

Палеонтологический журнал, 2022, № 4, стр. 12-17
Rusophycus (Hemisphaerichnus) communitas – новый таксон ихнофоссилий в ордовике Тывы
Н. В. Сенников *
Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
630090 Новосибирск, Россия
* E-mail: sennikovnv@ipgg.sbras.ru
Поступила в редакцию 20.07.2021
После доработки 11.01.2022
Принята к публикации 11.01.2022
- EDN: RXEEAY
- DOI: 10.31857/S0031031X22040110
Аннотация
В ордовикских терригенных образованиях Тывы нередко встречаются следы жизнедеятельности организмов. Наиболее богатое местонахождение ихнофоссилий обнаружено в разрезе Чиргакы в западной части региона. Там наряду со спорадически встречающимися ранее известными в других регионах мира таксонами ихнофоссилий, такими как Dimorphichnus Seilacher, Helmintopsis Heer, автором статьи определены преобладающие по численности ихнофоссилии, которые следует относить к новому виду нового подрода рода Rusophycus Hall.
ВВЕДЕНИЕ
Ордовикские отложения имеют в Тыве достаточно широкое распространение и представлены, главным образом, терригенными образованиями. Число местонахождений, содержащих в разрезах фаунистические остатки, крайне незначительно. В разрезах некоторых свит сами органические остатки и их следы полностью отсутствуют, а в других имеются только находки следов жизнедеятельности организмов. Информация об ихнофоссилиях и об обстановках седиментации ордовикских бассейнов Тывы содержится в ряде работ (Владимирская, 1960; Задорожная, 1961; Сенников В.М., 1977; Kulkov, 1991; Сенников Н.В., 2005, 2018, 2019; Sennikov, 2006; Сенников, Обут, 2018; Sennikov, Obut, 2019).
МАТЕРИАЛ
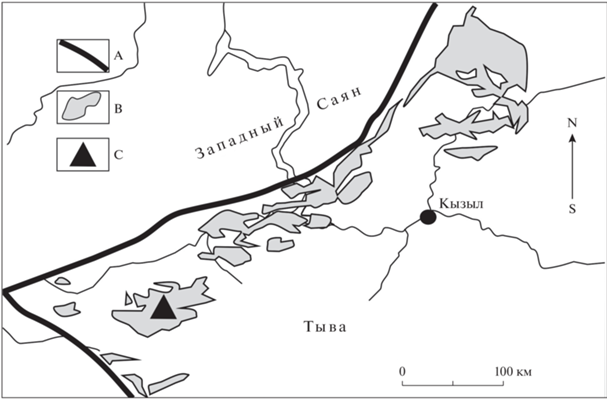
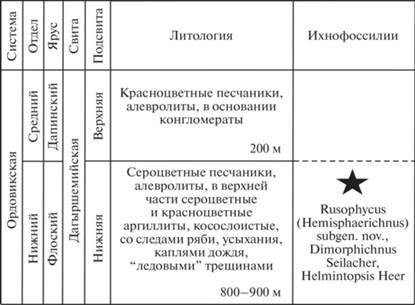
Наиболее значительные площади распространения ордовикских отложений в Тыве занимает шемушдагская серия, подразделяющаяся на дагыршемийскую, аянгатинскую и адырташскую свиты. В настоящей работе описываются ихнофоссилии, встреченные в стратотипе дагыршемийской свиты, в нижней ее подсвите в районе пос. Чиргакы (рис. 1, 2).
Рис. 1.
Схема расположения тувинского ордовикского разреза Чиргакы, содержащего рассмотренные ихнофоссилии: А – крупнейшие тектонические зоны, B – поля выходов ордовикских отложений, C – местоположение района исследований.
Литотипы пород дагыршемийской свиты шемушдагской серии Тывы, по которым можно судить о фациальных обстановках формирования осадочных тел со следами жизнедеятельности организмов, имеют следующие характеристики: а) грунты с воздействием приливов-отливов; б) грунты с высокой и средней гидродинамической энергией; в) мягкие грунты; г) грунты, подвергающиеся высыханию (субаэральные условия); д) линзовидные тела песчаных тел; е) косая слоистость; ж) регрессивный тренд осадконакопления; з) отсутствие самих фаунистических остатков; и) незначительное морфологическое разнообразие ихнофоссилий; к) биотурбация только отдельных слоев c умеренным и низким коэффициентами.
В изученной коллекции дагыршемийских следов жизнедеятельности организмов доминируют ихнофоссилии биоэрозионного типа на плотном (firmground) субстрате. Они имеют положительный гипорельеф (Hypichnia) и представляют из себя ямки-экскавации в виде неглубоких полусфер с желобками на их дне и боковых стенках. Такие ямки-экскавации относятся к структурам заполнения ямок-углублений в нижележащих слоях. Ордовикские тувинские Hypichnia соседствуют в сближенных группировках с другими ямками-экскавациями.
ОПИСАНИЕ ИХНОФОССИЛИЙ
Ихнород Rusophycus Hall, 1852
Типовой вид – Fucoides bilobata Vanuxem, 1842 из нижнего кембрия Пакистана.
Диагноз. Двухлопастные (bilobate) формы, напоминающие по конфигурации кофейные зерна, как правило, со срединным углублением. У ихнофоссилий Rusophycus Hall морщинистые пересекающиеся ребра образуют по отношению к центральной, обычно прогнутой и извилистой, осевой части углы 70°–80° (Vanuxem, 1842; Hall, 1852). У некоторых представителей ихнорода Rusophycus (ряд австралийских и боливийских форм ихновида R. latus) (Webby, 1983; Davies et al., 2007) срединное углубление не имеет прямолинейной направленности и в рельефе выражено крайне слабо.
Состав рода. Два подрода: Rusophycus Hall и новый ихноподрод Rusophycus (Hemisphaerichnus) isubgen. nov. Первый подрод имеет широкое стратиграфическое распространение – весь палеозой многих регионов мира, а второй пока известен только в нижнем ордовике Тывы.
Замечания. Другие известные ихнороды из этологических группировок Domichnia (жилища-норы и охотничьи ямы), Cubichnia (следы покоя), строящие неглубокие норы, и Repichnia (следы направленного передвижения) (Микулаш, Дронов, 2006; Seilacher, 2007; Buatois, Mangano, 2011), отличаются от нового ихноподродового таксона следующими параметрами. Представители ихнорода Bergaueria Prantl имеют неглубокие, цилиндрические, вертикальные норы с ровными стенками, заканчивающимися полусферическим дном. У этих ихнофоссилий каких-либо отпечатков желобков на стенках нет.
Представители этологической группировки Repichnia (следы направленного перемещения) в составе ихнорода Cruziana d’Orbigny имеют линейную двухлопастную (bilobate) форму, реже ямки-экскавации. Пересекающиеся ребра у ихнорода Cruziana образуют по отношению к центральной, обычно прогнутой и относительно прямой осевой части, углы 30°–60°.
Ихноподрод Rusophycus (Hemisphaerichnus) Sennikov, isubgen. nov.
Название подрода от hemisphaera греч. – полусфера (по форме нор-экскаваций).
Типовой вид – Rusophycus (Hemisphaerichnus) communitas isp. nov.
Диагноз. Простые, неглубокие, вертикальные или со слабым наклоном ямки-экскавации округлых очертаний, реже вытянуто-овальные. Общая форма ямок-экскаваций полусферическая, реже усеченной полусферы. На стенках нор видны отпечатки сегментов тела организма-хозяина. Ямки-экскавации могут быть одиночными и формировать группировки многочисленных сближенных (до соприкасающихся) углублений.
Организмы-хозяева выкапывали ямки-экскавации в плотном (firmground) осадке, оставшемся после отлива. Занимаемая ямками-экскавациями Rusophycus (Hemisphaerichnus) isubgen. nov. площадь поверхности слоя (плотность “ихнофоссильного освоения”) составляет не более 5–10%. В целом, такие “ихнофоссилиеносные” слои составляют не более 1–2% от общей мощности разреза.
Учитывая незначительную глубину ямок-экскаваций, нет оснований относить тувинские ихнофоссилии Rusophycus (Hemisphaerichnus) isubgen. nov. к норам-жилищам (Domichnia) или охотничьим норам. Предварительно их можно классифицировать как ямки отдыха-укрытия (Cubichnia), как и у всех представителей рода Rusophycus. На применение “организмами-хозяевами” исследуемых ямок-экскаваций для временного (не постоянного) отдыха-укрытия дополнительно указывает наблюдаемый феномен их многократного использования (незакономерно вкладывающиеся друг в друга отпечатки-слепки). При этом, как правило, “вторичное” и последующее использование ямок-экскаваций происходит организмами близкого по размеру к размеру организма первого “строителя-хозяина” такой ямки-экскавации.
Видовой состав. Типовой ихновид.
Сравнение. У форм рассматриваемого нового ихноподрода полностью отсутствует срединное углубление. Таким образом, это “не двухлопастные” (non-bilobate) формы, обособленные внутри рода Rusophycus Hall в отдельный подрод Rusophycus (Hemisphaerichnus) isubgen. nov. Классические двухлопастные (bilobate) формы в составе рода Rusophycus Hall составляют номинативный подрод Rusophycus (Rusophycus) Hall.
Замечания. Поведенческие функции организмов-хозяев, строящих ямки-экскавации Rusophycus (Hemisphaerichnus), выражаются в частой концентрации их в группировки, что может указывать на их предрасположенность к групповому образу обитания.
Следует отметить одно обстоятельство, пока не нашедшее логического объяснения. В разрезе с многочисленными дагыршемийскими ихнофоссилиями в виде ямок-экскаваций, созданных организмами с ярко выраженным сегментированным телом, наблюдаются структуры ползания (Repichnia), с которыми, предположительно, можно было бы каким-то образом соотносить передвижения таких крупных организмов. Но следы ползания встречаются крайне редко.
Фациальная приуроченность ихнофоссилий Rusophycus (Hemisphaerichnus) isubgen. nov. выражается в их встречаемости в тонких терригенных осадках – аргиллитах, реже алевролитах, вблизи их границ с песчаниками. По этологическим особенностям изученные тувинские ихнофоссилии представляют собой структуры заполнения ямок-углублений в нижележащих слоях.
Rusophycus (Hemisphaerichnus) communitas Sennikov, isp. nov.
Табл. II , фиг. 1–8 ; табл. III , фиг. 1–8 (см. вклейку)
Название вида communitas лат. – группировка.
Голотип – Музей ИНГГ, № 2107/5; Тыва, водораздел рр. Улуг-Чиргакы и Б. Аянгаты, разрез “Чиргакы”, 16-ая пачка; нижний ордовик, флоский ярус, дагыршемийская свита, нижняя подсвита.
Описание. Обособленные ямки-экскавации, созданные отдельным индивидом организма-хозяина в виде четких полусфер, чаще их соприкасающиеся группировки в более крупных округлых и овальных образованиях – углублениях-выемках усеченной полусферической формы.
Размеры. Ямки-экскавации имеют размеры по длинной оси от 6–8 мм до 10 см, а по короткой оси от 4–6 мм до 9 см. Отношение ширины ямки-экскавации к ее длине в среднем 0.75, реже 1.0. Углубления-выемки, в которые группируются экскавации, имеют размеры от 3–5 см до 12 см.
Изменчивость и параметры, которые можно интерпретировать как возрастные изменения организмов-хозяев – это, прежде всего, длина, а так как они все полусферической формы, то и глубина экскаваций. Ямки-экскавации по их размерам разделены на три условные категории: мелкие (длина до 0.2–0.3 см), средние (длина до 2–4 см) и крупные (длина до 5–7 см). Возможно, что такое разделение можно интерпретировать как следствие жизнедеятельности юных форм, взрослых особей видового таксона средних размеров и геронтологических особей этого же вида или, что менее вероятно, взрослых особей другого видового таксона крупных размеров.
Отношение глубины экскаваций к их длине: у мелких 0.75, у средних 0.75, у крупных 0.4. Отношение глубины экскаваций к их ширине: у мелких 0.5, у средних 0.5, у крупных 0.55. Число отпечатков сегментов тела в экскавациях: у мелких 6–9, у средних 10–15, у крупных 10–20. Границы сегментов иногда слабо изгибаются, чаще строго дугообразные. Ширина сегментов отпечатков тела в экскавациях: у мелких 0.5–0.7 мм, у средних 1.5–3.0 мм, у крупных до 7–8 мм. Глубина отпечатка сегмента тела в экскавациях по отношению к разделяющей сегменты более узкой борозде 0.1–0.4 мм. Ширина борозд, разделяющих сегменты в экскавациях: у мелких 0.4–0.5 мм, у средних 2 мм, у крупных 4–5 мм.
Замечания. Новый тувинский ихнотаксон, кроме отсутствия двухлопастного строения, по своим параметрам (отношение глубины экскаваций к их длине или ширине, характеризующем слабо усеченную полусферу) отличается от всех других ихновидов рода Rusophycus Hall. По отношению длины к ширине экскавации новый ихнотаксон можно сравнить с ихновидом R. leiferikssoni Bergström (Bergström, 1976; Mangano, Buatois, 2003; Seilacher, 2007). Сибирские ихнофоссилии R. communitas имеют величину этого параметра 0.75–1.0, а формы ихновида R. leiferikssoni – 0.5–0.8. Кроме того, характерной особенностью для R. leiferikssoni является четко выраженная двухлопастность строения и сильный наклон экскаваций с глубоким узким концом, а у тувинского ихнотаксона экскавации практически вертикальны.
ЗАКЛЮЧЕНИЕ
По полученным и приведенным выше материалам можно сделать некоторые выводы:
I. По условиям обитания и стратегии поведения нового тувинского ордовикского ихнотаксона изученная коллекция его следов жизнедеятельности позволяет констатировать следующее. Для организмов, оставивших такие следы, было характерно: 1) поселение не на твердом (hardground), а на плотном (firmground) грунте, который они могли использовать для создания ямок-экскаваций; 2) вероятно, группировки ямок-экскаваций (углубления-выемки) создавались животными в микропонижениях рельефа дна палеобассейна, где при отливах оставались лужи воды – такое местообитание можно называть лужей-лежбищем (возможно сходство с современными литоральными ваннами, не осушающимися при отливах); 3) создание компактных поселений (“оседлость”) – повышенная концентрация на ограниченной площади поверхности слоя; 4) зависимость от периодических процессов, выражающаяся в многократном посещении одних и тех же ямок-экскаваций (возможно, и одним и тем же индивидом или близким к нему по размерам сородичем), с пропечатыванием следов сегментов их тел поверх уже имеющихся сегментированных отпечатков; 5) “стайность” особей одного размера, фиксируемая в нахождении внутри одного углубления-выемки только отпечатков “сеголеток”, или в другом месте исключительно отпечатков взрослых особей, или в третьем месте малочисленных отпечатков гигантских геронтических экземпляров организмов-хозяев; 6) создание компактных группировок ямок-экскаваций, в которых могла быть температура выше, чем у окружающего воздуха, а такая температура, судя по присутствию в породах ихнофоссилиеносных слоев следов “морозных ледовых” трещин (Сенников, 2019), могла в ночное время опускаться ниже 0 градусов.
II. В отношении предположения о систематической принадлежности биологических объектов – организмов-хозяев следов жизнедеятельности ордовикских тувинских Rusophycus (Hemisphaerichnus) предварительно можно отметить следующее. В ряде работ убедительно показано, что многие Rusophycus Hall являются следами отдыха артропод – трилобитов (Orlowski et al., 1970; Orlowski, 1992; Aceñolaza, 2003; Mangano, Droser, 2004). Имеется информация (Mangano et al., 2020), что кроме трилобитов, такие следы в виде ямок-укрытий могли формировать (“создавать”) эуартроподы.
Ордовикские тувинские Rusophycus (Hemisphaerichnus) не могли “образовывать” не только отдыхающие артроподы (членистоногие), но и свернувшиеся их формы внутри ямок-экскаваций. В противном случае в тувинском материале должны были остаться отпечатки границ туловищных сегментов и плевральных шипов. Возможно, что “хозяевами” следов жизнедеятельности ордовикских тувинских Rusophycus (Hemisphaerichnus) были Malacostraca – высшие ракообразные, некоторые представители которых известны с кембрийского периода. Высшим ракообразным свойственны стайность и стратегия многократного использования собственных нор.
* * *
Описанная в работе коллекция ихнофоссилий хранится в ЦКП “Геохрон” при Ин-те нефтегазовой геологии и геофизики СО РАН в г. Новосибирск под № 2107. Исследования проводились в рамках Госзадания РАН FWZZ-2022-0003.
Автор выражает признательность специалистам по палеозойским ихнофоссилиям Л. Бато (L. Buatois, Канада) и Х. Гутиеррес-Марко (J.‑C. Gutiérrez-Marco, Испания) за ценные советы, полученные им при дискуссиях с этими коллегами в процессе работы над каменным материалом. Автор благодарит своих коллег О.Т. Обут, Н.Г. Изох, Т.В. Хлебникову, Д.А. Токарева за помощь в сборе тувинских ихнофоссилий. От А.В. Дронова и В.Б. Кушлиной на заключительном этапе работы над рукописью были получены ценные замечания, которые помогли улучшить качество изложения материала.
Список литературы
Владимирская Е.В. Ордовикские отложения Центральной и Западной Тувы // Зап. Ленингр. горн. ин-та. 1960. Т. 37. Вып. 2. С. 21–48.
Задорожная Н.М. Стратиграфия ордовикских отложений юго-западной оконечности Кортушубинского хребта // Матер. по региональной геологии Алтае-Саянской складчатой области. Л.: ВСЕГЕИ, 1961. С. 43–48 (Тр. ВСЕГЕИ. Нов. сер. Т. 58).
Микулаш Р., Дронов А. Палеоихнология. Введение в изучение ископаемых следов жизнедеятельности. Прага: Изд-во Геол. ин-та Акад. наук Чешской Республики, 2006. 122 с.
Сенников В.М. История развития структур южной части Алтае-Саянской складчатой области в ордовике. Барнаул: Алтайское книжн. изд-во, 1977. 135 с.
Сенников Н.В. Ихнофации и ихнофоссилии в нижнем палеозое Тувы // Эволюция жизни на Земле. Матер. III Междунар. симп. Томск: ТГУ, 2005. С. 158–160.
Сенников Н.В. Фациальная приуроченность, топономическая и этологическая характеристики ихнофоссилий дагыршемийской свиты среднего ордовика Тывы // Фундаментальная и прикладная палеонтология. Матер. LХIV сессии Палеонтол. об-ва при РАН. СПб.: Изд-во ВСЕГЕИ, 2018. С. 111–113.
Сенников Н.В. Лито- и ихнофации дагыршемийской свиты раннего–среднего ордовика Тывы // Тр. Палеонтол. об-ва. Т. 2. М.: ПИН РАН, 2019. С. 133–145.
Сенников Н.В., Обут О.Т. Экосистемы ордовикского бассейна Центральной Тывы // Матер. V Междунар. симп. “Эволюция жизни на Земле”, 12–16 ноября 2018. Томск: Издательский дом Томского госун-та, 2018. С. 117–119.
Aceñolaza G.F. Olenidae (Trilobita) – Rusophycus isp.: organismo productor-traza fósil resultante. Ejemplos para el análisis en el Cambro/Ordovícico de la Cordillera Oriental Argentina // Ameghinana. 2003. V. 40. № 4. P. 573–583.
Bergström J. Lower Paleozoic trace fossils from eastern Newfoundland // Can. J. Earth Sci. 1976. V. 13. P. 1613–1633.
Buatois L.A., Mangano M.G. Ichnology. Organism-Substrate Interactions in Space and Time. Cambridge: Cambridge Univ. Press, 2011. 358 p.
Davies N.S., Sansom I.J., Albanesi G.L. et al. Ichnology, palaeoecology and taphonomy of a Gondwanan early vertebrate habitat: Insights from the Ordovician Anzaldo Formation, Bolivia // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2007. V. 249. P. 18–35.
Hall J. Palaeontology of New-York. V. 2. Containing Descriptions of the Organic Remains of the Lower Middle Division of the New-York System (Equivalent in Part to the Middle Silurian Rocks of Europe). Albany: C. Van Benthuysen, 1852. 362 p.
Kulkov N.P. The trace fossil Thalassinoides from the Upper Ordovician of Tuva // Lethaia. 1991. V. 24. P. 187–190.
Mangano M.G., Buatois L.A. Rusophycus leiferikssoni Bergström en la Formación Campanario: implicancias en la paleoecologia de planicies marealis cámbricas // APA. 2003. Publ. Especial 9. P. 65–84.
Mangano M.G., Droser M.L. The ichnologic record of the Ordovician radiation // The Great Ordovician Biodiversification Event / Eds. B. Webby, F. Paris, M.L. Droser, I.G. Percival. N.-Y.: Columbia Univ. Press, 2004. P. 369–379.
Mángano M.G., Ortega-Hernández J., Piñuela L. et al. Trace fossil evidence for infaunal moulting in a Middle Devonian nontrilobite euarthropod // Sci. reports. 2020. 10:5316. 15 p. https://doi.org/10.1038/s41598-020-62019-6
Orlowski S. Trilobite trace fossils and their stratigraphical significance in the Cambrian sequence of the Holy Cross Mountains, Poland // Geol. J. 1992. V. 27. P. 15–34.
Orlowski S., Radwanski A., Roniewicz P. The trilobite ichnocoenoses in the Cambrian of the Holy Cross Mountains // Trace fossils / Eds. T.P. Crimes, J.C. Harper. Liverpool, 1970. P. 345–360 (Geol. J. Spec. Iss. № 3).
Seilacher A. Trace Fossil Analysis. Berlin, Heidelberg, N.Y.: Springer, 2007. 226 p.
Sennikov N.V. Lower Ordovician paleogeographical environments and ichnofossils from Dagyr-Shemi Formation (Tuva region, Siberia, Russia) // Intern. Symp. “Palaeogeography and Global Correlation of Ordovician Events”. IGCP 503 Project. Contributions. Novosibirsk, Russia (August 5–7, 2006). Novosibirsk: Publ. House of SB RAS, “Geo” Branch, 2006. P. 53–54.
Sennikov N.V., Obut O.T. Ecosystem reconstruction for the Ordovician Basin of the Central Tuva (South Siberia) // 13th Intern. Symp. on the Ordovician System: Contributions. Novosibirsk, Russia (July 19–22, 2019) / Eds. O.T. Obut, N.V. Sennikov. Novosibirsk: Publ. House of SB RAS, 2019. P. 183–185.
Vanuxem L. Geology of New York. Part. III. Comprising the Survey of the Third Geological District. Albany, 1842. 306 p.
Webby B.D. Lower Ordovician arthropod. Trace Fossils from western New South Wales // Proc. Linn. Soc. N.S.W. 1983. V. 107. № 2. P. 59–74.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал