

Палеонтологический журнал, 2022, № 3, стр. 83-91
Цикадовые в перми Субангариды
А. В. Гоманьков *
Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, Россия
* E-mail: gomankov@mail.ru
Поступила в редакцию 18.02.2021
После доработки 13.12.2021
Принята к публикации 14.12.2021
- EDN: QDYKQX
- DOI: 10.31857/S0031031X22030060
Аннотация
В казанских отложениях Восточно-Европейской платформы обнаружены остатки кладоспермов цикадовых, описанные как Dioonitocarpidium rossicum sp. nov. Эти находки подтверждают присутствие цикадовых в перми Субангарской палеофлористической области, предполагавшееся ранее на основе находок вегетативных листьев. Обсуждается роль рода Dioonitocarpidium как возможной исходной формы для последующей эволюции цикадовых в мезозое и кайнозое.
ВВЕДЕНИЕ
Цикадовые (порядок Cycadales подкласса Cycadopsida класса Gymnospermae) обычно считаются группой, характерной для мезозоя (Мейен, 1987; Thomas, Spicer, 1987; Юрина и др., 2010). Достигая максимального распространения и разнообразия в юре и раннем мелу, они постепенно угасают в кайнозое, и в настоящее время представлены примерно десятью родами, распространенными исключительно в тропиках и субтропиках. О происхождении и истории цикадовых в палеозое известно очень мало. С.В. Мейен (Meyen, 1984) на основании симметрии семян считал их потомками тригонокарповых, однако никаких переходных форм между тригонокарповыми и цикадовыми до сих пор не известно. С. Мамей (Mamay, 1976) в качестве возможных предков цикадовых рассматривал роды Spermopteis Cridland et Morris, Archaeocycas Mamay и Phasmatocycas Mamay, известные из приуралия США. Однако позже Гао Чжифен и Б. Томас (Gao, Thomas, 1989) описали из нижней перми Китая кладоспермы, фактически идентичные кладоспермам современного рода Cycas L. Таким образом, выяснилось, что в приуралии цикадовые существовали уже в “готовом виде” и их предков надо искать в более древних отложениях (Гоманьков, 1993). На роль примитивных цикадовых может претендовать также род Dioonitocarpidium Rühle von Lilienstern, с начала XX в. известный из средне–верхнетриасовых отложений Западной Европы (Pott, 2019). Сравнительно недавно (DiMichel et al., 2001) остатки этого рода были найдены также в приуралии Техаса, что ставит под сомнение представления об эволюции цикадовых не только Мамея, но также Гао и Томаса. Вне зависимости от пути происхождения цикадовых можно, по-видимому, утверждать, что в приуралии они были уже достаточно разнообразной (хотя и редкой) группой. Поэтому их присутствие представляется вполне возможным и для гвадалупия.
В гвадалупской флоре Субангарской палеофлористической области (Meyen, 2002) остатки, которые могли принадлежать цикадовым, были до недавнего времени представлены исключительно листьями (роды Taeniopteris Ad. Brongniart и Guramsania Vachrameev, E. Lebedev et Sodov). Таким образом, присутствие цикадовых в данной флоре, не будучи подтверждено находками генеративных органов, оставалось проблематичным. В 2006 г. в местонахождении Чепаниха в Удмуртии в пограничных казанско-уржумских отложениях мною было найдено два остатка, которые можно отнести к роду Dioonitocarpidium. Еще один остаток, который можно отнести к тому же роду, был обнаружен в коллекции, собранной в Чепанихе Е.И. Улановым в 1970 г. и хранящейся ныне в лаб. палеофлористики Геологического ин-та (ГИН) РАН. Ниже эти остатки описываются как новый вид рода Dioonitocarpidium. Они расширяют наши представления о разнообразии и распространении данного рода, а, следовательно, и всего порядка Cycadales.
При описании остатков принята терминология, предложенная Мейеном (1982) для фруктификаций голосеменных растений, а также использовавшаяся Х. Поттом (Pott, 2019) при описании видов рода Dioonitocarpidium. Обр. № 4036/87, являющийся оригиналом к настоящей статье, хранится в лаб. палеофлористики ГИН РАН (Москва). Остальные оригиналы к статье хранятся в лаб. палеоботаники Ботанического ин-та им. В.Л. Комарова РАН (БИН), С.-Петербург.
Автор выражает признательность сотрудникам лаб. артропод Палеонтологического ин-та им. А.А. Борисяка (ПИН) РАН А.С. Башкуеву, Д.С. Копылову и Д.Е. Щербакову за помощь в сборе материала, положенного в основу настоящей статьи, а также И.А. Игнатьеву и Ю.В. Мосейчик за предоставленную возможность ознакомиться с коллекциями, хранящимися в лаб. палеофлористики ГИН РАН. Работа выполнена в рамках темы госзадания № АААА-А19-119021190031-8 БИН РАН.
МЕСТОНАХОЖДЕНИЕ ЧЕПАНИХА
Местонахождение Чепаниха расположено на правобережье Камы (рис. 1, а) в верховьях оврага, впадающего слева в долину р. Россохи в д. Чепаниха, приблизительно в 1.3 км северо-восточнее центра деревни (56.678652° с.ш., 53.644367° в.д.). Растительные остатки в этом местонахождении более или менее равномерно распределены в слое темно-серого аргиллита видимой мощностью 7.2 м, который перекрывается примерно двадцатиметровой толщей пестроцветных терригенных пород (рис. 1, б) и имеет, вероятно, старичный генезис (Гоманьков, 2008). В одном слое с растительными остатками встречены также остатки остракод, конхострак и насекомых.
Рис. 1.
Географическое положение (а) и разрез (б) местонахождения Чепаниха: 1 – песчаники, 2 – пески, 3 – алевролиты, 4 – аргиллиты, 5 – растительные остатки.
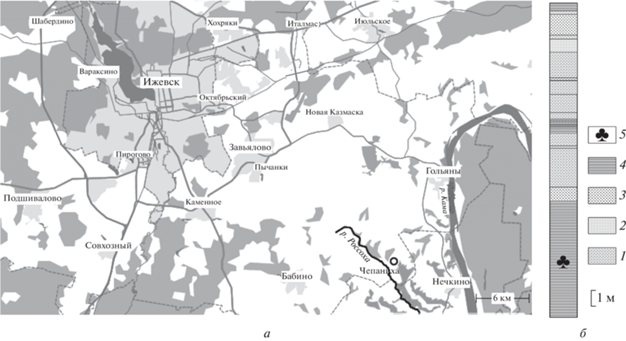
Стратиграфическое положение этого местонахождения подробно обсуждалось мною в недавно опубликованной статье (Gomankov, 2020). Привязка к “старой” стратиграфической шкале пермских отложений Восточно-Европейской платформы (Стратотипический разрез …, 2001) позволяла относить Чепаниху к пограничным казанско-уржумским отложениям, точный возраст которых представлялся не вполне определенным. Однако постановлением Межведомственного стратиграфического комитета от 8 апреля 2005 г. (Постановление …, 2006) эта шкала была существенно реорганизована: ее уржумский горизонт был повышен в ранге до яруса, а его лимитотип был перенесен с р. Вятки в Южное Приуралье и зафиксирован на значительно более высоком стратиграфическом уровне (Молостовская, 2009). При таком “новом” понимании положения казанско-уржумской границы (и, соответственно, объема казанского яруса) местонахождение Чепаниха заведомо должно относиться к верхнеказанскому подъярусу.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
ПОРЯДОК CYCADALES
СЕМЕЙСТВО CYCADACEAE PERSOON, 1807
Род Dioonitocarpidium Rühle von Lilienstern, 1928
Dioonitocarpidium rossicum Gomankov, sp. nov.
Табл. X , фиг. 1–3 (см. вклейку)
Рис. 2.
Общая форма кладоспермов рода Dioonitocarpidium (не в масштабе): а – тип 1 (D. rossicum sp. nov.), б – тип 2 (D. pennaeforme, D. titzei, Crossozamia yochelsonii), в – тип 3 (D. liliensternii), г – тип 4 (филлоспермы, описанные как D. pennaeforme: Kustatscher, van Konijnenburg-van Cittert, 2010).
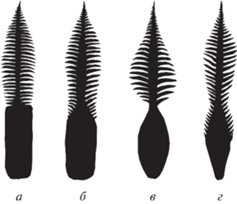
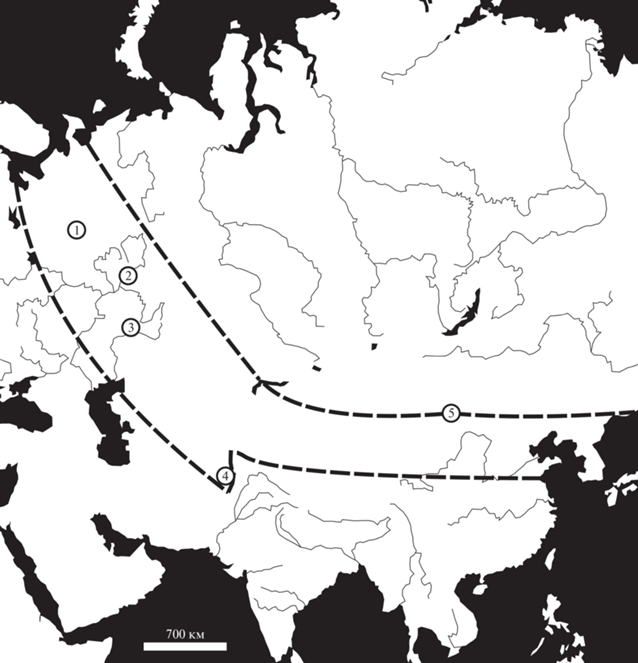
Рис. 3.
Местонахождения листьев, возможно, принадлежавших цикадовым, в пределах Субангарской палеофлористической области. Границы области, обозначенные пунктиром, проведены на основании работ Дуранте, Лувсанцедэн (2002) и Мейена (Meyen, 2002). Обозначения: 1 – Исады, Аристово, 2 – Чепаниха, Костоваты, 3 – Кичкас, 4 – Кабул, 5 – Яман-Ус.
Название вида – от Российской Федерации.
Голотип – БИН РАН, № 1853/243, отпечаток кладосперма; местонахождение Чепаниха; верхнеказанский подъярус; обозначен здесь (табл. X , фиг. 1).
Diagnosis. Fertile part of cladosperm is rectangular and possesses entire margins along the whole length. The central lamina of the sterile part of the cladosperm possesses no more than two longitudinal ridges; its maximum width does not exceed 7 mm. At the junction of the fertile and sterile parts, the central lamina of the sterile part is significantly narrower than the fertile part, so that the total width of the sterile part at its base is equal to the width of the fertile part or even slightly smaller. The maximum width of the cladosperm is in the middle of its sterile part.
Описание (рис. 2, а). Кладоспермы состоят из проксимальной фертильной и дистальной стерильной части. Фертильная часть имеет форму прямоугольника с закругленными углами. Ее ширина составляет около 12 мм, а длина, судя по общим пропорциям сохранившихся фрагментов, была не менее 20 мм. Поверхность кладосперма в его фертильной части почти гладкая; на ней можно заметить лишь очень тонкую штриховку в виде линий, изогнутых в сторону верхушки и веерообразно расходящихся от середины кладосперма к его краям. Семенных рубцов на сохранившихся фрагментах не видно.
Максимальная наблюдавшаяся (неполная) длина стерильной части кладосперма составляет 44 мм. Эта часть кладосперма состоит из центральной пластинки и обрамляющих ее по краям листочков. Центральная пластинка прикрепляется к дистальному краю фертильной части кладосперма (табл. X , фиг. 1). Ее основание (место соединения с фертильной частью) имеет ширину около 7 мм, дальше в сторону верхушки она сужается до 2 мм, а затем, вероятно, снова расширяется и в середине длины может достигать ширины 6 мм. В средней части центральной пластинки наблюдаются два продольных валика (на противоотпечатке, соответственно, желобка) шириной около 1 мм, отстоящих друг от друга примерно на 3 мм (табл. X , фиг. 3). Боковые листочки расположены под углом 40°–80° к продольной оси кладосперма (табл. X , фиг. 1, 2). Они вытянуто-треугольные, иногда слегка изогнутые в сторону верхушки кладосперма или в сторону его основания, с приостренными верхушками. Их ширина составляет около 1 мм. В основании стерильной части кладосперма длина листочков составляет 3–4 мм, так что общая ширина стерильной части кладосперма немного меньше, чем ширина его фертильной части, а в сторону верхушки длина листочков увеличивается до 6 мм, и общая ширина стерильной части оказывается немного больше, чем фертильной. В каждом листочке имеется единственная жилка, прослеживающаяся на всем его протяжении от основания до верхушки.
Сравнение и замечания. В составе рода Dioonitocarpidium к настоящему времени было описано пять видов: D. pennaeforme (Schenk) Rühle von Lilienstern (Schenk, 1864; Rühle von Lilienstern, 1928), D. titzei (Krasser) Pott (Krasser, 1917; Pott, 2019), D. liliensternii Kräusel (Kräusel, 1953), D. keuperianum Kräusel (Kräusel, 1953) и D. moroderi (Leonardi) Kustatscher, Wachtler et van Konijnenburg-van Cittert (Leonardi, 1953; Kustatscher et al., 2004). Вид D. keuperianum был впоследствии справедливо признан младшим синонимом D. liliensternii (Pott, 2019). Кроме того, к роду Dioonitocarpidium, по-видимому, следует относить единственный экземпляр, который был описан под видовым названием Cycadospadix yochelsonii Mamay (Mamay, 1976), но позже Гао и Томасом (Gao, Thomas, 1989) перенесен в род Crossozamia Pomel. Однако по имеющемуся изображению и описанию этого экземпляра трудно определить, действительно ли он представляет самостоятельный вид, или же принадлежит к какому-либо виду рода Dioonitocarpidium, описанному прежде.
От видов D. pennaeforme, D. titzei и D. liliensternii, а также от вида Crossozamia yochelsonii (Mamay) Gao et Thomas описываемый вид D. rossicum sp. nov. отличается формой фертильной части кладоспермов и характером соединения ее со стерильной частью. Среди остатков рода Dioonitocarpidium, описанных и изображенных в литературе, можно выделить несколько типов, различающихся формой фертильной части кладосперма (рис. 2).
Тип 1. Фертильная часть кладосперма имеет прямоугольную форму: ее верхний и боковые края прямые и ровные. Ширина центральной пластинки стерильной части в основании существенно меньше, чем ширина фертильной части, так что переход от фертильной части к стерильной сопровождается резким перегибом и очень ясно выражен. К этому типу относится только описываемый вид D. rossicum sp. nov.
Тип 2. Фертильная часть кладосперма имеет, в общем, прямоугольную форму, но наверху она постепенно сужается, и на краях появляются боковые листочки, сходные с листочками стерильной части. Переход от фертильной к стерильной части кладосперма, таким образом, оказывается постепенным: суженная фертильная часть переходит в центральную пластинку стерильной части. К этому типу относятся кладоспермы, описанные Г. Рюле фон Лилиенштерном как Dioonitocarpidium pennaeforme (Rühle von Lilienstern, 1928, табл. 5 , фиг. 1, 2), Х. Поттом как D. titzei (Pott, 2019, рис. 1, a, c, d) и Мамеем как Cycadospadix yochelsonii (Mamay, 1976, табл. 2 , фиг. 3). Вид D. pennaeforme (типовой вид рода) был описан лишь по стерильным частям кладоспермов (Schenk, 1864). Однако характер листочков в основании стерильной части на экземпляре, представленном на табл. V , фиг. 2 работы А. Шенка (Schenk, 1864), позволяет предположить, что форма фертильной части у этого кладосперма также соответствовала типу 2. Таким образом, можно (хотя и с некоторой долей условности) вслед за Рюле фон Лилиенштерном (автором рода Dioonitocarpidium и комбинации Dioonitocarpidium pennaeforme) считать, что для вида D. pennaeforme были характерны кладоспермы, относящиеся к типу 2. При этом самостоятельность видов Crossozamia yochelsonii и D. titzei (отличие их от D. pennaeforme) вызывает сомнения, а отнесение к виду D. pennaeforme кладоспермов, описанных и изображенных Э. Кустачер и И. ван Конийненбург-ван Циттерт (Kustatscher, van Konijnenburg-van Cittert, 2010) представляется неправомерным.
Тип 3. Фертильная часть кладосперма имеет овальную форму, сужаясь постепенно как к верхушке, так и к основанию; боковые края выпуклые и нигде не несут боковых листочков. Хотя в месте соединения фертильной и стерильной частей ширина фертильной части примерно равна ширине центральной пластинки стерильной части, но переход от фертильной части к стерильной имеет довольно резкий характер, благодаря появлению боковых листочков в основании стерильной части. К этому типу относится голотип вида D. liliensternii (Pott, 2019, рис. 3, a), а также, по-видимому, экземпляры, определенные У. Димайклом с соавт., как Dioonitocarpidium sp. (DiMichele et al. 2001, рис. 6.6, 6.7 ).
Тип 4. Так же, как в типе 3, фертильная часть кладосперма имеет овальную форму с выпуклыми боковыми краями, хотя, возможно, в основании имеет нечто вроде прямоугольного “черешка”. Однако, подобно типу 2, ее суженная верхняя часть несет боковые листочки, благодаря чему переход от фертильной части к стерильной оказывается постепенным, будучи выраженным лишь в уменьшении общей ширины кладосперма. К этому типу относятся кладоспермы, описанные Кустачер и ван Конийненбург-ван Циттерт как D. pennaeforme (Kustatscher, van Konijnenburg-van Cittert, 2010, рис. 7, A, C ), но заслуживающие, вероятно, выделения в отдельный вид.
Таким образом, поскольку описываемый вид оказывается единственным в своем типе, он отличается по форме фертильной части кладоспермов от всех других видов, у которых эта фертильная часть известна.
От вида D. moroderi, известного только по стерильным частям кладоспермов, описываемый вид отличается существенно более узкой центральной пластинкой стерильной части с меньшим числом продольных валиков: у D. moroderi эта центральная пластинка несет четыре валика, а ее максимальная ширина составляет 10 мм (Kustatscher et al., 2004), тогда как у D. rossicum центральная пластинка, по-видимому, не превышает в ширину 7 мм и несет только два валика.
Материал. Три отпечатка кладоспермов (два с противоотпечатками) из типового местонахождения.
НАХОДКИ ЛИСТЬЕВ ЦИКАДОВЫХ В ПЕРМИ СУБАНГАРИДЫ
В пермских отложениях с цикадовыми связывают обычно простые листья типа Taeniopteris. В триасе на основании ассоциации в одних и тех же местонахождениях листья Taeniopteris связываются, по крайней мере, с некоторыми видами Dioonitocarpidium (Kustatscher et al., 2004; Kustatscher, van Konijnenburg-van Cittert, 2010; Pott, 2019). Такие же листья были найдены в Чепанихе (табл. X , фиг. 4).
Рис. 4.
Yavorskia sp., экз. № 1860/195; местонахождение Яман-Ус (Монголия, аймак Умнеговь, окрестности колодца Яман-Ус в 25 км восточнее сомона Номгон); чансиний (?), яманусская свита, песчаниковая толща (см. табл. X , фиг. 6 ).
Находки листьев, которые можно было бы отнести к цикадовым, в перми Субангариды не единичны, хотя и немногочисленны (рис. 3). Мейен (1971) отмечал присутствие рода Taeniopteris в казанских отложениях Южного Приуралья, правда, без точного указания местонахождений. Помимо Чепанихи, листья Taeniopteris sp. (табл. X , фиг. 5) известны из местонахождения Костоваты, которое расположено на берегу Камы в 37 км северо-восточнее Чепанихи и синхронно этому местонахождению (Гоманьков, 2008). К тому же стратиграфическому уровню относится и местонахождение Кичкас в Оренбургской обл., откуда тоже известны листья рода Taeniopteris (Gomankov, 1995, 2020). В азиатской части Субангарской области остатки Taeniopteris sp. известны из позднепермской флоры района Кабула (Meyen, 2002).
Рис. 5.
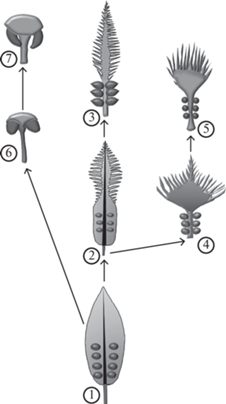
Схема предполагаемого эволюционного развития кладоспермов у цикадовых (кладоспермы и семезачатки показаны не в масштабе): 1 – гипотетическая предковая форма (Archaeocycas?), 2 – Dioonitocarpidium, 3 – Cycas revoluta Thunberg, 4 – Crossozamia chinensis, 5 – Crossozamia cucullata (Halle) Gao et Thomas, 6 – Beania, 7 – Zamia.
К роду Taeniopteris морфологически близки листья, описанные (Гоманьков, Мейен, 1986) из местонахождений Исады и Аристово, которые расположены в бассейне Северной Двины и относятся к вишкильскому и вятскому горизонту или, по новой схеме (Постановление …, 2006), соответственно, к северодвинскому и вятскому ярусам. Для этих листьев, относившихся первоначально к папоротникам, было введено родовое название Fefilopteris Gomankov, однако в дальнейшем выяснилось (Гоманьков и др., 2017), что оно является более поздним синонимом названия Rhabdotaenia Pant, а сами листья должны относиться, скорее, к голосеменным. Для них характерно большое количество одноклеточных волосков, а также сильноизвилистые радиальные стенки эпидермальных клеток. Одноклеточные волоски отмечались у кладоспермов Dioonitocarpidium pennaeforme (Rhüle von Lilienstern, 1928) и D. titzei (Pott, 2019). Возможно также, что отпечатками одноклеточных волосков являются тонкие изогнутые штрихи, наблюдавшиеся в фертильной части кладоспермов D. rossicum sp. nov. С другой стороны, извилистые радиальные стенки эпидермальных клеток характерны больше для беннеттитов, чем для цикадовых. Кладоспермы Dioonitocarpidium чрезвычайно похожи на базальные чешуи “цветков” беннеттитов, отличаясь от них лишь наличием семян (Mamay, 1976). На имеющихся экземплярах D. rossicum следов прикрепления семян также не наблюдается, что может указывать на их принадлежность к беннеттитам, однако, поскольку генеративные органы беннеттитов до сих пор никогда не встречались в перми, автор все же относит описываемые здесь остатки к цикадовым и, соответственно, включает их в род Dioonitocarpidium.
В пермских отложениях центральной Ангариды (Ангарской палеофлористической области) широко распространен род Yavorskia Radczenko (Meyen, 1982). Простоперистые листья этого рода также могли принадлежать цикадовым. Близкие по морфологии листья были найдены на юге Монголии в местонахождении Яман-Ус, располагающемся на границе Ангарской и Субангарской областей. Здесь в отложениях песчаниковой толщи яманусской свиты, имеющих, вероятно, чансинский возраст, был найден один отпечаток простоперистого листа, который можно определить как Yavorskia sp. (табл. X , фиг. 6 ; рис. 4). От типичных представителей рода Yavorskia он отличается длинными черешками перышек, изогнутыми почти под прямым углом к оси перышка и сильно низбегающими на рахис, а также единственной жилкой, входящей в каждый черешок. Вероятно, эти растения заслуживают выделения в отдельный вид, который, однако, не описывается здесь из-за недостаточности материала.
Из более древних отложений того же местонахождения, вероятно, синхронных вишкильскому горизонту, были описаны (Вахрамеев и др., 1986) простоперистые листья Guramsania hosbajari Vachrameev, E. Lebedev et Sodov, также морфологически сходные с родом Yavorskia. Все эти цикадоподобные листья могут рассматриваться как “центрально-ангарские” элементы в пограничном местонахождении Яман-Ус.
DIOONITOCARPIDIUM И ПРОИСХОЖДЕНИЕ ЦИКАДОВЫХ
Со времен работ Мамея (Mamay, 1973, 1976) считалось, что исходным типом кладосперма цикадовых является листоподобный орган с простой цельнокрайней пластинкой и сравнительно длинным черешком, к которому по бокам в два ряда прикрепляются семена. В качестве реализации этого типа рассматривались кладоспермы рода Phasmatocycas, известные из приуралия США. Однако в дальнейшем было показано (Gillespie, Pfefferkorn, 1986), что семена на кладоспермах Phasmatocycas прикреплялись не к черешку, а к средней жилке на абаксиальной стороне простого листоподобного органа, сходного с листьями рода Taeniopteris. Кроме того, в приуралии Китая были обнаружены кладоспермы рода Crossozamia, морфологически почти не отличимые от кладоспермов современного рода Cycas (Gao, Thomas, 1989). На основании этого Гао и Томас исключали Phasmatocycas из числа предковых форм цикадовых и в качестве исходной формы для эволюции кладоспермов этой группы голосеменных рассматривали род Crossozamia. Однако происхождение самого этого рода (весьма “продвинутого” в эволюционном смысле, если иметь в виду его морфологию) не обсуждалось. Также в эволюционной схеме цикадовых, предлагавшейся Гао и Томасом, не находилось места для рода Dioonitocarpidium, который выглядел “примитивнее”, чем Crossozamia, но имел более молодой (триасовый) возраст. Обнаружение остатков рода Dioonitocarpidium в пермских отложениях позволяет по-новому взглянуть на ранние стадии эволюции цикадовых.
Имея столь же древний возраст, как и кладоспермы Crossozamia, кладоспермы Dioonitocarpidium могут претендовать на роль исходной формы для эволюции цикадовых женских фруктификаций. С другой стороны, будучи более “листоподобными”, чем Crossozamia, кладоспермы Dioonitocarpidium, по-видимому, лучше подходят для этой роли, т.к. считается, что общая эволюция в цикадопсидной группе голосеменных шла в направлении все большей специализации кладоспермов, которые изначально мало чем отличались от вегетативных листьев (Meyen, 1984).
Стерильные части кладоспермов Crossozamia и Dioonitocarpidium сходны друг с другом в отношении расчленения на узкие линейные листочки; отличие же заключается в том, что у кладоспермов Dioonitocarpidium это расчленение имеет перистый характер, а у кладоспермов Crossozamia – скорее пальчатый. Однако у многих видов рода Crossozamia стерильная пластинка кладосперма имела ромбическую форму, и ее дистальная часть, суживающаяся к верхушке и несущая листочки на сходящихся боковых краях, вполне может быть сопоставлена с перисто рассеченной дистальной частью кладоспермов Dioonitocarpidium. А у видов C. chinensis (Zhu et Du) Gao et Thomas и C. minor Gao et Thomas, обладавших веерообразными кладоспермами, их дистальный край, помимо обычных листочков, нес еще центральный вырост, который, в свою очередь, делился перисто. Таким образом, стерильная пластинка кладоспермов Crossozamia может быть просто выведена из стерильной части кладоспермов Dioonitocarpidium путем расширения ее в основании.
Стерильные части кладоспермов у рода Cycas, так же как у рода Dioonitocarpidium, расчленены перисто. B этом отношении они ближе к Dioonitocarpidium, чем к Crossozamia, и могут быть выведены из кладоспермов Dioonitocarpidium путем редукции листовой пластинки в их фертильной части.
В итоге можно представить себе следующую картину эволюции кладоспермов у цикадовых (рис. 5). Исходные формы были Taeniopteris-подобными с семенами, прикреплявшимися к цельной листовой пластинке в ее базальной части двумя рядами вдоль средней жилки. Морфологически близкими к этой гипотетической форме, возможно, были кладоспермы Archaeocycas, описанные Мамеем (Mamay, 1973, 1976) из приуралия Северного Техаса, хотя позже сохранность оригинального материала этого рода была признана недостаточной для того, чтобы адекватно судить о характере прикрепления семян к пластинке кладосперма (Axsmith et al., 2003). В ходе дальнейшей эволюции дистальная часть пластинки стала перисто рассеченной, в результате чего возникли кладоспермы, известные как Dioonitocarpidium. Кстати, наиболее древние представители этого рода, описанные как Crossozamia yochelsonii (Mamay, 1976; Gao, Thomas, 1989) и Dioonitocarpidium sp. (DiMichele et al., 2001), происходят из тех же нижнепермских отложений Северного Техаса, что и род Archaeocycas. Эволюционный переход от цельнокрайней листовой пластинки к простоперистой происходил, вероятно, и в вегетативной сфере ранних цикадовых, т.к. среди древнейших листьев, связывающихся с цикадовыми, преобладают простые листья рода Taeniopteris. Последующая редукция пластинки в фертильной части кладоспермов Dioonitocarpidium привела к образованию кладоспермов такого типа, как у Cycas revoluta Thunberg, – простоперистых с семенами, прикрепляющимися к черешку. Параллельно с редукцией фертильной пластинки преобразование простоперистой стерильной части в пальчато рассеченную привело к появлению кладоспермов Crossozamia. Кладоспермы цикадовых с нерассеченной дистальной частью могли возникнуть через промежуточные формы, сходные с родом Beania Carruthers. Их можно вывести непосредственно из гипотетической предковой формы с цельной листовой пластинкой или из кладоспермов Crossozamia, как предполагалось Гао и Томасом (Gao, Thomas, 1989).
Список литературы
Вахрамеев В.А., Лебедев Е.Л., Содов Ж. Цикадовое (?) Guramsania gen. nov. из верхней перми Южной Монголии // Палеонтол. журн. 1986. № 3. С. 103–108.
Гоманьков А.В. Происхождение цикадовых: гипотезы и находки // Природа. 1993. № 4. С. 100–101.
Гоманьков А.В. Татарские пельтаспермовые Русской платформы: морфология, экология, эволюция // Вопросы палеофлористики и систематики ископаемых растений. СПб.: БИН РАН, 2008. С. 42–60 (Чтения памяти А. Н. Криштофовича. Вып. 6).
Гоманьков А.В., Кюнтцель М. К., Мейен С.В. Новые данные о растительных остатках из верхней перми и нижнего триаса Русской платформы // Lethaea rossica. Российский палеобот. журн. 2017. Т. 14. С. 28–63.
Гоманьков А.В., Мейен С.В. Татариновая флора (состав и распространение в поздней перми Евразии). М.: Наука, 1986. 174 с. (Тр. Геол. ин-та АН СССР. Вып. 401).
Дуранте М.В., Лувсанцэдэн У. Особенности развития южной ветви Субангарской палеофлористической области // Сб. памяти чл.-корр. АН СССР, проф. Всеволода Андреевича Вахрамеева (к 90-летию со дня рождения). М.: ГЕОС, 2002. С. 247–261.
Мейен С.В. Пермские флоры Русской платформы и Приуралья // Современные проблемы палеонтологии. М.: Наука, 1971. С. 294–308 (Тр. Палеонтол. ин-та АН СССР. Т. 130).
Мейен С.В. Органы размножения голосеменных и их эволюция (по палеоботаническим данным) // Журн. общ. биол. 1982. Т. 43. № 3. С. 303–323.
Мейен СВ. Основы палеоботаники. М.: Недра, 1987. 403 стр.
Молостовская И.И. Уржумский ярус и его лимитотип // Недра Поволжья и Прикаспия. 2009. Вып. 59. С. 40–45.
Постановление о модернизации верхнего отдела пермской системы Общей (Восточно-Европейской) стратиграфической шкалы // Постановления Межвед. Стратигр. комитета и его постоянных комиссий. 2006. Вып. 36. С. 14–16.
Стратотипический разрез татарского яруса на р. Вятке. М.: ГЕОС, 2001. 140 с.
Юрина А.Л., Орлова О.А., Ростовцева Ю.И. Палеоботаника. Высшие растения. М.: Изд-во МГУ, 2009. 222 с.
Axsmith B.J., Serbet R., Krings M. et al. The enigmatic Paleozoic plants Spermopteris and Phasmatocycas reconsidered // Amer. J. Bot. 2003. V. 90. № 11. P. 1585–1595.
DiMichele W.A., Mamay S.H., Chaney D.S. et al. An Early Permian flora with Late Permian and Mesozoic affinities from North-Central Texas // J. Paleontol. 2001. V. 75. № 2. P. 449–460.
Gao Zhifeng, Thomas B.A. A review of fossil cycad megasporophylls with new evidence of Crossozamia Pomel and its associated leaves from the Lower Permian of Taiyuan, China // Rev. Palaeobot. Palynol. 1989. V. 60. P. 205–223.
Gillespie W.H., Pfefferkorn H.W. Taeniopterid lamina on Phasmatocycas megasporophylls (Cycadales) from the Lower Permian of Kansas, U.S.A. // Rev. Palaeobot. Palynol. 1986. V. 49. № 1–2. P. 99–116.
Gomankov A.V. Kitchkas flora from the Lower Tatarian of the Southern Urals // Paleontol. J. 1995. V. 29. № 2A. P. 81–104.
Gomankov A.V. Flora of the Kazanian–Urzhumian boundary in the middle Permian of the Russian Platform // Palaeoworld. 2020. V. 29. P. 257–269.
Krasser F. Studien über die fertile Region der Cycadophyten aus den Lunzer-Schichten: Mikrosporophylle und männliche Zapfen // Denkschr. Kais. Akad. Wiss. Wien, Math.-Naturwiss. Kl. 1917. Bd 94. S. 489–553.
Kräusel R. Ein neues Dioonitocarpidium aus der Trias von Lunz // Senckenbergiana. 1953. Bd 34. S. 105–108.
Kustatscher E., van Konijnenburg-van Cittert J.H.A. Seed ferns and Cycadophytes from the Triassic Flora of Thale (Germany) // N. Jb. Geol. Paläontol. Abhandl. 2010. Bd 258. Hft. 2. S. 195–217.
Kustatscher E., Wachtler M., van Konijnenburg-van Cittert J.H.A. A number of additional and revised taxa from the Ladinian flora of the Dolomites, Northern Italy // GeoAlp. 2004. V. 1. P. 57–69.
Leonardi P. Flora continentale ladinica delle Dolomiti // Mem. Sci. Geol. Univ. Padova. 1953. V. 18. P. 1–22.
Mamay S.H. Archaeocycas and Phasmatocycas – new genera of Permian cycads // J. Res. U.S. Geol. Surv. 1973. V. 1. № 6. P. 687–689.
Mamay S.H. Paleozoic origin of the cycads // Prof. Pap. U. S. Geol. Surv. 1976. № 934. P. 1–48.
Meyen S.V. The Carboniferous and Permian floras of Angaraland (a synthesis) // Biol. mem. 1982. V. 7. № 1. P. 1–109.
Meyen S.V. Basic features of gymnosperm systematics and phylogeny as shown by the fossil record // Bot. Rev. 1984. V. 50. № 1. P. 1–111.
Meyen S.V. On the Subangara palaeofloristic area of the Permian // Сб. памяти чл.-корр. АН СССР, проф. Всеволода Андреевича Вахрамеева (к 90-летию со дня рождения). М.: ГЕОС, 2002. С. 232–246.
Pott C. The cycadalean megasporophyll Dioonitocarpidium in the Carnian (Late Triassic) flora of Lunz am See, Austria // Paläontol. Z. 2019. V. 93. Iss. 3 (spec. issue). P. 517–530.
Rühle von Lilienstern H. “Dioonites pennaeformis Schenk”, eine fertile Cycadee aus der Lettenkohle // Paläontol. Z. 1928. Bd 10. S. 91–107.
Schenk A. Beiträge zur Flora des Keupers und der rhätischen Formation // Ber. Naturforsch. Ges. Bamberg. 1864. Bd 7. S. 51–142.
Thomas B.A., Spicer R.A. The Evolution and Palaeobiology of Land Plants. L.–Sydney: Croom Helm, 1987. 309 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал