

Палеонтологический журнал, 2022, № 5, стр. 89-94
Новый вид рода Pseudauletes Voss (Coleoptera: Rhynchitidae) из балтийского янтаря
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
b Алтайский государственный университет
656049 Барнаул, Россия
c Томский государственный университет
Томск, 634050Россия
* E-mail: fossilweevils@gmail.com
Поступила в редакцию 27.01.2022
После доработки 04.04.2022
Принята к публикации 04.04.2022
- EDN: WBHSFA
- DOI: 10.31857/S0031031X22050075
Аннотация
В статье описан новый вид жуков-ринхитид Pseudauletes (Eopseudauletes) balticus sp. nov. из балтийского янтаря. Новый вид отличается от P. (E.) championi (Sharp, 1889) редко пунктированной переднеспинкой, чья длина больше ширины, и усиками, прикрепленными ближе к основанию головотрубки. Обсуждаются возможные связи между фауной Curculionoidea балтийского янтаря и американской фауной Limalophus и Toxorhynchus в эоцене, а также современными Pseudauletes, Caulophilus и Oxycraspedus на родовом уровне и Anypotactini на уровне трибы.
ВВЕДЕНИЕ
Представители трибы Auletini – самые базальные в надтрибе Rhynchititae подсемейства Rhynchitinae (Legalov, 2015a). К этой надтрибе относятся шесть триб (Auletini, Cesauletini, Eugnamptini, Deporaini, Rhynchitini и Byctiscini). Auletini отличаются от остальных триб совместно закругленными надкрыльями, как у Nemonychidae, самого древнего семейства долгоносикообразных жуков, известного с конца средней юры (Легалов, 2022). Хотя семейство Rhynchitidae появляется в палеонтологической летописи в сеномане (Legalov, 2015b), первые Auletini обнаружены в раннем–среднем эоцене Грин-Ривер (Legalov, 2020b). Остальные палеогеновые находки сделаны в позднеэоценовых балтийском и ровенском янтарях (Legalov, 2015b; Bukejs, Legalov, 2019, 2021), а также в терминальном эоцене Флориссанта (Scudder, 1893; Legalov, 2015b, 2020b). Неогеновые Auletini описаны из раннемиоценового доминиканского янтаря (Poinar, Legalov, 2015b).
Подтриба Pseudauletina включает в себя два рода: Eoropseudauletes Kania et Legalov, 2019 из балтийского янтаря (Kania, Legalov, 2019) и современный Pseudauletes Voss, 1922, включающий 14 видов (Legalov, 2007, 2018). В ископаемом состоянии виды этого рода не были известны.
В представленной работе описывается новый вид рода Pseudauletes из эоценового балтийского янтаря, который является первой находкой рода в палеогене.
МАТЕРИАЛ И МЕТОДЫ
Исследованный образец происходит из балтийского янтаря, добытого в окрестностях пос. Янтарный, расположенного на побережье Балтийского моря в Калининградской обл. (Россия). Продуцентом балтийского янтаря, вероятно, был один из видов рода Sciadopitys семейства Sciadopityaceae (Sadowski et al., 2016). Возраст балтийского янтаря из этой прусской свиты датируют от среднего эоцена, бартонского яруса (Bukejs et al., 2019), до позднего эоцена, приабонского яруса (Perkovsky et al., 2007).
Типовой материал хранится в коллекции Ин‑та систематики и экологии животных СО РАН, Новосибирск.
Описание, промеры и фотографии были сделаны при помощи бинокулярного микроскопа Zeiss Stemi 2000-C. Для обработки иллюстраций использовался Adobe Photoshop версии 21.1.3.
Автор благодарит М.В.Л. Баркли (M.V.L. Barclay) (Лондон), Л. Бэнэ (L. Behne) (Мюнхеберг), К.-Д. Класса (K.-D. Klass) (Дрезден), О. Йегера (O. Jaeger) (Дрезден), Р. Погги (R. Poggi) (Генуя), Й. Виллерс (J. Willers) (Берлин), и Д. Тельнова (Лондон) за возможность изучения материалов по роду Pseudauletes.
ТАКСОНОМИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО RHYNCHITIDAE GISTEL, 1848
ПОДСЕМЕЙСТВО RHYNCHITINAE GISTEL, 1848
Надтриба Rhynchititae Gistel, 1848
Триба Auletini Desbrochers des Loges, 1908
Подтриба Pseudauletina Voss, 1933
Род Pseudauletes Voss, 1922
Подрод Eopseudauletes Legalov, 2007
Pseudauletes (Eopseudauletes) balticus Legalov, sp. nov.
Название вида – от Балтики.
Голотип – BA2019/10 (жук в янтаре хорошо виден, снизу покрыт белым налетом; возле верхней левой части переднеспинки янтарь поврежден); балтийский янтарь, поздний эоцен.
Описание (рис. 1). Тело черное, блестящее, покрытое длинными торчащими темными щетинками. Головотрубка длинная, слабоизогнутая, явственно расширяющаяся на вершине, уплощенная сверху, мелко пунктированная. Длина головотрубки в 1.7 раза больше длины переднеспинки, в 4.1 раза больше ширины на вершине, в 7.3 раза больше ширины на середине и в 5.0 раз больше ширины на основании. Мандибулы с зубцом на внешнем крае. Глаза округленные, явственно выступают из контура головы, мелкофасеточные. Лоб довольно широкий, слабовыпуклый, мелко пунктированный. Ширина лба немного больше ширины головотрубки на основании. Длина висков примерно в 1.4 раза больше длины глаза. Темя слабовыпуклое, пунктированное. Усики не коленчатые, прикреплены перед серединой головотрубки. Первый–восьмой членики усиков удлиненно-конические. Длина первого членика в 1.8 раза больше его ширины на вершине. Длина второго членика примерно в 2.8 раза больше его ширины на вершине и равна длине первого членика. Ширина второго членика составляет примерно 0.6 ширины первого членика. Длина третьего членика в 5.2 раза больше его ширины на вершине и примерно в 1.4 раза больше длины второго членика. Ширина третьего членика составляет примерно 0.8 ширины второго членика. Длина четвертого членика примерно в 4.0 раза больше его ширины на вершине и составляет примерно 0.8 длины третьего членика. Ширина четвертого членика равна ширине третьего членика. Пятый–седьмой членики примерно равной ширины. Длина пятого членика примерно в 3.2 раза больше его ширины на вершине и немного меньше длины четвертого членика. Ширина пятого членика в 1.2 раза больше ширины четвертого членика. Длина шестого членика примерно в 2.2 раза больше его ширины на вершине и составляет примерно 0.7 длины пятого членика. Длина седьмого членика вдвое больше его ширины на вершине и немного меньше длины шестого членика. Длина восьмого членика примерно в 1.4 раза больше его ширины на вершине и составляет примерно 0.8 длины седьмого членика. Ширина восьмого членика примерно в 1.2 раза больше ширины седьмого членика. Булава крупная, не компактная. Длина девятого членика примерно в 1.5 раза больше его ширины на вершине и в два раза больше длины восьмого членика. Ширина девятого членика в 1.9 раза больше ширины восьмого членика. Длина десятого членика в 1.5 раза больше его ширины на вершине и составляет 0.9 длины девятого членика. Ширина десятого членика немного меньше ширины девятого членика. Длина 11-го членика примерно в 2.2 раза больше его ширины на вершине и примерно в 1.4 раза больше длины десятого членика. Ширина 11-го членика равна ширине десятого членика. Гулярный шов одинарный. Переднеспинка колоколовидная, редко и мелко пунктированная. Длина переднеспинки примерно вдвое больше ширины на вершине, немного больше ширины на середине и в 1.4 раза больше ширины на основании. Диск переднеспинки слабовыпуклый. Бока слабо округленные. Щиток явственный, примерно равной длины и ширины, пятиугольный. Надкрылья явственно выпуклые, почти прямоугольные, с наибольшей шириной за серединой. Плечи выпуклые. Длина надкрылий в два раза больше длины переднеспинки, примерно в 1.2 раза больше ширины на основании, в 1.2 раза больше ширины на середине и в 1.6 раза больше ширины в вершинной четверти. Надкрылья в спутанной пунктировке из редких и крупных точек. Промежутки между точками широкие, уплощенные, гладкие. Вершины надкрылий закруглены совместно. Эпиплевры явственные. Переднегрудь без заглазничных лопастей, пунктированная. Пре- и посткоксальные части переднегруди очень короткие. Тазиковые впадины переднегруди соединенные, расположены на середине переднегруди. Заднегрудь слабовыпуклая, пунктированная; ее длина равна длине заднего тазика. Эпистерны заднегруди узкие, в 4.0 раза больше ширины на середине. Брюшко выпуклое, пунктированное. Первый и второй вентриты удлиненные и сросшиеся, равной длины. Длина первого вентрита примерно равна максимальной длине заднего тазика. Длина третьего вентрита составляет 0.8 длины второго вентрита. Длина четвертого вентрита составляет 0.75 длины третьего вентрита. Длина пятого вентрита примерно в 1.6 раза больше длины четвертого вентрита. Ноги длинные. Передние тазики конические. Бедра булавовидные, без зубцов. Голени узкие и длинные, расширенные к вершине, с мелкозазубренным килем на наружном крае, без ункуса и мукро. Лапки длинные, немного короче половины длины голени. Первый и второй членики лапок удлиненно-трапециевидные. Третий членик лапок двухлопастный. Коготки не сросшиеся, слабо расставленные, с зубцами.
Рис. 1.
Pseudauletes (Eopseudauletes) balticus sp. nov., голотип BA2019/10, общий вид: а – дорсально; б – вентрально; в – латерально; балтийский янтарь, поздний эоцен. Длина масштабного отрезка 1.0 мм.
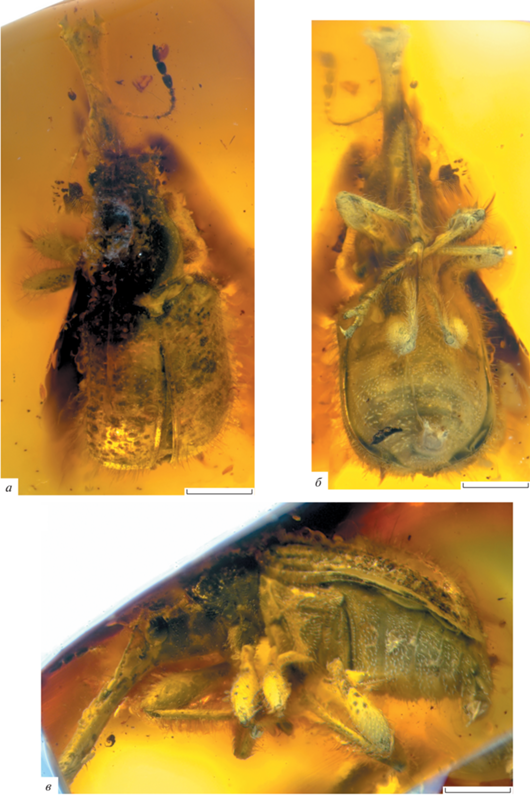
Размеры в мм: длина тела без головотрубки – 4.7; длина головотрубки – 2.5.
Сравнение. Новый вид широкими надкрыльями сходен с центральноамериканским P. (Eopseudauletes) championi (Sharp, 1889), но отличается редко пунктированной переднеспинкой, чья длина больше ширины, и усиками, прикрепленными ближе к основанию головотрубки.
Замечания. Новый вид отнесен к семейству Rhynchitidae, поскольку у него почти гомономные вентриты брюшка, не сросшиеся коготки лапок, одинарный гулярный шов, и голени без ункуса. Слабо расставленные коготки лапок являются признаком подсемейства Rhynchitinae. Новый вид помещен в надтрибу Rhynchititae, поскольку обладает мандибулами с зубцом на внешнем крае. Совместно закругленные вершины надкрылий указывают на принадлежность к трибе Auletini. Размещение нового вида в подтрибе Pseudauletina подтверждается голенями с килем на наружном крае и коготками с зубцами. Новый вид относится к роду Pseudauletes, поскольку характеризуется телом, покрытым густыми торчащими щетинками, а также длинной, уплощенной сверху головотрубкой. Темное брюшко и усики, прикрепленные перед серединой головотрубки, указывают на принадлежность к подроду Eopseudauletes Legalov, 2007.
Материал. Голотип.
ЗАКЛЮЧЕНИЕ
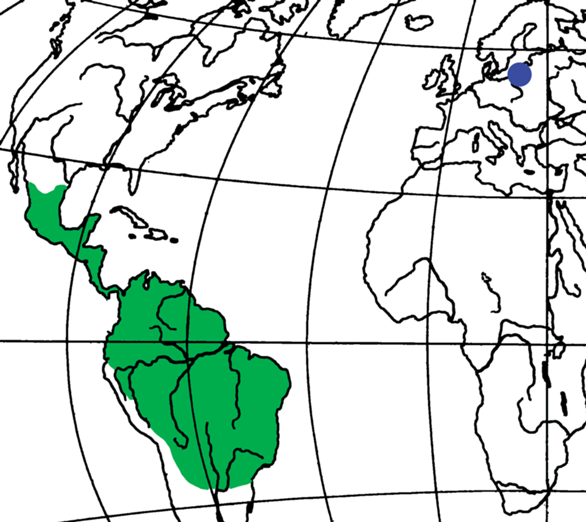
Находка нового вида из современного неотропического рода позволяет обсудить связи фауны долгоносикообразных жуков балтийского янтаря и Нового Света. Виды рода Pseudauletes распространены от Мексики до Аргентины (рис. 2). Они не заходят на севере на территорию США и в Чилийско-Патагонскую область на юге. Сходное современное распространение имеет род Caulophilus Wollaston, 1854, представленный тремя видами в балтийском янтаре (Legalov, 2020a) и двумя в ровенском (Legalov et al., 2019). Род Toxorhynchus Scudder, 1893 [один вид в балтийском и один вид в ровенском янтарях (Bukejs, Legalov, 2020; Легалов, 2021)] имеет более широкий ареал, охватывающий небореальную Северную Америку и Южную Америку. Представители этого рода описаны из позднего эоцена США и раннего миоцена Эспаньолы (Poinar, Legalov, 2015a). Связи с Чилийско-Патагонской областью демонстрирует род Oxycraspedus Kuschel, 1955, вымерший вид которого был обнаружен в балтийском янтаре (Legalov, 2016). Самые обычные долгоносики в балтийском янтаре – виды рода Paonaupactus Voss, 1953, который относится к преимущественно неотропической (на север до Техаса) трибе Anypotactini. Наряду с современными фаунистическими связями, Limalophus poinari Legalov, 2020 из балтийского янтаря (Legalov, 2020a), относящийся к вымершему роду Limalophus Scudder, 1893 из среднего–позднего эоцена США, подтверждает сходство эоценовых фаун Америки и Европы. Вероятно, связь между фаунами Северной Америки и Европы в эоцене осуществлялась через сухопутный “мост”, включавший Гренландию и располагавшийся в поясе теплых температур (Boucot et al., 2013), что позволяло долгоносикообразным жукам перемешаться с одного континента на другой.
Список литературы
Легалов А.А. Новые жуки-долгоносики семейства Brentidae (Coleoptera) из балтийского янтаря // Палеонтол. журн. 2021. № 1. С. 99–103.
Легалов А.А. Обзор юрских долгоносиков рода Belonotaris Arnoldi (Coleoptera: Nemonychidae) с прямой головотрубкой // Палеонтол. журн. 2022. № 2. С. 79–88.
Boucot A.J., Chen X., Scotese C.R. Phanerozoic paleoclimate: an atlas of lithologic indicators of climate. SEPM, 2013. 478 p. (Concepts in Sedimentol. and Paleontol. № 11).
Bukejs A., Alekseev V.I., Pollock D.A. Waidelotinae, a new subfamily of Pyrochroidae (Coleoptera: Tenebrionoidea) from Baltic amber of the Sambian peninsula and the interpretation of Sambian amber stratigraphy, age and location // Zootaxa. 2019. V. 4664. № 2. P. 261–273.
Bukejs A., Legalov A.A. The first record of Rhynchitidae (Coleoptera) from Rovno amber // Entomol. Fennica. 2019. V. 30. № 4. P. 168–172.
Bukejs A., Legalov A.A. The first record of Brentidae (Coleoptera) in Eocene Rovno amber with description of a new fossil species of Toxorhynchus Scudder, 1893 // Fossil Rec. 2020. V. 23. P. 169–177.
Bukejs A., Legalov A.A. Two new species of the family Rhynchitidae (Coleoptera: Curculionoidea) from Eocene Baltic amber, with key to species and assumed trophic relationships // Fossil Rec. 2021. V. 24. P. 117–127.
Kania J., Legalov A.A. A new genus of tooth-nosed snout weevils (Coleoptera: Rhynchitidae) in Baltic amber // Paleontol. J. 2019. V. 53. № 10. P. 1040–1044.
Legalov A.A. Leaf-rolling Weevils (Coleoptera: Rhynchitidae, Attelabidae) of the World Fauna. Novosibirsk: Agro-Siberia, 2007. 523 p.
Legalov A.A. The family Rhynchitidae (Insecta: Coleoptera) in the Himalayas // Biodiv. und Naturausst. im Himalaya. 2015a. Bd 5. P. 479–486.
Legalov A.A. Fossil Mesozoic and Cenozoic weevils (Coleoptera, Obrienioidea, Curculionoidea) // Paleontol. J. 2015b. V. 49. № 13. P. 1442–1513.
Legalov A.A. Two new genera and four new species of fossil weevils (Coleoptera: Curculionoidea) in Baltic amber // Entomol. Fennica. 2016. V. 27. № 2. P. 57–69.
Legalov A.A. Annotated key to weevils of the world. Part 1. Families Nemonychidae, Anthribidae, Belidae, Ithyceridae, Rhynchitidae, Brachyceridae and Brentidae // Ukr. J. Ecol. 2018. V. 8. № 1. P. 780–831.
Legalov A.A. A review of the Curculionoidea (Coleoptera) from European Eocene ambers // Geosciences. 2020a. V. 10. № 1(16). P. 1–74.
Legalov A.A. Fossil history of Curculionoidea (Coleoptera) from the Paleogene // Geosciences. 2020b. V. 10(9). № 358. P. 1–50.
Legalov A.A., Nazarenko V.Yu., Perkovsky E.E. New weevils (Coleoptera: Curculionidae) from Rovno amber // Paleontol. J. 2019. V. 53. № 10. P. 1045–1059.
Perkovsky E.E., Rasnitsyn A.P., Vlaskin A.P., Taraschuk M.V. A comparative analysis of the Baltic and Rovno amber arthropod faunas: representative samples // Afr. Invertebr. 2007. V. 48. P. 229–245.
Poinar G.Jr., Legalov A.A. New Apioninae (Coleoptera: Brentidae) in Dominican amber // Hist. Biol. 2015a. V. 27. № 2. P. 134–157.
Poinar G.Jr., Legalov A.A. Two new species of the genus Rhynchitobius Sharp, 1889 (Coleoptera: Rhynchitidae) in Dominican amber // Ann. Soc. Entomol. Fr. (N.S.). 2015b. V. 51. № 1. P. 70–77.
Sadowski E.-M., Schmidt A.R., Kunzmann L. et al. Sciadopitys cladodes from Eocene Baltic amber // Bot. J. Linn. Soc. 2016. V. 180. № 2. P. 258–268.
Scudder S.H. Tertiary rhynchophorus Coleoptera of the United States // Monogr. US Geol. Surv. Terr. 1893. V. 21. P. 1–206.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал