

Палеонтологический журнал, 2022, № 5, стр. 79-88
Обзор блаттинопсидовых насекомых (Insecta: Blattinopsida) и флоры конца ранней–начала средней перми европейской части России
Д. С. Аристов a, b, *, А. П. Расницын a, c, **, С. В. Наугольных d, ***
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
b Череповецкий государственный университет
162602 Череповец, Россия
c Музей естественной истории
SW7 5BD Лондон, Великобритания
d Геологический институт РАН
119017 Москва, Россия
* E-mail: danil_aristov@mail.ru
** E-mail: alex.rasnitsyn@gmail.com
*** E-mail: naugolnykh@list.ru
Поступила в редакцию 28.12.2021
После доработки 07.02.2022
Принята к публикации 15.02.2022
- EDN: PUZYZY
- DOI: 10.31857/S0031031X22050038
Аннотация
Рассмотрены состав блаттинопсидовых насекомых (Insecta: Blattinopsida) и флора конца ранней–начала средней перми России. Описаны Glaphyrophlebia semipenna sp. nov. и G. glinka sp. nov. из нижнепермских местонахождений Чекарда и Красная Глинка в Пермском крае России. Высказано предположение о том, что резкий скачок доли блаттинопсид в местонахождениях начала уфимского века, возможно, был связан с развитием специфических ландшафтных обстановок, в которых доминировали мохообразные и древовидные гетероспоровые плауновидные: предполагается, что блаттинопсидовые питались в основном спорами этих плауновидных.
Древнейшие блаттинопсидовые насекомые с территории России известны из нижнепермских отложений. Они найдены в кунгурских местонахождениях Чекарда и Красная Глинка в Пермском крае. Из Чекарды в Суксунском р-не (кошелевская свита) описаны Glaphyrophlebia uralensis (Martynov, 1940) (Мартынов, 1940; Расницын, 1980) и G. semipenna sp. nov. Из близковозрастного местонахождения Красная Глинка (Кишертский р-н) ниже описывается G. glinka sp. nov. Не определенный до вида представитель Glaphyrophlebia найден в кунгурских (лекворкутская свита) отложениях Печорского бассейна (Аристов и др., 2021). Из местонахождения Фокина (верхнебургуклинский подгоризонт верхнего кунгура Красноярского края России) описан Blattinopsis indefinitus Rasnitsyn et Aristov, 2021 (Rasnitsyn, Aristov, 2021). Это самая поздняя находка рода Blattinopsis Giebel, 1867 и единственный представитель рода из перми России, остальные Blattinopsida представлены родом Glaphyrophlebia.
Из более молодых, уфимских отложений Воркутского месторождения (интинская свита Печорского бассейна) описаны Glaphyrophlebia komia Aristov, Rasnitsyn et Naugolnykh, 2021, G. vorkutensis Aristov, Rasnitsyn et Naugolnykh, 2021 и G. borea Aristov, Rasnitsyn et Naugolnykh, 2021 (Аристов и др., 2021). Из близковозрастного местонахождения Тюлькино в Пермском крае (соликамская свита) описаны G. vaga Aristov et Rasnitsyn, 2022, G. rara Aristov et Rasnitsyn, 2022 и G. kamaensis Aristov et Rasnitsyn, 2022 (Аристов, Расницын, 2022).
Наиболее молодые представители отряда известны из нижнеказанских местонахождений Сояна в Архангельской обл. России и Тихие Горы в Татарстане (Россия). Из Сояны описаны G. subcostalis (Martynov, 1928) и G. iva Aristov et Rasnitsyn, 2021, из Тихих Гор – G. rossica (Martynov, 1930) (Аристов, Расницын, 2021). Из более молодых отложений блаттинопсидовые не известны.
Разнообразие блаттинопсидовых в пермских отложениях России невелико и составляет один–три вида в одном местонахождении вне зависимости от доли блаттинопсидовых среди других насекомых. Доля эта может составлять менее одного процента в кунгурских (Чекарда и отложения лекворкутской свиты) и нижнеказанских (Сояна и Тихие Горы) местонахождениях. В уфимских местонахождениях доля Blattinopsidae выше и составляет 14% в отложениях интинской свиты Печорского бассейна России (Расницын и др., 2005) и 22.5% в местонахождении Тюлькино в Пермском крае (соликамская свита: Аристов, Расницын, 2022). В нижней перми сходная доля блаттинопсидовых известна только в ассельском местонахождении Нидермошель (формация Майзенхайм, Рейнланд-Пфальц, Германия), где она составляет 11% (Hörnschemeyer, 1999). Вряд ли эта высокая (для блаттинопсидовых) доля объясняется климатическими причинами. Для Нидермошеля был характерен аридный континентальный климат, для Тюлькино – семиаридный прибрежноморской, для интинских местонахождений Печорского бассейна – гумидный прибрежноморской (Аристов и др., 2021).
БЛАТТИНОПСИДЫ И РАННЕУФИМСКИЙ ЭКОСИСТЕМНЫЙ КРИЗИС
Анализ доли блаттинопсид среди других насекомых в местонахождениях Чекарда, лекворкутской и интинской свит Печорского бассейна, Тюлькино и Сояна, совершенно отчетливо указывает на увеличение доли блаттинопсид в первой половине уфимского века, т.е. в соликамское время. Общая оценка изменения палеогеографической ситуации на востоке Русской (Восточно-Европейской) платформы и в Приуралье позволяет сделать вывод о том, что увеличение доли блаттинопсид в раннеуфимских местонахождениях интинской свиты Воркутского месторождения и Тюлькино могло соответствовать появлению в этом временном интервале специфических ландшафтных условий, благоприятствовавших развитию и распространению этой группы насекомых.
Как показывают исследования пермских отложений этого региона, именно в соликамское время произошло закрытие Приуральского морского бассейна, к кунгурскому веку превратившегося в гиперсоленую лагуну, в которой откладывались эвапориты – калийные соли и галит на севере в районе Соликамска и Березников (Пермский край), а также сульфаты (начиная с широты г. Пермь и далее на юг до Прикаспийской впадины; подробнее см.: Деревягин и др., 1981; Жарков, 2004). Эта ситуация, безусловно, прямо сказалась на палеофитогеографической и фитоценотической дифференциации растительности этого региона, а также на изменении типа и состава растительных сообществ от кунгурского века (местонахождение Чекарда) к казанскому (местонахождение Сояна; современные представления о трендах в эволюции растительности Приуралья и ее связях с другими палеофлористическими областями подробнее см. в: Naugolnykh, 2014, 2015, 2017, 2018; Tang et al., 2020).
В стратотипических разрезах и скважинах в Среднем Приуралье мощность соликамского горизонта составляет 100–300 м (Силантьев, 1996; Silantiev, 1998). Соликамские отложения в основном представлены плитчатыми мергелями и известняками (так называемыми “соликамскими плитняками”), иногда – с прослоями столбчатых и полусферических строматолитов (обзор см. в: Сонин, 2015; Наугольных, Литвинова, 2021) с подчиненными прослоями песчаников.
В соликамских отложениях Среднего Приуралья, включая разрез Тюлькино, широко распространены аридные и субаридные карбонатные палеопочвенные профили. Так, например, в отложениях этого возраста, обнажающихся в нижнем течении рек Чусовой и Сылвы, встречаются карбонатные палеопочвы с палеотакырами, разбитыми древними трещинами усыхания на характерные полигональные пластины (Наугольных, Литвинова, 2021). В профилях палеотакыров нередко присутствуют корни растений, сохранившиеся in situ, причем корни расположены либо рядом с трещинами, либо непосредственно в трещинах. Эта закономерность однозначно была вызвана тем, что именно в трещинах сохранялась влага, необходимая для растений в жарком и сухом климате соликамского времени.
Растительность первой половины уфимского века отличается очень низким таксономическим разнообразием, что, по всей видимости, было связано с региональным экологическим кризисом, имевшим место на рубеже кунгурского и уфимского веков (подробнее см.: Наугольных, 2007, с. 66–74).
Безусловным доминантом раннеуфимской растительности был древовидный гетероспоровый лепидофит Viatcheslavia vorcutensis Zalessky emend. Neuburg, остатки которого, в т.ч. фрагменты стволов, ризофоры, филлоиды, спорофиллы, мегаспоры и микроспоры, встречаются в Приуралье в большинстве местонахождений растительных остатков соликамского возраста. Известны находки стволов V. vorcutensis, достигающих 30 см в диаметре (подробнее о морфологии и формах роста V. vorcutensis см.: Наугольных, 2005). Высота самого растения при этом могла достигать 5 м. При этом важно помнить, что вторичные ткани у древовидных плауновидных были образованы не ксилемой, а корой, что совершенно не препятствует рассматривать форму роста этих растений как древовидную.
Вместе с вячеславиями часто встречаются листостебельные мхи Intia cf. variabilis Neuburg (рис. 1, а–д), а также хвощевидные Paracalamites aff. longisegmentatus Naug. В состав соликамского флористического комплекса также входят голосеменные: гинкгофиты Psygmophyllum sp., редкие хвойные, войновскиевые Rufloria derzavinii (Neub.) S. Meyen.
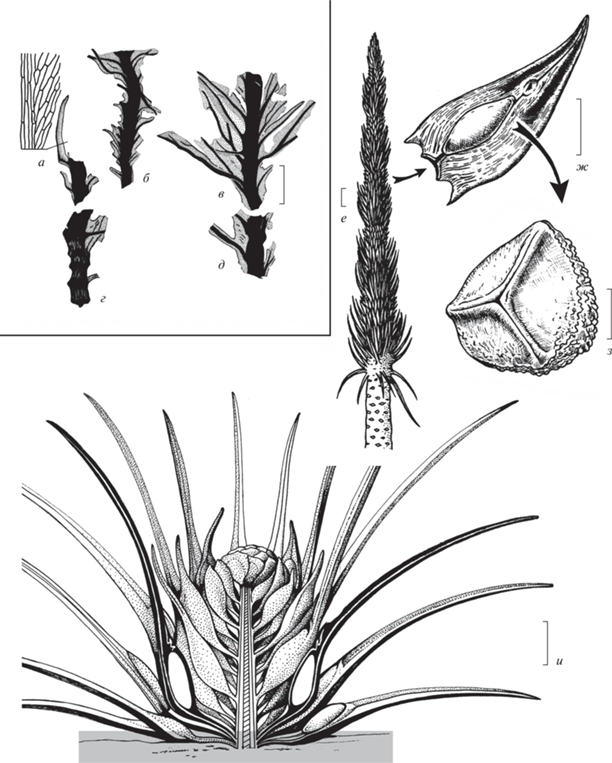
Рис. 1.
Основные типы ископаемых остатков мхов (а–д) и гетероспоровых плауновидных (е–и) из пермских отложений Пермского края, Россия: а–д – побеги листостебельного мха Intia cf. variabilis Neuburg (по: Наугольных, 2007, рис. 90 , с изм.), местонахождение Полазна, средняя пермь, уфимский ярус, соликамский горизонт; е – реконструкция апикальной части ствола гетероспорового древовидного лепидофита Viatcheslavia vorcutensis Zalessky; ж – спорофилл V. vorcutensis Zalessky; з – микроспора Densoisporites polaznaensis Naugolnykh et Zavialova, принадлежавшая V. vorcutensis Zalessky (по: Наугольных, 2016, рис. 8 , с изм.); местонахождение Полазна, средняя пермь, уфимский ярус, соликамский горизонт; и – Sadovnikovia belemnoides Naugolnykh, реконструкция травянистого гетероспорового плауновидного; нижняя пермь, кунгурский ярус, иренский горизонт. Длина масштабной линейки соответствует 2 мм (а–д); 10 см (е); 1 см (ж, и); 20 мкм (з).
Флористический комплекс интинской свиты существенно богаче синхронных ему флористических комплексов Среднего Приуралья, но и в нем присутствуют те же или близкие виды: Viatcheslavia vorcutensis, Paracalamites spp. и Rufloria derzavinii.
Таким образом, уфимские флористические комплексы различных районов Приуралья обладают в целом сходным таксономическим составом. Как уже было кратко отмечено выше, в качестве одной из наиболее ярких черт уфимских и, в особенности, раннеуфимских (соликамских) флор Приуралья можно назвать наличие древовидных лепидофитов Viatcheslavia vorcutensis (рис. 1, е–з).
Наиболее широко распространенной категорией ископаемых остатков V. vorcutensis являются фрагменты коры (табл. VI , фиг. 4, 7 ; табл. VII , фиг. 1–4 ; см. вклейку). Реже встречаются фрагменты стволов. Уникальное местонахождение крупных фрагментов стволов вячеславий, иногда сопровождающихся находками ризофоров, сохранившихся in situ, располагается в устьевой части р. Сылвы, напротив пос. Старые Ляды. Но, как правило, стволы вячеславий из этого местонахождения сильно декортицированы (табл. VI , фиг. 6 ). Гораздо реже встречаются спорофиллы вячеславий (табл. VI , фиг. 1, 8 ), строение которых удалось выяснить только через 60 лет после описания этого вида плауновидных (Наугольных, 1994, 2001). Путем аналитических исследований удалось установить, какие именно микроспоры продуцировались вячеславиями (Naugolnykh, Zavialova, 2004). Оказалось, что это каватные микроспоры с трилетным тетрадным рубцом (табл. VI , фиг. 2, 3, 5 ).
Рис. 2.
Реконструкция Glaphyrophlebia vaga Aristov et Rasnitsyn, на спорофилле Viatcheslavia vorcutensis Zalessky; Пермский край, местонахождение Тюлькино; средняя пермь, уфимский ярус, соликамский горизонт. Длина масштабной линейки 1 мм.
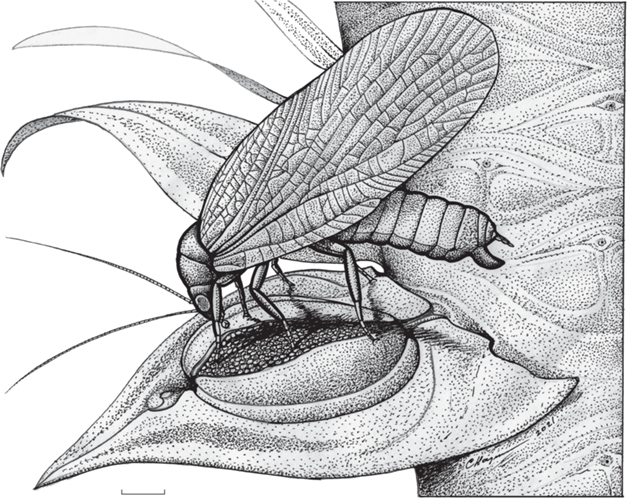
Рис. 3.
Новые представители рода Glaphyrophlebia из местонахождений Чекарда и Красная Глинка (нижняя пермь Пермского края России), передние крылья: а, б – G. semipenna sp. nov., голотип ПИН, № 1700/522: а – общий вид, б – прорисовка жилкования; в, г – G. glinka sp. nov., голотип ГГМ-1844-01/БП-12884: в – общий вид, г – прорисовка жилкования. Длина масштабной линейки 1 мм.

В качестве непосредственного предка вячеславий рассматривался род Sadovnikovia Naugolnykh из кунгурского яруса Приуралья. Этот монотипический род с единственным видом S. belemnoides Naugolnykh встречается очень редко, однако благодаря нескольким находкам хорошо сохранившихся спорофиллов, включая голотип (Наугольных, 1994), удалось выяснить, что садовниковия, в отличие от вячеславий, была невысоким, по всей видимости, травянистым растением (рис. 1, и), внешне очень сходным с современными гетероспоровыми плауновидными рода Isoetes L. По всей видимости, именно садовниковиям или каким-то близким плауновидным принадлежали дисперсные мегаспоры морфологического типа Laevigatisporites, изредка встречающиеся в отложениях кунгурского яруса Приуралья (табл. VII , фиг. 5 ). Подробное рассмотрение экологии пермских и триасовых гетероспоровых плауновидных приведено в работах одного из авторов (Naugolnykh, 2012, с. 398, 399; 2013, с. 2–5, 13).
Установление возможной трофической связи блаттинопсид и гетероспоровых плауновидных хорошо согласуется с той ролью, которую гетероспоровые плауновидные играли в пермских палеофитоценозах. Предполагается (Наугольных, 2007), что эти растения образовывали монодоминантные пионерные фитоценозы, широко распространившиеся в начале уфимского века. Именно поэтому вячеславии составили основной пищевой субстрат для блаттинопсид в соликамское время, что обусловило вспышку численности этих насекомых.
Подводя итог всему вышесказанному, можно предположить, что резкий скачок доли блаттинопсид в местонахождениях начала уфимского века, возможно, был связан с развитием специфических ландшафтных обстановок, в которых доминировали древовидные гетероспоровые плауновидные, а также мохообразные.
В отношении более общих данных, косвенно свидетельствующих о трофических преференциях блаттинопсид, следует процитировать В.В. Жерихина, писавшего, что водные насекомые (поденки, некоторые жуки и ручейники) питаются бентосными водорослями, но ни одного такого указания для наземных насекомых не приводится, и далее, что “…свидетельства питания (палеозойских) членистоногих живыми растениями существуют для ликопсид, хвощей (форм-род Schizoneura (Equisetopsida incertae sedis)), папоротников отряда Marattiales” (Zherikhin, 2002, с. 338, здесь и далее наш перевод). Далее даны дополнительные косвенные указания на трофические предпочтения палеозойских насекомых: “в этой связи интересно отметить, что современные папоротники, хвощи и особенно плауны имеют бедную и своеобразную фауну фитофагов. В последнем случае я вообще никогда не видел насекомых, питающихся современными Lycopodium или Selaginella, и смог найти только два опубликованных указания на насекомых-фитофагов, гусеницу бабочки Satyrinae в Сев. Америке и прямокрылое Eumastacidae в Индонезии” (Zherikhin, 2002, с. 342).
О диете ранних крылатых насекомых там же отмечено: “…ранние Pterygota питались преимущественно живыми тканями растений, наиболее богатыми азотом, такими как споры, пыльца и семена. Их пищевые стратегии первоначально должны были быть оппортунистическими и не слишком избирательными в отношении как таксономической принадлежности растений, так и используемых органов. Больные и поврежденные растительные ткани могли использоваться вместе со здоровыми, т.к. наличие там микроорганизмов и грибов могло служить дополнительным источником азота” (Zherikhin, 2002, с. 338).
Таким образом, авторы настоящей статьи сходятся во мнении, что древние наземные (не водные) насекомые, включая блаттинопсид, если и были фитофагами, то питались по преимуществу спорами, пыльцой и семенами/семязачатками, т.е., были фитэмбриофагами (рис. 2). Поскольку блаттинопсиды с их широкими крышевидно складывавшимися крыльями жили, вероятно, на растениях, и были не слишком приспособлены к обитанию в скважинах субстрата (включая тронутую грибами древесину), где больше всего мертвого растительного и животного материала, и не обнаруживают приспособлений для хищничества, скорее всего, они были фитэмбриофагами (Расницын, 1980; Rasnitsyn, Quicke, 2002).
Описанный в настоящей работе материал хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН) и Государственном геологическом музее им. В.И. Вернадского РАН (ГГМ). Авторы признательны А.Г. Пономаренко (ПИН РАН) и С.М. Снигиревскому (СПбГУ) за замечания по рукописи. Работа выполнена в рамках Госзадания ГИН РАН № 0135-2019-0044 “Фитостратиграфия, палеофлористика, кризисные события кайнозоя, мезозоя и палеозоя различных регионов Евразии, палеоклиматические, палеофитоценотические и палеофитогеографические реконструкции”.
ОТРЯД BLATTINOPSIDA
СЕМЕЙСТВО BLATTINOPSIDAE BOLTON, 1925
Род Glaphyrophlebia Handlirsch, 1906
Определительная таблица видов рода Glaphyrophlebia
В определительную таблицу не включены недостаточно сохранившиеся G. rossica Martynov, 1931 из местонахождения Тихие Горы (средняя пермь России) и G. glinka sp. nov. из местонахождения Красная Глинка (нижняя пермь России).
1(10) М ветвится (помимо конечных развилков).
2(3) Основание RS слито с R, RS отходит от R двумя или тремя стволами. R за основанием RS изогнут S-образно. Длина переднего крыла 10.5 мм. Местонахождение Мидко, нижняя пермь США........................................................ .................G. anderhalterorum Beckemeyer, 2013.
3(2) Основание RS не слито с R, RS отходит от R одним стволом. R за основанием RS не изогнут S-образно.
4(7) CuA правильно гребенчатая назад.
5(6) Ветви RS простые, лишь с концевыми развилками. R менее чем с 10 передними ветвями. Длина переднего крыла 8.5 мм. Местонахождение Обора, нижняя пермь Чехии................... .......................................G. clava Kukalová, 1965.
6(5) Ветви RS с основными и концевыми развилками. R более чем с 15 передними ветвями. Длина переднего крыла 21 мм. Местонахождение Нидермошель, нижняя пермь Германии.... ...............G. arnulfi Hörnschemeyer et Stapf, 2001.
7(4) CuA разделяется на переднюю ветвь с одним–двумя концевыми развилками и гребенчатую заднюю.
8(9) SC изогнута S-образно. Длина переднего крыла 12.5 мм. Местонахождение Нидермошель, нижняя пермь Германии.......................... .......G. jeckenbachi Hörnschemeyer et Stapf, 2001.
9(8) SC не изогнута S-образно. Длина переднего крыла 16.5–17.1 мм. Местонахождение Нидермошель, нижняя пермь Германии..................... ........G. rohwedderi Hörnschemeyer et Stapf, 2001.
10(1) M простая или с концевым развилком из более тонких, чем ствол, ветвей.
11(22) RS отходит от R двумя стволами.
12(15) CuA правильно гребенчатая назад.
13(14) Вершина SC далеко заходит за уровень нодальной складки. R в дистальной половине сильно S-образно изогнут. Длина переднего крыла 9.5 мм. Местонахождение Нидермошель, нижняя пермь Германии.......................... ..........G. parvavena Hörnschemeyer et Stapf, 2001.
14(13) SC короткая, едва достигает базального уровня нодальной складки. R слабо S-образно изогнут. Длина переднего крыла около 12 мм. Местонахождение Тюлькино, средняя пермь России................................................................ .........................G. rara Aristov et Rasnitsyn, 2022.
15(12) CuA перед своей дистальной третью разделяется на две ветви, задняя c не менее чем тремя ветвями.
16(19) RS правильно гребенчатый (только с концевыми развилками). SC короткая (едва достигает уровня основания нодальной складки). CuA с обособленной передней ветвью.
17(18) SC далеко заходит за уровень основания нодальной складки. В субкостальном поле поперечные жилки простые. Местонахождение Сояна, средняя пермь России............................ ...........................G. iva Aristov et Rasnitsyn, 2021.
18(17) SC едва достигает уровня нодальной складки. В субкостальном поле поперечные жилки ветвятся. Местонахождение Тюлькино, средняя пермь России............................................... ...............G. kamaensis Aristov et Rasnitsyn, 2022.
19(16) Ветви RS с основными и концевыми развилками. CuA без обособленной передней ветви.
20(21) Передние ветви SC и R прямые, слабо скошенные. Костальное поле в базальной трети крыла равно по ширине субкостальному. Длина переднего крыла 11–12 мм. Местонахождение Чекарда, нижняя пермь России.................. ..............................G. uralensis (Martynov, 1940).
21(20) Передние ветви SC и R сильно скошены и, последние, сильно изогнуты. Костальное поле в базальной трети крыла значительно шире субкостального. Длина переднего крыла около 9.5 мм. Воркутское месторождение, средняя пермь России...................................................... ......................G. vorkutensis Aristov, Rasnitsyn et Naugolnykh, 2021.
22(11) RS отходит от R одним стволом.
23(24) CuA с семью ветвями. SC очень короткая, далеко не достигает базального уровня нодальной складки. Длина переднего крыла 8 мм. Местонахождение Эльмо, нижняя пермь США........................G. speciosa (Sellards, 1909).
24(23) CuA с 10 и более ветвями. SC достигает базального уровня нодальной складки.
25(26) R с 25 ветвями на переднем крае крыла. Длина переднего крыла 8 мм. Местонахождение Эльмо, нижняя пермь США........................ ......................................G. ovata (Sellards, 1909).
26(25) R с менее чем 20 ветвями на переднем крае крыла.
27(32) CuA перед своей дистальной третью разделяется на две ветви, задняя c не менее чем тремя ветвями.
28(29) Основания двух задних ветвей CuA соединены в четкий развилок. Длина переднего крыла 9–10 мм. Воркутское месторождение, средняя пермь России........................................ ........G. borea Aristov, Rasnitsyn et Naugolnykh, 2021.
29(28) Область оснований задних ветвей CuA дезорганизована, соединения в четкий развилок нет.
30(31) Крыло не окрашено. Длина переднего крыла 8–11 мм. Местонахождение Тюлькино, средняя пермь России........................................ ........................G. vaga Aristov et Rasnitsyn, 2022.
31(30) Крыло темное, с большим светлым пятном. Длина переднего крыла 10–11 мм. Местонахождение Сояна, средняя пермь России....... ...........................G. subcostalis (Martynov, 1928).
32(27) CuA правильно гребенчатая назад.
33(36) RS с основными и конечными развилками.
34(35) R с менее чем 10 ветвями на переднем крае крыла. Длина переднего крыла 15.5 мм. Местонахождение Вайссиг, нижняя пермь Германии.. ...........................G. oblonga (Deichmüller, 1882).
35(34) R с более чем 10 ветвями на переднем крае крыла. Длина переднего крыла 10 мм. Местонахождение Чекарда, нижняя пермь России.......... .....................G. semipenna Aristov, Rasnitsyn et Naugolnykh, sp. nov.
36(33) RS правильно гребенчатый. R с более чем 10 ветвями на переднем крае крыла. Длина переднего крыла 9.5–11 мм. Воркутское месторождение, средняя пермь России...................... ...............G. komia Aristov, Rasnitsyn et Naugolnykh, 2021.
Glaphyrophlebia semipenna Aristov, Rasnitsyn et Naugolnykh, sp. nov.
Название вида от semi лат. – полу- и penna лат. – крыло. Название дано из-за характерной окраски.
Голотип – ПИН, № 1700/522, прямой и обратный отпечатки неполного переднего крыла; Россия, Пермский край, Суксунский р-н, левый берег р. Сылвы у д. Чекарда, местонахождение Чекарда; нижняя пермь, кунгурский ярус, иренский горизонт, кошелевская свита.
Описание (рис. 3, а, б). Мелкие насекомые. Передний край переднего крыла выпуклый. Костальное поле у оснований RS немного уже субкостального. SC заканчивается у середины крыла, изогнута параллельно его переднему краю, с простыми, соединенными поперечными жилками передними ветвями. RS начинается перед границей базальной трети крыла, отходит от R одним стволом. RS правильно гребенчатый (кроме одной дихотомирующей ветви), с восемью основными ветвями. М простая, почти прямая, с коротким концевым развилком. CuA начинает ветвиться после впадения М5, за своей серединой. В своей дистальной трети CuA разделена на переднюю ветвь с тремя окончаниями и гребенчатую заднюю более чем с семью ветвями. Поперечные жилки простые, Y- и Н-образные, образуют ряды ячеек в середине медиального и основании интеркубитального полей. Крыло темное, с большим светлым пятном за нодальной линией.
Размеры в мм: длина переднего крыла 10.
Сравнение. Новый вид наиболее сходен с G. jeckenbachi Hörnschemayer et Stapf, 2001 из ассельского местонахождения Нидермошель (Рейнланд-Пфальц, Германия). Новый вид отличается от него поздно ветвящейся М. У G. jeckenbachi М ветвится у своей середины (Hörnschemeyer, Stapf, 2001). От G. uralensis (Martynov, 1940) из Чекарды новый вид отличается RS, отходящим от R одним стволом. У G. uralensis RS отходит от R двумя стволами (Расницын, 1980).
Материал. Голотип.
Glaphyrophlebia glinka Aristov, Rasnitsyn et Naugolnykh, sp. nov.
Название вида от местонахождения Красная Глинка.
Голотип – ГГМ-1844-01/БП-12884, прямой и обратный отпечатки неполного переднего крыла; Россия, Пермский край, Кишертский р-н, правый берег р. Барды у д. Матвеево, местонахождение Красная Глинка; нижняя пермь, кунгурский ярус, филипповский горизонт, лекская свита.
Описание (рис. 3, в, г). Мелкие насекомые. Передний край переднего крыла выпуклый. Костальное поле у оснований RS в 1.5 раза уже субкостального. SC заканчивается за серединой крыла, не параллельна его переднему краю, с простыми передними ветвями. RS начинается перед серединой крыла, отходит от R одним стволом, гребенчатый назад, более чем с пятью ветвями. М до дистальной трети крыла простая. CuA начинает ветвиться перед впадением М5, гребенчатая. Задняя (отходящая перед вершиной М5) ветвь CuA простая. Основание простой А1 изогнуто S‑образно, А2 гребенчатая вперед, с четырьмя основными ветвями и более чем с 10 окончаниями. Поперечные жилки в интеррадиальном поле простые. Окраска в виде темной полосы в прерадиальном поле и темного пятна в основании анальной области.
Размеры в мм: ширина переднего крыла 4, длина около 10.
Сравнение. Новый вид наиболее сходен с G. borea Aristov, Rasnitsyn et Naugolnykh, 2021, от которого отличается простой задней ветвью CuA. У G. borea эта ветвь начинает ветвиться перед своей серединой (Аристов и др., 2021).
Материал. Голотип.
Список литературы
Аристов Д.С., Расницын А.П. Новый и малоизвестные виды Blattinopsidae (Insecta: Blattinopsida) из средней перми России // Палеонтол. журн. 2021. № 3. С. 49–55.
Аристов Д.С., Расницын А.П. Новые Blattinopsidae (Insecta: Blattinopsida) из средней перми европейской части России // Палеонтол. журн. 2022. № 2. С. 69–73.
Аристов Д.С., Расницын А.П., Наугольных С.В. Новые Blattinopsidae (Insecta: Blattinopsida) из пермских отложений Печорского бассейна (республика Коми, Россия) в контексте эволюции ландшафта и растительности // Палеонтол. журн. 2021. № 6. С. 55–62.
Деревягин В.С., Свидзинский С.А., Седлецкий В.И. и др. Нижнепермская галогенная формация Северного Прикаспия. Ростов-на-Дону: Изд-во Ростовского ун-та, 1981. 397 с.
Жарков М.А. Палеогеографические перестройки и обстановки седиментации в перми и раннем триасе // Климат в эпохи крупных биосферных перестроек / Ред. Семихатов М.А., Чумаков Н.М. М.: Наука, 2004. С. 158–180 (Тр. Геол. ин-та РАН. Вып. 550).
Мартынов А.В. Пермские ископаемые насекомые Чекарды // Тр. Палеонтол. ин-та АН СССР. 1940. Т. 11. Вып. 1. С. 5–62.
Наугольных С.В. Новый лепидофит из кунгура Среднего Приуралья // Палеонтол. журн. 1994. № 4. С. 131–136.
Наугольных С.В. Viatcheslavia vorcutensis Zalessky (плауновидные): морфология, систематика, палеоэкология // Палеонтол. журн. 2001. № 2. С. 97–102.
Наугольных С.В. Ископаемые растения из верхней перми Пермского Приуралья (коллекция Г.Т. Мауэра) в Государственном Геологическом музее им. В.И. Вернадского РАН // VM-Novitates. Новости из Геол. музея им. В.И. Вернадского. 2005. № 13. 44 с.
Наугольных С.В. Пермские флоры Урала. М.: Геос, 2007. 322 с. (Тр. Геол. ин-та РАН. Вып. 524).
Наугольных С.В. Flora Permica. Растительный мир пермского периода. Приуралье. М.: Геос, 2016. 336 с. (Тр. Геол. ин-та РАН. Вып. 612).
Наугольных С.В., Литвинова Т.В. Условия формирования отложений соликамского горизонта (уфимский ярус) и новые данные о пермских строматолитах Среднего Приуралья (бассейн р. Чусовой, Пермский край) // Тр. Палеонтол. об-ва. Т. 4. М.: ПИН РАН, 2020. С. 57–72.
Расницын А.П. Надотряд Caloneuroidea. Отряд Blattinopsida // Историческое развитие класса насекомых / Ред. Родендорф Б.Б., Расницын А.П. М.: Наука, 1980. С. 38–41 (Тр. Палеонтол. ин-та АН СССР. Т. 178).
Расницын А.П., Сукачева И.Д., Аристов Д.С. Пермские насекомые воркутской серии Печорского бассейна и их стратиграфическое значение // Палеонтол. журн. 2005. № 4. С. 63–75.
Силантьев В.В. Соликамский горизонт Пермского Приуралья // Стратотипы и опорные разрезы верхней перми Поволжья и Прикамья. Казань: Экоцентр, 1996. С. 13–55.
Сонин Г.В. Морфогенез жизненных форм и эколого-морфологическая классификация строматолитов // Научные перспективы XXI века. Достижения и перспективы нового столетия. Часть 3. 5(12). Новосибирск, 2015. С. 158–166.
Hörnschemeyer T. Fossil insects from the Lower Permian of Niedermoschel (Germany) // Proc. 1st Intern. Palaeoentomol. Conf. Moscow. 1998. Bratislava: AMBA projects, 1999. P. 57–60.
Hörnschemeyer T., Stapf H. Review of Blattinopsidae (Protorthoptera) with description of new species from the Lower Permian of Niedermoschel (Germany) // N. Jb. Geol. Paläontol. Abhand. 2001. V. 221. № 1. P. 81–109.
Naugolnykh S.V. Sporophyll morphology and reconstruction of the heterosporous lycopod Tomiostrobus radiatus Neuburg emend. from the Lower Triassic of Siberia (Russia) // Palaeobot. 2012. V. 61. P. 387–405.
Naugolnykh S.V. The heterosporous lycopodiophyte Pleuromeia rossica Neuburg, 1960 from the Lower Triassic of the Volga River basin (Russia): organography and reconstruction according to the ‘Whole-Plant’ concept // Wulfenia. 2013. V. 20. P. 1–16.
Naugolnykh S.V. Fossil flora and stratigraphy of the terrigenous Kungurian beds (Lower Permian) of the basin of the Barda River (Urals, Perm krai) // Stratigr. Geol. Correlation. 2014. V. 22. №. 7. P. 680–707.
Naugolnykh S.V. Fossil flora from the Kazanian (Middle Permian) Iva-Gora locality, Soyana River, Arkhangelsk Region, Russia // Paleontol. J. 2015. V. 49. № 11. P. 1193–1205.
Naugolnykh S.V. Lower Kungurian shallow-water lagoon biota of the Middle Cis-Urals, Russia: towards the paleoecological reconstruction // Global Geol. 2017. V. 20. № 1. P. 1–13.
Naugolnykh S.V. Lower Permian conifers of the Urals: taxonomic and morphological diversity and paleoecology // Paleontol. J. 2018. V. 52. № 7. P. 34–51.
Naugolnykh S.V., Zavialova N.E. Densoisporites polaznaensis sp. nov.: with comments on its relation to Viatcheslavia vorcutensis Zalessky // Palaeobot. 2004. V. 53. P. 21–33.
Rasnitsyn A.P., Aristov D.S. New species of the genus Blattinopsis Giebel, 1867 (Insecta: Blattinopsida: Blattinopsidae) from the Permian of Taimyr Peninsula, Russia // Far East. Entomol. 2021. № 437. P. 6–9.
Rasnitsyn A.P., Quicke D.L.J. (eds.). History of Insects. Dordrecht: Kluwer Acad. Publ., 2002. 417 p.
Silantiev V.V. Solikamskian Horizon of the Permian in the Fore-Urals // Stratotypes and reference sections of the Upper Permian in the regions of the Volga and Kama Rivers. Moscow: Geos, 1998. P. 11–31.
Tang Z., Zhang Y., Naugolnykh S.V. et al. Ufadendron elongatum sp. nov., an Angaran lycopsid from the Upper Permian of Inner Mongolia, China // J. Earth Sci. 2020. V. 31. № 1. P. 1–8.
Zherikhin V.V. Ecological history of terrestrial insects // History of Insects / Eds. Rasnitsyn A.P., Quicke D.L.J. Dordrecht: Kluwer Acad. Publ., 2002. P. 331–388.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал