

Палеонтологический журнал, 2022, № 5, стр. 58-78
Некоторые новые остракоды из келловея–нижнего оксфорда Русской плиты (роды Camptocythere Triebel и Procytherura Whatley) и их стратиграфическое значение
Е. М. Тесакова a, b, *, В. Б. Сельцер c, **
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
b Геологический институт РАН
119017 Москва, Россия
c Саратовский национальный исследовательский государственный университет им. Н.Г. Чернышевского
410012 Саратов, Россия
* E-mail: ostracon@rambler.ru
** E-mail: seltservb@mail.ru
Поступила в редакцию 20.12.2021
После доработки 21.02.2022
Принята к публикации 21.02.2022
- EDN: UGASFC
- DOI: 10.31857/S0031031X22050129
Аннотация
Изучены остракоды родов Camptocythere Triebel и Procytherura Whatley из отложений нижнего и среднего келловея опорного разреза ТЭЦ-5 (Саратовская обл.) и среднего келловея–нижнего оксфорда опорного разреза Михайловцемент (Рязанская обл.). Проведена ревизия рода Camptocythere; из его состава исключены представители рода Aparchitocythere Swain et Peterson, который вновь рассматривается в качестве самостоятельного рода, и напротив, род Palaeoloxoconcha Dreyer впервые включается в качестве подрода. Описаны новые виды: C. (Camptocythere) quinta Tesakova, sp. nov., C. (Palaeoloxoconcha) caudata Tesakova, sp. nov. [с подвидами C. (P.) caudata caudata Tesakova, subsp. nov. и C. (P.) caudata nuda Tesakova, subsp. nov.], C. (P.) ryazanica Tesakova, sp. nov. и Procytherura ippolitovi Tesakova, sp. nov. В пределах Восточно-Европейской платформы (ВЕП) выделены две остракодовые зоны совместного распространения. В нижнем келловее (аммонитовая зона Subpatruus, биогоризонты Subpatruus–Saratovensis) Украины, Курской обл. и Поволжья выделена зона A. milanovskyi–P. cinicinnusa. В нижнем келловее (аммонитовая зона Subpatruus, биогоризонт Saratovensis)–среднем келловее (зона Jason) Украины, Белоруссии, Центрально-Черноземной и Курской обл. и Среднего Поволжья выделена зона Pr. wartae–Pl. kurskensis. По эволюции подрода C. (Palaeoloxoconcha) предложены новые филозоны. Зона C. (P.) bathonica установлена в Северо-Западной Европе в верхнем байосе–бате. Зона C. (P.) caudata прослеживается на ВЕП (Белоруссия, Курская и Саратовская обл.) в нижнем келловее (зона Subpatruus, биогоризонт Saratovensis)–нижней части среднего келловея (зона Jason). Зона C. (P.) ryazanica выделена в Рязанской обл. в верхней части среднего келловея (зона Coronatum)–нижнем оксфорде (зона Mariae).
ВВЕДЕНИЕ
Многолетнее изучение юрских ракушковых раков Восточно-Европейской платформы (ВЕП) показало высокую насыщенность ими осадочных пород и возможность использования группы в биостратиграфии. К концу XX в. был накоплен солидный блок информации о систематическом составе остракодовой фауны Поволжья, Центрально-Черноземной области, Днепровско-Донецкой впадины, Общего Сырта, а также юго-западного (Донбасс) и юго-восточного (Эмбинский р-н, Мангышлак, Узбекистан) обрамлений ВЕП (обзор соответствующей литературы см.: Тесакова, 2020). Последующее изучение остракод на сканирующем микроскопе выявило необходимость ревизии старых таксонов (Тесакова, 2013а, б и др.) и открыло новые пути для классификации. В современных публикациях, наряду с описаниями новых видов из центральных районов ВЕП, Поволжья и Тимано-Печорской провинции (Колпенская, 1993; Tesakova, 2003 и др.), появилось большое число таксонов, оставленных в открытой номенклатуре (Тесакова, Сельцер, 2013; Tesakova, 2013 и др.). Их упоминание в составе объемных зональных комплексов до последнего времени не создавало затруднений для биостратиграфии, поскольку юрская шкала по остракодам ВЕП состоит в основном из комплексных зон (Колпенская, 1999; Тесакова, 2014). Но разработка новых параллельных филогенетических шкал на основе эволюции разных родов (напр., Никитенко, 1994, 2009) повлекла за собой необходимость валидизировать стратиграфически значимые виды. Особое внимание уделялось остракодам из отложений, хорошо охарактеризованных аммонитами и расчлененных с точностью до биогоризонтов.
Одним из таких важных для стратиграфии разрезов – опорным для нижнего–среднего келловея Среднего Поволжья – является обнажение ТЭЦ-5 (Саратовская обл.) с богатыми комплексами аммонитов (Попов и др., 2004; Митта, Сельцер, 2006; Gulyaev, Ippolitov, 2021) и микрофауной (Глинских и др., 2022). По распределению остракод в этом разрезе ранее было выделено четыре комплекса, предположительно, связанных с менявшимися экологическими условиями келловейского бассейна (Глинских и др., 2022), но также могущих иметь стратиграфическое значение. Для двух из них в качестве индексов были выбраны представители рода Camptocythere Triebel, 1950 потому, что по его эволюции ранее были реконструированы филолинии подродов C. (Camptocythere) Triebel, 1950 и C. (Anabarocythere) Nikitenko, 1994, легшие в основу филогенетических шкал нижней и средней юры для Западной Европы и Сибири (Triebel, 1950; Plumhoff, 1963; Никитенко, 1994, 2009). Анализ распространения камптоцитер из впервые рассматриваемой в настоящей работе филолинии подрода C. (Palaeoloxoconcha) Dreyer, 1967 может оказаться полезным для разработки региональных филозон в средней юре ВЕП.
В задачи настоящей статьи входит ревизия рода Camptocythere, описание некоторых новых камптоцитер (ранга вида и подвида) и одного представителя семейства Cytheruridae G.W. Mueller из нижнего и среднего келловея разреза ТЭЦ‑5 и среднего келловея–нижнего оксфорда разреза Михайловцемент (Рязанская обл.). Другой задачей статьи является определение стратиграфического диапазона остракодовых комплексов, предложенных Е.М. Тесаковой (Глинских и др., 2022), и объема выделяемых камптоцитеровых филозон путем сопоставления с аммонитовыми биогоризонтами.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для статьи послужили остракоды из отложений нижнего (аммонитовые зоны Subpatruus, Koenigi и Calloviense) и среднего (аммонитовая зона Jason) келловея разреза ТЭЦ-5 (Саратовская обл.) (рис. 1), встреченные в семи из 24 образцов глин, отобранных в разные годы различными рабочими коллективами (Глинских и др., 2022); а также коллекции остракод из среднего келловея–нижнего оксфорда (аммонитовые зоны Coronatum–Mariae) разреза Михайловцемент (Рязанская обл.) (Tesakova, Shurupova, 2018; настоящая работа). Микрофауна выделялась из проб стандартным методом (кипячением с содой с последующим промыванием в воде на ситах с ячеей 0.01 или 0.05 мм), а ее отбор производился под бинокулярами Stemi 2000-С (Carl Zeiss) и Advance ICD (Bresser). Разрез ТЭЦ-5 оказался обеднен остракодами, большей частью разрушившимися при выветривании и представленными единичными экземплярами немногих видов (в общей сложности 14 таксонами) на семи уровнях обнажения (рис. 1). Найденные виды, по-видимому, были доминантами в комплексах на соответствующих интервалах разреза.
Рис. 1.
Расчленение разреза ТЭЦ-5 по остракодам. Литологическая колонка, карта-схема с расположением разреза (отмечен стрелкой) и аммонитовая зональность приводится по (Gulyaev, Ippolitov, 2021, с добавлениями В.Б. Сельцера). Условные обозначения: 1 – глины, 2 – глины алевритовые, 3 – алевриты глинистые, 4 – мергели, 5 – размывы (перерывы в осадконакоплении), 6 – скопления раковинных остатков, 7 – скопления ростров белемнитов, 8 – фосфориты, 9 – линзы и конкреции. Римскими цифрами в кружках обозначены Т/Р циклиты, связанные с трансгрессией из: I – Бореального бассейна, II – Западной Европы.
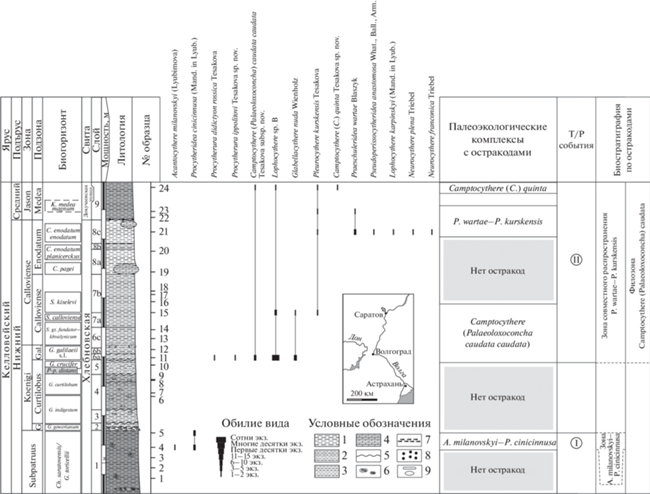
Фотографирование производилось на сканирующих электронных микроскопах в ИГМ СО РАН (LEO-1430VP) и в кабинете приборной аналитики ПИН РАН (TESCAN VEGA-II XMU) (Глинских и др., 2022). Изображения всех таксонов остракод, изученных из разреза ТЭЦ-5, представлены на таблицах IV и V (см. вклейку); экземпляры из разреза Михайловцемент – на рис. 4.
Поскольку представители новых видов имеют широкое распространение за пределами стратотипической местности и уже определялись в открытой номенклатуре в нижнем келловее Гомельского р-на Белоруссии (Махнач, Тесакова, 2015) и Курской области (Tesakova, 2013), при написании статьи, кроме остракод из ТЭЦ-5 и Михайловцемента, использовались коллекции Е.М. Тесаковой, собранные в упомянутых регионах ранее (рис. 2).
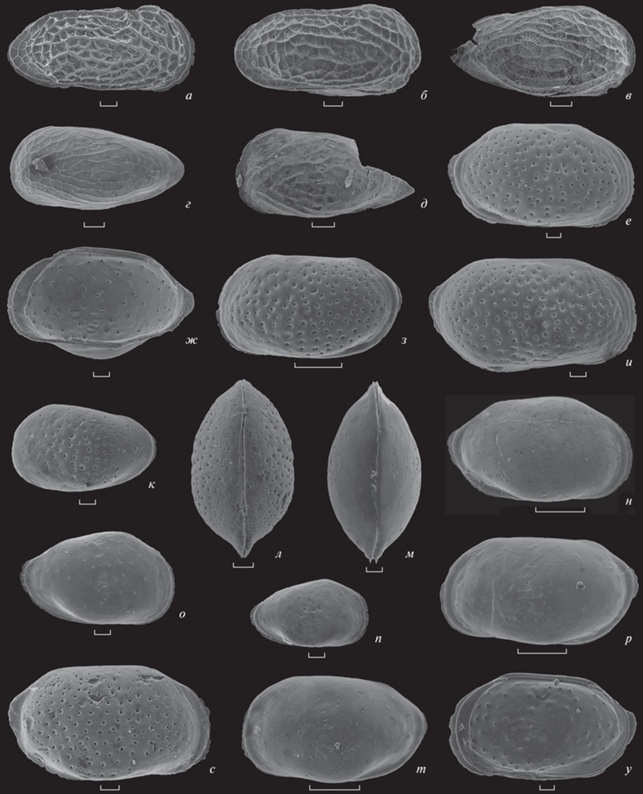
Рис. 2.
Остракоды: а–д – Procytherura ippolitovi Tesakova, sp. nov.: а – экз. МГУ, № 300-Kusk-199, пс самки, скв. 7, гл. 134 м [=Procytherura pleuraperiousios in: Tesakova, 2013, табл. 5 , фиг. 14 ]; б – экз. МГУ, № 370-Belor-1-25, ц.р. самки справа [=Procytherura pleuraperiousios in: Махнач, Тесакова, 2015, табл. 1 , фиг. 10 ]; в–д – [=Procytherura tenuicostata in: Махнач, Тесакова, 2015]: в – экз. МГУ, № 370-Belor-1-141, пс juv., г – экз. МГУ, № 370-Belor-2-14, цр juv. слева (Махнач, Тесакова, 2015, табл. 1 , фиг. 15, 16 ), д – экз. МГУ, № 370-Belor-1-130, обл. лс juv.; е–л, с – Camptocythere (Palaeoloxoconcha) caudata caudata Tesakova, sp. et subsp. nov.: е – голотип МГУ, № 300-Ky-1-31, пс самки, скв. 7, гл. 137 м; ж – экз. МГУ, № 300-Kusk-131, пс самки изнутри, скв. 4, гл. 125 м; з – экз. МГУ, № 300-Ky-1-32, лс самки, скв. 7, гл. 137 м; и – экз. МГУ, № 300-Kusk-179, пс самца, скв. 7, гл. 131 м; к – экз. МГУ, № 300-Ky-1-37, лс juv., скв. 7, гл. 137 м; л – экз. МГУ, № 370-Belor-1-32, цр juv. со спинной стороны; с – экз. МГУ, № 370-Belor-1-52, пс самца; м–р, т, у – C. (P.) caudata nuda Tesakova, sp. et subsp. nov.: м – экз. МГУ, № 300-Kusk-124, цр juv. со спинной стороны, скв. 4, гл. 124 м; н – голотип МГУ, № 300-Ky-2-90, пс самки, скв. 7, гл. 138 м; о – экз. МГУ, № 300-Kusk-125, пс juv., скв. 4, гл. 124 м; п – экз. МГУ, № 300-Ку-2-57, пс juv. скв. 7, гл. 138 м; р – экз. МГУ, № 300-Ky-2-89, лс самца, скв. 7, гл. 138 м; т – экз. МГУ, № 300-Ky-2-88, лс самки, скв. 7, гл. 138 м; у – экз. МГУ, № 300-Kusk-90, пс самки изнутри, скв. 4, гл. 119 м. Экз. на фиг. б–д, л, с происходят из нижнего келловея, остракодовая зона P. wartae–P. kurskensis, скв. 792, Гомельский р-н Белоруссии (Махнач, Тесакова, 2015). Экз. на фиг. а, е–к, м–р, т, у из нижнего келловея, остракодовая филозона Palaeocytheridea parabakirovi, интервал-зона P. wartae–P. kurskensis Курской обл. (Tesakova, 2013). Длина шкалы на фиг. а–ж, и‒м, о, п, с, у составляет 30 мкм; на фиг. з, н, р, т – 100 мкм.
Коллекции остракод из разреза ТЭЦ-5 хранятся в лаб. микропалеонтологии ИНГГ СО РАН под № ИНГГ ТЭЦ-5-О и на каф. региональной геологии и истории Земли МГУ под № МГУ ТЭЦ-5. На той же кафедре хранятся остракоды под №№ МГУ МС-2017 и МГУ MC2 (Рязанская обл., разрез Михайловцемент), МГУ 370 (Белоруссия, Гомельский р-н, скв. 792) и МГУ 300 (Курская обл., Фатежский р-н, скв. 4 и 7).
Для научных учреждений, упомянутых в настоящей статье, приняты следующие аббревиатуры: ГИН (Геологический ин-т РАН, Москва), ИНГГ (Ин-т нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН, Новосибирск), ИГМ (Ин-т геологии и минералогии им. В.С. Соболева СО РАН, Новосибирск), МГУ (Московский государственный ун-т им. М.В. Ломоносова), ПИН (Палеонтологический ин-т им. А.А. Борисяка РАН, Москва).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Систематика надродовых таксонов принята по “Практическому руководству…” (1999) и приведена в соответствии с “Международным кодексом…” (2004), терминология различных элементов раковин остракод взята по “Практическому руководству…” (1989). Для обозначения измерений раковин приняты следующие сокращения: Д – длина, В – высота, Т – толщина раковины, Д/Т – отношение длины к высоте, ВПК – высота переднего конца, ВЗК – высота заднего конца, juv. – ювенильная особь.
ОТРЯД PODOCOPIDA
ПОДОТРЯД CYTHEROCOPINA
НАДСЕМЕЙСТВО PROGONOCYTHEROIDEA SYLVESTER-BRADLEY, 1948
СЕМЕЙСТВО PROGONOCYTHERIDAE SYLVESTER-BRADLEY, 1948
ПОДСЕМЕЙСТВО PROGONOCYTHERINAE SYLVESTER-BRADLEY, 1948
Род Camptocythere Triebel, 1950
Camptocythere: Triebel, 1950, с. 198; Лев, 1958, с. 41; Кашеварова и др., 1960, с. 376; Howe, 1961, с. 257; Plumhoff, 1963, с. 41 [part.]; Никитенко, 1994, с. 51 [part.]; Практическое …, 1999, с. 64 [part.].
Типовой вид – С. praecox Triebel, 1950 из средней юры, нижнего аалена, аммонитовой зоны Opalinus Северо-Западной Германии (Triebel, 1950, с. 199, табл. 1 , фиг. 1–10 ; табл. 2 , фиг. 11, 12 ).
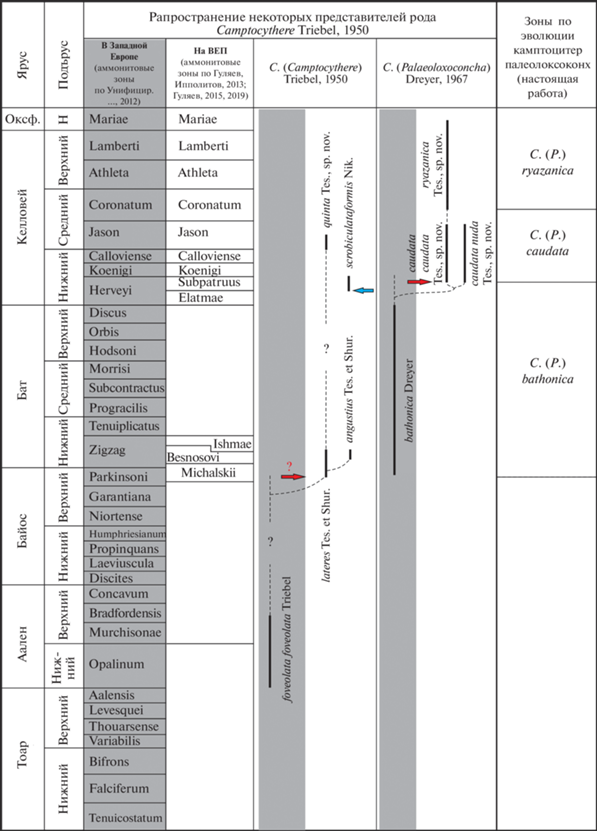
Рис. 3.
Реконструкция филолиний изученных камптоцитер. Территория Северо-Западной Европы выделена серым цветом, территория Восточно-Европейской платформы – белым. Синими стрелками показаны миграции камптоцитер на ВЕП из Западной Сибири и Баренцевоморского шельфа, красными – из Северо-Западной Европы. Сплошной чертой показано достоверное распространение вида.
Диагноз. Раковина среднего размера, субовальная (яйцевидная), умеренно выпуклая, с наибольшей толщиной в середине створки. При виде со спинной стороны контур раковины линзовидный, эллипсовидный или цилиндрический. Передний конец плавно закруглен, задний конец тоже плавно закруглен или округло-треугольный; у самок передний конец выше заднего, у самцов обратное соотношение. Спинной край левой створки прямой, у правой створки – выпуклый. Левая створка больше правой и охватывает ее по брюшному краю или по переднеспинному и заднеспинному углам, правая охватывает левую по спинному краю. Замок лофодонтный, краевые пластинчатые зубы правой створки слабо насечены: передний на три–четыре отдела, задний на четыре–пять, между ними расположен гладкий желобок. Отпечаток аддуктора в виде слабовыпуклого к заднему концу вертикального ряда из четырех продолговатых рубцов. Порово-канальная зона умеренно широкая или узкая. Может быть развита краевая кайма. Поверхность гладкая, ямчатая или ячеистая, может нести мускульный бугорок. В заднебрюшной части створки (одной или обеих) могут быть развиты валики, бугорки или шипы. Поры простые и ситовидные. Половой диморфизм выражается в бóльших размерах раковин и более высоком заднем конце у самцов.
Видовой состав. Более 30 видов и подвидов в составе трех подродов: C. (Camptocythere) Triebel, 1950, C. (Anabarocythere) Nikitenko, 1994 и C. (Palaeoloxoconcha) Dreyer, 1967 из нижней и средней юры (тоар–келловей) бореальной и суббореальной палеобиогеографических провинций Северного полушария (Канада, Западная и Восточная Европа, север Сибири) (Шарапова, 1940; Triebel, 1950; Лев, 1958, 1961, 1966; Plumhoff, 1963; Bate, Coleman, 1975; Riegraf, 1985; Никитенко, 1994, 2009; Басов и др., 2009; Тесакова, Сельцер, 2013; Dietze et al., 2017, 2018; Tesakova, 2017; Franz et al., 2018; Wannenmacher et al., 2021 и др.). Один вид, C. (P.) ryazanica Tesakova sp. nov., (из отложений аммонитовых зон Coronatum–Mariae в Рязанской обл.) проходит в верхнюю юру.
Сравнение. От наиболее похожего по форме раковины и лофодонтному замку рода Aparchitocythere Swain et Peterson, 1952 из верхней юры Сев. Америки (Кашеварова и др., 1960, с. 376; Howe et al., 1961, с. 290) отличается слабо насеченными (против гладких) краевыми зубами замка.
Замечания. Сравнение первоописания Camptocythere (где внимание акцентировалось на слабо насеченные краевые пластинчатые зубы замка; Triebel, 1950, с. 199) со всеми последующими описаниями этого рода (Кашеварова и др., 1960; Howe, 1961; Plumhoff, 1963; Никитенко, 1994; Практическое …, 1999) заставило нас пересмотреть его объем. Во-первых, из состава Camptocythere исключены представители рода Aparchitocythere Swain et Peterson, 1952, введенные туда Ф. Плюмовым (Plumhoff, 1963) и упомянутые в составе описываемого рода Б.Л. Никитенко (1994) и в “Практическом руководстве…” (1999), и восстановлен статус Aparchitocythere в качестве самостоятельного рода. Во-вторых, в состав рода Camptocythere в качестве подрода нами включена группа видов, обладающих всеми соответствующими родовыми признаками, но ранее выделявшихся в самостоятельный род Palaeoloxoconcha Dreyer, 1967 (см. ниже).
Кроме того, новые виды из подрода C. (Palaeoloxoconcha) расширяют стратиграфический диапазон рода до низов верхней юры (нижнего оксфорда).
Во всех публикациях, упомянутых в синонимике [кроме Никитенко (1994), с первоописанием подрода C. (Anabarocythere)], содержится описание морфологии и скульптуры раковин камптоцитер, относящихся только к номинативному подроду. Поэтому в настоящей статье приводится обновленный диагноз рода, учитывающий морфологические особенности всех подродов, а также содержащий актуализированную информацию об их видовом составе, стратиграфическом и географическом распространении.
Подрод Camptocythere (Camptocythere) Triebel, 1950
Сamptocythere (Camptocythere): Никитенко, 1994, с. 51 [part.].
Типовой вид – как у рода Camptocythere.
Диагноз. Наиболее полно и детально этот подрод описан Е. Трибелем (Triebel, 1950) и О.М. Лев (1958). Общая форма раковины, замок, мускульные отпечатки, порово-канальная зона и поры на боковой поверхности стенки – как у рода в целом. К особенностям подрода относятся: передний и задний концы закруглены одинаково плавно, поверхность створок гладкая или ямчатая.
Видовой состав. Более 20 видов, относящихся к номинативному подроду, известны из нижней–средней юры (нижний тоар–келловей) Северного полушария: Западной Европы (Triebel, 1950; Plumhoff, 1963; Bate, Coleman, 1975; Riegraf, 1985 и др.), Северной Сибири (Шарапова, 1940; Лев, 1958, 1961, 1966; Никитенко, 1994, 2009 и др.), Канады (Nikitenko, Mickey, 2004 и др.), Баренцевоморского шельфа и Притиманья (Лев, Кравец, 1982; Атлас …, 1995; Басов и др., 2009; Унифицированная …, 2012), а также ВЕП (Shurupova, Tesakova, 2017; Тесакова и др., 2020).
Сравнение. От подрода C. (Anabarocythere), сходного по округлой форме заднего конца, отличается отсутствием на створках шипов, бугорков или валиков, присущих только анабароцитерам. От C. (Palaeoloxoconcha), представители которого также обладают гладкой или ямчатой поверхностью раковин, отличается плавно закругленным задним концом против округло-треугольного у палеолоксоконх.
Camptocythere (Camptocythere) quinta Tesakova, sp. nov.
Табл. IV , фиг. 14–16
Camptocythere (C.) sp. 1: Глинских и др., 2022, табл. 2 , фиг. 6 .
Название вида quinta лат. – пятая, по типовому местонахождению, разрезу ТЭЦ-5.
Голотип – МГУ ТЭЦ-5-17, левая створка ювенильной особи; Саратовская обл., разрез ТЭЦ-5, обр. 24; средний келловей, аммонитовая зона Jason, подзона Medea (табл. IV, фиг. 14, 16 ).
Описание. Раковина средних размеров, эллипсовидная (яйцевидная), умеренно выпуклая, выполаживается к переднему концу более плавно, чем к заднему. Левая створка больше правой и охватывает ее по передне- и заднеспинному углам и (незначительно) по брюшному краю. Наибольшая длина чуть выше середины высоты, наибольшая высота в передней трети раковины, наибольшая толщина в заднебрюшной части. Спинной край прямой, брюшной слабовыпуклый, и они конвергируют к заднему концу. Передний и задний концы симметричные, равномерно дугообразно закруглены, с брюшным краем сопрягаются плавно, в спинной край переходят через неясные тупые углы; задний конец ниже переднего и уплощен отчетливо. Поры простые и ситовидные. Поверхность створки ямчатая; размер ямок уменьшается на периферии; различаются ямки трех скульптурных типов (располагаются поштучно, выстраиваются в ряды и группируются розетками).
Размеры в мм:
Изменчивость. Незначительно выражается в большем (табл. IV , фиг. 16 ) или меньшем (табл. IV , фиг. 15 ) размере ямок. Половую и возрастную изменчивость проследить не удалось.
Сравнение. От видов с ВЕП, наиболее сходных по форме раковины и ямчатой скульптуре с аналогичными скульптурными типами – C. (C.) lateres Tesakova et Shurupova из верхнего байоса и нижнего бата Саратовской обл. (Shurupova, Tesakova, 2017, с. 127, табл. 1 , фиг. 1–4; табл. 2 , фиг. 1–9 ; табл. 3 , фиг. 1–3) и нижнего бата Пензенской обл. (рабочие коллекции Тесаковой из трех скважин и разреза Плетневского карьера), а также от C. (C.) angustius Tesakova et Shurupova из верхнего байоса Саратовской обл. (Shurupova, Tesakova, 2017, с. 129, табл. 2 , фиг. 10–15 ) отличается более округлым и симметричным задним концом и гораздо меньшим числом ямок, далеко отстоящих друг от друга (в отличие от густо расположенных ямок у сравниваемых видов). От другого ямчатого вида, C. (C.) scrobiculataformis Nikitenko, распространенного в Сибири и на Баренцевоморском шельфе в бате и нижнем келловее (Никитенко, 1994, с. 51, табл. 1 , фиг. 16–19 ; 2009, табл. 0–4, фиг. 11–15 , табл. 0-5, фиг. 1) и мигрировавшего на ВЕП в раннем келловее в фазе Subpatruus (Тесакова и др., 2020, рис. 3), отличается скульптурой: существенно меньшим числом ямок и их группировкой в скульптурные типы.
Замечания. Поскольку в материале авторов имелись только ювенильные экземпляры, полностью развитую порово-канальную зону наблюдать не удалось. Также и замок был развит не полностью: на левой створке различается трехчленное подразделение переднезубной ямки и четырехчленное – задней, что свойственно представителям A-1 и A-2 (две предпоследние ювенильные стадии) C. (C.) lateres и C. (C.) angustius (Shurupova, Tesakova, 2017).
Судя по весьма сходной скульптуре и деталям замка, C. (C.) quinta Tesakova, sp. nov. являлся потомком C. (C.) lateres, который мог проникнуть на ВЕП с позднебайосской трансгрессией только из Западной Европы, а не потомком северосибирского C. (C.) scrobiculataformis – более подходящего в качестве предка из-за времени миграции на ВЕП, но морфологически отстоящего гораздо дальше (рис. 3). Между последними представителями C. (C.) lateres из нижнего бата (зона Besnosovi) (Shurupova, Tesakova, 2017) и первыми экземплярами C. (C.) quinta sp. nov. имеется большой гиатус (почти весь бат и весь нижний келловей), что ставит под вопрос их прямое родство. Также пока нет ясности с прямыми предками C. (C.) lateres, поскольку между его появлением на ВЕП и наиболее близким по морфологии подвидом C. (C.) foveolata foveolata Triebel из нижнего и нижней части верхнего аалена Западной Европы (Triebel, 1950; Plumhoff, 1963; Никитенко, 2009, рис. 155 ) тоже существует большой стратиграфический перерыв. Тем не менее, предлагается следующая филогенетическая линия: C. (C.) foveolata foveolata (J2a1-21) → ? → C. (C.) lateres (J2b3-bt1) → → ? → C. (C.) quinta sp. nov. (J2cl21), где вопросительным знаком показаны предполагаемые, но неизвестные пока, ее члены.
Материал. Две створки хорошей сохранности из типового местонахождения.
Подрод Camptocythere (Palaeoloxoconcha) Dreyer, 1967
Palaeoloxoconcha: Dreyer, 1967, с. 53.
Типовой вид – Palaeoloxoconcha bathonica Dreyer, 1967 из верхнего байоса (верхи зоны Parkinsoni) и бата (зоны Wuerttembergica – Aspidoides) Сев. Германии (Dreyer, 1967, с. 54, табл. 1 , фиг. 1–3).
Диагноз. Габитус раковины, соотношение створок, мускульные отпечатки и порово-канальная зона как у рода. Замок: передний зуб рассечен на три части, задний – на четыре (рис. 2, ж, у). Передний конец плавно закруглен, задний конец округло-треугольного очертания. Развита широкая краевая кайма, обычно лучше сохраняющаяся на заднем конце. Поверхность створок гладкая или ямчатая. Поры простые и ситовидные.
Видовой состав. C. (P.) bathonica (Dreyer, 1967) из терминального байоса (зона Parkinsoni) – бата (зоны Wuerttembergica–Aspidoides) Сев. Германии (Dreyer, 1967, с. 54, табл. 1 , фиг. 1–3), C. (P.) caudata Tesakova, sp. nov. (с двумя подвидами) из нижнего келловея (зона Subpatruus) – нижней части среднего келловея (зона Jason) ВЕП (Tesakova, 2013; Махнач, Тесакова, 2015; настоящая работа), а также C. (P.) ryazanica Tesakova, sp. nov. из верхней части среднего келловея – нижнего оксфорда (зоны Coronatum–Mariae) Рязанской обл. (Тесакова и др., 2017; Tesakova, Shurupova, 2018).
Сравнение. От номинативного подрода и C. (Anabarocythere) отличается округло-треугольным задним концом. От подрода C. (Anabarocythere) также отличается отсутствием заднебрюшных шипов, бугорков или валиков.
Замечания. Среди келловейских остракод ВЕП Тесаковой были выделены необычные камптоцитеры, которые она считала эндемиками, не связанными родством ни с западноевропейскими, ни с арктическими/сибирскими фаунами, и долгое время оставляла в открытой номенклатуре как “Camptocythere sp. A ssp. B” и “C. sp. A ssp. A” или просто “Camptocythere sp.” (для таксонов из нижнего келловея; Tesakova, 2013; Махнач, Тесакова, 2015), и как “Camptocythere sp. A” (для таксона из среднего–верхнего келловея; Тесакова и др., 2017; Tesakova, Shurupova, 2018). Первоначально планировалось выделить новый подрод с двумя новыми видами (они описаны в настоящей статье, один – с двумя подвидами), которые составляли бы короткую доказанную филолинию. Но в литературе удалось выявить более раннего представителя той же филолинии – C. (P.) bathonica (Dreyer, 1967) (по которому Э. Драйер описала новый род Palaeoloxoconcha), поэтому новые виды Тесаковой должны быть отнесены к тому же надвидовому таксону. Однако самостоятельность Palaeoloxoconcha как рода была нами подвергнута сомнению, поскольку представители этой филолинии обладают всеми основными родовыми признаками Camptocythere (как то: размер и габитус раковины, перекрытие створок, строение замка, поверхностных пор, порово-канальной зоны и мускульных отпечатков). Тем не менее, таксон Palaeoloxoconcha целесообразно сохранить, поскольку его представители объединены родственными связями предок–потомок (что отвечает эволюции самостоятельной филолинии внутри рода) и хорошо отличаются от других камптоцитер по форме заднего конца.
Camptocythere (Palaeoloxoconcha) caudata Tesakova, sp. nov.
Табл. IV , фиг. 12, 13
Название вида – по форме заднего конца, напоминающего каудальный (хвостовой) отросток; сauda лат. – хвост.
Голотип – МГУ 300-Ky-1-31, правая створка самки; Курская обл., скв. 7, гл. 137 м; нижний келловей, остракодовая зона P. wartae–P. kurskensis (рис. 2, е).
Описание (рис. 2, е–у). Раковина средних размеров, умеренно выпуклая, равномерно выполаживается к переднему и заднему концам, оба конца уплощены (рис. 2, л, м). По свободному краю развита кайма, которая лучше сохраняется на переднем и заднем концах. Наибольшая длина расположена на середине высоты створки; наибольшая высота одинаковая на переднем и заднем концах у раковин самок и больше на заднем конце у раковин самцов; наибольшая толщина находится посередине брюшной стороны. Левая створка больше правой и охватывает ее на переднеспинном и заднеспинном углах; правая створка незначительно охватывает левую в средней части спинного края. Спинной край прямой на левой створке и слабовыпуклый на правой; брюшной край прямой, вогнут посередине створки ближе к переднему концу. Передний конец равномерно дугообразно закруглен, слабо скошен сверху (на правых створках сильнее), в брюшной край переходит плавно, а в спинной край через неясный уступ. Задний конец одной высоты с передним у самок и выше переднего конца у самцов, округло-треугольной формы; сильнее, чем передний, скошен сверху, а за счет каймы производит впечатление приподнятого кверху.
Замок и мускульные отпечатки типичные для подрода, порово-канальная зона широкая, поры простые и ситовидные. Поверхность ямчатая или гладкая.
Средние размеры в мм:
| Д | ВПК | ВЗК | Д/В | |
|---|---|---|---|---|
| Самки | 0.33–0.38 | 0.20–0.22 | 0.18–0.21 | 1.7–1.9 |
| Самцы | 0.32–0.40 | 0.16–0.20 | 0.18–0.22 | 1.8–1.9 |
Изменчивость. Довольно сильно варьируют размеры раковин у взрослых представителей. Треугольный контур заднего конца может меняться от более длинного, заостренного и приподнятого кверху до укороченного, округлого и симметричного.
Возрастная изменчивость. В зависимости от возрастной стадии экземпляра закономерно изменяется отношение длины к высоте (при взрослении оно уменьшается). Высота заднего конца раковин ювенильных экземпляров всегда ниже переднего, поэтому спинной и брюшной края у них отчетливо конвергируют к заднему концу, в то время как на раковинах взрослых особей они приблизительно параллельны из-за примерно одинаковой высоты концов (табл. IV , фиг. 13 ; рис. 2, к, о, п).
Половая изменчивость. Высота и выпуклость заднего конца самцов (рис. 2, и, р, с) превышают те же параметры у самок (рис. 2, е–з, н, т, у): если наибольшая толщина раковины самок находится посередине брюшной стороны, то у самцов – в заднебрюшной части створки, что свидетельствует о наличии зенкерова органа. Поскольку размерный разброс как у самок, так и у самцов довольно велик, самым надежным критерием различения полов является соотношение высоты переднего (ПК) и заднего (ЗК) концов: у самок всегда ПК > ЗК, у самцов всегда ПК < ЗК.
Сравнение. От весьма похожего по очертаниям раковины и ямчатой скульптуре вида C. (P.) bathonica (Dreyer, 1967) из верхнего байоса (верхи зоны Parkinsonia) и бата (зоны Wuerttembergica–Aspidoides) Северной Германии (Dreyer, 1967, с. 54, табл. 1 , фиг. 1–3) отличается меньшими размерами, более низким и длинным задним концом, более скошенным сверху, а также менее плотным расположением ямок, которые к тому же уменьшаются в размерах к периферии. От другого сходного вида C. (P.) ryazanica Tesakova, sp. nov. из среднего келловея – нижнего оксфорда (зоны Coronatum–Mariae) разреза Михайловцемент Рязанской обл. (рис. 4), отличается бóльшими размерами и параллельными спинным и брюшным краями.
Видовой состав. Два подвида: C. (P.) caudata caudata Tesakova, subsp. nov. и C. (P.) caudata nuda Tesakova, subsp. nov.
Замечания. (1) Различия между сравниваемыми видами описывают эволюцию путем педоморфоза от C. (P.) bathonica (верхний байос–бат Западной Европы) через C. (P.) caudata sp. nov. (нижний и нижняя часть среднего келловея ВЕП) к C. (P.) ryazanica sp. nov. (верхняя часть среднего келловея–нижний оксфорд ВЕП). Во-первых, размеры каждого следующего вида уменьшаются относительно таковых у предка. Во-вторых, со временем растет отношение ВПК/ВЗК, что приводит к “омоложению” контура створок у каждого последующего вида (наклон спинного и брюшного краев к заднему концу увеличивается, что наблюдается на ранних стадиях онтогенеза у всех остракод). Количество и размер ямок на поверхности створок также уменьшается от предка к потомкам, что свойственно онтогенезам видов C. (P.) caudata sp. nov. и C. (P.) ryazanica sp. nov. (чем младше возрастная стадия, тем меньше число и размер ямок, и выше тенденция к их измельчанию на периферии створки). В геологической летописи значимые гиатусы между этими тремя видами не наблюдаются (рис. 3). Все перечисленное позволяет считать последовательность из трех видов C. (P.) bathonica → C. (P.) caudata sp. nov. → → C. (P.) ryazanica sp. nov. единой филолинией, и по первому появлению в летописи каждого из этих таксонов установить нижние границы соответствующих филозон.
(2) Поскольку в коллекциях Тесаковой нет экземпляров, демонстрирующих постепенный переход между ямчатыми и почти гладкими морфами [которые ниже описаны как подвиды C. (P.) caudata caudata subsp. nov. и C. (P.) caudata nuda subsp. nov.], их нельзя считать единым видом с широкой изменчивостью. С другой стороны, обе морфы наблюдались в нижнем келловее в одной и той же биофации А, как правило, в одних и тех же образцах (Tesakova, 2013, рис. 2, 5 ). Они характеризовали один и тот же временной интервал, хорошо распознаваемый по ним на большой территории ВЕП, вследствие чего породы, насыщенные экземплярами вида C. (P.) caudata sp. nov. (s.l.), ранее выделялись в слои с P. wartae–P. kurskensis (см. раздел Биостратиграфия). При этом в образцах присутствовали либо обе морфы, с отчетливым доминированием одной из них, либо какая-нибудь одна с высоким обилием, что позволяет судить о нюансах палеоэкологии, интерпретировать эти две морфы как экологический полиморфизм и описывать в ранге подвида.
Распространение. Нижний келловей, аммонитовая зона Subpatruus, биогоризонт Saratovensis – средний келловей, нижняя часть аммонитовой зоны Jason, подзона Medea; остракодовая зона P. wartae–P. kurskensis; остракодовая филозона C. (P.) caudata; Белоруссия, Курская и Саратовская обл.
Camptocythere (Palaeoloxoconcha) caudata caudata Tesakova, subsp. nov.
Табл. IV , фиг. 12, 13
Camptocythere sp. A ssp. B: Tesakova, 2013, табл. 6 , фиг. 2.
Camptocythere sp.: Махнач, Тесакова, 2015, табл. 2 , фиг. 7, 9, 11, 12 (non табл. 2 , фиг. 8 ).
Camptocythere (P.) sp. A ssp. B sensu Tesakova, 2013: Глинских и др., 2022, табл. 2 , фиг. 5 .
Название подвида caudata лат. – хвостатая.
Голотип – как у вида.
Описание (рис. 2, е–л, с). Габитус раковины, общие очертания обеих створок, замок, мускульные отпечатки, зона сращения, краевая кайма и строение пор как у вида. Поверхность створок покрыта многочисленными некрупными ямками, на дне которых расположены ситовидные поры (рис. 2, е, з–к, с). Наиболее крупные из ямок выстраиваются в два параллельных субвертикальных ряда, оконтуривающих отпечатки аддуктора; эти ряды слабо выгибаются к заднему концу, расположены вблизи центра створки и лучше всего заметны на ювенильных экземплярах (табл. IV , фиг. 13 ). Ямки в передней половине створки группируются в неясные ряды, пересекающие ее наискось по направлению к нижней части переднего края; в задней половине – в неясные субвертикальные ряды с тенденцией выгибаться в сторону заднего края (рис. 2, з, с). Размер ямок уменьшается на периферии, где они выстраиваются в ряды параллельно свободному краю, что воспринимается как тонкая слабая ребристость по контуру створки (рис. 2, е, з, и).
Размеры в мм:
| Д | ВПК | ВЗК | Т | Д/В | |
|---|---|---|---|---|---|
| Голотип МГУ 300-Ky-1-31 (самка) | 0.38 | 0.21 | 0.19 | – | 1.8 |
| Экз. МГУ ТЭЦ-5-27 (самка) | 0.38 | 0.22 | 0.20 | – | 1.7 |
| Экз. МГУ ТЭЦ-5-26 (juv.) | 0.33 | 0.20 | 0.15 | – | 1.7 |
| Экз. МГУ 300-Kusk-131 (самка) | 0.33 | 0.20 | 0.18 | – | 1.7 |
| Экз. МГУ 300- Ky-1-32 (самка) | 0.38 | 0.22 | 0.21 | – | 1.7 |
| Экз. МГУ 300- Ky-1-37 (juv.) | 0.27 | 0.15 | 0.11 | – | 1.8 |
| Экз. МГУ 370- Belor-1-32 (juv.) | 0.26 | – | – | 0.15 | – |
| Экз. МГУ 300- Kusk-179 (самец) | 0.36 | 0.17 | 0.19 | – | 1.9 |
| Экз. МГУ 370- Belor-1-52 (самец) | 0.32 | 0.16 | 0.18 | – | 1.8 |
| Среднее (самцы) | 0.32–0.36 | 0.16–0.17 | 0.18–0.19 | – | 1.8–1.9 |
| Среднее (самки) | 0.33–0.38 | 0.20–0.22 | 0.18–0.21 | – | 1.7–1.8 |
Изменчивость у взрослых представителей проявляется в размерах ямок от более крупных (рис. 2, и) до более мелких (рис. 2, с) и отчетливости рядов. Ямки могут быть развиты по всей поверхности створки равномерно (рис. 2, е–и, с) или концентрироваться в ее центральной части, как это свойственно молодым экземплярам (табл. IV , фиг. 12, 13 ; рис. 2, к). На ювенильных раковинах число и размер ямок возрастает с каждой линькой; сначала на поверхности появляются поры, с возрастом они погружаются на дно ямок. Возрастная и половая изменчивость как у вида.
Сравнение. От подвида C. (P.) caudata nuda Tesakova, subsp. nov. отличается ямчатой скульптурой.
Распространение. Как у вида.
Материал. 190 створок ювенильных и взрослых особей самок и самцов хорошей и прекрасной сохранности из нижнего келловея, остракодовая зона P. wartae–P. kurskensis, Курская обл. (рис. 2, е–к; Tesakova, 2013, рис. 2, 5 ); девять створок самок, самцов и ювенилов из нижнего келловея, аммонитовые зоны Subpatruus (биогоризонт Saratovensis) и Koenigi, подзоны Gowerianus (биогоризонт K. g. gowerianus) и Curtilobus [биогоризонт K. (G.) indigestus] Курской обл. (рабочие коллекции Тесаковой; датирование по: Тесакова и др., 2009); 12 створок разных возрастных генераций обоих полов из нижнего келловея, зона P. wartae–P. kurskensis, Белоруссия (рис. 2, л, с; Махнач, Тесакова, 2015, рис. 2); пять створок ювенильных особей разных возрастов хорошей сохранности из нижнего келловея, аммонитовая зона Koenigi, подзона и биогоризонт K. galilaeii и среднего келловея, зона Jason, подзона Medea, Саратовская обл. (табл. IV , фиг. 12, 13 ; рис. 1).
Camptocythere (Palaeoloxoconcha) caudata nuda Tesakova, subsp. nov.
Camptocythere sp. A ssp. A: Tesakova, 2013, табл. 6 , фиг. 1.
Camptocythere sp.: Махнач, Тесакова, 2015, табл. 2 , фиг. 8 (non табл. 2 , фиг. 7, 9, 11, 12 ).
Название подвида nuda лат. – голая.
Голотип – МГУ-300-Ky-2-90, правая створка самки; Курская обл., скв. 7, гл. 138 м; нижний келловей, остракодовая зона P. wartae–P. kurskensis (рис. 2, н).
Описание (рис. 2, м–р, т, у). Габитус раковины, общие очертания обеих створок, замок, мускульные отпечатки, зона сращения, краевая кайма и строение пор как у вида. Поверхность створок воспринимается в отраженном свете лишенной скульптуры, гладкой. Однако на сканирующем микроскопе видны мелкие поры, расположенные в том же порядке, как у подвида C. (P.) caudata caudata Tesakova, subsp. nov., но в меньшем количестве (рис. 2, м–р, т).
Размеры в мм:
| Д | ВПК | ВЗК | Т | Д/В | |
|---|---|---|---|---|---|
| Голотип МГУ 300-Ky-2-90 (самка) | 0.37 | 0.21 | 0.19 | – | 1.8 |
| Экз. МГУ 300- Kusk-124 (juv.) | 0.32 | – | – | 0.16 | – |
| Экз. МГУ 300- Kusk-125 (juv.) | 0.30 | 0.17 | 0.14 | – | 1.8 |
| Экз. МГУ 300- Ку-2-57 (juv.) | 0.23 | 0.12 | 0.08 | – | 1.9 |
| Экз. МГУ 300- Ky-2-88 (самка) | 0.36 | 0.20 | 0.19 | – | 1.9 |
| Экз. МГУ 300- Kusk-90 (самка) | 0.38 | 0.21 | 0.18 | – | 1.8 |
| Экз. МГУ 300- Ky-2-89 (самец) | 0.40 | 0.20 | 0.22 | – | 1.8 |
| Среднее (самцы) | 0.40 | 0.20 | 0.22 | – | 1.8 |
| Среднее (самки) | 0.36–0.38 | 0.20–0.21 | 0.18–0.19 | – | 1.8–1.9 |
Изменчивость. Слабо варьирует выраженность пор – от мелких, редко рассеянных по поверхности (рис. 2, н, т), до разновеликих и более обильных (рис. 2, р) и даже погруженных в единичные ямки, иногда наблюдающиеся у ювенильных особей (рис. 2, п).
Сравнение. Сравнение с номинативным подвидом сделано при его описании.
Распространение. Как у вида.
Материал. 65 створок ювенильных и взрослых особей самок и самцов хорошей и прекрасной сохранности из нижнего келловея, остракодовая зона P. wartae–P. kurskensis Курской обл. (рис. 2, м–р, т, у; Tesakova, 2013, рис. 2, 5 ); пять створок самок и ювенилов из нижнего келловея, аммонитовые зоны Subpatruus (биогоризонт Saratovensis) и Koenigi, подзоны Gowerianus (биогоризонт K. g. gowerianus) и Curtilobus (биогоризонт K. (G.) indigestus) Курской обл. (рабочие коллекции Тесаковой; датирование по: Тесакова и др., 2009); семь створок разных возрастных генераций обоих полов из нижнего келловея, зона P. wartae–P. kurskensis Белоруссии (Махнач, Тесакова, 2015, рис. 2).
Camptocythere (Palaeoloxoconcha) ryazanica Tesakova, sp. nov.
Camptocythere sp. A: Тесакова и др., 2017, табл. 3 , фиг. 8, 9 ; Tesakova, Shurupova, 2018, табл. 10 , фиг. 12 .
Название вида – по первому нахождению в Рязанской обл.
Голотип – МГУ, № MC-309, правая створка самки; Рязанская обл., разрез Михайловцемент, обр. 12 (положение обр. в разрезе приводится по: Степанов и др., 2019); верхний келловей, аммонитовая зона Athleta, подзона Proniae (рис. 4, л).
Описание (рис. 4). Раковина мелкого и среднего размера, умеренно выпуклая, равномерно выполаживается к переднему и заднему концам, слабо неравностворчатая – левая створка незначительно перекрывает правую на переднеспинном и заднеспинном углах. Максимальная длина находится на середине высоты раковины, наибольшая высота – в начале переднего конца, максимальная толщина – в средней части брюшной стороны. Спинной край прямой, на правых створках немного короче, чем на левых, незначительно конвергирует к заднему концу, в передний и задний концы переходит через тупые сглаженные углы, на правых створках выраженные сильнее. Брюшной край прямой, слабо вогнут в средней части (иногда вогнутость находится ближе к переднему концу: рис. 4, к), с передним и задним концами сопрягается плавно. Передний конец высокий, плавно дугообразно закруглен, слабо скошен в верхней части на левых створках (рис. 4, а, в, г, к) и значительно сильнее на правых (рис. 4, б, д, ж, з, л), вдоль края уплощен. Задний конец ниже переднего на левых створках, а на правых почти одинаковой с ним высоты; округло-треугольной формы (на правых створках более заостренный и приподнятый кверху, чем на левых); уплощенный. Вдоль свободного края развита фестончатая кайма (рис. 4, ж, к, л). На поверхности створки имеются простые и ситовидные поры; главная пора последних смещена к краю ситовидной пластинки, а сама ситовидная пластинка погружена ниже поверхности стенки (рис. 4, и). Совокупность ситовидных пор воспринимается как ямчатая скульптура, занимающая в основном центральную часть створки, но иногда распространяющаяся на периферию. Ямки мелкие, а их размер уменьшается от центра к периферии. Уплощенные части переднего и заднего концов, а также спинная сторона створки – гладкие, с редкими простыми порами. На брюшной стороне развиты тонкие короткие продольные ребрышки (рис. 4, е). Порово-канальная зона умеренной ширины, мускульные отпечатки и замок как у подрода (рис. 4, б).
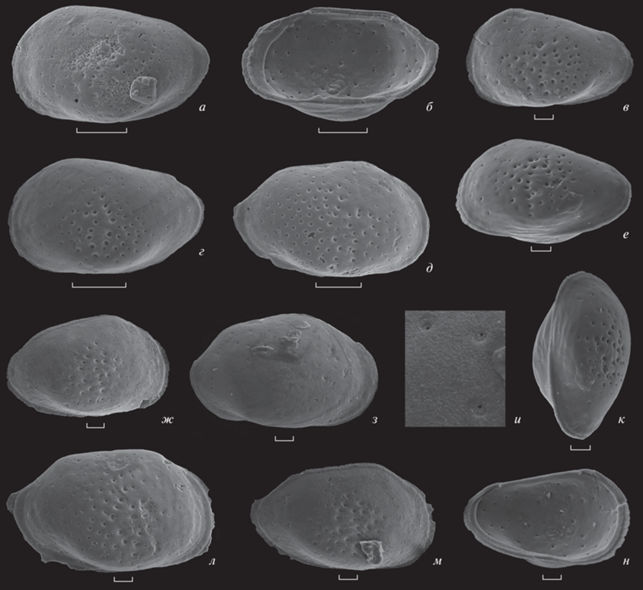
Рис. 4.
Camptocythere (Palaeoloxoconcha) ryazanica Tesakova, sp. nov.: а – экз. МГУ, № MC2-1-120, лс самки, зона Lamberti, подзона Henrici, обр. 93; б – экз. МГУ, № MC2-1-196, пс самки изнутри, обр. 74; в – экз. МГУ, № МС-2017-23, лс juv., обр. 40; г – экз. МГУ, № МС-2017-69, лс самки, обр. 71; д – экз. МГУ, № МС-2017-103, пс самки, обр. 41; е, к – экз. МГУ, № MC2-1-104, цр juv., обр. 41: е – слева, к – с брюшной стороны; ж – экз. МГУ, № MC2-149, пс juv., обр. 13; з, и – экз. МГУ, № MC2-54, пс самки, зона Coronatum, подзона и биогоризонт Grossouvrei, обр. 8: з – снаружи, и – ситовидные поры; л – голотип МГУ, № MC-309, пс самки, обр. 12; м – экз. МГУ, № MC2-147, пс juv., обр. 13; н – экз. МГУ, № MC2-1-23, лс juv. изнутри, обр. 40. Все изображенные остракоды происходят из среднего и верхнего келловея разреза Михайловцемент Рязанской области. Положение образца в разрезе для фиг. ж–и, л, м см. в: (Тесакова и др., 2017, рис. 3), для фиг. а–е, к, н – см. в: (Степанов и др., 2017, рис. 3–5). Остракоды на фиг. б, г, ж, л, м происходят из аммонитовой зоны Athleta, подзоны Proniae; на фиг. в, д, е, к, н – из зоны Athleta, подзоны Phaeinum. Длина шкалы для фиг. а, б, г, д составляет 100 мкм; для фиг. в, е–з, к–н – 30 мкм.
Размеры в мм:
| Д | ВПК | ВЗК | Т | Д/В | |
|---|---|---|---|---|---|
| Голотип МГУ MC-309 (самка) | 0.35 | 0.19 | 0.18 | – | 1.84 |
| Экз. МГУ MC2-1-120 (самка) | 0.40 | 0.23 | 0.18 | – | 1.74 |
| Экз. МГУ MC2-1-196 (самка) | 0.41 | 0.19 | 0.17 | – | 2.16 |
| Экз. МГУ МС-2017-23 (juv.) | 0.26 | 0.15 | 0.11 | – | 1.73 |
| Экз. МГУ МС-2017-69 (самка) | 0.38 | 0.20 | 0.16 | – | 1.90 |
| Экз. МГУ МС-2017-103 (самка) | 0.43 | 0.24 | 0.21 | – | 1.79 |
| Экз. МГУ MC2-1-104 (juv.) | 0.26 | 0.15 | 0.11 | 0.11 | 1.73 |
| Экз. МГУ MC2-149 (juv.) | 0.28 | 0.17 | 0.12 | – | 1.65 |
| Экз. МГУ MC2-54 (самка) | 0.32 | 0.18 | 0.15 | – | 1.78 |
| Экз. МГУ MC2-147 (juv.) | 0.30 | 0.17 | 0.14 | – | 1.76 |
| Экз. МГУ MC2-1-23 (juv.) | 0.27 | 0.16 | 0.11 | – | 1.69 |
Изменчивость. Значительно различаются линейные параметры створок взрослых самок (что определяется не только изменчивостью, но также различиями между левыми и правыми створками): длина в пределах 0.32–0.43 мм, ВПК от 0.18 до 0.24 мм, отношение Д/В 1.74–2.16 мм. Количество ситовидных пор у взрослых экземпляров варьирует от множественных (рис. 4, д) до малочисленных (рис. 4, а, г, л) и редких (рис. 4, з), а их выраженность от хорошей (рис. 4, г, д, л) до слабой (рис. 4, а, з). Половая изменчивость на изученных экземплярах не наблюдалась (в материале авторов имелись только самки). Возрастная изменчивость проявляется в неравномерном увеличении линейных размеров при каждой следующей линьке (Д возрастает быстрее, чем ВПК и ВЗК; ВЗК возрастает быстрее, чем ВПК), что отражается на контуре створок – наклон спинного и брюшного краев к заднему концу уменьшается (на ранних стадиях он максимальный) (рис. 4, в, ж, к). Также в онтогенезе происходит постепенное увеличение числа ситовидных пор и их распространение от центральной части створки (рис. 4, в, е, ж) на периферию (рис. 4, д, л).
Сравнение. Сравнение с наиболее схожими по форме раковины и скульптуре видами C. (P.) bathonica и C. (P.) caudata sp. nov. сделано при описании последнего.
Замечания. Сохранность изученных экземпляров сильно влияет на восприятие их морфологии. Так, наличие или отсутствие краевой каймы увеличивает или, соответственно, уменьшает длину створки (см. Размеры), чем можно объяснить столь сильный разброс этого параметра. От сохранности каймы на заднем конце зависит степень его заостренности и приподнятости.
При обрастании створки микритом (карбонат) под ним скрываются простые поры и мелкие ситовидные, и тогда поверхность может казаться гладкой (рис. 4, з) или скульптированной только в центральной части створки (рис. 4, а, г, л).
Распространение. Верхняя часть среднего келловея–низы нижнего оксфорда, зоны Coronatum–Mariae Рязанской обл.
Материал. Весь материал происходит из разреза Михайловцемент Рязанской обл.: 11 створок ювенильных особей и взрослых самок хорошей сохранности из среднего и верхнего келловея (зоны Coronatum и Athleta) (Тесакова и др., 2017, рис. 3; Tesakova, Shurupova, 2018, рис. 2); 143 экземпляра (целые раковины и отдельные створки) ювенильных и взрослых особей обоего пола из верхнего келловея, зона Athleta и семь створок из нижнего оксфорда, зона Mariae (рабочие коллекции Тесаковой).
НАДСЕМЕЙСТВО CYTHEROIDEA BAIRD, 1850
СЕМЕЙСТВО CYTHERURIDAE G.W. MUELLER, 1894
Род Procytherura Whatley, 1970
Procytherura ippolitovi Tesakova, sp. nov.
Табл. V , фиг. 18
Procytherura pleuraperiousios: Tesakova, 2013, табл. 5 , фиг. 14 ; Махнач, Тесакова, 2015, табл. 1 , фиг. 10 (non табл. 1 , фиг. 11, 12 ).
Procytherura tenuicostata: Махнач, Тесакова, 2015, табл. 1 , фиг. 15, 16 (non табл. 1 , фиг. 14 ).
Procytherura sp.: Глинских и др., 2022, табл. 2 , фиг. 17 .
Название вида – в честь российского палеонтолога А.П. Ипполитова.
Голотип – МГУ, № ТЭЦ-5-22, правая створка самки; Саратовская обл., разрез ТЭЦ-5; нижний келловей, зона Koenigi, подзона Galilaeii, биогоризонт K. galilaeii s.l. (табл. V , фиг. 18 ).
Описание (рис. 2, а–д). Раковина маленькая, удлиненно-овальная, умеренно равномерно выпуклая, с уплощенными передним и задним концами (плоская часть заднего конца шире, чем у переднего). Левая створка незначительно больше правой, перекрывает ее на переднеспинном и заднеспинном углах. Максимальная длина расположена на середине высоты; максимальная высота – в начале переднего конца и может совпадать с высотой в задней трети раковины; максимальная толщина – в заднебрюшной части створки (в задней половине створки). Спинной край прямой или слабо вогнут посередине за счет незначительно возвышающегося над ним продольного спинного ребра, может слабо конвергировать к заднему концу; в последний переходит плавно на правых створках и через небольшой уступ на левых; в передний конец переходит на обеих створках плавно или через небольшой уступ. Брюшной край прямой, в средней части вогнут, с обоими концами сочленяется плавно (в задний конец переходит более полого, чем в передний). Передний конец раковины высокий, равномерно дугообразно закруглен и весьма незначительно скошен сверху на правой створке. Задний конец раковины ниже переднего, округло-треугольного очертания; на правых створках скошен сверху сильнее, чем на левых. Глазной бугорок отсутствует. Отпечаток аддуктора в виде вертикального ряда из четырех удлиненных бугорков или ямок. На всей поверхности раковины развиты сравнительно крупные, отчетливые, четырехгранные (редко трех- или пятигранные) ячейки, размер и форма которых почти не меняются на периферии, а поверхность створки внутри них покрыта обильными мелкими простыми порами (рис. 2, б, в). Ячейки обладают толстыми гранями, которые, сливаясь, образуют скульптуру из продольных ребер. Примерно посередине створки (над отпечатками аддуктора) различается неглубокая пологая депрессия, нарушающая непрерывность продольных ребер. Спереди от нее два–три коротких ребра наискось пересекают передний конец, достигая его середины. В задней половине створки продольные ребра могут выгибаться к спинной стороне; они плавно огибают задний конец, на брюшной стороне располагаются вдоль брюшного края, а на переднем конце, вновь отклоняясь кверху, упираются в нижнее из косых ребер. В результате в задней половине створки возникает характерный концентрический рисунок из ребер, а в средней части створки они могут, сливаясь, формировать эллиптический контур. В передней части эллипса, приходящейся на мускульные отпечатки, регулярность ячеек и соответствующая скульптура нарушены, а в задней части эллипса различается короткое продольное ребрышко.
Порово-канальная зона узкая, с широкой бесструктурной пластинкой, развитой на обоих концах и вдоль брюшного края. Замок лофодонтный, на правых створках представлен гладким желобком, по краям которого расположены два простых зуба: удлиненный сзади, округлый спереди.
Размеры в мм:
| Д | В | |
|---|---|---|
| Голотип МГУ ТЭЦ-5-22 | 0.38 | 0.18 |
| Экз. МГУ 300-Kusk 199 | 0.33 | 0.15 |
| Экз. МГУ 370-Belor-1-25 | 0.30 | 0.14 |
| Экз. МГУ 370-Belor-1-141 (juv.) | 0.26 | 0.13 |
| Экз. МГУ 370-Belor-1-130 (juv.) | 0.24 | 0.11 |
| Экз. МГУ 370-Belor-2-14 (juv.) | 0.25 | 0.15 |
Изменчивость. Весьма незначительная и выражается в большей (табл. V , фиг. 18 ) или меньшей (рис. 2, б) толщине ребер. Возрастная изменчивость. Сначала на створках ранних стадий проявляется тончайший ретикулюм. При каждой следующей линьке преимущественное развитие получают субгоризонтальные грани ячеек, и в центральной части створки скульптура приближается к продольно ребристой, с отдельными фрагментами перемычек (рис. 2, г). По мере взросления межреберные перемычки развиваются на всей поверхности створки, и скульптура сначала становится сетчатой (рис. 2, в, д), а впоследствии ячеистой.
Сравнение. Из других келловейских процитерур с сетчатой скульптурой новый вид более всего похож на P. didictyon rossica Tesakova из нижнего келловея Саратовской обл. и Гомельского р-на Белоруссии (Тесакова, Сельцер, 2013, с. 61, рис. 6 , фиг. 7–11 ; Махнач, Тесакова, 2015, табл. 1 , фиг. 6–9, 13 ), от которого отличается толстыми гранями ячеек, сливающимися в ребра. От ребристого вида P. pleuraperiousios Whatley, Ballent et Armitage из верхнего келловея Дорсета, Англия (Whatley et al., 2001, с. 146, табл. 2 , фиг. 15–18 ; табл. 3 , фиг. 1, 2), скульптуре которого свойственна регулярность продольных ребер, отличается концентрическим расположением ребер на заднем конце и косыми ребрами на переднем конце.
Распространение. Нижний келловей, остракодовая зона Pr. wartae–P. kurskensis; Белоруссия и Европейская Россия (Курская и Саратовская обл.).
Материал. 23 экз.: один экз. (голотип) прекрасной сохранности из нижнего келловея, зона Koenigi, разрез ТЭЦ-5, Саратовская обл.; 20 экз. хорошей и удовлетворительной сохранности взрослых и ювенильных особей из нижнего келловея, остракодовая зона Pr. wartae–P. kurskensis, разрез скв. 792, Гомельский р-н, Юго-Восточная Белоруссия (Махнач, Тесакова, 2015, рис. 2); два экз. прекрасной сохранности из нижнего келловея, зона Pr. wartae–P. kurskensis, разрез скв. 7, Курская обл. (Tesakova, 2013).
БИОСТРАТИГРАФИЯ
Неравномерное распределение остракод по разрезу ТЭЦ-5 позволило выделить четыре различных комплекса (рис. 1).
Комплекс с A. milanovskyi–P. cinicinnusa состоит только из видов-индексов и определен в терминальной части нижнекелловейской зоны Subpatruus, в биогоризонте Ch. saratovensis (сл. 1, обр. 4 и 5). Вид Procytheridea cinicinnusa (Mand. in Lyub.) представлен единичными ядрами раковин (табл. IV , фиг. 1–6 ), как и сильно деформированная раковина Acantocythere milanovskyi (Lyub.) (табл. V , фиг. 13 ). В ассоциации с ними в отложениях зоны Subpatruus в разрезах Починки (Нижегородская обл.) (Тесакова и др., 2020) и Бартоломеевка (Саратовская обл.) (Тесакова, Сельцер, 2013) наблюдалось присутствие вида Camptocythere (A.) starcevae Tes. in Tesakova et Seltser, 2013, который маркирует контакт Печорского и Среднерусского морей и относительно холодноводные условия (Тесакова, 2022). По совместному распространению индексов предложено установить одноименную зону, отвечающую аммонитовым биогоризонтам Subpatruus–Saratovensis (Тесакова и др., 2020) и отнести к ней соответствующий интервал разреза ТЭЦ-5.
Комплекс с Camptocythere (P.) caudata caudata выделяется в разрезе ТЭЦ-5 в слоях 6а–7а (обр. 11 и 15), отвечает верхней части зоны Koenigi, подзоне и биогоризонту Galilaeii и нижней части зоны и подзоны Calloviense, биогоризонтам Calloviense и Kiselevi, и имеет совершенно иной состав: Procytherura didictyon rossica (табл. V , фиг. 16, 17 ), P. ippolitovi sp. nov. (табл. V , фиг. 18 ), C. (P.) caudata caudata sp. et subsp. nov. (табл. IV , фиг. 12, 13 ), Glabellacythere nuda Wienholz (табл. IV , фиг. 17 ), Lophocythere sp. B (табл. V , фиг. 4–8 ) и Pleurocythere kurskensis Tes. (табл. V , фиг. 11, 12 ). Все перечисленные виды характерны для остракодовой зоны P. wartae–P. kurskensis.
В комплексе с P. wartae–P. kurskensis состав обновлен полностью (за исключением транзитного P. kurskensis) и состоит из: Praeschuleridea wartae Błaszyk (табл. IV , фиг. 7–11 ), Pseudoperissocytheridea anastomosa Whatley, Ballent et Armitage (табл. V , фиг. 14, 15 ), Lophocythere karpinskyi (Mand. in Lyub.) (табл. V , фиг. 9, 10 ), Neurocythere plena (Triebel) (табл. V , фиг. 1) и N. franconica (Triebel) (табл. V , фиг. 2, 3). Комплекс выделен в слое 8с и нижней части слоя 9 (обр. 21 и 23) и отвечает верхней части зоны Calloviense и подзоны Enodatum (биогоризонт C. enodatum enodatum) и низам среднекелловейской зоны Jason, подзоне Medea (биогоризонт K. medea magnum). Перечисленные виды также являются характерными для зоны P. wartae–P. kurskensis.
Комплекс с Camptocythere (C.) quinta выделяется в слое 9 (обр. 24), в низах зоны Jason в подзоне Medea, но выше биогоризонта K. medea magnum. В нем зафиксировано только четыре вида: C. (P.) caudata caudata sp. et subsp. nov., L. sp. B, P. kurskensis и C. (C.) quinta sp. nov. Первые три входят в состав второго и третьего комплексов и являются фоновыми таксонами зоны Ps. wartae–P. kurskensis.
Между нижним комплексом и тремя верхними существует очень важное отличие – принадлежность входящих в них камптоцитер к разным подродам. Появление на ВЕП анабароцитер свидетельствует о проникновении их с севера. Вид C. (A.) starcevae из первого комплекса вселился в Среднерусское море из Печорского моря, Баренцевоморского шельфа и севера Сибири (Тесакова и др., 2020; Тесакова, 2022). С другой стороны, палеолоксоконхи C. (P.) caudata caudata sp. et subsp. nov. из второго и четвертого комплексов могли прийти только по западному миграционному коридору. Вид C. (C.) quinta sp. nov. мог развиваться в Среднерусском море или мигрировать из Западной Европы, но не мог попасть на ВЕП по северному коридору, так как среди хорошо известных камптоцитер циркумполярной области Северного полушария нет ни одного похожего таксона.
ОПИСАНИЕ СТРАТОНОВ
Зона Acantocythere (P.) milanovskyi–Procytheridea cinicinnusa Tesakova, 2020 (зона совместного распространения)
слои с Acantocythere: Тесакова, Сельцер, 2011, с. 218, рис. 2, nom. nud.
комплекс с Acantocythere milanovskyi–P. cinicinnusa в слоях с Acantocythere: Тесакова, Сельцер, 2011, с. 218, рис. 2, nom. nud.
слои с Palaeocytheridea pavlovi: Унифицированная…, 2012, лист 2, с. 17, part.
слои с Acantocythere (P.) milanovskyi: Тесакова, Сельцер, 2013, с. 58, рис. 2.
комплекс с P. cinicinnusa в слоях с Acantocythere (P.) milanovskyi: Тесакова, Сельцер, 2013, с. 58, рис. 2.
подзона A. milanovskyi–P. cinicinnusa: Тесакова, 2014, с. 20, рис. 4; Устинова, Тесакова, 2015, с. 74, рис. 2, 3.
слои с A (P.) milanovskyi: Тесакова, 2014, с. 20, рис. 4; Устинова, Тесакова, 2015, с. 74, рис. 2, 3.
зона A. milanovskyi–P. cinicinnusa: Тесакова и др., 2020, с. 223, рис. 3; Глинских и др., 2022, рис. 3, 4.
Состав комплекса: кроме индексов P. cinicinnusa (Mand. in Lyub.) и A. (P.) milanovskyi (Lyub.), характерно присутствие: Camptocythere (A.) starcevae Tes. in Tes. et Selts., C. (C.) scrobiculataformis Nikitenko, Palaeocytheridea (P.) pavlovi (Lyubimova), Fuhrbergiella (P.) archangelskyi (Mand. in Lyub.), Galliaecytheridea prodromos Whatley, Ballent et Arm., Fastigatocythere interrupta directa Wienholz, Lophocythere karpinskyi, Lophocythere sp., Neurocythere cruciata (Trieb.), Patellacythere sp., Macrocypris aequabilis Oertli, Parariscus octoporalis Błaszyk, Procytherura sokolovi (Lyub.), P. didictyon rossica Tes. in Tes. et Selt., Aaleniella sp., Praeschuleridea sp. и др. (изображения некоторых характерных видов приведены в: Тесакова, Сельцер, 2013; наиболее полный список таксонов приведен в: Тесакова и др., 2020, рис. 3).
Лектостратотип: разрез Починки Нижегородской обл. (Тесакова и др., 2020, рис. 3). Темно-серые глины елатьминской свиты (Унифицированная…, 2012, лист 9), слой 5 (обр. 1–13). Мощность 5.7 м. Комплекс выделяется по совместной встречаемости видов-индексов.
Границы: нижняя – по первому появлению A. (P.) milanovskyi; верхняя – по исчезновению обоих индексов. Фактически, объем зоны совпадает с интервалом распространения первого индекса.
Стратиграфическое положение: отвечает аммонитовой зоне Subpatruus, биогоризонтам Subpatruus, Uzhovkensis и Saratovensis (Тесакова и др., 2020); средней части фораминиферовой зоны Haplophragmoides infracalloviensis–Guttulina tatarensis (Глинских и др., 2022); верхней части остракодовой филозоны Palaeocythe-ridea (Р.) pavlovi (Тесакова, 2014).
Замечания. (1) Хотя стратон используется авторами в работах, начиная с 2011 г., и характерный комплекс был отмечен во многих разрезах на территории ВЕП, ни в одной из предшествующих работ не был формально обозначен стратотип. Пользуясь этим обстоятельством, в настоящей работе мы считаем необходимым зафиксировать лектостратотип зоны. В качестве такового нами выбран наиболее представительный из известных на сегодняшний день и вскрывающий почти полную последовательность зоны Subpatruus – разрез Починки, несмотря на то, что первоначально стратон был выделен по материалам, собранным преимущественно в Саратовской обл. (в разрезе Бартоломеевка).
(2) Для зоны чрезвычайно характерно массовое развитие обоих видов-индексов, имеющих узкое стратиграфическое распространение в интервале зоны Subpatruus, а порядок их появления в других, датированных по аммонитам, разрезах – Бартоломеевка (Саратовская обл.; Тесакова, Сельцер, 2013) и Починки (Нижегородская обл.; Тесакова и др., 2020) – одинаков: P. cinicinnusa появляется в самом низу зоны, в биогоризонте Surensis, в то время как A. milanovskyi позже, в биогоризонте Subpatruus.
(3) Три наиболее типичных вида зоны – A. (P.) milanovskyi, P. cinicinnusa и P. (P.) pavlovi – выявлены П.С. Любимовой (1955), Т.Н. Хабаровой (1961) и Тесаковой (Тесакова, Сельцер, 2013; Глинских и др., 2022) в нижнем келловее Саратовской, Нижегородской (Тесакова и др., 2020) областей и в Татарстане (Любимова, 1955, с. 125, 126). Зональный вид A. (P.) milanovskyi отмечен как типичная форма нижнего келловея Украины (Пяткова, Пермякова, 1978), Курской обл. (Преображенская, 1964) и Самарской Луки (Любимова, 1955). P. cinicinnusa указан из нижнего келловея Украины и Самарской Луки, а P. (P.) pavlovi – из нижнего келловея Самарской Луки и Татарстана (Любимова, 1955; Пяткова, Пермякова, 1978).
(4) Присутствие в зональном комплексе C. (A.) starcevae и C. (C.) scrobiculataformis – камптоцитер, связанных своим географическим происхождением с палеобассейнами севера России – маркирует, вероятно, некоторое снижение температуры вод и устойчивую связь Среднерусского и Печорского палеоморей (Тесакова и др., 2020; Тесакова, 2022).
Распространение: Украина, Курская обл., Нижнее и Среднее Поволжье.
Зона Praeschuleridea wartae–Pleurocythere kurskensis Tesakova, 2003 (зона совместного распространения)
слои с Praeschuleridea wartae–Pleurocythere regularis: Tesakova, 2003, с. 137, табл. 4 .
слои с Praeschuleridea wartae–Pleurocythere kurskensis: Тесакова и др., 2009, с. 30; Тесакова, Сельцер, 2011, с. 281; Глинских и др., 2022, c. 66, рис. 3, 4.
подзона Praeschuleridea wartae–Pleurocythere kurskensis: Tesakova, 2013, с. 1216; Тесакова, 2014, с. 20, рис. 4; Махнач, Тесакова, 2015, с. 91, рис. 2.
Состав комплекса: кроме видов-индексов, известно большое число других: Cytherella fullonica Jones et Sherborn, Polycope sububiquita What., Paracypris sp., Macrocypris aequabilis, Bythoceratina scrobiculata (Triebel et Bart.), B. ssp., Patellacythere aliena (Lyub.), Camptocythere (P.) caudata sp. nov., Glabellacythere nuda, G. dolabra (Jones et Scherb.), Mandelstamia aff. ventrocornuta Lyub., Camptocythere (P.) caudata sp. nov., Palaeocytheridea (M.) parabakirovi Malz, Lophocythere scabra Triebel, L. karpinskyi, Fuhrbergiella archangelskyi, Fastigatocythere interrupta interrupta (Triebel), F. interrupta directa, Galliaecytheridea prodromos, Neurocythere franconica, N. flexicosta (Triebel), Procytherura tenuicostata Whatley, P. didiction What., Ball. et Arm., P. pleuraperiousios, P. sokolovi, P. ippolitovi sp. nov., Eucytherura acostata (Tes.), E. paula (Lyub.), E. ssp., Tethysia bathonica Sheppard, Pedicythere anterodentina What., Metacytheropteron aff. sutherlandensis What., Nodophthalmocythere sp., Parariscus octoporalis, Rubracea artis Lyub. и др. (Тесакова и др., 2009; Tesakova, 2013; Махнач, Тесакова, 2015).
Наиболее характерными именно для этой зоны являются: Pl. kurskensis, P. wartae, C. (P.) caudata sp. nov., G. nuda, G. prodromos, N. flexicosta, N. franconica, M. aff. sutherlandensis, Nodophthalmocythere sp. и P. ippolitovi sp. nov.
Стратотип: разрез Михайловского рудника КМА, темно-серые глины фатежской свиты, нижняя подсвита, сл. 5–9 (Tesakova, 2003, рис. 15 ). Мощность ≈ 36 м.
Границы: нижняя граница выделяется по первому появлению Pl. kurskensis; верхняя – по исчезновению обоих индексов.
Замечания. (1) Зона Pr. wartae–Pl. kurskensis впервые была установлена в Курской обл. как слои с остракодами только в нижнекелловейской зоне Koenigi, хотя характерный комплекс, включая виды-индексы, был прослежен в стратотипе также и в среднем келловее (зона Jason) (Tesakova, 2003). Впоследствии ее интервал был расширен за счет верхней части зоны Subpatruus (биогоризонт Saratovensis) (Тесакова и др., 2009). Позже она была идентифицирована в зонах Koenigi, Calloviense и Jason в Саратовской обл. (Глинских и др., 2022).
(2) Характерный комплекс был установлен В.Н. Преображенской в Центрально-Черноземной (1966а, б) и Курской (1964) областях, а также Любимовой (1956) и М.Н. Пермяковой (Пяткова, Пермякова, 1978) в Белоруссии и на Украине.
Стратиграфическое положение: нижний келловей (зона Subpatruus, биогоризонт Saratovensis)–средний келловей (зона Jason) (Tesakova, 2003; Тесакова и др., 2009; Глинских и др., 2022); верхняя часть фораминиферовой зоны H. infracalloviensis‒G. tatarensis и нижняя часть зоны L. pseudocrassa‒L. cultratiformis (Глинских и др., 2022); средняя часть остракодовой филозоны Palaeocytheridea (M.) parabakirovi (Тесакова, 2014).
Распространение: Украина, Белоруссия, Центрально-Черноземная область, Курская обл. и Среднее Поволжье.
Выбор камптоцитер в качестве индексов для описанных ниже филозон не случаен – для юры севера России и Северо-Западной Европы были разработаны филогенетические шкалы по эволюции подродов C. (Camptocythere) и C. (Anabarocythere). Экземпляры номинативного подрода были встречены в разрезе ТЭЦ-5 только в среднем келловее в зоне Jason и описаны как новый вид. К сожалению, скудость материала и стратиграфическая обособленность наиболее позднего вида C. (C.) quinta sp. nov. не позволяет пока выделить по этой филолинии биостратоны (в том числе в ранге слоев). А эволюция другого подрода, C. (Palaeoloxoconcha), проходившая только в палеобассейнах Северо-Западной и Восточной Европы, дала основания для выделения новых филозон со смыкаемыми границами: C. (P.) bathonica (J2b2-bt) → C. (P.) caudata sp. nov. (J2cl1-21) → C. (P.) ryazanica sp. nov. (J2cl22-J3ox1). Филозона C. (P.) bathonica устанавливается в Северо-Западной Европе в отложениях верхнего байоса–бата и охватывает терминальную часть верхнебайосской аммонитовой зоны Parkinsoni и все зоны бата Wuerttembergica–Aspidoides Северной Германии (Dreyer, 1967).
Зона Camptocythere (Palaeoloxoconcha) caudata Tesakova, nov. (филозона)
Вид-индекс. C. (P.) caudata Tesakova, sp. nov.
Состав комплекса: такой же, как у зоны Pr. wartae–Pl. kurskensis.
Стратотип: разрез Михайловского рудника КМА, темно-серые глины фатежской свиты, нижняя подсвита, сл. 5–9 (Tesakova, 2003, рис. 15 ). Мощность ≈36 м.
Границы: нижняя – по появлению вида индекса, а верхняя по появлению преемственного вида.
Стратиграфическое положение: от нижнего келловея (с верхней части зоны Subpatruus, биогоризонт Saratovensis) до нижней части среднего келловея (зона Jason).
Распространение. Прослеживается на ВЕП в Белоруссии, Курской и Саратовской обл.
Зона Camptocythere (Palaeoloxoconcha) ryazanica Tesakova, nov. (филозона)
Вид-индекс. C. (P.) ryazanica Tesakova, sp. nov.
Состав комплекса. Кроме зонального вида-индекса, известно большое число видов: Palaeocytheridea (M.) parabakirovi, Cytherella perennis Błasz., C. oblonga Permjakova, Lophocythere acrolophos Wh., Bal. et Arm., L. sp. A, Fuhrbergiella archangelskyi, Fastigatocythere sp. A., Galliaecytheridea imprimus What., Ball. et Arm., Schuleridea translucida (Lyub.), Patellacythere calloveica (Mand. in Lyub.), Neurocythere cruciata cruciata (Trieb.), N. flexicosta labyrinthos What., Ball. et Arm., N. flexicosta ssp. A, Pseudoperissocytheridea ex gr. anastomosa What., Ball. et Arm., Procytherura baculumbajula (Mand. in Lyub.), Eucytherura acostata, E. grandipyga (Tes.), E. paula, Pedicythere anterodentina, Exophthalmocythere pilosa Tes. и др. (Тесакова и др., 2017; Tesakova, Shurupova, 2018).
Наиболее характерными являются: C. (P.) ryazanica sp. nov., L. acrolophos, L. sp. A, Fastigatocythere sp. A., G. imprimus, S. translucida, N. flexicosta labyrinthos и P. ex gr. anastomosa.
Стратотип: разрез Михайловцемент (Рязанская обл.), темно-серые глины чулковской свиты (Унифицированная …, 2012, лист 9), сл. 4 – нижняя часть сл. 6 (по: Степанов и др., 2019, рис. 5 ). Мощность ≈ 11 м.
Границы: нижняя – по появлению вида-индекса; верхняя – по его исчезновению.
Стратиграфическое положение: отвечает верхней части среднего келловея (зона Coronatum, подзона Grossouvrei) – нижнему оксфорду (зона Mariae, подзона Scarburgense); верхней части фораминиферовой зоны L. pseudocrassa–L. cultratiformis, зоне L. tumida–E. elschankaensis и нижней части зоны O. sagittum–E. volgensis; а также верхней части остракодовой филозоны P. (M.) parabakirovi.
Распространение. Прослеживается на ВЕП в Рязанской обл.
ЗАКЛЮЧЕНИЕ
Из нижнего и среднего келловея разреза ТЭЦ-5 (Саратовская обл.) и среднего келловея–нижнего оксфорда разреза Михайловцемент (Рязанская обл.) описано четыре новых вида остракод: Camptocythere (Camptocythere) quinta Tesakova, sp. nov., C. (Palaeoloxoconcha) caudata Tesakova, sp. nov. [с подвидами C. (P.) caudata caudata Tesakova, subsp. nov. и C. (P.) caudata nuda Tesakova, subsp. nov.], C. (P.) ryazanica Tesakova, sp. nov. и Procytherura ippolitovi Tesakova, sp. nov. Проведена ревизия рода Camptocythere Triebel, изменен его систематический состав, расширен стратиграфический объем и составлено обновленное описание; также переописаны подроды C. (Camptocythere) и C. (Palaeoloxoconcha).
Составлено описание остракодовой зоны совместного распространения A. milanovskyi–P. cinicinnusa, которая отвечает аммонитовой зоне Subpatruus, биогоризонтам Subpatruus, Uzhovkaensis и Saratovensis; средней части фораминиферовой зоны H. infracalloviensis–G. tatarensis; верхней части остракодовой филозоны P. (Р.) pavlovi; и прослеживается на Украине, в Курской обл. и по всему Поволжью.
Вторая зона совместного распространения Pr. wartae–Pl. kurskensis выделена для нижнего келловея (аммонитовая зона Subpatruus, биогоризонт Saratovensis) – среднего келловея (зона Jason, подзона Medea) Украины, Белоруссии, Центрально-Черноземной области, Курской обл. и Среднего Поволжья. Также отвечает верхней части фораминиферовой зоны H. infracalloviensis‒G. tatarensis и нижней части зоны L. pseudocrassa‒L. cultratiformis; средней части остракодовой филозоны P. (M.) parabakirovi и верхней части новой филозоны Camptocythere (P.) caudata.
По эволюции камптоцитер подрода C. (Palaeoloxoconcha) в средней юре Северо-Западной и Восточной Европы предложены новые филозоны. Зона C. (P.) bathonica устанавливается в Северо-Западной Европе в отложениях верхнего байоса–бата (Dreyer, 1967). Зона C. (P.) caudata охватывает нижний келловей (зона Subpatruus, биогоризонт Saratovensis)–нижнюю часть среднего келловея (зона Jason) и прослеживается на ВЕП повсеместно. Зона C. (P.) ryazanica выделена для верхней части среднего келловея (зона Coronatum)–нижнего оксфорда (зона Mariae) Рязанской обл.
* * *
Авторы признательны Я.А. Шуруповой (МГУ) за отмывку образцов, отобранных из разреза ТЭЦ-5 Сельцером и Тесаковой в 2011 г., и Р.А. Ракитову (ПИН) за неизменную помощь в работе на СЭМ. Также мы глубоко благодарны нашим рецензентам Л.М. Мельниковой (ПИН) и А.П. Ипполитову (ГИН) за конструктивные замечания, способствующие улучшению этой статьи.
Работа выполнена в рамках тем госзадания №№ 0135-2019-0062 (ГИН РАН) и АААА-А16-116033010096-8 (МГУ).
Список литературы
Атлас руководящих ископаемых мезозоя баренцевского шельфа и его островного обрамления. Кн. 3. Юрская и меловая системы: отчет о НИР тема 150 / ВНИИОкеангеология; рук. Н.И. Шульгина. СПб.: ВНИИОкеангеология, 1995. 175 с. № ГР 467622 (Росгеолфонд).
Басов В.А., Никитенко Б.Л., Куприянова Н.В. Стратиграфия и микрофауна (фораминиферы и остракоды) нижней и средней юры баренцевоморского шельфа // Геол. и геофизика. 2009. Т. 50. № 5. С. 525–549.
Глинских Л.А., Тесакова Е.М., Сельцер В.Б. О биостратиграфии нижнего–среднего келловея Саратовского Поволжья по микрофауне // Стратигр. Геол. корреляция. 2022. Т. 30. № 2. С. 60–70.
Кашеварова Н.П., Мандельштам М.И., Шнейдер Г.Ф. Надсемейство Cytheracea // Основы палеонтологии. Членистоногие – трилобитообразные и ракообразные. М.: Изд-во АН СССР, 1960. С. 365–411.
Колпенская Н.Н. Новые виды остракод из кимериджских и волжских отложений в бассейне р. Печоры // Палеонтол. журн. 1993. № 4. С. 80–86.
Колпенская Н.Н. Юра // Практическое руководство по микрофауне. Т. 7. Остракоды мезозоя. СПб.: Изд-во ВСЕГЕИ, 1999. С. 125–129.
Лев О.М. Нижнеюрские остракоды Нордвикского и Лено-Оленекского районов // Сборник статей по палеонтологии и биостратиграфии. Л., 1958. С. 23–49 (Тр. НИИГА. Вып. 12).
Лев О.М. Микрофауна нижне- и среднеюрских отложений Лено-Оленекского района // Сборник статей по палеонтологии и биостратиграфии. Л., 1961. С. 35–71 (Тр. НИИГА. Вып. 26).
Лев О.М. Комплексы остракод из юрских отложений Анабарского района // Палеонтология и биостратиграфия. Л., 1966. С. 25–41 (Уч. записки НИИГА. Вып. 15).
Лев О.М., Кравец B.C. Юрские остракоды Тимано-Печорского региона и их стратиграфическое значение // Стратиграфия триасовых и юрских отложений нефтегазоносных бассейнов СССР. Л.: ВНИГРИ, 1982. С. 65–78.
Любимова П.С. Остракоды мезозойских отложений Среднего Поволжья и Общего Сырта // Тр. ВНИГРИ. Нов. сер. 1955. Вып. 84. С. 3–190.
Любимова П.С. Триасовые и юрские остракоды восточных районов Украины // Тр. ВНИГРИ. Нов. сер. 1956. Вып. 98. С. 533–583.
Махнач В.В., Тесакова Е.М. Палеогеографические реконструкции природной среды Юго-Восточной Белоруссии в батском–оксфордском веках // Вестн. Моск. ун-та. Сер. 4. Геол. 2015. № 2. С. 84–93.
Международный кодекс зоологической номенклатуры. 4 изд. М.: Т-во научн. изданий КМК, 2004. 223 с.
Митта В.В., Сельцер В.Б. О среднеюрских гетероморфных аммонитах России // Современные проблемы изучения головоногих моллюсков. Матер. всеросс. совещ. М.: ПИН РАН, 2006. С. 25–28.
Никитенко Б.Л. Ранне- и среднеюрские остракоды севера Сибири: основные закономерности эволюции и зональная шкала // Стратигр. Геол. корреляция. 1994. Т. 2. № 4. С. 38–55.
Никитенко Б.Л. Стратиграфия, палеобиогеография и биофации юры Сибири по микрофауне (фораминиферы и остракоды). Новосибирск: Параллель, 2009. 680 с.
Попов Е.В., Сельцер В.Б., Волков А.В. О находке зубов эласмобранхий (Chondrichthyes: Elasmobranchii) в нижнем келловее Саратова // Тр. НИИГео СГУ им. Н.Г. Чернышевского. Нов сер. 2004. Т. 16. С. 133–154.
Практическое руководство по микрофауне СССР. Т. 3. Остракоды кайнозоя / Ред. Б.С. Соколов. Л.: Недра, 1989. 233 с.
Практическое руководство по микрофауне. Т. 7. Остракоды мезозоя / Ред. Б.С. Соколов. СПб.: ВСЕГЕИ, 1999. 244 с.
Преображенская В.Н. Значение остракод для стратиграфии юрских отложений территории КМА // Геология и полезные ископаемые Центрально-Черноземных областей. Тр. межобластного геол. совещ. Воронеж: Изд-во Воронеж. гос. ун-та, 1964. С. 179–183.
Преображенская В.Н. Условия существования и захоронения фораминифер и остракод в юрское и нижнемеловое время на территории ЦЧО // Тр. Третьего совещ. по проблемам изучения Воронежской антеклизы (7–11 апреля 1964 г.). Воронеж: Изд-во Воронеж. гос. ун-та, 1966а. С. 261–272.
Преображенская В.Н. Стратиграфия отложений юры и низов нижнего мела территории ЦЧО. Воронеж: Изд‑во Воронеж. гос. ун-та, 1966б. 281 с.
Пяткова Д.М., Пермякова М.Н. Фораминиферы и остракоды юры Украины. Киев: Наук. думка, 1978. 288 с.
Степанов И.А., Казанский А.Ю., Киселев Д.Н. и др. Возможности петромагнитных методов при палеоэкологических реконструкциях на примере юрских отложений (келловей–нижний оксфорд) разреза Михайловцемент (Рязанская область) // Вестн. Моск. ун-та. Сер. 4. Геол. 2019. № 2. С. 30–38.
Тесакова Е.М. Остракоды рода Palaeocytheridea Mandelstam, 1947 в средней и верхней юре Европы. 1. Развитие представлений об объеме рода и результаты его ревизии // Палеонтол. журн. 2013а. № 3. С. 25–38.
Тесакова Е.М. Остракоды рода Palaeocytheridea Mandelstam, 1947 в средней и верхней юре Европы. 2. Описание таксонов // Палеонтол. журн. 2013б. № 5. С. 28–36.
Тесакова Е.М. Юрские остракоды Русской плиты: стратиграфическое значение, палеоэкология и палеогеография. Автореф. дис. … докт. геол.-мин. наук. М.: МГУ, 2014. 48 с.
Тесакова Е.М. Юрские ракушковые раки (Ostracoda) европейской части бывшего СССР: трудности и успехи изучения // Успехи совр. биол. 2020. Т. 140. № 1. С. 73–86. https://doi.org/10.31857/S004213242001007X
Тесакова Е.М. Позднебайосские и раннебатские остракоды Русской плиты. Часть III. Роды Camptocythere Triebel и Procytheridea Peterson // Палеонтол. журн. 2022. № 4. С. 48–58.
Тесакова Е.М., Глинских Л.А., Федяевский А.Г. и др. Микрофоссилии из нижнекелловейской аммонитовой зоны Subpatruus Нижегородской области // Юрская система России: проблемы стратиграфии и палеогеографии. Матер. VIII Всеросс. совещ. с международным участием. Онлайн-конф., 7–10 сентября 2020 г. / Ред. В.А. Захаров, М.А. Рогов, Е.В. Щепетова, А.П. Ипполитов. Сыктывкар: ИГ Коми НЦ УрО РАН, 2020. С. 222–230.
Тесакова Е.М., Сельцер В.Б. Стратиграфическое значение остракод нижнего келловея Центральной России // Научн. матер. 4-го Всеросс. совещ. “Юрская система России: проблемы стратиграфии и палеогеографии” / Ред. В.А. Захаров. СПб.: ООО “Изд-во ЛЕМА”, 2011. С. 218–220.
Тесакова Е.М., Сельцер В.Б. Остракоды и аммониты нижнего келловея разреза Бартоломеевка (Саратовская обл.) // Бюлл. Моск. об-ва испыт. прир. Отд. геол. 2013. Т. 88. Вып. 2. С. 50–68.
Тесакова Е.М., Стреж А.С., Гуляев Д.Б. Новые остракоды из нижнего келловея Курской обл. // Палеонтол. журн. 2009. № 3. С. 25–36.
Тесакова Е.М., Шурупова Я.А., Устинова М.А. Стратиграфия келловея и нижнего оксфорда разреза Михайловцемент (Рязанская обл.) по микрофауне и наннопланктону // Тр. Геол. ин-та РАН. 2017. Вып. 615. С. 264–300.
Унифицированная региональная стратиграфическая схема юрских отложений Восточно-Европейской платформы. Объяснительная записка. М.: ПИН РАН–ФГУП “ВНИГНИ”, 2012. 64 с.
Устинова М.А., Тесакова Е.М. Сравнительный анализ распространения раннекелловейских фораминифер и остракод из разреза Бартоломеевка (Саратовская область) // Вестн. Моск. ун-та. Сер. 4. Геол. 2015. № 3. С. 73–84.
Хабарова Т.Н. О микрофауне юрских отложений Саратовской области // Тр. ВНИГРИ. 1961. Вып. 29. Т. 3. С. 177–184.
Шарапова Е.Г. Остракоды батского яруса полуострова Нордвик (Юрюнг-Тумус) // Тр. НГРИ. Нов. сер. 1940. Вып. 10. С. 120–130.
Bate R.H., Coleman B.E. Upper Lias ostracoda from Rutland and Huntingdonshire // Bull. Geol. Surv. G.B. 1975. № 55. P. 1–42.
Dietze V., Franz M., Kutz M., Waltschew A. Stratigraphy of the Middle Jurassic Sengenthal Formation of Polsingen-Ursheim (Nördlinger Ries, Bavaria, Southern Germany) // Palaeodiversity. 2017. V. 10. P. 49–95.
Dietze V., Wannenmacher N., Franz M., Weis R. Neue Erkenntnisse über die Wedelsandstein-Formation der Zollernalb (Schwäbische Alb, SW-Deutschland) // Zitteliana. 2018. V. 93. S. 47–80.
Dreyer E. Einige neue Ostracoden aus dem Ober-Bajoce und Bath der Deutschen Demokratischen Republik // Freiberger Forschungshefte, Pal. Reihe C. 1967. № 213. S. 53–61.
Gulyaev D.B., Ippolitov A.P. Lower Callovian of Kanev Dislocations, Cherkasy Oblast, Ukraine: Ammonites and stratigraphy // Stratigr. Geol. Correlation. 2021. V. 29. № 7. P. 767–847.
Howe H.V. Family Cytheridae Baird, 1850 // Treatise on Invertebrate Paleontology. Pt Q. Arthropoda. Lawrence: Geol. Soc. Amer.; Univ. Kansas Press, 1961. P. 254–259.
Howe H.V., van den Bold W.A., Reyment R.A. Family Cytherideidae Sars, 1925 // Treatise on Invertebrate Paleontology. Pt Q. Arthropoda. Lawrence: Geol. Soc. Amer.; Univ. Kansas Press, 1961. P. 272–290.
Franz M., Ebert M., Stulpinaite R. Aalenian–Lower Bajocian (Middle Jurassic) ostracods from the Geisingen clay pit (SW Germany) // Palaeodiversity. 2018. V. 11. P. 59–105.
Malz H. Zur Kenntnis einiger Ostracoden-Arten der Gattungen Kinkelinella und Praeschuleridea. // Senckenb. leth. 1966. V. 47. № 4. S. 385–404.
Nikitenko B.L., Mickey M.B. Foraminifera and ostracodes across the Pliensbachian–Toarcian boundary in the Arctic Realm // Geol. Soc. London. Spec. Publ. 2004. V. 230. P. 137–173.
Plumhoff F. Ostracoden des Oberaalenium und tiefen Unterbajocium (Jura) des Gifhorner Troges, Nordwestdeutschland // Abh. Senckenb. Naturf. Ges. 1963. Bd 503. S. 1–100.
Riegraf W. Microfauna, Biostratigraphie und Fazies im Unteren Toarcium Sudwestdeutschlands und Vergleiche mit benachbarten Gebieten // Tubinger Micropaläontol. 1985. Bd 3. S. 1–232.
Shurupova Y.A., Tesakova E.M. Detailed biostratigraphic scales as based on the palaeobiogenetical approach (an example of the Upper Bajocian-Lower Bathonian ostracod scale of the Russian Platform) // Volumina Jurassica. 2017. V. 15. P. 1–17.
Tesakova E.M. Callovian and Oxfordian ostracodes from the central region of the Russian Plate // Paleontol. J. 2003. V. 37. Suppl. 2. P. 107–227.
Tesakova E.M. Ostracode-based reconstruction of paleodepths in the Early Callovian of the Kursk Region, Central Russia // Paleontol. J. 2013. V. 47. № 10. P. 1214–1230.
Tesakova E.M. Biostratigraphie du Jurassique moyen des sondages Benken et Schlattingen-1 (Nord de la Suisse) ètablie sur la base de la répartition des ostracodes // NAGRA Arbeitsbericht NAB. 2017. V. 17–41. 77 p.
Tesakova E.M., Shurupova Ya.A. Ostracod analysis of Callovian and Lower Oxfordian deposits of the Mikhailovtsement Section (Ryazan Region): methods and results // Paleontol. J. 2018. V. 52. № 13. P. 1547–1568.
Triebel E. Camptocythere, eine neue Ostracoden-Gattung aus dem Dogger Norddeutschlands // Senckenbergiana. 1950. Bd 31. № 3/4. S. 197–208.
Wannenmacher N., Dietze V., Franz M., Schweigert G. New records of ostracods and ammonites from the Aalenian (mainly Concavum Zone) of the Zollernalb (Swabian Alb, SW Germany) // Zitteliana. 2021. V. 95. P. 1–55. https://doi.org/10.3897/zitteliana.95.56296
Whatley R.C., Ballent S., Armitage J. Callovian Ostracoda from the Oxford Clay of Southern England // Rev. Esp. Micropaleontol. 2001. V. 33. № 2. P. 135–162.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал