53.860° с.ш., 44.693° в.д.) (рис. 1, а), любезно переданы автору Л.А. Глинских (ИНГГ СО РАН) из проб, отобранных А.П. Ипполитовым (ГИН РАН) во время полевого сезона 2016 г. (Гуляев, Ипполитов, 2017; Гуляев, 2019). Из отложений нижнего бата было отобрано 13 образцов на микрофауну; фораминиферы встречены во всех (устное сообщение Глинских), а остракоды найдены только в двух, оба из слоя 1b (аммонитовый биогоризонт Orani-ceras issae) (Гуляев, 2019). Комплекс остракод был представлен единичными экземплярами удовлетворительной сохранности видов G. bathonica sp. nov. (табл. XII , фиг. 10 а, 10б; см. вклейку), P.? bajociensis (Khabarova, 1955) и Camptocythere (C.) cf. lateres Tesakova et Shurupova, 2017.
Фотографирование экземпляров G. aspera и G. bathonica sp. nov. из разрезов скважин Сокурская и Обвал, а также открытой части Сокурского и Плетневского карьеров производилось в кабинете приборной аналитики ПИН РАН на сканирующем электронном микроскопе TESCAN VEGA-III. Их изображения представлены на табл. XI и XII , см. вклейку.
Для выявления полных ареалов изученных видов на РП привлечены литературные данные (Хабарова, 1955, 1961; Преображенская, 1966; Пермякова, 1970, 1975; Масумов, 1973; Колпенская, 2002; Салтыков и др., 2008 и др.), ревизованные автором настоящей работы.
Коллекции остракод хранятся на каф. региональной геологии и истории Земли МГУ им. М.В. Ломоносова (г. Москва) под №№ МГУ Sokur, МГУ Sokur-Ya и МГУ Sokur-LG (Саратовская обл., Сокурский сводный разрез), МГУ Pnz-12 (Пензенская обл., скв. Обвал) и МГУ ISSA (Пензенская обл., разрез Плетневского карьера).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Систематика надродовых таксонов принята по “Практическому руководству …” (1999) и приведена в соответствие с “Международным кодексом …” (2004), терминология различных элементов раковин остракод взята по “Практическому руководству…” (1989). При измерении параметров раковин или отдельных створок приняты следующие сокращения: Д – длина, ВПК – высота переднего конца, ВЗК – высота заднего конца, Т – толщина, Д/ВПК – отношение длины к высоте переднего конца (здесь – к максимальной высоте), Д/ВЗК – отношение длины к высоте заднего конца, juv. – ювенильная особь. В онтогенезе остракоды остаются неполовозрелыми (ювенильными) до предпоследней жизненной стадии; половая зрелость наступает после последней линьки. Счет ювенильных стадий начинают вести от предпоследней (juv. I) до самых первых (начальных) стадий жизненного цикла (juv. VIII, IX).
ОТРЯД PODOCOPIDA
ПОДОТРЯД CYTHEROCOPINA
НАДСЕМЕЙСТВО PROGONOCYTHEROIDEA SYLVESTER-BRADLEY, 1948
СЕМЕЙСТВО PROGONOCYTHERIDAE SYLVESTER-BRADLEY, 1948
ПОДСЕМЕЙСТВО PROGONOCYTHERINAE SYLVESTER-BRADLEY, 1948
Род Glyptocythere Brand et Malz, 1962
Glyptocythere aspera (Khabarova, 1955)
Табл. XI , фиг. 1–18 ; табл. XII , фиг. 1–4
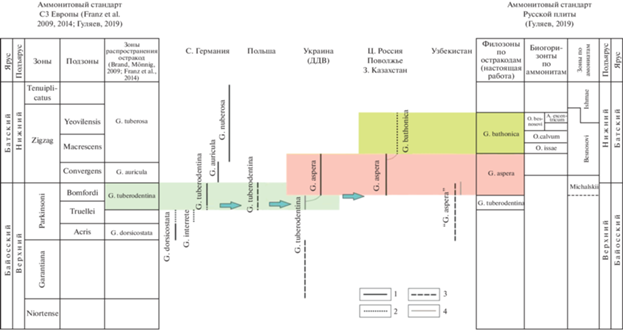
Рис. 2.
Распространение некоторых остракод рода Glyptocythere в пограничных отложениях верхнего байоса–нижнего бата Западной и Восточной Европы. Обозначения: 1 – достоверный интервал распространения вида (обильные экземпляры), 2 – достоверный интервал распространения вида (единичные экземпляры), 3 – примерный интервал распространения вида, установленный на основе литературных данных, 4 – предполагаемый интервал распространения вида.
Palaeocytheridea aspera: Хабарова, 1955, с. 196, табл. 1, фиг. 2.
Таблица 1.
Разброс линейных параметров раковин видов G. tuberodentina, G. aspera и G. bathonica sp. nov.
| Параметры раковины | G. tuberodentina | G. aspera | G. bathonica sp. nov. |
|---|---|---|---|
| Д самок, мм | 0.72–0.86 | 0.71–0.79 | 0.74–0.88 |
| Д самцов, мм | 1.00–1.09 | 0.76–0.85 | 0.84–0.88 |
| ВПК самок, мм | 0.47–0.50 | 0.40–0.49 | 0.41–0.56 |
| ВПК самцов, мм | 0.51–0.54 | 0.40–0.48 | 0.48–0.50 |
| Д/ВПК самок | 1.54–1.72 | 1.54–1.88 | 1.57–1.80 |
| Д/ВПК самцов | 1.89–2.04 | 1.73–1.93 | 1.75–1.76 |
| ВЗК самок, мм | 0.35–0.4 | 0.26–0.33 | 0.27–0.36 |
| ВЗК самцов, мм | 0.43–0.5 | 0.30–0.43 | 0.35–0.36 |
Progonocythere aspera: Преображенская, 1966, табл. 12 , фиг. 116–117 .
Glyptocythere aspera: Пермякова, 1970, с. 66, табл. 1, фиг. 5 ; 1975, с. 39; Пяткова, Пермякова, 1978, с. 140, табл. 56 , фиг. 2.
Macrodentina aspera: Масумов, 1973, с. 86, табл. 9 , фиг. 2.
Glyptocythere tuberodentina: Шурупова и др., 2016, рис. 2, 5 /13–16; Shurupova, Tesakova, 2017, рис. 3 .
Голотип – ЦНИЛ объединения “Саратовнефть” № 81, левая створка самки; Волгоградская обл., с. Жирное (ныне г. Жирновск); верхний байос (Хабарова, 1955, табл. 1, фиг. 2) [Место хранения голотипа в настоящий момент неизвестно, с высокой долей вероятности он не сохранился (Тесакова, 2022а, с. 58)].
Неотип – МГУ Sokur-Ya-138, раковина самки; Саратовская обл., Сокурская скв., гл. 42.9 м; верхний байос–нижний бат, аммонитовая зона Michalskii (табл. XI , фиг. 6 ).
Описание. Раковина средних размеров, продолговато-овальная, умеренно выпуклая, уплощенная, с параллельными боковыми сторонами. Левая створка больше правой и перекрывает ее по всему свободному краю, особенно сильно вдоль брюшного края и по переднеспинному и заднеспинному углам, в то время как правая створка перекрывает левую в средней части спинного края. Максимальная длина находится на середине высоты, максимальная высота – в передней трети, максимальная толщина – в задне-брюшной части раковины. Брюшная, наиболее выпуклая часть створки нависает над брюшным краем, закрывая его при виде снаружи. Спинной край левой створки слабо ундулирует (он выпуклый в районе замочных ушек и посередине); на правой створке – отчетливо выпуклый только в средней части (у самок сильнее, чем у самцов); а у ювенильных особей спинной край прямой на обеих створках. Брюшной край прямой, у самок и ювенилов вогнут посередине, ближе к передней трети, а у самцов – в передней трети. При виде снаружи он почти параллелен спинному краю, а при виде изнутри слабо конвергирует к заднему концу. У самцов спинной и брюшной края почти параллельны; а у самок и ювенильных особей оба эти края сходятся к заднему концу (как при виде изнутри, так и снаружи). На правой створке в вогнутой части брюшного края расположена крупная продолговатая выемка с частыми поперечными тонкими ребрышками (около двух–трех десятков) (табл. XI , фиг. 18 ; табл. XII , фиг. 1а, 2), куда входит, как в замок, утолщенная выпуклость брюшного края левой створки для обеспечения дополнительной прочности при смыкании створок. Передний конец высокий, уплощенный вдоль края, плавно дугообразно закруглен, в верхней трети на левых створках скошен слабо, на правых сильнее, с выемкой; в спинной край на левых створках переходит плавно через замочное ушко, на правых – через уступ; с брюшным краем сопрягается плавно на обеих створках. Задний конец ниже переднего, вдоль края уплощен, почти симметричный, на левых створках округло-треугольный и слабо скошен сверху, на правых – треугольный, скошен сверху с выемкой; в спинной край переходит через маленькое ушко на левой створке и через уступ на правой; в брюшной край на обеих створках переходит плавно. Скульптура развита на всей поверхности створки, кроме уплощенной части переднего и заднего концов, замочных ушек и узкой полосы вдоль спинного края. Она представлена 12–14 невысокими тонкими поперечными ребрами, которые соединяются между собой многочисленными перемычками, образуя сетчатую скульптуру из четырехгранных ячеек. Поперечные ребра, расположенные перед мускульными отпечатками, отклоняются своей нижней частью к переднему концу. Поперечные ребра сзади мускульных отпечатков параллельны друг другу, слабо выгибаются в сторону заднего конца и пересекаются в задней верхней трети створки коротким слабым продольным ребрышком. В нижней трети створки расположено четыре продольных ребра, верхнее и нижнее из которых – особенно длинные и хорошо развитые – плавно переходят в вертикальные ребра на переднем конце. Ниже них (на брюшной стороне раковины) присутствуют тонкие продольные ребрышки. Все продольные ребра также связаны перемычками.
На гладкой и скульптированной поверхности (иногда на стенках ребер, иногда внутри ячеек) развиты немногочисленные мелкие ситовидные поры. На заднем конце створки некоторые из них расположены на вершине маленьких конусов (конулей) (обычно различаются три таких конуля). Простые поры, также немногочисленные, наблюдаются на всей поверхности, в т.ч. на ребрах.
Порово-канальная зона широкая, с прямыми, редко расположенными краевыми каналами: восемь на переднем конце (табл. XII , фиг. 1в), четыре(?) на заднем (табл. XII , фиг. 1г). Отпечатки аддуктора представлены вертикальным, слабовыпуклым в сторону заднего конца рядом из четырех округлых или овальных рубцов практически одинакового размера (верхний и нижний могут иметь очертания, приближенные к округло-треугольным, и быть немного крупнее двух средних). Спереди от них выделяются два отчетливых круглых отпечатка: антеннальный (более крупный) расположен примерно на уровне середины ряда, мандибулярный (более мелкий) – ниже ряда (табл. XII , фиг. 1д).
Замки самок и самцов отличаются. Здесь приводится описание замка самки (а замок самца см. в разделе Половой диморфизм). Замок правой створки выражен массивными, крупными и удлиненными краевыми зубами, разделенными на семь длинных узких двувершинных зубцов – передний зуб, и восемь-девять таких же зубцов – задний (табл. XII , фиг. 1а, 1б). Узкие зубцы имеют тенденцию к дополнительному продольному подразделению, что более заметно на самых коротких из них, расположенных по краям заднего зуба. Средняя часть замка – желобок с широкой передней и узкой задней частями – осложнен ямкогруппами, которые комплементарны группировкам из мелких зубчиков на валике противоположной створки. Краевые части замка левой створки представлены крупными продолговатыми насеченными ямками. Задняя из них рассечена на восемь узких отделов; иногда на пять, но широких – за счет близкого расположения некоторых зубцов противоположной сворки и слияния их нижних частей. Передняя ямка рассечена на семь отделов. В расширенной передней части валика наблюдаются: один мелкий проксимальный зубчик и четыре крупных зубчика, образованных более мелкими, слившимися по два. Узкая задняя часть валика орнаментирована мелкими удлиненными зубчиками, которые объединяются по три и демонстрируют пять таких групп. Строение замка в виде зубной формулы (ЗФ) выглядит так: 7[1+4:2+5:3]8–9 где цифры спереди и сзади квадратной скобки означают число зубцов/подразделений латеральных (краевых) зубов/ямок, цифры внутри квадратной скобки отражают строение желобка/валика: первая цифра – мелкий одиночный проксимальный зубчик/ямка; первая группа цифр, разделенная двоеточием, означает четыре крупных зубчика/ямки, слившихся из двух мелких; цифры, подчеркнутые линией, означают зубчики/ямки, оконтуренные арковидным расширением валика/желобка; последняя группа цифр, разделенная двоеточием – пять более мелких дистальных зубчиков/ямок, слившихся из трех мелких.
Размеры в мм:
| Д | ВПК | ВЗК | Т | Д/ВПК | Д/ВЗК | |
|---|---|---|---|---|---|---|
| неотип Sokur- Ya-138 самка | 0.76 | 0.47 | 0.33 | – | 1.62 | 2.30 |
| экз. Sokur-Ya-111 самец | 0.80 | 0.46 | 0.43 | – | 1.74 | 1.86 |
| экз. Sokur-Ya-055 juv. VII-VIII | 0.31 | 0.18 | 0.09 | – | 1.72 | 3.44 |
| экз. Sokur-Ya-145 самец | 0.76 | 0.44 | 0.37 | – | 1.73 | 2.05 |
| экз. Sokur-Ya-075 самка | 0.76 | 0.49 | 0.31 | – | 1.55 | 2.45 |
| экз. Sokur-Ya-131 самец | 0.85 | 0.48 | 0.37 | – | 1.77 | 2.30 |
| экз. Sokur-Ya-094 juv. III-IV | 0.57 | 0.33 | 0.17 | – | 1.72 | 3.35 |
| экз. Sokur-Ya-054 juv. VII-VIII | 0.39 | 0.24 | 0.16 | – | 1.63 | 2.44 |
| экз. Sokur-Ya-137 juv. I-II | 0.67 | 0.38 | 0.22 | – | 1.76 | 3.04 |
| экз. Sokur-Ya-073 juv. V-VI | 0.46 | 0.28 | 0.14 | – | 1.64 | 3.29 |
| экз. Sokur-Ya-095 самец | 0.77 | 0.40 | 0.30 | – | 1.93 | 2.57 |
| экз Sokur-Ya-093 juv. I-II | 0.63 | 0.38 | 0.20 | – | 1.66 | 3.15 |
| экз. Sokur-Ya-139 juv. V-VI | 0.50 | 0.29 | 0.14 | – | 1.72 | 3.57 |
| экз. Sokur-Ya-146 самка | 0.71 | 0.46 | 0.28 | – | 1.54 | 2.54 |
| экз. Sokur-Ya-107 самка | 0.71 | 0.40 | 0.26 | – | 1.77 | 2.73 |
| экз. Sokur-Ya-053 самец | 0.81 | 0.46 | 0.32 | – | 1.76 | 2.53 |
| экз. Sokur-Ya-228 juv. V-VI | 0.48 | 0.30 | 0.17 | – | 1.60 | 2.83 |
| экз. Sokur-51 самка | 0.79 | 0.42 | 0.26 | – | 1.88 | 3.04 |
| экз. Sokur-139 самка | 0.78 | 0.44 | 0.27 | – | 1.77 | 2.89 |
| экз. Sokur-140 juv. I-II | 0.64 | 0.36 | 0.18 | – | 1.78 | 3.56 |
| экз. Sokur-70 juv. V-VI | 0.45 | – | – | 0.21 | – | – |
| экз. Sokur-Ya-147 самка | 0.75 | – | – | 0.34 | – | – |
Изменчивость. Длина раковин изученных самок слабо варьирует в диапазоне 0.71–0.79 мм, у самцов сильнее – от 0.76 до 0.85 мм; соотношение Д/В у самок составляет 1.54–1.88, а у самцов 1.73–1.93 (табл. 1). Незначительно меняется степень выраженности короткого продольного ребрышка в задней верхней трети створки. Задний краевой элемент замка может быть рассечен на пять–восемь частей.
Онтогенез. С каждой следующей возрастной стадией возрастают такие параметры раковины, как ее длина, высота переднего и заднего концов. При достижении половой зрелости у самок эти показатели стабилизируются, а их разброс становится незначительным (для Д в пределах 0.08 мм, для ВПК – 0.09, а для ВЗК – 0.07) (табл. 1). У взрослых самцов разброс параметров Д – 0.09 мм и ВПК – 0.08 очень схож с таковым у самок, в то время как вариация ВЗК – 0.13 вдвое превышает таковую у самок. Также педоморфизм самцов проявляется в пределах разброса параметра Д/В: если у ювенильных особей он составляет 1.60–1.78, самок 1.54–1.88 и самцов 1.73–1.93, то пределы разброса составляют у неполовозрелых 0.18, у самок 0.34, а у самцов 0.20 – что сходно с ювенильными особями.
Переднее замочное ушко низкое у всех ювенильных стадий, а высоким оно становится только на поздней – половозрелой – стадии. Скульптура ювенильных раковин (табл. XI , фиг. 2, 7–10, 12, 13, 17 ; табл. XII , фиг. 2, 3 ) отличается преимущественным развитием поперечных ребер, что делает ее скорее поперечно-ребристой, нежели сетчатой, как у взрослых самок. Продольные перемычки между ребрами становятся все более заметными с каждой следующей линькой. Короткое продольное ребрышко, объединяющее поперечные ребра в задней верхней части створки, лучше всего проявлено на самых ранних возрастных стадиях, а по мере взросления нивелируется и слабо различается на раковинах как самок, так и самцов.
Половой диморфизм. Самцы (табл. XI , фиг. 1, 3, 5, 11, 16 ) отличаются от самок (табл. XI , фиг. 4, 6, 14, 15, 18 ) более крупными и длинными раковинами, значительно более высоким задним концом (0.26–0.33 мм у самок, 0.30–0.43 мм у самцов) и низким передним замочным ушком левой створки. Отличия в замке самцов выражены более длинной узкой дистальной частью среднего элемента, на котором располагается бóльшее, чем у самок, число ямкогрупп/зубчиков, сгруппированных по три. Кроме того, в скульптуре самцов поперечные ребра развиты сильнее продольных перемычек, что делает ее скорее поперечно-ребристой, нежели сетчатой, как у самок. Низкое замочное ушко и слабое развитие продольных перемычек, как и разброс размерных параметров раковин (см. раздел Онтогенез), характерны для ювенильных стадий этого вида, т.е., развитие самцов шло по пути педоморфоза.
Сравнение. От видов, более всего похожих по форме раковины и скульптуре из поперечных ребер, пересекающихся двумя продольными ребрами в брюшной части створки и/или одним в спинной/заднеспинной части створки, G. aspera отличается следующим. От G. costata Bate, 1965 из нижнего байоса С.-В. Англии (остракодовая зона G. polita по: Bate, 1965; аммонитовая зона Sauzei по: Bate, 2009) (Bate, 1965, с. 106, табл. 5 , фиг. 1–7 ) отличается существенно более коротким и округлым задним концом, высоким передним ушком на левых створках самок, бóльшим развитием продольных перемычек (что делает скульптуру G. aspera скорее сетчатой, нежели поперечно ребристой, что свойственно G. costata) и наличием тонкого и извилистого продольного ребрышка в заднеспинной части створки (против плохо различимого или отсутствующего у G. costata).
От G. dorsicostata Brand et Malz in Brand et Fahrion, 1962 из верхов верхнего байоса (аммонитовая зона Parkinsoni) Сев. Германии (Brand, Fahrion, 1962, с. 145, табл. 21 , фиг. 10 ; Brand, Malz, 1966, с. 511, табл. 58 , фиг. 76–81 ; Luppold, 2012, с. 234, табл. 6 , фиг. 11 ), а в Юж. Германии известного с начала верхнего байоса (аммонитовая зона Niortense) по нижний бат (аммонитовая зона Zigzag) (Franz et al., 2009, с. 139, табл. 3 , фиг. 10, 11 ), отличается более высоким задним концом, высоким замочным ушком на левых створках (особенно у самок), слабо развитым и коротким продольным ребрышком в заднеспинной части створки (против более длинного и сильного спинного ребра у G. dorsicostata), сетчатой скульптурой, а не поперечно ребристой, как у G. dorsicostata, и отсутствием в передней половине створки короткого горизонтального вилочковидного ребра.
От G. auricula Brand et Malz in Brand et Fahrion 1962 из нижнего бата (зона Zigzag) С.-З. Германии (Brand, Fahrion, 1962, с. 143, табл. 18 , фиг. 5 , табл. 21 , фиг. 16 ; Brand, Malz, 1966, с. 516, рис. 16–18 , табл. 58 , фиг. 82, 83 ) отличается высоким передним замочным ушком на левых створках самок и сетчатой скульптурой (против поперечно-ребристой у G. auricula).
От наиболее похожего по сетчатой скульптуре вида G. tuberodentina Brand et Malz in Brand et Fahrion, 1962 из верхнего байоса С.-З. Германии (верхняя часть зоны Parkinsoni) (Brand, Fahrion, 1962, с. 143, табл. 21 , фиг. 11, 12 ; Brand, Malz, 1966, с. 485, табл. 52 , фиг. 1–13 , табл. 53 , фиг. 14, 15 ), Польши (средний “Kujavian”, зоны Parkinsonia parkonsoni и P. schloenbachi) (Bielecka, Styk, 1969, c. 622, табл. 2 и Украины (аммонитовая зона Garantiana11) (Пермякова, 1975, с. 37; Пяткова, Пермякова, 1978, с. 142, табл. 58 , фиг. 3 ) отличается меньшими размерами раковин (0.72–0.86 мм у самок и 1.00–1.09 мм у самцов G. tuberodentina). Кроме того, раковины самок сравниваемых видов отличаются гораздо сильнее, чем таковые самцов (табл. 1). Самки G. aspera отличаются менее выпуклой и более удлиненной раковиной (Д/В у G. aspera > Д/В у G. tuberodentina), более высоким замочным ушком на левых створках, более длинным спинным краем и, следовательно, более длинной узкой (проксимальной) частью среднего элемента замка: ЗФ самок G. aspera 7[1+4:2+5:3]8–9 против 6-7[1+6:2+5]7–8 у самок G. tuberodentina (Brand, Malz, 1962).
Замечания. Распространение G. aspera в узком стратиграфическом интервале зоны Michalskii зафиксировано в Днепровско-Донецкой впадине (ДДВ) (Пермякова, 1975, с. 39), а в Поволжье – в зонах Michalskii и нижней части Besnosovi (ниже биогоризонта O. issae) (Шурупова и др., 2016, рис. 2; Shurupova, Tesakova, 2017, рис. 3 ). В публикациях XX в. зона Michalskii считалась нижнебатской (Пермякова, 1970, 1975), позже была перенесена в верхний байос (Унифицированная …, 2012), а в настоящей работе рассматривается отвечающей верхней части верхнего байоса (референсный биогоризонт P. michalskii)–нижней части нижнего бата (референсный биогоризонт P. mazarowichi) (Гуляев, 2019). На обширной территории Нижнего Поволжья и Зап. Казахстана находки G. aspera упоминались из аммонитовой зоны Parkinsonia doneziana (Хабарова, 1955, 1961). Также этот таксон известен в Узбекистане из отложений зоны Parkinsonia parkinsoni (Масумов, 1973). Во всех упомянутых публикациях отмечается высокое насыщение остатками G. aspera этих пород, как и то, что в вышележащих отложениях нижнего бата они присутствуют очень редко (кроме Узбекистана, где в верхнем байосе отмечены единичные экземпляры, а в нижнем бате их нет совсем).
Батские представители из верхней части зоны Besnosovi (биогоризонты O. issae–A. excentricum) выделены нами в новый вид G. bathonica sp. nov. (см. ниже).
Распространение. Верхний байос – нижний бат, зона Michalskii, ДДВ (Пермякова, 1970, 1975); зона Michalskii – нижняя часть зоны Besnosovi (референсные биогоризонты O. mojarowskii и O. scythicum) Центральной России (Курская обл.) (Преображенская, 1964, 1966), Нижнего Поволжья (Саратовская, Волгоградская обл.) и Зап. Казахстана (Хабарова, 1955, 1961; Салтыков и др., 2008; Шурупова и др., 2016), а также зона Parkinsoni Узбекистана (Масумов, 1973).
Материал. 130 целых раковин и отдельных створок хорошей и удовлетворительной сохранности самок, самцов и ювенильных особей из верхнего байоса–нижнего бата, зона Michalskii и нижняя часть зоны Besnosovi (референсные биогоризонты O. mojarowskii и O. scythicum) Сокурской скважины.
Glyptocythere bathonica Tesakova, sp. nov.
Табл. XII , фиг. 5–13
Glyptocythere aff. tuberodentina: Shurupova, Tesakova, 2017, рис. 3 .
Название вида – по появлению таксона с начала батского века.
Голотип – МГУ SOK-LG-02, раковина самки; Саратовская обл., Сокурский карьер, обр. 12f; нижний бат, верхняя часть аммонитовой зоны Besnosovi, биогоризонт O. besnosovi (расчленение разреза по: Гуляев, 2019) (табл. XII , фиг. 5 а, 5б).
Описание. Раковина средних размеров, продолговато-овальная, умеренно выпуклая, уплощенная, с параллельными боковыми сторонами (табл. XII , фиг. 6 б). Левая створка больше правой и перекрывает ее по брюшному краю, переднеспинному и заднеспинному углам, а правая створка перекрывает левую в средней части спинного края (табл. XII , фиг. 5 а, 6б). Максимальная длина находится на середине высоты, максимальная высота – в передней трети, максимальная толщина – в заднебрюшной части раковины. Брюшная, наиболее выпуклая часть створки нависает над брюшным краем. Спинной край левой створки ундулирующий (выпуклый в районе переднего замочного ушка и посередине); на правой створке – отчетливо выпуклый в средней части. Брюшной край – при виде изнутри – прямой, вогнут слабо (у самок и ювенильных особей примерно посередине, а у самцов – в передней трети) и слабо конвергирует к заднему концу. При виде снаружи он кажется слабо выпуклым и почти параллелен спинному краю. У самцов спинной и брюшной края параллельны, у самок конвергируют к заднему концу, но очень слабо; а у ювенильных особей оба эти края сходятся к заднему концу (как при виде изнутри, так и снаружи). На правой створке в вогнутой части брюшного края расположена крупная продолговатая выемка с частыми поперечными тонкими ребрышками, куда входит, как в замок, утолщенная выпуклость брюшного края левой створки. Передний конец высокий, уплощенный вдоль края, плавно дугообразно закруглен, в верхней трети на левых створках скошен слабо, на правых сильнее, с выемкой; в спинной край на левых створках переходит плавно через замочное ушко, на правых – через уступ; с брюшным краем сопрягается плавно на обеих створках. Задний конец почти равной высоты с передним (у самок; у самцов он ниже переднего), вдоль края уплощен, округло-треугольный, симметричный, немного сильнее скошен сверху на правых створках; в спинной край на обеих створках переходит через уступ, а в брюшной край – плавно. Скульптура из продольных и поперечных ребер более всего похожа на таковую G. aspera, но более слабое развитие соединительных перемычек приводит к формированию ребристой скульптуры. Простые и ситовидные поры, порово-канальная зона, мускульные отпечатки и замок такие же, как у G. aspera.
Размеры в мм:
| Д | ВПК | ВЗК | Т | Д/ВПК | Д/ВЗК | |
|---|---|---|---|---|---|---|
| голотип SOK-LG-02 самка | 0.88 | 0.55 | 0.36 | – | 1.60 | 2.44 |
| экз. SOK-LG-01 самка | 0.88 | 0.56 | 0.36 | 0.44 | 1.57 | 2.44 |
| экз. PNZ-12-37 самец | 0.88 | 0.50 | 0.36 | – | 1.76 | 2.44 |
| экз. SOK-LG-43 самка | 0.83 | 0.52 | 0.36 | – | 1.69 | 2.31 |
| экз. SOK-LG-48 самка | 0.75 | 0.46 | 0.32 | – | 1.63 | 2.34 |
| экз. ISSA-01 самка | 0.74 | 0.41 | 0.27 | – | 1.80 | 2.74 |
| экз. SOK-LG-49 самка | 0.75 | 0.42 | 0.27 | – | 1.79 | 2.78 |
| экз. PNZ-12-02 самец | 0.84 | 0.48 | 0.35 | – | 1.75 | 2.40 |
| экз. PNZ-12-36 самка | 0.83 | 0.51 | 0.34 | – | 1.63 | 2.44 |
Изменчивость. Длина раковины у самок может меняться в пределах 0.74–0.88 мм, у самцов 0.84–0.88 мм, а Д/В у самок составляет 1.57–1.80, у самцов 1.75–1.76. На левых створках, как самцов, так и самок, выпуклость спинного края может варьировать от округлой (табл. XII , фиг. 5 б, 8, 9, 12) до угловатой (табл. XII , фиг. 6 а, 7).
Половой диморфизм. У самок длина раковины в среднем меньше, чем у самцов. Переднее замочное ушко на левых створках самок (табл. XII , фиг. 5 б, 6а, 8, 9, 13) существенно выше, чем у самцов (табл. XII , фиг. 7, 12 ). Передний конец у самок практически симметричный и равномерно дугообразно закруглен, против скошенного сверху у самцов. Интересно, что ВЗК у самцов (0.35–0.36 мм) и самок (0.27–0.36 мм) этого вида находятся в одном диапазоне.
Сравнение. От наиболее похожего по форме раковины, размерам и скульптуре G. aspera отличается большей длиной раковины, более высоким задним концом самок, примерно равной ВЗК самцов и самок (что нетипично для представителей этого рода), более угловатым спинным краем, превалирующим развитием поперечных ребер над перемычками (т.е. поперечно-ребристой скульптурой, в отличие от сетчатой у G. aspera).
От сходного по ребристой скульптуре G. auricula из нижнего бата (зона Zigzag) северо-западной Германии отличается высоким передним замочным ушком на левых створках самок, а также слабым коротким заднеспинным ребрышком против сильного и длинного спинного у G. auricula.
Замечания. Изученный материал датирован по аммонитам в разрезах Сокурского и Плетневского карьеров. В Плетневском карьере остракоды были встречены только в зоне Besnosovi в биогоризонте O. issae, а в других стратиграфических интервалах не найдены. В Сокурском карьере они известны из отложений, отнесенных В.В. Митта к аммонитовым зонам Besnosovi и Ishmae (Mitta et al., 2014; Shurupova, Tesakova, 2017), а Гуляевым к зоне Besnosovi, биогоризонтам O. besnosovi и A. excentricum (Гуляев, 2019).
В нижнем бате Курской (Преображенская, 1966), Белгородской (Колпенская, 2002), Саратовской, Волгоградской областей и Зап. Казахстана (Хабарова, 1955, 1961; Салтыков и др., 2008) отмечались редкие находки вида G. aspera s.l. Но в результате сравнительно-морфологического и онтогенетического анализов, выполненных автором настоящей статьи, удалось подразделить этот вид на два – G. aspera и G. bathonica sp. nov., связанных отношением предок–потомок и характеризующих разные стратиграфические уровни верхнего байоса и нижнего бата. В связи с этим, упоминание в литературе редких находок “G. aspera” в нижнем бате следует понимать, как присутствие в этом интервале вида G. bathonica sp. nov.
Распространение. Нижний бат, верхняя часть зоны Besnosovi, биогоризонты O. issae–A. excentricum, РП (по: Гуляев, 2019): Курская, Пензенская, Белгородская, Саратовская, Волгоградская области и Западный Казахстан.
Материал. 18 раковин и отдельных створок хорошей и удовлетворительной сохранности самок и самцов из нижнего бата открытой части Сокурского карьера, верхняя часть аммонитовой зоны Besnosovi (биогоризонты O. besnosovi и A. excentricum); 64 экз. самок и самцов (отдельные створки и целые раковины) хорошей и удовлетворительной сохранности из нижнего бата, остракодовая зона G. bathonica, скв. Обвал; один экз. удовлетворительной сохранности из нижнего бата, зона Besnosovi, биогоризонт O. issae (по: Гуляев, 2019) из разреза Плетневского карьера.
ЭВОЛЮЦИЯ
Морфологическое сходство, обусловленное эволюционной преемственностью, в сочетании с непрерывной стратиграфической последовательностью, позволяет реконструировать филолинию G. tuberodentina (b2, зона Parkinsoni) → G. aspera (bj2-bt1, зона Michalskii – нижняя часть зоны Besnosovi, референсные биогоризонты O. mojarow-skii и O. scythicum) → G. bathonica sp. nov. (bt1, верхняя часть зоны Besnosovi, биогоризонты O. issae–A. excentricum), где два последних вида развивались в обособленных акваториях Восточной Европы как потомки G. tuberodentina, вселившегося на РП из Западной Европы в позднем байосе. Рассмотрим основания для выделения филолинии, особенности морфогенеза и эволюции полов.
Скульптура. У всех глиптоцитер этой филолинии скульптура представлена продольными и поперечными ребрами, которые развивались на базе первичных ячеек. У каждого вида на ранних онтогенетических стадиях выделяются хорошо сформированные поперечные ребра, в то время как продольные представлены тонкими стеночками первичных ячеек и начинают развиваться позже – сначала как перемычки между поперечными, потом как продольные ребра. В онтогенезе G. tuberodentina поперечные ребра наиболее ярко выражены у ювенильных особей (Brand, Malz, 1966, табл. 52 , фиг. 6, 9 ) и самок (Brand, Malz, 1966, табл. 52 , фиг. 3, 5, 7 ). Продольные межреберные перемычки, весьма слабые у ювенильных особей, у самок развиты уже настолько хорошо, что их скульптура может казаться не только продольно ребристой, но иногда и сетчатой (Brand, Malz, 1966, табл. 52 , фиг. 10, 11 ). Наибольшее развитие перемычек у этого вида наблюдается у самцов, поэтому их скульптура в основном сетчатая (Brand, Malz, 1966, табл. 52 , фиг. 4, 13b ). У дочернего таксона G. aspera дальнейшее усиление перемычек демонстрирует их максимальную высоту и толщину у самок (табл. XI , фиг. 4, 6, 14, 15 ), сетчатая скульптура которых приблизилась к таковой самцов G. tuberodentina. У самцов же G. aspera наблюдается регрессивный тренд развития этого признака – их скульптура становится, скорее, ребристой, сходной с ювенильной (табл. XI , фиг. 1, 5, 11, 16 ). Вслед за самцами предкового вида, самки G. bathonica sp. nov. вторично приобрели ребристую скульптуру, и теперь скульптура обоих полов у этого вида сравнялась.
То, что наблюдается в эволюции скульптуры данной филолинии [первоначальное усиление признака (развитие продольных ребер) с последующей редукцией] описано Е.И. Шорниковым как “цикличность морфогенеза” (Шорников, Михайлова, 1990, с. 75–79). Более раннее проявление этапов этого цикла у самцов свидетельствует о разных темпах эволюции полов у этих видов.
Высокое сходство скульптуры взрослых представителей G. bathonica, как самцов (табл. XII , фиг. 7, 12 ), так и самок (табл. XII , фиг. 5, 6, 8–11, 13 ), с таковой неполовозрелых представителей G. aspera (табл. XI , фиг. 7, 9, 10, 12, 13 ) и G. tuberodentina (Brand, Malz, 1966, табл. 52 , фиг. 6, 9 ) является еще одним доказательством преемственности в реконструированной филолинии.
Форма и размер раковины. Во время миграции представителей G. tuberodentina в Восточную Европу, в результате экологического стресса произошло уменьшение размеров представителей вида-потомка (эффект лилипутов), что особенно сильно сказалось на самцах (табл. 1). У самцов G. aspera уменьшение произошло значительно сильнее, чем у самок, что привело к сближению размеров и контура раковин полов. Последующее возрастание этих параметров у G. bathonica sp. nov. также происходило у самок и самцов с разной скоростью (у первых быстрее, чем у вторых). В результате, сглаживание половых различий оказалось наиболее ярко выражено у последнего члена филолинии – G. bathonica sp. nov., что крайне затрудняет различение самок и самцов этого вида в ископаемом состоянии.
В табл. 1 продемонстрирован высокий диапазон вариации Д/ВПК самцов, против низкого самок. Другими словами, самцы сильнее подвержены разбалансировке старой видовой нормы, поэтому эволюционируют быстрее, чем самки (еще одно подтверждение опережающей эволюции самцов этой филолинии). Кроме того, эта таблица показывает, что при переходе от G. tuberodentina к G. aspera у самок увеличилась относительная длина раковины, что было более свойственно самцам, нежели самкам предка. Соответственно, у самок G. aspera и G. bathonica sp. nov. удлинился спинной край, что привело к трансформации замка.
Замок. При переходе от G. tuberodentina (ЗФ 6-7[1+6:2+5]7–8) к G. aspera (ЗФ 7[1+4:2+5:3]7–8) (табл. XII , фиг. 1а, 1б) у самок увеличилась длина спинного края, вследствие чего увеличился передний краевой зуб и появилось высокое замочное ушко над приемной ямкой на левых створках; также существенно возросла длина валика/желобка, а осложняющие его элементы укрупнились за счет удвоения или утроения своих составляющих. У самок наиболее молодого вида G. bathonica sp. nov. (ЗФ 7[1+4:2+5:3]7–8) (табл. XII , фиг. 10 б) дальнейшая эволюция замка, по-видимому, не происходила (не наблюдалась). Другими словами, сближение морфологии замков самок и самцов в этой филолинии произошло при первом эволюционном преобразовании (в период миграции западноевропейского таксона на РП) и приостановилось в эволюции потомков восточной группы.
Таким образом, развитие этой филолинии сопровождалось опережающей эволюцией самцов путем педоморфоза, а самки каждого следующего вида при эволюции скульптуры, замка и линейных параметров раковин изменялись вслед за самцами предков. У самцов контур и линейные размеры раковин эволюционировали с ретардацией, поэтому со временем сближались с таковыми самок и практически сравнялись у полов самого молодого вида G. bathonica sp. nov. Другими словами, эволюция полового диморфизма сначала привела к снижению его выраженности у промежуточного таксона G. aspera, а у последнего члена филолинии почти к нивелировке. Наиболее крупное эволюционное событие произошло при миграции западноевропейского G. tuberodentina на РП и возникновении G. aspera; оно выразилось в эффекте лилипутов и перестройке замка. Впоследствии замок не менялся, а уменьшение размеров сохранялось у самцов следующего члена филолинии на фоне их роста у самок.
СТРАТИГРАФИЯ
В отложениях байоса и бата Сев. Германии были распространены морфологически сходные и стратиграфически сменяющие друг друга виды (возможно, связанные филогенетическим родством), по распространению которых Э. Бранд и Э. Мённинг выделили биозоны (зоны распространения вида): Glyptocythere dorsicostata Brand et Malz in Brand et Fahrion, 1962 (bj2, зона Parkinsoni, подзона Acris), G. tuberodentina (bj2, зона Parkinsoni, верхняя часть подзоны Truellei и подзона Bomfordi), G. auricula Brand et Malz in Brand et Fahrion, 1962 (bt1, зона Zigzag, подзона Convergens) и G. tuberosa Brand et Malz in Brand et Fahrion, 1962 (bt1, зона Zigzag, подзоны Macrescens, Yeovilensis и аммонитовая зона Tenuiplicatus) (рис. 2) (Brand, Mönnig, 2009; Franz et al., 2014).
Восточнее, в Польше, глиптоцитеры появились лишь начиная с позднего байоса и сформировали здесь совершенно другие эндемичные виды (Bielecka et al., 1988). Однако из средней части местного подразделения “Kujavian” (зоны Parkinsonia parkonsoni и P. schloenbachi) был определен немецкий таксон G. tuberodentina (Bielecka, Styk, 1969, c. 622, табл. 2), что позволяет проследить в Центральной Европе соответствующую зону (рис. 2).
В ДДВ нахождение G. tuberodentina в зоне Garantiana (Пермякова, 1975, с. 37; Пяткова, Пермякова, 1978, с. 142) (рис. 2) кажется проблематичным, поэтому в первую очередь требуется переизучение украинских аммонитов для уточнения стратиграфии. Экземпляр самца, изображенный в (Пяткова, Пермякова, 1978, табл. 58 , фиг. 3 ), отвечает описанию вида в полной мере и не вызывает сомнений в определении. Но проникновение G. tuberodentina на Украину не могло произойти раньше второй половины фазы Parkinsoni, т.к. в ходе позднебайосской трансгрессии, облегчившей миграции остракод между акваториями Северо-Западной, Центральной и Восточной Европы, широкое (панъевропейское) распространение получили лишь те остракоды (некоторые виды соответствующих фаун), которые существовали именно в это время. Однако уже в терминальном байосе – раннем бате, в фазе Michalskii, на Украине получил распространение дочерний таксон G. aspera, а представители G. tuberodentina перестали встречаться (Пермякова, 1975, с. 39). Таким образом, интервал разреза ДДВ с остатками G. tuberodentina должен отвечать нижней части западноевропейской остракодовой зоны G. tuberodentina и верхней части зоны Parkinsoni. Зоне Michalskii соответствует восточноевропейская остракодовая филозона G. aspera (рис. 2).
Эволюция этой филолинии продолжилась в Среднерусском море в раннем бате и завершилась последним членом G. bathonica sp. nov., поэтому в верхней части зоны Besnosovi (биогоризонты O. issae–A. excentricum) следует выделить соответствующую филозону. Восточнее – в Центральной России, Поволжье и Западном Казахстане – прослежены только две верхние филозоны, G. aspera и G. bathonica, а зона G. tuberodentina не установлена (рис. 2).
В опорном Сокурском разрезе вид G. aspera обильно представлен в верхнебайосских слоях с Camptocythere (C.) lateres и слоях с C. (C.) angustius и не встречен в разрезе выше уровня 10.5 м (Shurupova, Tesakova, 2017). Между его последними находками и первым появлением единичных экземпляров дочернего таксона имеется слепой интервал (примерно в 10 м), не охарактеризованный представителями этой филолинии. Остатки G. bathonica sp. nov. встречены в Сокурском разрезе в зоне Besnosovi, начиная с уровня обр. 12f (маркировка и позиция образца дается по: Mitta et al., 2014). Этот образец отобран между биогоризонтами Oraniceras calvum (=Oraniceras sp. B) и O. besnosovi, судя по статье (Shurupova, Tesakova, 2017), и непосредственно в биогоризонте O. besnosovi, судя по публикации (Гуляев, 2019). В более древних отложениях зоны Besnosovi, сопоставленных с биогоризонтами O. calvum и O. issae (=Oraniceras sp. A) (Гуляев, 2019), находок G. bathonica sp. nov. не было. Гиатус между последними находками G. aspera и первыми – G. bathonica в Сокурском разрезе объясняется неполнотой материала (недостаточным объемом проб из керна скважины). А в Плетневском карьере G. bathonica sp. nov. был найден в зоне Besnosovi в биогоризонте O. issae.
Следует отметить значительное обилие и частоту находок G. aspera в отложениях зон Michalskii – нижней части Besnosovi на обширной территории РП: ДДВ (Пермякова, 1970, 1975), центральные районы России (Преображенская, 1966; Колпенская, 2002), Нижнее Поволжье (Саратовская, Волгоградская обл.) и Зап. Казахстан (Хабарова, 1955, 1961; Салтыков и др., 2008; Shurupova, Tesakova, 2017), а также Узбекистан (Масумов, 1971). Тогда как G. bathonica sp. nov. представлен в более мелководных фациях верхней части зоны Besnosovi на той же площади (кроме Узбекистана и, вероятно, ДДВ) единичными экземплярами на отдельных уровнях разреза, о чем говорится во всех вышеперечисленных публикациях (с поправкой на то, что новый таксон ошибочно определялся в ранних публикациях, как G. aspera).
В результате стратиграфического анализа двух родственных филолиний остракод рода Glyptocythere (рис. 2) получены следующие выводы. Зона распространения вида G. tuberodentina, характеризующая верхний байос (зона Parkinsoni, верхняя часть подзоны Truellei и подзона Bomfordi) Сев. Германии, прослежена в Польше и на Украине. На Украине объем зоны G. tuberodentina сокращен за счет выделения новой остракодовой зоны в зоне Michalskii.
На основе филолинии восточноевропейской ветви изученных глиптоцитер предлагаются новые филозоны для территории РП. Филозона G. tuberodentina характеризует верхний байос, зону Parkinsoni ДДВ; зона G. aspera (верхний байос–нижний бат, зоны Michalskii–нижняя часть Besnosovi) прослежена на Украине, в Центральной России, Поволжье и Зап. Казахстане; зона G. bathonica (нижний бат, верхняя часть зоны Besnosovi) установлена в Центральной России, Поволжье и Зап. Казахстане.
ОПИСАНИЕ СТРАТОНОВ
Зона Glyptocythere aspera Tesakova, nov. (филозона)
Вид-индекс. G. aspera (Khabarova, 1955).
Состав комплекса. Кроме зонального вида-индекса, известно большое число других таксонов: Procytheridea? bajociensis (Khabarova, 1955), Procytherura iyae Tesakova, 2022, P. sokolovi (Lyub., 1955), Plumhoffia tricostata (Khab., 1955), Camptocythere (C.) lateres Tesakova et Shurupova, 2017, C. (C.) angustius Tes. et Shur., 2017, C. (A.) praemuricata Tes., 2022, Aaleniella franzi Tes., 2022, A. volganica Tes., 2022, A.? ovoidea Tes., 2022, Palaeocytheridea (P.) kalandadzei Tes., 2013, Nanacythere octum Tes., 2022, N. sp. 2, Acrocythere sokurensis Tes., 2022, Glyptocythere praerimosa (Khab., 1955), G. strigatus (Khab., 1955), G. tuberosa (Khab., 1955), Paracypris bajociana Bate, 1963, Trachycythere peculiaris Tes., 2022, Pseudohutsonia clivosa (Khab., 1955), P. cf. subtilis (Oertli, 1959) , Ljubimovella? sp. 1.
Самые обильные и часто встречающиеся виды: G. aspera, Pr.? bajociensis, P. iyae, Pl. tricostata, C. (C.) lateres, C. (A.) praemuricata и Pal. (P.) kalandadzei.
Стратотип: разрез скважины Сокурская (Саратовская обл.), серые и голубовато-серые глины починковской свиты (Унифицированная …, 2012, лист 12), интервал 43.0–10.5 м в пачках I и II (по: Mitta et al., 2014, рис. 4 ; Shurupova, Tesakova, 2017, рис. 3 ). Мощность 32.5 м.
Границы: нижняя – по появлению вида-индекса; верхняя – по появлению вида-потомка.
Замечания. (1) Для зоны характерно высокое разнообразие глиптоцитер (G. aspera, G. praerimosa, G. strigatus и G. tuberosa) и аалениелл (A. franzi, A. volganica и A.? ovoidea).
(2) Только в этой зоне были встречены виды G. praerimosa, G. tuberosa, Ps. сlivosa, P. subtilis и A. sokurensis, не перешедшие в более высокие горизонты бата. Перечисленные таксоны могут служить дополнительными маркерами этого стратиграфического интервала.
(3) Обилие экземпляров G. aspera в отложениях зоны Michalskii позволило считать его одной из руководящих форм для верхнего байоса–нижнего бата Центральной России (Преображенская, 1964, 1966; Колпенская, 2002), но никакие стратиграфические подразделения выделены не были, поэтому синонимика отсутствует.
Стратиграфическое положение: верхний байос–нижний бат, аммонитовые зоны Michalskii–нижняя часть Besnosovi (референсные биогоризонты O. mojarowskii, O. sciticum) (настоящая работа); фораминиферовая зона Lenticulina volganica–Vaginulina dainae; а также верхняя часть остракодовых слоев с Camptocythere (C.) lateres, слои с Pseudohutsonia и с C. (C.) angustius и средняя часть остракодовой филозоны P. (M.) parabakirovi (по: Shurupova, Tesakova, 2017).
Распространение. Прослеживается на РП повсеместно (ДДВ, Центральная Россия, Поволжье), в Зап. Казахстане и, возможно, в Узбекистане.
Зона Glyptocythere bathonica Tesakova, nov. (филозона)
Вид-индекс. G. bathonica Tesakova, sp. nov.
Состав комплекса. Кроме зонального вида-индекса, Camptocythere (A.) arangastachiensis Nikitenko, 1994, Fuhrbergiella (P.) kizilkaspakensis (Mandelstam, 1947) и Glyptocythere aff. praerimosa, присутствующих только в этой зоне, известны таксоны, перешедшие в нее из зоны G. aspera: Procytheridea? bajociensis, Plumhoffia tricostata, Camptocythere (C.) lateres, C. (A.) praemuricata, Aaleniella franzi, A. volganica, A.? ovoidea, Palaeocytheridea (P.) kalandadzei, Glyptocythere tuberosa, G. strigatus, Paracypris bajociana.
Стратотип: разрез открытой части Сокурского карьера (Саратовская обл.), светло-серые и серые глины починковской свиты (Унифицированная …, 2012, лист 12), пачка III (по: Mitta et al., 2014, рис. 4 ; Shurupova, Tesakova, 2017, рис. 3 ). Мощность около 8 м.
Границы: нижняя – по появлению вида-индекса; верхняя – по его исчезновению.
Замечания. (1) Для зоны характерно низкое таксономическое разнообразие и низкая численность всех представителей.
(2) Вид-индекс был встречен в зоне Besnosovi в отложениях биогоризонтов O. issae (Плетневский карьер), O. besnosovi и A. excentricum (Сокурский карьер).
(3) Появление на РП бореального вида C. (A.) arangastachiensis из верхней части нижнего байоса–низов нижнего бата севера Сибири и севера Европейской России (Басов и др., 2009; Никитенко, 2009) маркирует миграционное событие, связанное с трансгрессией из бореального бассейна.
Стратиграфическое положение: нижний бат, верхняя часть аммонитовой зоны Besnosovi, биогоризонты O. issae–A. excentricum (настоящая работа); фораминиферовая зона Trochammina jakovlevae (=Trochammina aff. praesquamata, по: Shurupova, Tesakova, 2017); а также верхняя часть остракодовой зоны C. (A.) arangastachiensis (по: Никитенко, 2009).
Распространение. В нижнем бате РП: Курская, Пензенская, Белгородская, Саратовская, Волгоградская области и Зап. Казахстан.
ЗАКЛЮЧЕНИЕ
Проведена ревизия вида Glyptocythere aspera (Khabarova, 1955), описанного из верхнего байоса–нижнего бата Волгоградской обл. и распространенного в этом интервале повсеместно на РП, в Западном Казахстане и, возможно, Узбекистане; выделен неотип и сделано расширенное актуализированное переописание таксона. Вид G. bathonica из нижнего бата РП описан как новый.
Сравнительно-морфологический анализ с учетом онтогенетических данных позволил реконструировать на РП филолинию: G. tuberodentina (bj2, зона Parkinsoni) → G. aspera (bj2-bt1, зона Michalskii–нижняя часть зоны Besnosovi) → G. bathonica (bt1, верхняя часть зоны Besnosovi), которая развивалась в акваториях Украины и европейской России как потомки G. tuberodentina, проникшего на РП из Северной Германии через Польшу.
Интересно, что в этой филолинии происходила опережающая эволюция самцов (путем педоморфоза), а самки каждого следующего таксона приобретали новую гетерохронию вслед за самцами предкового таксона. Эволюционировали следующие признаки: контур и размеры раковины, замок и скульптура. У самцов эволюция проходила с ретардацией, поэтому морфологическое различие между полами со временем уменьшалось, пока не стало минимальным у G. bathonica sp. nov. Другими словами, в развитии этой филолинии наблюдается паттерн снижения полового диморфизма. Наиболее значимое эволюционное событие произошло при переходе от G. tuberodentina к G. aspera, при котором дочерний таксон, кроме изменений в скульптуре, претерпел эффект лилипутов и перестройку замка. Различия по тем же параметрам между G. aspera и G. bathonica (хронологические виды) оказались гораздо меньшими.
Северогерманская зона распространения вида G. tuberodentina, характеризующая верхний байос (зона Parkinsoni, верхняя часть подзоны Truellei и подзона Bomfordi), впервые прослежена в Польше и на Украине. На Украине объем зоны G. tuberodentina сокращен за счет выделения в зоне Michalskii новой остракодовой филозоны.
По эволюции восточноевропейской ветви изученных глиптоцитер, начавшейся с миграции G. tuberodentina в акваторию ДДВ в позднем байосе (в зоне Parkinsoni), для территории РП предлагаются новые филозоны. Филозона G. tuberodentina характеризует верхний байос, зону Parkinsoni в ДДВ; зона G. aspera (верхний байос – нижний бат, зона Michalskii–нижняя часть зоны Besnosovi с референсными биогоризонтами O. mojarowskii и O. sciticum) прослежена на Украине, в Центральной России, Нижнем Поволжье и Зап. Казахстане, а зона G. bathonica (нижний бат, верхняя часть зоны Besnosovi с биогоризонтами O. issae–A. excentricum) установлена в Центральной России, Нижнем Поволжье и Зап. Казахстане.
* * *
Автор весьма признательна Н.Н. Колпенской (Геологоразведка, СПб.), Я.А. Шуруповой (МГУ), Л.А. Глинских (ИНГГ СО РАН), А.П. Ипполитову (ГИН РАН), А.С. Алексееву (МГУ), А.В. Черешинскому (ВГУ), Р.А. Воиновой (МГУ), В.Б. Сельцеру (СНИГУ) и А.В. Иванову (ИГ РАН) за разнообразную помощь в сборе и обработке материала, легшего в основу этой статьи (Тесакова, 2022а, б). Отдельная благодарность моим рецензентам Л.М. Мельниковой (ПИН РАН) и А.П. Ипполитову (ГИН РАН) за замечания к тексту статьи, позволившие его улучшить.
Работа выполнена в рамках тем госзадания №№ 0135-2019-0062 (ГИН РАН) и АААА-А16-116033010096-8 (МГУ).
Список литературы
Басов В.А., Никитенко Б.Л., Куприянова Н.В. Стратиграфия и микрофауна (фораминиферы и остракоды) нижней и средней юры баренцевоморского шельфа // Геол. и геофизика. 2009. Т. 50. № 5. С. 525–549.
Гуляев Д.Б. Аммониты и инфразональная стратиграфия зоны Besnosovi нижнего бата Русской плиты // Стратигр. Геол. корреляция. 2019. Т. 27. № 1. С. 103–125.
Гуляев Д.Б., Ипполитов А.П. Новые данные о морском нижнем бате Центральной России // Юрская система России: проблемы стратиграфии и палеогеографии. Седьмое Всеросс. совещ.: научн. материалы. М.: ГИН РАН, 2017. С. 42–46.
Колпенская Н.Н. Комплексы остракод средней – верхней юры Белгородской области (юго-западный склон Воронежской антеклизы) // Тр. НИИГеологии СГУ. Нов. сер. 2002. Т. 11. С. 116–122.
Масумов А.С. Юрские остракоды Узбекистана. Ташкент: Фан, 1973. 197 с.
Международный кодекс зоологической номенклатуры. Изд. 4. СПб.: Зоол. ин-т РАН, 2000. 221 с.
Никитенко Б.Л. Стратиграфия, палеобиогеография и биофации юры Сибири по микрофауне (фораминиферы и остракоды). Новосибирск: Параллель, 2009. 680 с.
Пермякова М.Н. Остракоды рода Glyptocythere из среднеюрских отложений Днепрово-Донецкой впадины // Палеонтол. сб. Львов. ун-та. 1970. Вып. 1. № 7. С. 61–67.
Пермякова М.Н. Юрские остракоды платформенной Украины // Обоснование стратиграфических подразделений мезо-кайнозоя Украины по микрофауне. Киев: Наук. думка, 1975. С. 30–47.
Практическое руководство по микрофауне СССР. Т. 3. Остракоды кайнозоя / Ред. Б.С. Соколов. Л.: Недра, 1989. 233 с.
Практическое руководство по микрофауне. Т. 7. Остракоды мезозоя / Ред. Б.С. Соколов. СПб.: ВСЕГЕИ, 1999. 244 с.
Преображенская В.Н. Значение остракод для стратиграфии юрских отложений территории КМА // Геология и полезные ископаемые Центрально-Черноземных областей. Тр. межобл. геол. совещ. Воронеж: Изд-во Воронеж. ун-та, 1964. С. 179–183.
Преображенская В.Н. Стратиграфия отложений юры и низов нижнего мела территории ЦЧО. Воронеж: Изд‑во Воронеж. ун-та, 1966. 281 с.
Пяткова Д.М., Пермякова М.Н. Фораминиферы и остракоды юры Украины. Киев: Наук. думка, 1978. 288 с.
Салтыков В.Ф., Старцева Г.Н., Троицкая Е.А. Стратиграфия байос–батских морских отложений Нижнего Поволжья // Изв. Саратов. ун-та. 2008. Т. 8. Сер. Науки о Земле. Вып. 2. С. 56–71.
Тесакова Е.М. Позднебайосские и раннебатские остракоды Русской плиты. Часть I. Роды Plumhoffia Brand и Aaleniella Plumhoff // Палеонтол. журн. 2022а. № 2. С. 57–68. https://doi.org/10.31857/S0031031X2202012X
Тесакова Е.М. Позднебайосские и раннебатские остракоды Русской плиты. Часть II. Роды Procytherura Whatley, Pseudohutsonia Wienholz, Acrocythere Neale, Nanacythere Herrig и Trachycythere Triebel et Klingler // Палеонтол. журн. 2022б. № 3. С. 19–30. https://doi.org/10.31857/S0031031X22030151
Тесакова Е.М. Позднебайосские и раннебатские остракоды Русской Плиты. Часть III. Роды Camptocythere Triebel и Procytheridea Peterson // Палеонтол. журн. 2022в. № 4. С. 48–58.
Унифицированная региональная стратиграфическая схема юрских отложений Восточно-Европейской платформы (14 листов). Объяснительная записка. М.: ПИН РАН–ФГУП “ВНИГНИ”, 2012. 64 с.
Хабарова Т.Н. Остракоды из отложений средней юры Саратовской области и северных районов Сталинградской области // Тр. ВНИГРИ. Нов. сер. 1955. Вып. 84. С. 192–197.
Хабарова Т.Н. О микрофауне юрских отложений Саратовской области // Тр. ВНИГРИ. 1961. Вып. 29. Т. 3. С. 177–184.
Шорников Е.И., Михайлова Е.Д. Остракоды Bythocytheridae раннего этапа развития: Сравнительная морфология, палеоэкология, пути эволюции. М.: Наука, 1990. 200 с.
Шурупова Я.А., Тесакова Е.М., Колпенская Н.Н. и др. Саратовское Поволжье в позднем байосе (средняя юра): палеогеография, реконструированная по остракодам // Жизнь Земли. 2016. Т. 38. № 1. С. 22–37.
Bate R.H. Middle Jurassic Ostracoda from the Grey Limestone Series, Yorkshire // Bull. Brit. Mus. (Natur. Hist.) Geol. 1965. V. 11. № 3. P. 73–133.
Bate R.H. The Bathonian Upper Estuarine Series of Eastern England. Part I: Ostracoda // Bull. Brit. Mus. (Natur. Hist.) Geol. 1967. V. 14. № 2. P. 21–66.
Bate R.H. Some Bathonian ostracoda of England with a revision of the Jones, 1884 and Jones et Sherborn, 1888 collections // Bull. Brit. Mus. (Natur. Hist.) Geol. 1969. V. 17. № 8. P. 379–437.
Bate R.H. Middle Jurassic (Aalenian – Bathonian) // Ostracods in British Stratigraphy / Eds. J.E. Whittaker, M.B. Hart. L.: The Micropaleontol. Soc., Spec. Publ. Geol. Soc, 2009. P. 199–223.
Bielecka W., Styk O. Assemblage of foraminifers and ostracods in the deposits of clay-siltstone-arenaceous facies of Kujavian and Bathonian age within the Polish Lowlands // Kwart. Geol. 1969. V. 13. № 3. P. 619–628.
Bielecka W., Styk O., Błaszyk J., Kopik J. Middle Jurassic. Ostracoda Latreille, 1806 // Geology of Poland. Atlas of guide and characteristic fossils. Part 2b. Mesozoic Jurassic / Ed. L. Malinowska. Warsaw, 1988. P. 165–179.
Błaszyk J. Middle Jurassic ostracods of the Czestochowa region (Poland) // Acta Palaeontol. Pol. 1967. V. 12. № 1. P. 1–75.
Brand E. Biostratigraphische Untergliederung des Ober-Bathonium im Raum Hildesheim, Nordwestdeutschland mittels Ostracoden und Korrelation ihrer Vertikalreichweiten mit Ammoniten-Zonen // Jb. Geol. B.-A. 1990. Bd 121. S. 119–273.
Brand E., Fahrion H. Dogger NW-Deutschlands // Arbeitskreis Deutscher Mikropaläontologen. Leitfossilien der Mikropaläontologie. Berlin: Borntraeger, 1962. S. 123–158.
Brand E., Malz H. Ostracoden-Studium im Dogger, 5: Glyptocythere n. g. // Senckenb. leth. 1962. V. 43. № 5. S. 433–435.
Brand E., Malz H. Die Arten der Gattung Glyptocythere Brand & Malz 1962 im NW-deutschen Dogger // Senckenb. leth. 1966. V. 47. № 5/6. S. 481–535.
Brand E., Mönnig E. Litho- und Biostratigraphie des Mittel-Jura (Dogger) in Bohrungen Norddeutschlands // Schr. Dtsch. Ges. Geowiss. 2009. Bd 54. S. 5–73.
Dépêche F. Les ostracodes d’une plate-forme continentale au Jurassique: recherches sur le bathonien du Bassin Parisien // Mém. Sci. Terre. Univ. Pierre-et-Marie-Curie, Paris. N 84 38. P., 1984. 419 p. [unpubl. thesis].
Dépêche F. Lias supérieur, Dogger, Malm // Atlas des ostracodes de France. Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine. 1985. Mém. 9. P. 119–145.
Dietze V., Franz M., Kutz M., Waltschew A. Stratigraphy of the Middle Jurassic Sengenthal Formation of Polsingen-Ursheim (Nördlinger Ries, Bavaria, Southern Germany) // Palaeodiversity. 2017. V. 10. P. 49–95.
Dietze V., Wannenmacher N., Franz M., Weis R. Neue Erkenntnisse über die Wedelsandstein-Formation der Zollernalb (Schwäbische Alb, SW-Deutschland) // Zitteliana. 2018. V. 93. P. 47–80.
Franz M., Beher E., Dietl G. The Bathonian and Early Callovian Ostracoda of Baden-Wuerttemberg, southern Germany // N. Jb. Geol. Paläontol. Abh. 2014. V. 274. № 2–3. P. 149–185.
Franz M., Tesakova E.M., Beher E. Documentation and revision of the index ostracods from the Lower and Middle Jurassic in SW Germany according to Buck (1954) // Palaeodiversity. 2009. V. 2. P. 119–167.
Luppold F.W. Ostracod assemblages from the Middle Jurassic of NW Germany with special reference to the Sowerbyi ammonite Zone (Early Bajocian, Jurassic) // N. Jb. Geol. Paläontol. Abh. 2012. V. 266. № 3. P. 217–238.
Lutze G.F. Glyptocythere obtusa n. sp. (Ostrac., Dogger) // Senckenb. leth. 1966. V. 47. № 5/6. P. 536.
Mitta V., Kostyleva V., Dzyuba O. et al. Biostratigraphy and sedimentary settings of the Upper Bajocian-Lower Bathonian in the vicinity of Saratov (Central Russia) // N. Jb. Geol. Paläontol. Abh. 2014. V. 271. № 1. P. 95–121.
Sheppard L.M. Middle Jurassic Ostracoda from Southern England and Northern France. Thesis Ph.D. Univ. of London. 1981. 214 p.
Shurupova Y.A., Tesakova E.M. Detailed biostratigraphic scales as based on the palaeobiogenetical approach (an example of the Upper Bajocian-Lower Bathonian ostracod scale of the Russian Platform) // Volumina Jurassica. 2017. V. 15. P. 1–17.
Sylvester-Bradley P.C. Bathonian ostracods from the Boueti Bed of Langton Herring, Dorset // Geol. Mag. 1948. V. 85. P. 185–204.
Tesakova E.M. Biostratigraphie du Jurassique moyen des sondages Benken et Schlattingen-1 (Nord de la Suisse) ètablie sur la base de la répartition des ostracodes // NAGRA Arbeitsbericht NAB. 2017. V. 17–41. 77 p.
Tesakova E.M., Franz M., Baykina E., Beher E. A new view on Bathonian ostracods of Poland // Senckenb. leth. 2008. V. 88. № 1. P. 55–65.
Wakefield M. Ostracoda and palaeosalinity fluctuations in the Middle Jurassic Lealt Shale Formation, Inner Hebrides, Scotland // Palaeontology. 1995. V. 38. P. 583–617.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал