

Палеонтологический журнал, 2022, № 4, стр. 23-31
Пористые девонские и раннекарбоновые ринхонеллиды (Brachiopoda) Закавказья
А. В. Пахневич *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: alvpb@mail.ru
Поступила в редакцию 06.04.2021
После доработки 16.12.2021
Принята к публикации 16.12.2021
- EDN: CZKCYQ
- DOI: 10.31857/S0031031X22040079
Аннотация
Представлены данные о разнообразии позднедевонских и раннекарбоновых пористых ринхонеллид Закавказья. Рассмотрены различные возможные причины происхождения пористости. Она появилась в разных надсемействах отряда Rhynchonellida в результате адаптации к обеднению кислородом придонного слоя морской воды. Первые пористые ринхонеллиды появились после фран–фаменского экосистемного кризиса в Закавказье. Это виды Greira transcaucasica Erlanger, 1993 и Sharovaella mirabilis Pakhnevich, 2012, найденные в отложениях зоны Cyrtospirifer asiaticus–Mesoplica meisteri нижнего фамена. Большинство пористых ринхонеллид расселилось вдоль северного побережья Гондваны. И только виды рода Araratella Abramian, Plodowski et Sartenaer, 1975 населяли моря вдоль южного побережья Лавруссии. Из закавказских видов только Tchanakhtchirostrum araraticum (Abramian, 1957) продолжал существовать в раннем карбоне, но быстро вымер.
ВВЕДЕНИЕ
Для большинства ископаемых и всех современных брахиопод отряда Rhynchonellida характерна непористая раковина. Всего несколько родов за миллионы лет существования отряда, начиная с раннего ордовика, имели пористую раковину. Появление пор в раковинах этих брахиопод совпало с крупной экосистемной перестройкой. Первые представители пористых ринхонеллид появились сразу после девонского вымирания на границе франа и фамена. Наибольшее их разнообразие приходится на фамен (пять родов и 10 видов, не считая спорных и слабо изученных): Greira transcaucasica Erlanger, 1993, Sharovaella mirabilis Pakhnevich, 2012, S. morini (Drot, 1964), Tchanakhtchirostrum araraticum (Abramian, 1957), T. centralis (Sartenaer et Plodowski, 2003), Zaigunrostrum iranicum (Gaetani, 1965), Z. nakhichevanensе Pakhnevich, 2018, Araratella dichotomians (Abramian, 1954) [в т.ч. подвиды A. dichotomians assimulata (Abramian, 1954), A. moresnetensis (de Koninck, 1887) и, возможно, A. dichotomians kazakhstanica (Rozman, 1962)], A. anatolica Sartenaer et Plodowski, 2003.
Помимо этого, к надсемейству Rhynchoporoidea различными авторами относилось еще несколько родов: Rariella Zhang, 1981 и Yingtangella Bai et Ying, 1977 из нижнего девона Южного Китая (Rong, Zhang, 1994), Momarhynchus Baranov et Sartenaer, 1996 из отложений фамена Якутии (Баранов, 1996), Paryphorhynchopora Simorin, 1956 из турнейских отложений Казахстана (Симорин, 1956), Striatorhynchus Pushkin, 1986 из фаменских отложений Белоруссии (Пушкин, 1986), Porostictia Cooper, 1955 из верхнего фамена США (Нью-Мексико) (Cooper, 1955). Но некоторые из них (Rariella, Yingtangella) затем были отнесены к другим отрядам. Для других (Momarhynchus) не было достаточно доказательств для отнесения их к пористым ринхонеллидам, в частности, не проиллюстрировано наличие пор. Поры остальных спорных родов (Paryphorhynchopora, Striatorhynchus, Porostictia) оказались экзопорами, т.е., ямками на наружной поверхности раковины. Экзопористость возникла независимо в разных эволюционных стволах ринхонеллид. Более подробно о спорных родах пористых ринхонеллид можно посмотреть в публикациях (Sartenaer, Plodowski, 2003; Pakhnevich, 2013).
Цель настоящей работы проанализировать динамику биоразнообразия пористых ринхонеллид Закавказья, пути их расселения, центры происхождения, причины возникновения пористости.
К ВОПРОСУ ПРОИСХОЖДЕНИЯ ПОР У RHYNCHONELLIDA
На границе двух веков девонского периода, франского и фаменского, у нескольких родов брахиопод отряда Rhynchonellida появилась необычная особенность в микроструктуре раковины – пористость. Известно семь родов пористых ринхонеллид, пять из которых характерны для фамена. Поры относятся к типу эндопор. Что стало движущим толчком для изменений микроструктуры раковины? Ответ на этот вопрос следует искать в физиологических изменениях у брахиопод, которые произошли в начале фаменского века. Следует рассмотреть версии о функции пор. У современных теребратулид пора прорастает через вторичный и первичный слои раковины и соприкасается с периостракумом с помощью выростов кэки (англ. caeca), которые заканчиваются пептидными трубочками (Williams, 1997). Поскольку у пор есть контакт с наружной водой, и в них заходят клетки наружного эпителия мантии, их может характеризовать полифункциональность. Есть предположение, что поры, а точнее, те ткани, которые в них заходят, участвуют в дыхании (Shumway, 1982), то есть эти ткани можно назвать дыхательными (Peck et al., 1986). Вероятно, клетки эпителия в порах могут запасать некоторые вещества, напр., липиды и полисахариды (Pérez-Huerta et al., 2009). Существует предположение, что пористость раковины отрицательно влияет на обрастателей, поселяющихся на поверхности раковин (Alexander, Scharpf, 1990). Вероятно, защита осуществляется с помощью веществ-репеллентов, которые “отпугивают” личинок. В девоне в различных акваториях планеты обрастатели на раковинах брахиопод были многочисленны (Hoare, Steller, 1967; Thayer, 1974; Пахневич, 2008). Ч. Тейер предполагал, что брахиоподы таким образом могли “отпугивать” и хищников (Thayer, 1974). Еще одна версия о функции пор связана с возможностью их деятельности как органов чувств (Pérez-Huerta et al., 2009). Считается, что эндопоры палеозойских ринхонеллид заканчивались во вторичном слое, и тогда логично было бы предположить, что они выполняли функцию запасания. Но в результате новых исследований (рис. 1) появилось предположение, что у ринхонеллид также были выросты (caeca), которые распространялись в первичный слой и, возможно, подходили к периостракуму. Но первичной слой раковин пористых ринхонеллид не изучен.
Рис. 1.
Следы выростов кэки в поре (обозначены стрелками), обнаруженной в створке Sharovaella mirabilis Pakhnevich, 2012, голотип ПИН, № 3744/804; Нахичеванская АР, гора Геран-Kаласы, обн. 1241; нижний фамен, зона Cyrtospirifer asiaticus–Mesoplica meisteri.
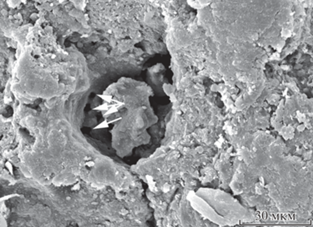
Какие же события могли привести к значительным физиологическим изменениям в различных группах ринхонеллид? С границей франа и фамена связано массовое вымирание, произошедшее в различных группах морской фауны. О его причинах высказывалось несколько версий. На границе двух веков в атмосфере увеличивается содержание углекислого газа. При этом содержание кислорода в придонном слое воды падает. Для данного времени характерно накопление в донных осадках большого количества органического углерода. Одна из возможных причин повышения содержания углерода – снос органического вещества с континентов (Joachimski et al., 2001). В связи с этим на границе франа и фамена встречаются черные углеродистые породы (Buggisch, 1991; Язиков и др., 2013). В фамене в Южной Гондване происходит оледенение (Caputo, 1985). Конец франа – начало фамена характеризуется трансгрессивно-регрессивным морским циклом, который связан с тектоническими процессами на планете (Мизенс, 2003). С этим же связана и активная вулканическая активность, вплоть до конца франа (Маслов и др., 2008). О возможности импактного события на границе франа и фамена могут свидетельствовать иридиевые аномалии (Nicoll, Playford, 1993) и микротектиты (Claeys, Casier, 1994) в породах этого возраста. В девоне на суше продолжается экспансия высших растений. Их разнообразие увеличивается. По молекулярно-генетическим данным, в позднем девоне отмечено шесть событий, связанных с дивергенцией линий наземных растений (Рубан, 2013). Еще в раннем девоне происходит активное почвообразование, формирование углей (Броушкин, Горденко, 2012). В связи с этим в океан поступает больше органического вещества с суши. Таким образом, опираясь на предположение, что поры связаны с дыханием брахиопод, можно предположить, что обеднение кислородом придонного слоя морской воды могло стать определяющим фактором для независимого появления пор в раковинах нескольких групп позднедевонских ринхонеллид. На это могли повлиять и вулканическая активность, и повышение уровня углекислого газа, и увеличившийся снос органики с суши, и падение космического тела. Данная проблема остается одной из самых сложных в эволюции девонских ринхонеллид, и ей необходимо посвятить отдельное исследование.
Если обратиться непосредственно к описанию литологических особенностей отложений последней франской зоны Закавказья Uchtospirifer subarchiaci–Cyphoterorhynchus arpaensis, то выясняется, что верхняя часть зоны сложена черными аргиллитами с маломощными прослоями органогенно-детритовых известняков (Grechishnikova, 2018, c. 852). В вышележащей пачке кварцитов тоже содержатся углистые примазки, а также слои черных аргиллитов, т.e., присутствует много обугленного органического вещества. Первые три пачки фаменских отложений зоны Cyrtospirifer asiaticus–Mesoplica meisteri сложены темно-серыми и черными известняками или аргиллитами (Grechishnikova, 2018, c. 854–855), т.e., отложения вновь богаты обугленной органикой. Из этого можно сделать предположение, что на границе франа и фамена в исследуемом районе в море было недостаточно кислорода из-за большого поступления органики, и появление пористости у ринхонеллид совпадает с этим временным периодом. При этом ринхонеллиды были обитателями нижнего придонного слоя воды, где и накапливалось органическое вещество.
СИСТЕМАТИКА ПОРИСТЫХ РИНХОНЕЛЛИД
Х. Мюр-Вуд (Muir-Wood, 1955) выделила пористых ринхонеллид в отдельное надсемейство Rhynchoporoidea. Для них характерны простые и дихотомирующие эндопоры, располагающиеся в фиброзном слое двух- или трехслойной раковины. На начало 2000-х гг. (Savage, 2002) пористые ринхонеллиды рассматривались в рамках единого надсемейства Rhynchoporoidea. Основным объединяющим признаком данной группы была пористость раковины, хотя положение пор в слоях раковины внутри надсемейства отличалось. Анализируя характер пористости и строение стенки раковины, ряд исследователей (Эрлангер, 1986; Savage, 2002) не придал большoго значения серьезным различиям во внутреннем строении раковин пористых ринхонеллид. B первую очередь они сравнивали их друг с другом по признакам микроструктуры, тогда как по внутреннему строению можно было отметить, что надсемейство состоит из таксонов с различными внутренними морфологическими структурами, собранных вместе по единственному признаку. По внутреннему строению хорошо видно, что объединенные в одно надсемейство ринхонеллиды не принадлежали одной группе. За основу проведенной ревизии было взято сравнение родов по таким признакам, как особенность развития зубных пластин, длина и высота септы в спинной створке, открытый или закрытый септалий, слитная или разделенная замочная пластина, форма круральных оснований и крур. Поэтому после ревизии (Pakhnevich, 2013) данное надсемейство было признано сборным, и сделан вывод, что пористость появилась в разных группах девонских ринхонеллид, семействах Trigonirhynchiidae, Leiorhynchidae, Septalariidae, параллельно.
В настоящее время представляется, что систематическое положение этих брахиопод выглядит следующим образом (Pakhnevich, 2013):
Надсемейство Rhynchotrematoidea Schuchert, 1913, Семейство Trigonirhynchiidae Schmidt, 1965, Подсемейство Rhynchoporinae Muir-Wood, 1955: Род Rhynchopora King, 1865; Подсемейство Greirinae Erlanger, 1993: Роды: Greira Erlanger, 1993, Tchanakhtchirostrum Sartenaer et Plodowski, 2003, Sharovaella Pakhnevich, 2012, Zaigunrostrum Sartenaer et Plodowski, 2003.
Надсемейство Camarotoechioidea Schuchert, 1929, Семейство Leiorhynchidae Stainbrook, 1945, Подсемейство Tretorhynchinae Savage, 2002: Род Tretorhynchia Brunton, 1971; Семейство Septalariidae Havliček, 1960, Подсемейство Araratellinae Erlanger, 1986: Род Araratella Abramian, Plodowski et Sartenaer, 1975.
Из представленных родов в фамене Закавказья встречаются Greira, Tchanakhtchirostrum, Sharovaella, Zaigunrostrum, Araratella. Они представлены видами: G. transcaucasica, S. mirabilis, T. araraticum, Z. nakhichevanensе, A. dichotomians (рис. 2).
Рис. 2.
Пористые ринхонеллиды Закавказья: а–д – Sharovaella mirabilis Pakhnevich, 2012, голотип ПИН, № 3744/804; цельная раковина (×1): а – со стороны брюшной створки, б – со стороны спинной створки, в – со стороны замочного края, г – сбоку, д – со стороны переднего края; Нахичеванская АР, гора Геран-Каласы, обн. 1241; нижний фамен, зона Cyrtospirifer asiaticus–Mesoplica meisteri; е–к – Zaigunrostrum nakhichevanense Pakhnevich, 2018, экз. ПИН, № 3744/861, цельная раковина (×1.5): е – с брюшной створки, ж – со спинной створки, з – сбоку, и – со стороны замочного края, к – со стороны переднего края; левобережье р. Арпы, обн. 20, сл. 6; верхний фамен, зона Cyrtospirifer pamiricus–Enchondrospirifer ghorensis; л–п – Greira transcaucasica Erlanger, 1993, экз. ПИН, № 3744/856, цельная раковина, трехмерная модель, полученная с помощью рентгеновского микротомографа Skyscan 1172 (×1.7): л – с брюшной створки, м – со спинной створки, н – сбоку, о – со стороны замочного края, п – со стороны переднего края; Нахичеванская АР, гора Геран-Каласы, обн. 1241; нижний фамен, зона Cyrtospirifer asiaticus–Mesoplica meisteri; р–ф – Tchanakhtchirostrum araraticum (Abramian, 1957); экз. ПИН, № 3744/855, цельная раковина, трехмерная модель, полученная с помощью рентгеновского микротомографа Skyscan 1172 (×1.5): р – с брюшной створки, с – со спинной створки, т – сбоку, у – со стороны замочного края, ф – со стороны переднего края; Нахичеванская АР, р-н с. Кярки; верхний фамен, зона Sphenospira julii–Spinocarinifera nigra; х – Araratella dichotomians (Abramian, 1954), экз. ПИН, № 3744/864, цельная раковина (×1), вид с брюшной створки; Нахичеванская АР, восточнее горы Геран-Каласы, обн. 10, сл. 3; верхний фамен, зона Sphenospira julii–Spinocarinifera nigra.

ПОЯВЛЕНИЕ И РАСПРОСТРАНЕНИЕ ПОРИСТЫХ РИНХОНЕЛЛИД В ЗАКАВКАЗЬЕ
В отложениях девона Закавказья встречена разнообразная фауна ринхонеллид, начиная с верхов эмса и до границы девона и карбона. Высоко разнообразие ринхонеллид и в фаменских отложениях. В конце девона территория Закавказья находилась в Южном полушарии недалеко от экватора в составе континента Гондваны (Лордкипанидзе и др., 1984) или было небольшим микроконтинентом (Grigoryan et al., 2019). Это было его северное побережье, омываемое водами океана Палеотетис (Torsvik, Cocks, 2011). Рядом с изучаемым районом располагался микроконтинент “Иран–Афганистан” (Gamkrelidze, 1986; Ruban et al., 2007). Сходство фаун брахиопод Ирана, Афганистана и Армении (правда, на родовом уровне) подробно изучено с помощью коэффициента Жаккара (Brock, Yazdi, 2000). Они имеют несомненное сходство, так же как и позднедевонская фауна Турции, микроконтиненты Тауриды [Taurides (Nazik et al., 2018)], которые также располагались географически близко к Закавказью (Ruban et al., 2007, рис. 7, 8 ). Среди общих видов фаменских ринхонеллид Ирана (Jafarian, 2000; Kebria-Ee Zadeh, 2009) и Закавказья (Pakhnevich, 2018) отмечаются Araratella dichotomians (Abramian, 1954) (оба подвида), Paurogastroderhynchus nalivkini (Abramian, 1957), Sartenaerus charakensis (Brice, 1967), а также виды родов Stenaulacorhynchus Sartenaer, 1968 и Leptocaryorhynchus Sartenaer, 1970. Среди общих видов ринхонеллид с фаменом Афганистана почти все те же представители отряда, что известны из фамена Ирана (кроме видов Stenaulacorhynchus), плюс к ним добавляется Paropamisorhynchus kotalensis (Brice, 1970) (Brice, 1967, 1970).
Первый вид пористых ринхонеллид Greira transcaucasica Erlanger, 1993 (семейство Trigonirhynchiidae) отмечается для нижнего фамена зоны Cyrtospirifer asiaticus–Mesoplica meisteri (Эрлангер, 1993) (табл. 1). Для него характерно типичное для тригониринхиид внутреннее строение раковины: хорошо развитые зубные пластины в макушке брюшной створки, септа с закрытым септалием и единая замочная пластина. Вид и род являются эндемиками, поскольку нигде более не встречаются.
Таблица 1.
Стратиграфическая последовательность брахиоподовых зон, в отложениях которых встречаются пористые ринхонеллиды
| Ярус | Брахиоподовые зоны |
|---|---|
| Турнейский | Parallelora praeulbanensis–Rhytiophora curtirostris |
| Фаменский | Sphenospira julii–Spinocarinifera nigra |
| Paurogastroderhynchus nalivkini | |
| Cyrtospirifer pamiricus–Enchondrospirifer ghorensis | |
| Dmitria seminoi | |
| Cyrtospirifer asiaticus–Mesoplica meisteri |
В то же время появляется еще один род, представленный в Закавказье одним видом, Sharovaella mirabilis (семейство Trigonirhynchiidae). Для Sharovaella характерны наличие зубных пластин в макушке брюшной створки, септы с закрытым септалием в спинной створке и уплощенные с боков круральные основания и круры. Не совсем понятно, какой из двух родов, Sharovaella или Greira, появился раньше. Первый характерен для нижнего и верхнего фамена, начиная с зоны Cyrtospirifer asiaticus–Mesoplica meisteri и заканчивая Cyrtospirifer pamiricus–Enchondrospirifer ghorensis. В других частях побережья Северной Гондваны и за ее пределами данный вид неизвестен. Вид S. mirabilis можно считать эндемиком Закавказья, но не род Sharovaella. Другой вид этого рода, S. morini (Drot, 1964), известен из верхнего фамена Марокко, зоны IV (Drot, 1964), что соответствует низам верхнего фамена (Kaiser et al., 2007) близко к зоне Cyrtospirifer pamiricus–Enchondrospirifer ghorensis. Т.е., S. mirabilis появился раньше в море на территории Закавказья и расселился вдоль северного побережья Гондваны в северо-западном направлении. Виды сходны друг с другом, но у S. mirabilis раковина более выпуклая.
В верхнем фамене появляются сразу три рода и три вида пористых ринхонеллид: Zaigunrostrum nakhichevanensе (семейство Trigonirhynchiidae), Araratella dichotomians (семейство Septalariidae), Tchanakhtchirostrum araraticum (семейство Trigonirhynchiidae). Z. nakhichevanensе известен в Закавказье в отложениях зон Cyrtospirifer pamiricus–Enchondrospirifer ghorensis и Sphenospira julii–Spinocarinifera nigra. Для Zaigunrostrum характерно присутствие зубных пластин в макушке брюшной створки, в спинной створке – закрытый септалий, единая изогнутая замочная пластина, треугольные в сечении круральные основания, вытянутые в дорсо-вентральном направлении круры. К роду Zaigunrostrum принадлежит еще вид Z. iranicum, описанный в Северном Иране из отложений, изначально (Gaetani, 1965) относимых к нижнему фамену, зоне Ptychomaletoechia ? deltidialis, но позже – к верхнему девону (Jafarian, 2000). Поскольку он появляется позже зонального для Закавказья вида Paurogastroderhynchus nalivkini Sartenaer, 1970, эти отложения, скорее, соответствуют зоне Sphenospira julii–Spinocarinifera nigra. Т.е., Z. nakhichevanensе является предковым для Z. iranicum. Поскольку территория Ирана располагалась недалеко от исследуемой территории (Gamkrelidze, 1986; Ruban et al., 2007), расселение произошло недалеко. Z. nakhichevanensе отличается от Z. iranicum более уплощенной и вытянутой в ширину раковиной, более вытянутой макушкой брюшной створки, менее глубоким синусом и не очень высоким седлом.
Из верхов фамена Закавказья известен вид T. araraticum (зоны Paurogastroderhynchus nalivkini и Sphenospira julii–Spinocarinifera nigra). Этот вид является эндемиком, но род распространен более широко. Для рода Tchanakhtchirostrum характерно наличие длинных зубных пластин в макушке брюшной створки, закрытый септалий в спинной створке, а также тонкая единая замочная пластина, круральные основания и круры в форме полумесяцев или дуговидные. Интересная морфологическая особенность раковин данного рода – ребра образуются путем дихотомии или интеркаляции. Этим он похож на A. dichotomians, но значительно отличается по внутреннему строению. Вид T. araraticum также характерен для зоны турне, нижнего карбона Parallelora praeulbanensis–Rhytiophora curtirostris. Очень немногие виды ринхонеллид пережили девон – карбоновое массовое вымирание и перешли в турнейский век, но T. araraticum вымирает в начале карбона. Помимо него, к тому же роду относится вид T. centralis, известный из самых верхов девона Центрального Марокко (Sartenaer, Plodowsky, 2003), вероятно, соответствующих зоне Sphenospira julii–Spinocarinifera nigra Закавказья. И вновь, T. araraticum является более ранним видом рода Tchanakhtchirostrum, вероятно, предковым для T. centralis. Как и в случае с S. mirabilis, представители рода расселились вдоль северного побережья Гондваны северо-западнее территории Закавказья.
Гораздо шире расселился по северному побережью Гондваны еще один вид пористых ринхонеллид, A. dichotomians. В Закавказье он характерен для зон Cyrtospirifer pamiricus–Enchondrospirifer ghorensis–Sphenospira julii–Spinocarinifera nigra. Род Araratella отличается некоторыми интересными особенностями внешнего и внутреннего строения раковины. Скульптура сходна с видами рода Tchanakhtchirostrum и представлена дихотомирующими или интеркалирующими складками (у Tchanakhtchirostrum – ребра). Именно поэтому виды T. araraticum и A. dichotomians объединяли в пределах рода Araratella. В брюшной створке зубные пластины короткие неутолщенные, сближенные со стенками макушки, а ближе к ее кончику срастающиеся со стенками. Стенки макушек створок утолщены. В спинной створке есть многолопастный замочный отросток, с зубовидными лопастями. Септалий Y-образный, широкий, открытый. Под септалием в септе образуется расширение и полость. Она формирует ложнозакрытый септалий. Замочные пластины короткие. Ближе к макушке они соединены с септалием. В момент образования полости в септе они разъединяются с септалием. Продолжением септалия является вогнутый к спинной створке выступ, по краям которого формируются V-образные круральные основания, открытые в сторону спинной створки. Круры ближе к макушке в виде удлиненной запятой, а затем удлиняются параллельно боковым краям и становятся дуговидными. Ближе к переднему краю они распрямляются. Замочный отросток – черта, не характерная для остальных девонских пористых ринхонеллид. До нашей публикации (Пахневич, 2015) эта особенность никем не отмечалась. Еще одна своеобразная черта строения раковины – ложно-закрытый септалий. Почти на всем своем протяжении он открыт, но под его дном образуется полость в утолщенной септе, что приводит к заблуждению, что септалий имеет крышечку. В связи с этим вид A. centralis был отнесен (Пахневич, 2015) к роду Tchanakhtchirostrum, поскольку не имел ряд черт внутреннего строения (замочный отросток, ложно-закрытый септалий), характерных для рода Araratella. Интересно, что раковина данного рода трехслойная, в отличие от всех остальных пористых ринхонеллид, что также заставляет рассматривать этот род как стоящий особняком от остальных закавказских родов.
Araratella – единственный представитель семейства Septalariidae (надсемейство Camarotoechioidea), у которого появилась пористая раковина. A. dichotomians характерен для близких по возрасту отложений верхнего девона Ирана и Афганистана, возможно, и Казахстана. В Иране он известен из фамена (Biozone 11 и 12). Судя по нахождению (Kebria-Ee Zadeh, 2009) в комплексе Biozone 11 зонального вида Enchondrospirifer ghorensis Brice, 1971 (табл. 1), эта биозона соответствует закавказской брахиоподовой зоне Cyrtospirifer pamiricus–Enchondrospirifer ghorensis, тем более для Biozone 10 характерен вид Dmitria seminoi (Verneui, 1850), который является зональным для одноименной подстилающей зоны. А в комплексе брахиопод Biozone 12 есть вид Paurogastroderhynchus nalivkini (Abramian, 1957), по видовому названию которого именуется вышележащая закавказская зона. То есть, A. dichotomians появляется в районе современных территорий Ирана и Закавказья одновременно, и определить область его происхождения не представляется возможным. Тем более, эти территории располагались близко.
Также в конце фамена оба подвида A. dichotomians обитали в акватории на территории Афганистана. По возрасту зоны, в которых распространены брахиоподы, соответствуют иранским (Sartenaer, Plodowsky, 2003).
К верхам фамена приурочены и другие виды рода. A. anatolica известен из верхнедевонских отложений Турции [“middle praesulcata zone” (Sartenaer, Plodowsky, 2003)], что, вероятно, соответствует брахиоподовой зоне Sphenospira julii–Spinocarinifera nigra (Гречишникова, Левицкий, 2011). Вид очень похож на A. dichotomians assimulata, но раковина у A. anatolica вытянута в длину, синус у закавказского вида развит слабо и находящаяся в нем дихотомирующая срединная складка крупнее, чем у турецкого вида; есть отличия в скульптуре (Sartenaer, Plodowsky, 2003). A. anatolica имеет более выпуклую раковину (Pakhnevich, 2018).
Род Araratella наиболее широко расселился не только в пределах северной оконечности Гондваны, но также попал в южные акватории Лавруссии. A. moresnetensis внешне очень похож на A. dichotomians, но отличается большим количеством радиальных складок. A. moresnetensis известен из отложений зоны Quasiendothyra kobeitusana Бельгии и Франции, зоны Siphonodella praesulcata Испании, зоны Wocklumeria Германии (Sartenaer, Plodowsky, 2003; Nicollin, Brice, 2004) – примерно в то же время, что в Турции известен A. anatolica. Но отложения, где находят A. dichotomians в Закавказье, Иране, Афганистане, старше – они характерны для зоны Cyrtospirifer pamiricus–Enchondrospirifer ghorensis, которая соответствует конодонтовым зонам: верхней части marginifera и trachytera (Гречишникова, Левицкий, 2011), что в современной стандартной конодонтовой шкале относится к среднему фамену (Becker et al., 2020). То есть, вид A. dichotomians в районе Закавказья и микроконтинента “Иран–Афганистан” появился еще в среднем фамене, но более точный район происхождения определить пока невозможно. Он стал родоначальником еще двух видов, которые обитали на территории современной Турции и у южных берегов Лавруссии. Возможно, он проник даже в акватории континента Казахстан. Из верхнего фамена Каратау описан подвид A. dichotomians kazakhstanica Rozman, 1962 (Розман, 1962), однако его принадлежность к роду Araratella остается неуточненной. В описании поры в стенке раковины не упоминаются, на иллюстрациях – не изображены. У A. dichotomians kazakhstanica септалий открыт и не утолщен, ложно-закрытый септалий не образуется, выступа как продолжения септалия нет, основания крур уплощенные, а круры стержневидные (Розман, 1962, рис. 26 ). Материал, описанный Х.С. Розман, требует дополнительного изучения.
Вероятно, к роду Araratella относится неопределенный до рода и вида экземпляр Rhynchonellacea gen. et sp. indet (Balinski, 1995), с дихотомирующими ребрами, который А. Балински сближал с видами Liorhynchus dichotomians и Camarotoechia (?) araratica. Он описан из фамена юга Польши, при этом в тексте нет никаких указаний на поры. Наличие дихотомирующих ребер или складок не является доказательством принадлежности к роду Araratella.
Почти все пористые ринхонеллиды в исследуемом районе вымирают на границе девона и карбона, кроме T. araraticum. Но и тот просуществовал недолго, исчезнув в турне. Последние его находки – в отложениях зоны Parallelora praeulbanensis–Rhytiophora curtirostris. Также до турнейского века в море на территории современного Ирана доживает вид Z. iranicum. В турне Франции и США (Mottequin et al., 2014) появляется новый род пористых ринхонеллид Rhynchopora, который широко расселяется в морях позднего палеозоя, вымирая только в пермском периоде. Данный род относится к семейству Trigonirhynchiidae. Вероятно, он является продолжением ветви пористых ринхонеллид-тригониринхиид, но какой род предковый для него, неизвестно. В самом конце фамена на территории современной Франции обитает пористая ринхонеллида A. moresnetensis. Но род Araratella не может быть предковым для Rhynchopora, так как значительно отличается по внутреннему строению – у ринхопоры стенки макушек не утолщенные, настоящий закрытый септалий, единая замочная пластина. Возможно, предком Rhynchopora был какой-то закавказский род, расселившийся в конце фамена на южные берега Лавруссии, который еще не найден.
С середины до конца визе в Западной Европе обитает еще один род пористых ринхонеллид из семейства Leiorhynchidae – Tretorhynchia, но он исчезает до серпуховского века. Пористость у данного рода возникла независимо.
ЗАКЛЮЧЕНИЕ
Таким образом, в фамене Закавказья появились первые пористые ринхонеллиды, относящиеся к семейству Trigonirhynchiidae, подсемейству Greirinae. То есть, данную территорию можно считать центром происхождения подсемейства и пористости у ринхонеллид. Причиной появления пористости можно предполагать кислородное обеднение придонного слоя морской воды. Первые пористые ринхонеллиды относятся к родам Greira и Sharovaella. Здесь же пористые тригониринхииды достигли максимального разнообразия. Позже подсемейства Greirinae в данном районе появились и первые пористые представители семейства Septalariidae, подсемейства Araratellinae – род Araratella. Для него характерна очень необычная морфологическая особенность – ложно-закрытый септалий, что отличает его от всех остальных упомянутых родов. Если род Greira является эндемиком Закавказья, то род Sharovaella распространился вдоль северного побережья Гондваны (до территории современного Марокко). Это направление расселения характерно и для другого рода пористых тригониринхиид, Tchanakhtchirostrum. Род Zaigunrostrum не распространился дальше побережий ближайших микроконтинентов, помимо территории современного Закавказья; его представители жили в акватории около микроконтинента “Иран–Афганистан”.
В отличие от тригониринхиид, род Araratella расселился гораздо шире. Вид A. dichotomians известен из ближайших акваторий северного побережья Гондваны (отложения Афганистана, Ирана), а другие виды данного рода заселили не только ближайшие воды (территория современной Турции), но и южные побережья Лавруссии, а возможно, и континента Казахстан. Девон–карбоновое массовое вымирание пережили два рода: Tchanakhtchirostrum и Zaigunrostrum. Они оба известны из акватории современного Закавказья и микроконтинента “Иран – Афганистан”, но оба вымирают в турне. Центр видообразования карбоновых пористых ринхонеллид переместился в моря, омывающие Лавруссию. Новые, карбоновые, пористые ринхонеллиды появились в семействах Leiorhynchidae и Trigonirhynchiidae. Пористость стенки раковины у рода Tretorhynchia возникла независимо от других обладателей этой необычной для ринхонеллид особенности. Связь девонских пористых тригониринхиид Северной Гондваны и космополитного карбон–пермского рода Rhynchopora достоверно не просматривается. За время существования ринхонеллид пористость стенки раковины возникала у них независимо в разных семействах не менее трех раз за всю историю отряда.
Список литературы
Баранов В.В. Отряд Rhynchonellida // Атлас девонских брахиопод Северо-Востока России. М.: Наука, 1996. С. 55–112 (Тр. Палеонтол. ин-та РАН. Т. 266).
Броушкин А.В., Горденко Н.В. Древнейшие сосудистые растения и их влияние на окружающую среду // Ранняя колонизация суши. Серия “Гео-биологические системы в прошлом”. М.: ПИН РАН, 2012. С. 157–176.
Гречишникова И.А., Левицкий Е.С. Опорный разрез фамена и нижнего карбона Геран-Каласи (Нахичеванская АР, Азербайджан) // Стратигр. Геол. корреляция. 2011. Т. 19. № 1. С. 24–46.
Лордкипанидзе М.Б., Адамия Ш.А., Асанидзе Б.З. Эволюция активных окраин океана Тетис (на примере Кавказа) // Докл. 27 Междунар. геол. конгр. Палеоокеанология. Т. 3. М.: Наука, 1984. С. 72–83.
Маслов В.А., Артюшкова О.В., Якупов Р.Р., Мавринская Т.М. Проблемные вопросы стратиграфии нижнего и среднего палеозоя Южного Урала // Геол. сборник ИГ УНЦ РАН. 2008. № 7. С. 193–204.
Мизенс Г.А. Колебания уровня Мирового океана и осадконакопление в девонских глубоководных бассейнах юга Урала // Литосфера, 2003. № 4. С. 43–64.
Пахневич А.В. Обрастатели беломорских брахиопод Hemithyris psittacea // Матер. науч. конф., посвященной 70-летию Беломорской биологической станции МГУ: Сборн. статей. М.: Изд.-во “Гриф и К”, 2008. С. 94–97.
Пахневич А.В. К ревизии пористых ринхонеллид рода Araratella (Rhynchonellida, Brachiopoda) // Палеонтол. журн. 2015. № 4. С. 47–54.
Пушкин В.И. Фаменские ринхонеллиды (Brachiopoda) Белоруссии. Семейства Trigonirhynchiidae, Pugnacidae, Hadrorhynchiidae // Новые и малоизученные виды ископаемых животных и растений Белоруссии. Минск: Наука и техника, 1986. С. 63–96.
Розман Х.С. Стратиграфия и брахиоподы фаменского яруса Мугоджар и смежных районов. М.: Наука, 1962. 196 с. (Тр. Геол. ин-та АН СССР. Т. 50).
Рубан Д.А. Дивергенции линий высших растений в палеозое в свете молекулярно-генетических исследований и глобальные условия их проявления // Палеоботан. временник. Прилож. к “Lethaea rossica”. 2013. Вып. 1. С. 18–20.
Симорин А.М. Стратиграфия и брахиоподы Карагандинского бассейна. Алма-Ата: Изд-во АН Каз. ССР, 1956. 300 с.
Эрлангер О.А. К систематике пористых ринхонеллид (брахиоподы) // Палеонтол. журн. 1986. № 3. С. 52–59.
Эрлангер О.А. Greira gen. nov. – древнейший род пористых брахиопод // Палеонтол. журн. 1993. № 1. С. 118–122.
Язиков А.Ю., Изох Н.Г., Сараев С.В. и др. Новые данные по биостратиграфии и седиментологии верхнедевонских отложений о. Столб (дельта р. Лена) // Геол. и геофизика. 2013. Т. 54. № 8. С. 1013–1027.
Alexander R.R., Scharpf C.D. Epizoans on late Ordovician brachiopods from Southeastern Indiana // Hist. Biol. 1990. V. 4. № 3–4. P. 179–202.
Balinski A. Brachiopods and conodont stratigraphy of the Famennian from the Debnik anticline, southern Poland // Palaeontol. Pol. 1995. V. 54. 88 p.
Becker R.T., Marshall J.E.A., Da Silva A.-C. Chapter 22 – The Devonian Period // Geologic Time Scale 2020. V. 2 / Eds Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg G.M. Amsterdam: Elsevier, 2020. P. 733–810.
Brice D. Deux nouvelles espèces de Rhynchonelloidea dans le Dévonien supèrieur d’Afghanistan central // Ann. Soc. Géol. Nord. 1967. V. 87. № 2. P. 95–105.
Brice D. Étude paléontologique et stratigraphique du Dévonien de ľ Afghanistan. Contribution a la connaissance des brachiopodes et des polypiers rugueux // Notes Mém. Moyen- Orient. 1970. T. 11. 364 p.
Brock G.A., Yazdi M. Palaeobiogeographic affinities of Late Devonian brachiopods from Iran // Rec. West. Austral. Mus. 2000. V. 58. Suppl. P. 321–334.
Buggisch W. The global Frasnian – Famennian “Kellwasser Event” // Geol. Rundschau. 1991. V. 80. № 1. P. 49–72.
Caputo M.V. Late Devonian glaciation in South America // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1985. V. 51. P. 291–317.
Claeys P., Casier J.-G. Microtektite-like impact glass associated with the Frasnian–Famennian boundary mass extinction // Earth Planet. Sci. Lett. 1994. V. 122. P. 303–315.
Cooper G.A. New genera of Middle Paleozoic brachiopods // J. Paleontol. 1955. V. 29. № 1. P. 45–63.
Drot J. Rhynchonelloidea et Spiriferoidea Siluro-Dévoniens du Maroc Pré-Saharien // Notes Mém. Serv. Géol. Maroc. 1964. № 178. 284 p.
Gaetani M. Brachiopods and mollusks from Geirud formation, Member A (Upper Devonian and Tournaisian). The geology of the Upper Djadjerud and Lar Valleys (North Iran). II. Palaeontology // Riv. Ital. Paleontol. 1965. V. 71. № 3. P. 679–770.
Gamkrelidze I.P. Geodynamic evolution of the Caucasus and adjacent areas in Alpine time // Tectonophysics. 1986. V. 127. P. 261–277.
Grechishnikova I.A. Biostratigraphy of the Devonian and Carboniferous of Transcaucasia (Brachiopod Zonal Scale) // Devonian and Carboniferous Brachiopods and Biostratigraphy of Transcaucasia. N.Y., 2018. P. 832–873 (Paleontol. J. V. 52. № 8).
Grigoryan A., Serobyan V., Randon C. et al. A Famennian (Late Devonian) conodont assemblage from brachiopod–rich limestones of the Djravank section (Southern Armenia) // Proc. NAS RA, Earth Sci. 2019. V. 72. № 3. P. 3–12.
Hoare R.D., Steller D.L. A Devonian brachiopod with epifauna // Ohio J. Sci. 1967. V. 67. № 5. P. 291–297.
Jafarian M.A. Late Devonian index Brachiopoda of north-east Esfahan in correlation with other regions // J. Sci. I. R. Iran. 2000. V. 11. № 3. P. 221–231.
Joachimski M.M., Ostertag-Henning C., Pancost R.D. et al. Water column anoxia, enhanced productivity and concomitant changes in δ13C and δ34S across the Frasnian–Famennian boundary (Kowala – Holy Cross Mountains / Poland) // Chem. Geol. 2001. V. 175. P. 109–131.
Kaiser S.I., Becker R.T., El Hassani A. Middle to Late Famennian successions at Ain Jemaa (Moroccan Meseta) – implications for regional correlation, event stratigraphy and synsedimentary tectonics of NW Gondwana // Geol. Soc. Lond. Spec. Publ. 2007. V. 278. P. 237–260.
Kebria-Ee Zadeh M. Biozonation of Geirud Formation in the Eastern Alborz (Damghan Region) based on the Brachiopoda // Sci. Quart. J. Geosci. 2009. V. 18. № 71. P. 19–30. (in Farsi).
Mottequin B., Brice D., Legrand-Blain M. Biostratigraphic significance of brachiopods near the Devonian–Carboniferous boundary // Geol. Mag. 2014. V. 151. № 2. P. 216–228.
Muir-Wood H.M. A history of the classification of the phylum Brachiopoda. L.: Brit. Museum (Natur. History), 1955. 124 p.
Nazik A., Groos-Uffenorde H., Olempska E. et al. Late Silurian and Devonian ostracods of the Istanbul Zone (Western Pontides) and the Taurides: palaeogeographical implications // Palaeobiodiv. Palaeoenv. 2018. V. 98. № 4. P. 593–612. https://doi.org/10.1007/s12549-018-0340-5
Nicoll R.S., Playford P.E. Upper Devonian iridium anomalies, conodont zonation and the Frasnian–Famennian boundary in the Canning Basin, Western Australia // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1993. V. 104. P. 105–113.
Nicollin J.-P., Brice D. Biostratigraphical value of some Strunian (Devonian, uppermost Famennian) Productidina, Rhynchonellida, Spiriferida, Spiriferinida brachiopods // Geobios. 2004. V. 37. P. 437–453.
Pakhnevich A.V. Revision of the Superfamily Rhynchoporoidae Muir-Wood, 1955 // Paleontol. J. 2013. V. 47. № 1. P. 36–43.
Pakhnevich A.V. Order Rhynchonellida // Devonian and Carboniferous Brachiopods and Biostratigraphy of Transcaucasia. N.Y., 2018. P. 892–956 (Paleontol. J. V. 52. № 8).
Peck L.S., Morris D.J., Clarke A., Holmes L.J. Oxygen consumption and nitrogen excretion in the Antarctic brachiopod Liothyrella uva (Jackson, 1912) under simulated winter conditions // J. Exper. Var. Biol. Ecol. 1986. V. 104. Iss. 1–3. P. 203–213.
Pérez-Huerta A., Cusack M., McDonald S. et al. Brachiopod punctae: a complexity in shell biomineralisation // J. Struct. Biol. 2009. V. 167. P. 62–67.
Rong J.-Y., Zhang Y. Rariellidae, a new family of Rhynchoporoidea (Brachiopoda) with a restudy of the type genus Rariella Zhang 1981, from the Emsian (Early Devonian) of Inner Mongolia, north China // Alcheringa. 1994. V. 18. P. 135–146.
Ruban D.A., Al-Husseini M.I., Iwasaki Y. Review of Middle East Paleozoic plate tectonics // GeoArabia. 2007. V. 12. № 3. P. 35–56.
Sartenaer P., Plodowski G. Reassessment of the Strunian genus Araratella Abrahamian, Plodowski & Sartenaer 1975 in the northern Gondwanaland (Rhynchonellida, Brachiopoda) // Cour. Forsch.-Inst. Senckenb. 2003. V. 242. P. 329–348.
Savage N.M. Superfamily Rhynchoporoidea Muir-Wood, 1955 // Treatise on Invertebrate Paleontology. Pt H. Brachiopoda. Vol. 4: Rhynchonelliformea (part). Lawrence: Kansas Univ. Press, 2002. P. 1232–1235.
Shumway S.E. Oxygen consumption in brachiopods and the possible role of punctae // J. Exp. Mar. Biol. Ecol. 1982. V. 58. P. 207–220.
Thayer C.W. Substrate specificity of Devonian Epizoa // J. Paleontol. 1974. V. 48. № 5. P. 881–894.
Torsvik T.H., Cocks L.R.M. The Paleozoic palaeogeography of central Gondwana // Geol. Soc. Lond. Spec. Publ. 2011. V. 357. P. 137–166.
Williams A. Shell structure // Treatise on Invertebrate Paleontology. Pt H. Brachiopoda, Revised, Vol. 1. Introduction / Ed. Kaesler R.L. Boulder (Co)–Lawrence (Ks): Geol. Soc. Amer.–Univ. Kansas Press, 1997. P. 267–320.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал