

Палеонтологический журнал, 2022, № 4, стр. 3-11
Цианобактериальные сообщества завханской ассоциации позднего венда и палеообстановки формирования вендо-кембрийских отложений Западной Монголии
А. Л. Рагозина a, Д. Доржнамжаа b, *, Е. А. Лужная (Сережникова) a, **, Л. В. Зайцева a, Б. Энхбаатар b
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
b Палеонтологический институт Монгольской академии наук
5826128 Улаанбаатар, Монголия
* E-mail: ddorj2001@yahoo.com
** E-mail: serezhnikova@mail.ru
Поступила в редакцию 25.11.2021
После доработки 14.02.2022
Принята к публикации 21.02.2022
- EDN: MHJTIV
- DOI: 10.31857/S0031031X22040092
Аннотация
В составе завханской циано-альго-бактериальной ассоциации верхнего венда верхней части терригенно-карбонатного разреза цаганоломской свиты (<632 ± 14 млн лет) Дзабханской структурно-фациальной зоны запада Монголии установлены остатки бентосных цианобактериальных сообществ, сформированных в мелководных литоральных условиях. Основными строителями матов являлись нитчатые цианобактерии рода Siphonophycus. Бентосные цианобактериальные сообщества маркируют мелководные фации регрессивного этапа развития обширного постледникового вендо-кембрийского Дзабханского палеобассейна запада Монголии.
ВВЕДЕНИЕ
Цианобактериальные сообщества являются самыми древними экосистемами в геологической летописи Земли, их возраст свыше двух млрд лет. Наряду с абиотическими факторами, эти сообщества играли существенную роль в формировании облика планеты, в частности, седиментогенеза и климата. Это во многом предопределило процесс становления биосферы современного типа. Поэтому интерес к их изучению становится особенно активным с середины XX в., когда наступает время мультидисциплинарных исследований древних и современных геобиоценозов (Бактериальная …, 2002; Заварзин, 2004 и др.).
Как известно, глобальные позднепротерозойские оледенения оказали огромное влияние на древние экосистемы и, как считается, способствовали постепенной смене преимущественно прокариотной биосферы на биосферу современного типа (Федонкин, 2003 и др.). Одним из ключевых моментов обоснования таких построений является изучение непрерывных вендо–кембрийских последовательностей, с тиллитами (диамиктитами) в основании, богатых разнообразными остатками растительной и животной жизни.
В настоящей работе приведены результаты биостратиграфических исследований авторского коллектива в Дзабханской структурно-фациальной зоне Западной Монголии, которую можно назвать одним из полигонов для решения целого ряда комплексных геологических задач (Доржнамжаа и др., 2018; Dorjnamjaa et al., 2021).
МАТЕРИАЛ И МЕТОДИКА
Материал для изучения (кремнисто-карбонатные, фосфатизированные и углефицированные сланцы) был собран участниками Российско-Монгольской палеонтологической экспедиции Д. Доржнамжаа, А.В. Краюшкиным, А.Л. Рагозиной, Е.А. Лужной (Сережниковой) и Б. Энхбаатаром из естественных обнажений цаганоломской свиты Дзабханского района: из ручьев Баян Гол, Цаган Гол, района Тайшир сум (села), хр. Хэвтээ Цахир. Исследование пород проводили в тонких петрографических шлифах на световом биологическом микроскопе Carl Zeiss, а также методом сканирующей электронной микроскопии (SEM) на электронном микроскопе EVO-50 Zeiss c микроанализатором INCA Oxford 350 (Лондон) при 15–20 кВ с применением EDX (энергодисперсионного рентгеновского микроанализа). Полуколичественный элементный анализ состава участков микронного и меньшего размеров проводили с использованием EDX-спектрометра.
Коллекция № 5492 хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН РАН).
СТРАТИГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ, ПАЛЕООБСТАНОВКИ
В Дзабханской структурно-фациальной зоне, в районе руч. Цаган Гол находится один из наиболее полных разрезов венда (майханульская и цаганоломская свиты) с характерными ледниковыми образованиями в основании (Khomentovsky, Gibsher, 1996; Lindsay et al., 1996; Кузнецов, Семихатов, 2014; Рагозина и др., 2016; Доржнамжаа и др., 2018) (рис. 1). Этот разрез охватывает почти всю вендскую систему. Абсолютный возраст надтиллитовых карбонатов цаганоломской свиты составляет 632 ± 14 млн лет (Овчинникова и др., 2012).
Рис. 1.
Схема расположения фосфоритовых месторождений и проявлений в Дзабханском бассейне. V – месторождение Алагийн Даваа и Цахир Уул, проявления: I – Баруун Арц, II – Зуун Арц, III – Буурал Уул, IV – Богдын Гол, VI – Аргалант Уул, VII – Хэвтээ Цахир, VIII – Баян Гол, IX – Салааны Гол.
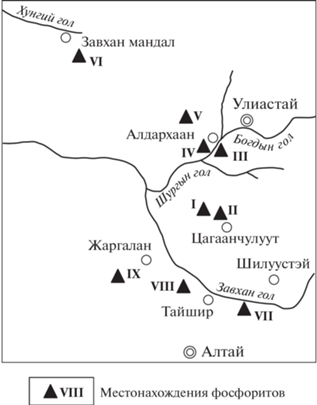
Накопление терригенно-карбонатных пород цаганоломской свиты сопровождалось образованием маломощных горизонтов фосфоритоносных отложений, главным образом, в северной части Дзабханского бассейна (Алагиндабанское, Цахирулинское), отдельные выходы отмечены в районах зон Зуун Арц, Баруун Арц, Баян Гол и Хэвтээ Цахир (рис. 1).
Постледниковая морская трансгрессия в конце венда сменилась регрессией, в результате которой произошла смена бесструктурных и доломитистых фосфоритов сначала вивианитовыми зернистыми, а позже, в подошве баянгольской свиты, песчаниковыми [высокоглиноземистыми (22–23, реже до 33% Al2O3) каменистыми или коллофанитовыми] фосфоритами (Доржнамжаа и др., 2018; Dorjnamjaa et al., 2021). Формирование кремнисто-карбонатных толщ с фосфоритоносными горизонтами, начавшееся в конце цаганоломского времени в результате периодической трансгрессии и расширения морского бассейна происходило и в раннекембрийское время (баянгольская и саланыгольская свиты). Начало томмотского времени характеризуется расцветом органической жизни и появлением многочисленных скелетных организмов (SSFs). Наибольшее количество остатков фауны томмотской ассоциации приурочено к нижней и средней части баянгольской свиты (Коробов, Миссаржевский, 1977; Воронин и др., 1982; Есакова, Жегалло, 1996). В этих отложениях также содержатся многочисленные следы жизнедеятельности древних организмов, свидетельствующие о разнообразии органической жизни вендо-кембрийского бассейна (Goldring, Jensen, 1996; Рагозина и др., 2007). Чрезмерное мелководье Дзабханского палеобассейна допускало, но не стимулировало накопление водорослевой органики, необходимой для формирования крупных фосфатных месторождений. Обильный привнос кварцевого песка во время фосфоритообразования и строматолитовые биогермы свидетельствуют о литоральной обстановке, которая перемежалась лагунными углублениями, когда формировались черные сланцы. В это время фосфоритообразование прекращалось. Дзабханский вендо-кембрийский морской бассейн был столь же мелководным, как и продуктивные фосфоритоносные бассейны Каратау, Прихубсугулья и Китая (Dorjnamjaa et al., 2021).
Вендо-кембрийский период на территории Западной Монголии характеризуется прогибанием и обширной постледниковой морской трансгрессией. Этот этап отличается единым субплатформенным седиментационным циклом развития осадконакопления. В венде и в начале кембрия формировались шельфовые мелководные морские бассейны (рис. 2). Одним из них был вендо-кембрийский Дзабханский бассейн. В начале стабилизации происходили кратковременные блоковые подвижки, о чем свидетельствуют маломощные конгломераты и гравелиты в трансгрессивной серии терригенно-карбонатных пород.
Рис. 2.
Палеогеографическая схема цаганоломско–баянгольского времени венда–нижнего кембрия Монголии. Обозначения: 1 – суша – области питания; 2 – мелководное субплатформенное шельфовое море; 3 – эффузивы преимущественно среднего и кислого состава, пирокласты смешанного состава и их туфы; 4 – участки фосфатонакопления; 5 – глубоководные вулканогенные и кремнистые осадки (офиолитовые образования); 6 – мощность по разрезам отложения.
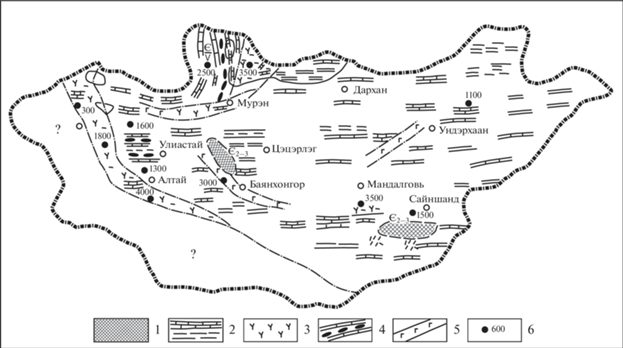
Дзабханская структурно-фациальная зона является специфической биогеографической провинцией, близкой к Алтае-Саянской складчатой области, с одной стороны, и Сибирской платформе – с другой.
ЗАВХАНСКАЯ АССОЦИАЦИЯ. ЦИАНО-БАКТЕРИАЛЬНЫЕ МАТЫ
Завханская ассоциация водорослей, микрофоссилий и проблематик позднего венда (эдиакария) была установлена в верхней части цаганоломской свиты в маркирующем кремнисто-фосфоритовом горизонте (рис. 3), распространенном по всей территории Дзабханского района (Рагозина и др., 2007, 2016, 2019 и др.; Доржнамжаа и др., 2018). Последние палеобиологические исследования вендских (эдиакарских) отложений в Дзабханской структурно-фациальной зоне также подтверждают наличие богатой ассоциации микрорастительных остатков и разнообразных микрофоссилий в Дзабханском морском палеобассейне (Anderson et al., 2017).
Рис. 3.
Маркирующий кремнисто-фосфоритовый горизонт Дзабханской структурной зоны. Западная Монголия, разрез Хэвтээ Цахир; верхний венд, цаганоломская свита.

Завханская ассоциация содержит многочисленные минерализованные сфероморфные микрофоссилии Tasmanites mongolicus Ragozina, 2016, Megasphaera sp. (?), Archaeooides sp., Leiosphaeridia sp., акантоморфные акритархи родов Tanarium (рис. 4, а) и Cavaspina (рис. 4, б) (Рагозина и др., 2016), фрагменты слоевищ вендотениевых водорослей (рис. 4, в, г), водоросли Rodophyta (?), Douchantuophyton zavkhanicus Ragozina et Luzhnaya (Serezhnikova), 2019, проблематики Octoedrixium truncatum Rud., 1989, Beltanelliformis (?) brunsae Menner, 1974.
Рис. 4.
а, б – акантоморфные акритархи завханской ассоциации: а – Tanarium sp.; б – Cavaspina sp.; в, г – фрагменты тканеподобных вендотениевых водорослей; Западная Монголия, разрез сухого русла р. Баян Гол; верхний венд, цаганоломская свита.
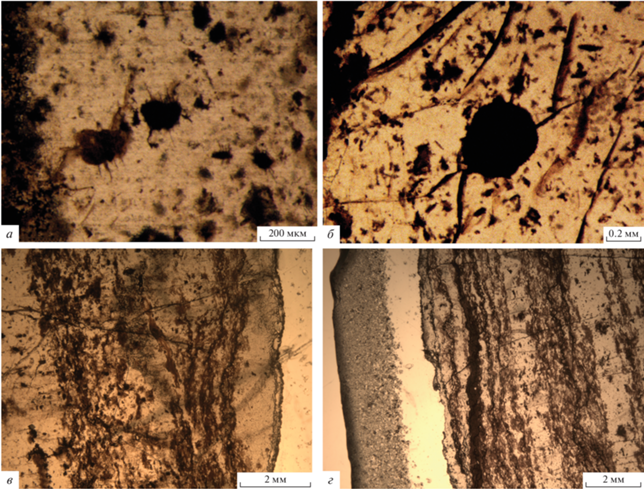
В позднем венде богатые комплексы ассоциации Doushantuo-Pertatataka резко сокращаются, и акантоморфные микрофоссилии (акритархи) в ней представлены мелкими формами со слаборазвитой орнаментацией (Бактериальная …, 2002). По комплексу сфероморфных микрофоссилий завханская ассоциация может рассматриваться как поздневендская (позднеэдиакарская), что согласуется с биостратиграфическими, хемостратиграфическими и радиометрическими характеристиками цаганоломской свиты. Эта ассоциация коррелируется с биотами верхней части формации Doushantuo-Pertatataka Южного Китая, а также с поздневендской редкинско-котлинской ассоциацией севера Восточно-Европейской платформы. Завханская биота может рассматриваться как поздневендская и сопоставляться с редкинским горизонтом Общей стратиграфической шкалы России или с верхней частью эдиакарской системы МСШ (Рагозина и др., 2016). Ледниковые отложения майханульской свиты нижнего венда коррелируются с отложениями формации Nantuo (эпоха Marino) (Khomentovsky, Gibsher, 1996; Кузнецов и др., 2014), что согласуется с нашими биостратиграфическими данными.
Характерной особенностью завханской ассоциации является широкое развитие минерализованных цианобактериальных матов и биопленок. Бентосные цианобактериальные сообщества занимали главенствующее место в вендских мелководных морских палеобассейнах.
Основными компонентами цианобактериальных матов Дзабханского палеобассейна являются цианобактерии, относящиеся к роду Siphonophycus (Schopf, 1968) emend. Knoll et al., 1991. Они представляют собой полые трубчатые несептированные неветвящиеся образования. Структура наружной поверхности варьирует от гладкой до мелкозернистой. Диаметр этих образований от 0.5 до 8–10 мкм, 25–30, реже до 60 мкм. Цианобактерии Siphonophycus формально разделены на несколько видов, имеющих разный диаметр (Butterfield et al., 1994). В Дзабханском вендском морском бассейне распространены, в основном, цианобактерии Siphonophycus tipicum (Hermann) Butterfield et al., 1994 и S. robustum (Schopf, 1968) Knoll et al., 1991.
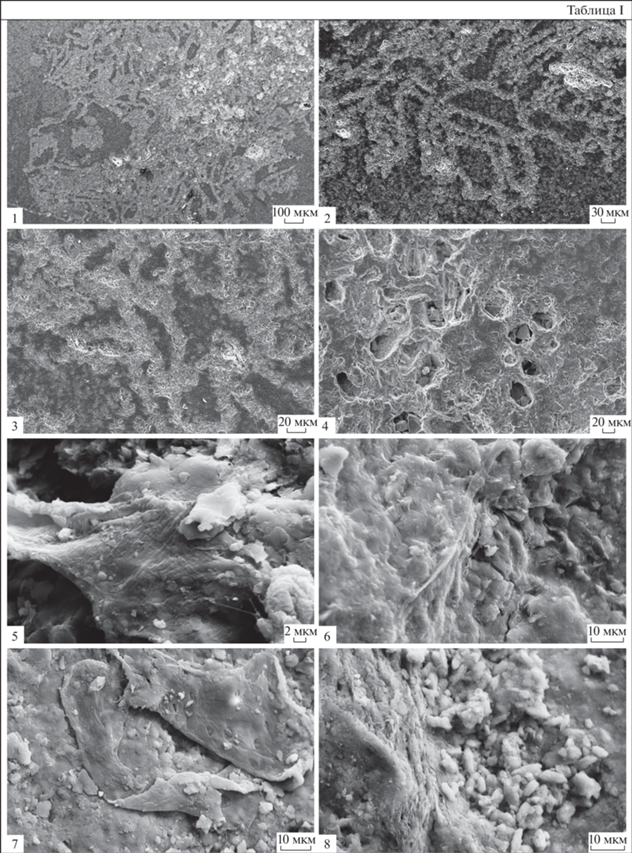
На сканирующем электронном микроскопе исследованы цианобактерии S. robustum (Schopf, 1968) Knoll et al., 1991 из разных частей мата. Они образуют скопления из многочисленных сложно-переплетенных трубчатых нитей (табл. I , фиг. 1–4). Размеры цианобактерий составляют 1.7–2 мкм в диаметре. Обнаружено различие в составе цианобактерий и вмещающих кремнисто-карбонатных сланцев (рис. 5, в): цианобактерии отличаются повышенным содержанием кальция и фосфора (рис. 5, б).
Рис. 5.
Цианобактериальные сообщества завханской ассоциации: а – минерализованный цианобактериальный мат, сложенный нитчатыми формами Siphonophycus; б, в – результаты анализов. Западная Монголия, разрез Хэвтээ Цахир; верхний венд, цаганоломская свита.

Ископаемые остатки Siphonophycus обнаруживают сходство с бентосными гормогониевыми цианобактериями Microcoleus, широко распространенными в современных мелководных бассейнах (Бактериальная …, 2002). Современные цианобактериальные маты представляют собой бентосные сообщества разнородных организмов, объединенных общим внеклеточным веществом (гликокаликс), выделяемым самими цианобактериями. Устойчивость цианобактериальным матам придают экзополимеры. Цианобактерии образуют “псевдоткань”, в которой могут развиваться другие микроорганизмы (Герасименко, Заварзин, 1993; Заварзин, 2004 и др.). Слизистый чехол цианобактерий выполняет защитную функцию. Толщина чехла может увеличиваться в зависимости от различных факторов. В современных матах при благоприятных условиях цианобактерии располагаются горизонтальными слоями в верхней части мата. При экстремальных условиях они выползают из общего чехла. Они способны пробуравливать верхний слизистый слой мата и выходить на поверхность, образуя новый слой (Бактериальная …, 2002 и др.).
В завханской ассоциации сохраняется только верхняя часть минерализованного цианобактериального мата. Здесь наблюдаются процессы, аналогичные происходящим в современных матах. Установлены фрагменты минерализованного мата с трубчатыми пустыми чехлами цианобактерий Siphonophycus и трихомами водорослей, пробуравивших верхний слой и вышедших из чехлов на поверхность мата (табл. I , фиг. 4).
Объяснение к таблице I
Фиг. 1–8 . Цианобактериальные сообщества завханской ассоциации; Западная Монголия, разрез Хэвтээ Цахир; верхний венд, цаганоломская свита.
Многочисленные биопленки, содержащиеся в кремнисто-карбонатных породах, в отличие от слоистых цианобактериальных матов, представляют собой различные микробиальные структуры на поверхности цианобактериальных сообществ (Заварзин, 2004).
В завханской ассоциации присутствуют также минерализованные фрагменты верхней части цианобактериальных матов, представленные корковидными образованиями разнообразной формы (табл. I , фиг. 5–7 ). Поверхность их обычно гладкая, морщинистая или ребристая. Верхние слои современных цианобактериальных матов в неблагоприятных условиях сильно ослизняются, создают пленку, которая при подсыхании образует корки. Корковидные образования завханской ассоциации являются фрагментами литифицированной экзополимерной матрицы верхней части цианобактериальных матов.
В Дзабханской структурно-формационной зоне широко распространены строматолитовые биогермы с доминирующим видом Boxonia grumulosa Komar. Они наряду с вышележащими кремнисто-карбонатными фосфоритами являются хорошим стратиграфическим маркером. В результате исследования строматолитов верхней части цаганоломской свиты были обнаружены ультрамикроскопические структуры (табл. I , фиг. 8 ), представленные нитчатыми, столбчатыми, палочковидными формами, которые принимали участие в образовании цианобактериального мата (Рагозина и др., 2018). Древние строматолиты, как и современные, развивались в мелководных бассейнах. Форма построек была обусловлена гидродинамическими условиями. Строматолиты формировались за счет минерализации бентосных цианобактериальных сообществ (матов) и доминировали в докембрийских мелководно-морских обстановках.
Быстрое захоронение микрофоссилий и минерализация полимерной матрицы матов обеспечивали хорошую сохранность органических остатков. Первоначальное органическое вещество (кероген) минерализовано кремнистыми карбонатами с участием фосфатов.
ЗАКЛЮЧЕНИЕ
В венде и в начале кембрия, в постледниковое время, на территории Монголии существовали шельфовые мелководные морские бассейны, которые были частично изолированы участками суши. Одним из них был вендо-кембрийский Дзабханский бассейн. Поздневендская завханская ассоциация, установленная в верхней части цаганоломской свиты, содержит многочисленные остатки цианобактериальных сообществ: сфероморфные микрофоссилиии, цианобактериальные маты, биопленки, корковидные образования. Цианобактериальные сообщества, разнообразные водоросли и обилие кислорода создавали благоприятные условия для развития богатой органической жизни.
Работа выполнена при поддержке гранта РФФИ 20-55-44010 Монг_а.
Список литературы
Бактериальная палеонтология / Ред. Розанов А.Ю. М.: ПИН РАН, 2002. 188 с.
Воронин Ю.И., Воронова Л.Г., Григорьева Н.В. и др. Граница докембрия и кембрия в геосинклинальных областях (опорный разрез Саланы-Гол, МНР). М.: Наука, 1982. 152 с. (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 18).
Герасименко Л.М., Заварзин Г.А. Реликтовые циано-бактериальные сообщества // Проблемы эволюции ведущих компонентов биосферы. М.: Наука, 1993. С. 221–252.
Гниловская М.Б. О древнейшей тканевой дифференциации докембрийских (вендских) водорослей // Палеонтол. журн. 2003. № 2. С. 92–98.
Доржнамжаа Д., Бат-Ирээдуй Я., Рагозина А.Л., Алтаншагай Г. Бактериальная палеонтология Монголии. Улаанбаатар, 2018. 175 с.
Есакова Н.В., Жегалло Е.А. Биостратиграфия и фауна нижнего кембрия Монголии. М.: Наука, 1996. 216 с. (Тр. Совм. Росс.-Монгол. палеонтол. экспед. Вып. 46).
Заварзин Г.А. Лекции по природоведческой микробиологии. М.: Наука, 2004. 348 с.
Коробов М.Н., Миссаржевский В.В. О пограничных слоях кембрия и докембрия Западной Монголии (хребет Хасагт-Хайрхан) // Беспозвоночные палеозоя Монголии. М.: Наука, 1977. С. 7–9 (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 5).
Кузнецов А.Б., Семихатов М.А., Горохов И.М. Возможности стронциевой изотопной хемостратиграфии в решении проблем стратиграфии верхнего протерозоя (рифея и венда) // Стратигр. Геол. корреляция. 2014. Т. 22. № 6. С. 3–25.
Овчинникова Г.В., Кузнецов А.Б., Васильева И.М. и др. U-Pb возраст и Sr-изотопная характеристика надтиллитовых известняков неопротерозойской цаганоломской свиты, бассейн р. Дзабхан, Западная Монголия // Стратигр. Геол. корреляция. 2012. Т. 20. № 6. С. 28–40.
Рагозина А.Л., Доржнамжаа Д., Краюшкин А.В., Сережникова Е.А. Органические остатки и ихнофоссилии в вендо-кембрийских отложениях Дзабханской структурной зоны Западной Монголии. Рассвет и закат вендской (эдиакарской биоты) // Происхождение современной биосферы. Тр. Междунар. конф. по проекту 493 МПГК, 20–31 августа 2007 г. М.: ГЕОС, 2007. С. 57–64.
Рагозина А.Л., Доржнамжаа Д., Лужная (Сережникова) Е.А., Зайцева Л.В. Вендская (эдиакарская) ассоциация водорослей, микрофоссилий и проблематик Западной Монголии и вендо-кембрийская граница // Палеонтология, палеогеография и биостратиграфия Монголии / Pед. Лопатин А.В. М.: ПИН РАН, 2019. С. 206–229 (Тр. Совм. Pocc.-Монгол. палеонтол. экспед. Вып. 48).
Рагозина А.Л., Доржнамжаа Д., Сережникова Е.А. и др. Ассоциация макро- и микрофоссилий в вендских (эдиакарских) постледниковых отложениях западной Монголии // Стратигр. Геол. корреляция. 2016. Т. 24. № 3. С. 27–37.
Рагозина А.Л., Зайцева Л.В., Доржнамжаа Д. Микробиальные ультрамикроскопические структуры запада Монголии // Эволюция жизни на Земле. Матер. V Междунар. симп. (Томск, Томский гос. ун-т, 12–16 ноября 2018 г.). Томск: Издат. дом Томского гос. ун‑та, 2018. С. 114–116.
Федонкин М.А. Сужение геохимического базиса жизни и эвкариотизация биосферы: причинная связь // Палеонтол. журн. 2003. № 6. С. 33–40.
Anderson R.P., McMahon S., Bold U. et al. Palaeobiology of the early Ediacaran Shuurgat Formation, Zavkhan Terrane, south-western Mongolia // J. Syst. Palaeontol. 2017. V. 15. № 11. P. 947–968.
Butterfield N.J., Knoll A.H., Swett K. Paleobiology of the Neoproterozoic Svanbergfjellet Formation, Spitsbergen // Fossils and Strata. 1994. V. 34. 84 p.
Dorjnamjaa D., Enkhbaatar B., Altanshagai G. Sedimentological and bacterial paleontological peculiarity of the vivianite phosphorites in the Zavkhan basin, Western Mongolia // Mongolian J. Paleontol. 2021. V. 4. P. 5–22.
Goldring R., Jensen S. Trace fossils and biofabrics at the Precambrian–Cambrian boundary interval in western Mongolia // Geol. Mag. 1996. V. 133. № 4. P. 403–415.
Khomentovsky V.V., Gibsher A.S. The Neoproterozoic–Lower Cambrian in northern Govi-Altay, western Mongolia: regional setting, lithostratigraphy and biostratigraphy // Geol. Mag. 1996. V. 133. № 4. P. 371–390.
Lindsay J.F., Brasier M.D., Shields G. et al. Glacial facies association in a Neoproterozoic back-arc setting, Zavkhan Basin, Western Mongolia // Geol. Mag. 1996. V. 133. № 4. P. 391–402.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал