

Палеонтологический журнал, 2021, № 5, стр. 98-106
Миоценовые пищухи рода Bellatona (Lagomorpha, Mammalia) из Долины Озер, Центральная Монголия
М. А. Ербаева a, *, Б. Баярмаа b
a Геологический институт Сибирского отделения РАН
Улан-Удэ, Россия
b Институт палеонтологии Монгольской академии наук
Улаанбаатар, Монголия
* E-mail: erbajeva@ginst.ru
Поступила в редакцию 03.07.2020
После доработки 01.09.2020
Принята к публикации 01.09.2020
Аннотация
Изучены миоценовые пищуховые из отложений свиты Лу Долины Озер Центральной Монголии. Выявлено присутствие представителей трех видов рода Bellatona (B. forsythmajori Dawson, 1961, B. yanghuensis Zhou, 1988, B. kazakhstanica Erbajeva, 1988). Приводится описание и сравнение их с номинативным видом, дано стратиграфическое и географическое распространение и прослежено эволюционное развитие рода Bellatona.
ВВЕДЕНИЕ
Род Bellatona описан впервые из Тайрум Нор Китая в 1961 г. М. Доусон по материалам Центрально-Азиатской экспедиции Американского музея естественной истории (AMNH) из отложений свиты Тунггур, без точного указания горизонта находок. Указано, что возрастом осадков геологи считали поздний миоцен (Dawson, 1961, c. 6). Фауна пищуховых этого местонахождения включала в себя две формы, принадлежавшие новым родам – Bellatona и Alloptox. К новому роду Alloptox был отнесен вид Ochotona gobiensis, описанный ранее Ч. Яном (Young, 1932). Вторая форма описана как новый род Bellatona и новый вид B. forsythmajori Dawson, 1961. Изучение богатого материала позволило Доусон обнаружить значительные вариации в размерах и строении зубов рода Bellatona, в частности, в строении p3, и установить, что, несмотря на существующую вариабельность, весь материал принадлежит одному виду (Dawson, 1961, с. 14). В связи с отсутствием в Азии таксонов раннего миоцена, которых можно было бы отнести к предковым формам рода Bellatona, Доусон предположила, что таковым может быть позднеолигоценовый род Sinolagomys, представители которого обладают рядом прогрессивных черт в строении зубов. Дальнейшие детальные морфологические исследования позволили ей обнаружить сходство родов Bellatona и Ochotona, с преобладанием примитивных признаков у первого, и установить, что Bellatona принадлежит эволюционной линии, ведущей к формированию рода Ochotona (Dawson, 1961, с. 13).
Позднее остатки B. forsythmajori были найдены М.Б. Борисоглебской и В.И. Жегалло в Монголии во время совместных работ отрядов Советско-Монгольской палеонтологической и геологической экспедиций в 1970–1972 гг. Они обнаружены в осадках формации Лу детально стратифицированного разреза Улан-Тологой, возраст которого был определен как средний миоцен. Материал был представлен только одним фрагментом нижнечелюстной кости с p3-m3 (Ербаева, 1981).
Новый вид B. yanghuensis Zhou, 1988 был описан из раннемиоценового местонахождения Янху в провинции Шанси, Китай (Zhou, 1988). Отмечено, что эта форма в целом близка к Bellatona forsythmajori из Тунггура, но отличается от нее рядом примитивных признаков, и происходит она из отложений более раннего возраста, который определен как орлеаний (Orleanian), а возраст номинативной формы – астараций (Astaracian). Автор предположил, что Bellatona yanghuensis возможно является предковой формой B. forsythmajori.
Остатки пищух рода Bellatona были установлены в местонахождениях нижнего миоцена Зайсанской впадины Казахстана. Изучение богатого материала показало, что они принадлежат новому виду B. kazakhstanica Erbajeva, 1988 (Ербаева, 1988, 1994). Этот таксон обладал более примитивными признаками, чем B. forsythmajori и B. yanghuensis, и является наиболее ранним представителем рода.
Детальные палеонтологические исследования, проведенные З. Чу (Qiu, 1996) в новых местонахождениях (Moergen II, Moergen V, Mandelin Chaba II) района Тунггур позволили выявить богатую фауну пищуховых, в том числе – Bellatona forsythmajori. В этих местонахождениях, как и в Тайрум Нор, у B. forsythmajori прослежена значительная изменчивость в строении p3 (Qiu, 1996, рис. 72, 73 ). Сравнительно-морфологический анализ материала позволил автору предположить, что предковой формой Bellatona является род Bohlinotona.
Новые сведения о находке в Китае формы, близкой к Bellatona forsythmajori (B. cf. B. forsythmajori) были представлены в работе Ж. Жанг и др. (Zhang et al., 2012). В местонахождении Дамяо (Damiao), Внутренняя Монголия, коллективом исследователей обнаружен богатый материал, представленный преимущественно изолированными зубами. Авторы отмечают значительную вариабельность в строении p3, что позволило им установить в танатоценозе Дамяо присутствие представителей трех родов: Bellatona, Bellatonoides и Ochotona. Первый род включает в себя Bellatona cf. B. forsythmajori, p3 которого сильно варьирует: имеется p3, сходный с таковым из местонахождения Тайрум Нор (Zhang et al., 2012, рис. 2 A), и дополнительно обнаружен другой p3 с необычным морфотипом, для которого характерна глубокая передненаружная складка, заполненная обильным цементом (Zhang et al., 2012, рис. 2 B, C). Следующий род, Bellatonoides, представлен видом B. eroli Sen, 2003 (Zhang et al., 2012, рис. 2 D-F), известным из позднего миоцена Турции (Sen, 2003); наконец, третий род, Ochotona, судя по размерам зубов, включает в себя два вида – O. lagreli Schlosser, 1924 (Zhang et al., 2012, рис. 2 G, H, K) и O. minor (Bohlin, 1942) (Zhang et al., 2012, рис. 2 I, J).
Впервые богатый материал, включающий в себя все три вида рода Bellatona, был собран участниками Австрийско-Монгольских проектов во время полевых работ с 1995 по 2012 гг. Известны они из девяти местонахождений нижнего и среднего миоцена. Наиболее многочисленны остатки Bellatona yanghuensis (пять местонахождений), B. kazakhstanica представлен в четырех местонахождениях, и B. forsythmajori известен только из одного. Исследования миоценовых пищух из местонахождений Монголии являются целью данной работы.
Детальный сравнительно-морфологический анализ пищуховых трех родов из местонахождений Китая (Qiu, 1996; Zhang et al., 2012), отмеченных выше, позволяет проследить значительные изменения в структуре жевательной поверхности зубов слагающих видов и представить последовательное эволюционное развитие единой линии пищух родов Bellatona, Bellatonoides и Ochotona. Из них наиболее архаичной формой является род Bellatona, отдельные представители которого могли постепенно трансформироваться в род Bellatonoides. Дальнейшее развитие последнего, очевидно, привело к формированию современного рода Ochotona. Это прослеживается на материалах из местонахождения Дамяо, возраст которого 12 млн лет. Здесь в одном танатоценозе установлено присутствие Bellatona cf. B. forsythmajori, представленного прогрессивной формой рода Bellatona, а также вида Bellatonoides eroli Sen, который считается промежуточным звеном в линии, ведущей к формированию рода Ochotona. В танатоценозе присутствует O. cf. O. lagreli, для которого характерны типичные признаки рода Ochotona – строение p3 и наличие хорошо развитого талона на M2. Находка в танатоценозе трех таксонов, вероятно, единой линии позволяет предполагать значительную морфологическую изменчивость при достаточно высокой скорости эволюционного развития этой группы пищуховых на протяжении раннего и среднего миоцена.
Высокая скорость эволюционного развития была обусловлена, вероятно, существованием благоприятных условий для процветания этой группы животных. В начале миоцена, отмечает В.М. Синицын (1965), наблюдалась общая аридизация климата Евразии, произошло расширение засушливых областей, сформировались разнообразные степи. Это подтверждается исследованиями М. Харцхаузера и др. (Harzhauser et al., 2016, с. 3) в Центральной Азии, где начало миоцена совпало с “новым пульсом аридизации и широким распространением пустынных формаций”. Это было связано с позднеолигоценовым похолоданием, продолжавшимся на протяжении транзитного олигоцен-миоценового интервала и оказавшего существенное влияние на изменение биоты раннего миоцена (Harzhauser et al., 2017).
В начале неогена в составе фауны зайцеобразных Центральной Азии полностью исчезли олигоценовые роды Desmatolagus и Bohlinotona, сократилось количество представителей зайцевых, и появились новые роды Amphilagus, Alloptox и Bellatona. Из олигоценовых зайцеобразных сохранился лишь род Sinolagomys, представленный прогрессивными видами S. pachygnathus и S. ulungurensis, полностью утратившими корни зубов; они завершили свое существование в конце раннего миоцена. Род Bellatona дожил, вероятно, до конца среднего миоцена, т.к. остатки его в отложениях позднего миоцена не известны. Можно считать вероятным, что Bellatona был вытеснен прогрессивными формами родов Ochotona и Ochotonoides, получившими расцвет в позднем миоцене и населявшими обширную территорию от Китая и Монголии до Казахстана, включительно.
МАТЕРИАЛ И МЕТОДЫ
Исследованный палеонтологический материал происходит из отложений свиты Лу, вскрывающихся в ряде миоценовых местонахождений Долины Озер Центральной Монголии (Daxner-Höck et al., 2017). Собран он участниками Австрийско-Монгольской экспедиции в рамках проектов FWF, P-10505-GEO, P-15724-N06 и P‑23061-N19 (рук. G. Daxner-Höck) методом промывки породы, содержащей костные остатки, однако редкие экземпляры последних известны из поверхностных сборов. Материал хранится в Венском музее естественной истории (NHMW, Австрия), представлен он фрагментами верхне- и нижнечелюстных костей и изолированными зубами. Типовые экземпляры Bellatona forsythmajori Dawson, 1961 из Тайрум Нор, хранящиеся в коллекционном фонде AMNH, были изучены первым автором. Промеры зубов проводились по общепринятым стандартным методикам, даны в мм и приводятся в таблицах. Названия местонахождений, их сокращения и возраст приведены в разделе текста “Материал”. Терминология зубов дана по Н. Лопес-Мартинес (Lopez Martinez, 1989).
Авторы выражают глубокую благодарность Г. Дакснер-Хёк и У. Гёлих (NHMW) за неоценимую помощь и консультации. М. Ербаева благодарит профессоров Р. Тедфорда† и Л. Маркуса† за предоставленную возможность работать с коллекциями в AMNH. Авторы признательны рецензентам за ценные замечания.
Сокращения, приведенные в тексте и таблицах: ПИН – Палеонтологический ин-т им. А.А. Борисяка РАН, Москва, Россия; AMNH – Американский музей естественной истории, Нью Йорк, США; IVPP – Ин-т палеонтологии и палеоантропологии АН Китая, Пекин, Китай; NHMW – Венский музей естественной истории, Вена, Австрия; P–M – премоляры и моляры верхних зубов; p–m – премоляры и моляры нижних зубов; n – число изученных образцов; m – среднее значение промеров; min, max – минимальное и максимальное значение промеров; sd – стандартное отклонение; L, W – длина и ширина зубов; trig L – длина тригонида; trig W – ширина тригонида; tal L – длина талонида; tal W – ширина талонида; a-loph – антеролоф; hyp – гипострия.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
КЛАСС MAMMALIA
ОТРЯД LAGOMORPHA
СЕМЕЙСТВО OCHOTONIDAE THOMAS, 1897
ПОДСЕМЕЙСТВО SINOLAGOMYINAE GUREEV, 1960
Род Bellatona Dawson, 1961
Bellatona: Dawson, 1961, с. 8.
Типовой вид – Bellatona forsythmajori Dawson, 1961; Китай, Тунггур; вторая половина среднего миоцена (астараций).
Диагноз (по Dawson, 1961, с. 8–9; Ербаева, 1988, с. 58; Zhou, 1988, с. 149). Пищухи мелких и средних размеров, корни на зубах отсутствуют. P2 овальной формы, передняя входящая складка одна или отсутствует. На P3 антеролоф проходит до середины ширины зуба, парафлекса умеренной глубины. На P4–M2 глубина гипострии варьирует, заполнена она цементом; p3 треугольной формы, одна задненаружная входящая складка, глубина которой менее ширины зуба, заполнена цементом. На p4–m2 тригонид незначительно шире талонида; m3 мелкий, овальной формы.
Видовой состав. B. forsythmajori Dawson, 1961, Тайрум Нор (Tairum Nor), Тунггур (Tunggur), Китай, вторая половина среднего миоцена; B. yanghuensis Zhou, 1988, Янху (Yanghu), Синьчжо (Xinzhou), Шанси (Shanxi), Китай, вторая половина раннего миоцена; В. kazakhstanica Erbajeva, 1988, Ашутас, Зайсанская впадина, Казахстан, ранний миоцен.
Bellatona forsythmajori Dawson, 1961
Bellatona forsythmajori: Dawson, 1961, с. 8, рис. 3–7 ; Ербаева, 1988, с. 58, рис. 13–10 .
Bellatona cf. forsythmajori: Erbajeva, Daxner-Höck, 2014, с. 239.
Голотип – AMNH, № 26770, фрагмент правой нижнечелюстной кости с p3-m1; Китай, Тунггур, Тайрум Нор; вторая половина среднего миоцена.
Описание (рис. 1). Пищухи средних размеров, P2 овальной формы с одной передней входящей складкой, заполненной небольшим количеством цемента. На P3 (рис. 1, а) антеролоф доходит до середины ширины зуба; парафлекса начинается и заканчивается на уровне 1/3 ширины зуба; гипострия короткая, заполнена небольшим количеством цемента. Наружные края зуба слабо заостренные, внутренний – выпрямленный. Эмаль хорошо развита на переднем и внутреннем краях зуба. На P4–M2 гипострия глубокая, на P4 (рис. 1, б) достигает 2/3 ширины зуба, на M1 и M2 она проходят практически до наружного края зуба. На заднем крае M2 у экз. из Тайрум Нор (№ 26242; Dawson, 1961, рис. 3) имеется небольшой выступ – слабо развитый талон. Последний достаточно хорошо развит у экз. из Мёрген V (Qiu, 1996, рис. 72 E ).
Рис. 1.
Bellatona forsyhtmajori Dawson, 1961, зубы, вид со стороны жевательной поверхности: а – правый P3 (NHMW 2013/0378/0003); б – правый P4 (NHMW 2013/0378/0006); в – левый p3 (NHMW 2013/0378/0002); г – левый p4 (NHMW 2013/0378/0004); Улан-Толгой (UTO-A/6).
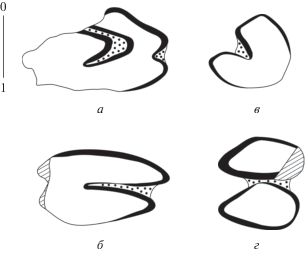
Зуб p3 имеет треугольные очертания со слабо заостренной вершиной; одна задне-наружная входящая складка, заполненная цементом, достигает менее ширины зуба (рис. 1, в). Тригонид с выпрямленным наружным краем, талонид слабо заостренный. Внутренний край p3 слабо закругленный. Эмаль развита по всему периметру зуба неравномерно, иногда отсутствует на задневнутреннем крае. Тригониды и талониды p4–m2 (рис. 1, г) соединены цементом, тригонид несколько шире талонида.
Размеры. См. табл. 1.
Таблица 1.
Промеры зубов Bellatona forsythmajori Dawson
| Длина | Ширина | ||||
|---|---|---|---|---|---|
| min-max | m | n | min-max | m | |
| P3 | 1.5 | 1 | 2.75 | ||
| P4 | 1.4–1.5 | 1.45 | 2 | 2.25 | |
| M1 | 1.25–1.4 | 1.32 | 2 | 2.1–2.25 | 2.17 |
| p3 | 1.25–1.35 | 1.3 | 2 | 1.5–1.6 | 1.55 |
| p4 | 1.6 | 1 | trig. | 1.6 | |
| tal. | 1.5 | ||||
| m2 | 1.65 | 1 | trig. | 1.75 | |
| tal. | 1.65 | ||||
| m3 | 1.0 | 1 | 1.1 | ||
Сравнение. По размерам и строению p3 B. forsythmajori из Долины Озер сходна с типовым материалом из Тайрум Нор, зуб треугольной формы с заостренной вершиной. Однако, они незначительно различаются: p3 пищухи из Монголии треугольной формы с заостренной вершиной, тригонид его с выпрямленным наружным краем. У формы из Тайрум Нор у некоторых экземпляров (AMNH №№ 26236, 26762) на внешней стороне тригонида p3 имеется слабая выемка, у отдельных особей эта выемка углубляется и появляется дополнительная передненаружная складка без цемента (AMNH № 26765) или с цементом (AMNH № 26766). Сходный тип строения p3 прослежен у B. cf. B. forsythmajori из местонахождения Дамяо (Damiao), Уланьхуа (Wulanhua), Внутренняя Монголия, Китай (Zhang et al., 2012, рис. 2 A) и у экземпляров p3 B. forsythmajori из среднемиоценовых местонахождений Мёрген II и Мёрген V, расположенных в бассейне Тунггур (Qiu, 1996, рис. 72 F, H ). Кроме того, в местонахождениях Мёрген II и Мёрген V встречается p3, тригонид которого имеет выпрямленный внешний край, как у B. forsythmajori из Монголии (Qiu, 1996, рис. 2 G, J). Значительная вариабельность в строении p3 прослежена у пищух из Мёрген II и Мёрген V, а также местонахождения Дамяо; на передневнутреннем крае p3 некоторых особей появляется дополнительное небольшое углубление без цемента (Qiu, 1996, рис. 72 H ; Zhang et al., 2012, рис. 2 A). В дальнейшем происходит усложнение этого зуба, передненаружная складка p3 становится очень глубокой и включает большое количество цемента, что прослежено у B. cf. B. forsythmajori (Zhang et al., 2012, рис. 2 B, C). Возможно, отдельные особи с таким морфотипом p3 представляют собой базальную форму зарождающегося нового вида.
B. forsythmajori из Долины Озер близок по размеру к B. kazakhstanica, однако, в среднем этот вид несколько мельче последнего и отличается строением p3, имеющего заостренную вершину, у казахстанской формы этот зуб крупнее и имеет закругленный передний край. От B. yanghuensis отличается более крупными размерами и заостренным передним краем p3.
Распространение. Центральная Монголия, Долина Озер, свита Лу; средний миоцен, зона D 1/2 (Улан Толгой – UTO-A/6) (Daxner-Höck et al., 2017); Улан Тологой (Ербаева, 1981). Китай, свита Тунггур, Тайрум Нор, Мёрген II, Мёрген V и Манделин Чаба II, Дамяо, Уланьхуа, Сыцзыван, Ци (Siziwang, Qi); средний миоцен (Dawson, 1961; Qiu, 1996; Zhang et al., 2012).
Материал. Улан Толгой (UTO-A/6): 1 P3 (NHMW 2013/0378/0003), 2 P4 (NHMW 2013/0378/ 0006-0007), 2 M1 (NHMW 2013/0378/0008-0009), 2 p3 (NHMW 2013/0378/0001-0002), 1 p4 (NHMW 2013/0378/0004), 1 m2 (NHMW 2013/0378/0005), 1 m3 (NHMW 2013/0378/0016); 1 p3-m3 (ПИН № 3218/22).
Bellatona kazakhstanica Ербаева, 1988
Bellatona kazakhstanica: Ербаева, 1988, с. 60, рис. 16-1, 2 .
Bellatona cf. kazakhstanica: Erbajeva, Daxner-Höck, 2014, с. 238, рис. 16-1, 2.
Голотип – ПИН, № 3462/46, фрагмент левой верхнечелюстной кости с P3–M2; Казахстан, Ашутас; ранний миоцен, акжарская свита.
Описание (рис. 2). Пищуха средних размеров; P2 овальной формы с одной передней входящей складкой. На P3 (рис. 2, а) гипострия мелкая, заполнена небольшим количеством цемента; парафлекса умеренной глубины, начинается и заканчивается на уровне середины ширины зуба; длина антеролофа немного превышает половину ширины зуба; протокон и гипокон равной величины. На P4–M2 (рис. 2, а) глубина гипострии варьирует: на P4 проходит до середины ширины зуба, на M1 и M2 гипострия немного глубже, заполнена цементом. Задний край M2 гладкий. На всех верхних зубах эмаль развита на переднем и внутреннем краях зубов.
Рис. 2.
Bellatona kazakhstanica Erbajeva, 1988, зубы, вид со стороны жевательной поверхности: а – правый P3–M2 (NHMW 2011/0188/0007); б – правый p3 (NHMW 2011/0188/0011); в – левый p3 (NHMW 2011/0188/0003); г – левый p3 (NHMW 2011/0188/0004); д – левый m1–m3 (NHMW 2011/0188/0014); Унхэльцэг (UNCH-A/3).
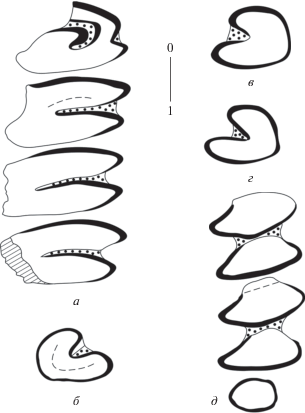
На p3 (рис. 2, б–г) одна передненаружная входящая складка, заполнена цементом. Зуб имеет закругленные гладкие края, эмаль развита по всему периметру зуба. Тригонид и талонид p4–m2 соединены цементом, эмаль отсутствует только на переднем крае конидов (рис. 2, д); m3 овально-округлой формы, эмаль развита по всему периметру зуба (рис. 2, д).
Размеры. См. табл. 2, 3.
Таблица 2.
Промеры верхних зубов Bellatona kazakhstanica Erbajeva
| Промеры | n | m | min | max | sd | |
|---|---|---|---|---|---|---|
| 1 | P3-M2 | 1 | 7.2 | |||
| 2 | P3-M1 | 2 | 5.25 | 5.0 | 5.5 | 0.354 |
| 3 | P3-P4 | 2 | 3.4 | 3.2 | 3.6 | 0.283 |
| 4 | P2 L | 1 | 0.6 | |||
| 5 | P2 W | 1 | 1.0 | |||
| 6 | P3 L | 5 | 1.56 | 1.5 | 1.65 | 0.082 |
| 7 | P3 W | 5 | 2.44 | 2.0 | 2.8 | 0.365 |
| 8 | P3 a-loph | 5 | 1.45 | 1.25 | 1.75 | 0.226 |
| 9 | P4 L | 2 | 1.75 | 1.5 | 2.0 | 0.354 |
| 10 | P4 W | 2 | 3.0 | |||
| 11 | P4 hyp | 2 | 1.95 | 1.6 | 2.3 | 0.495 |
| 12 | M1 L | 5 | 1.49 | 1.35 | 1.85 | 0.21 |
| 13 | M1 W | 5 | 2.59 | 2.25 | 3.25 | 0.46 |
| 14 | M1 hyp | 5 | 1.72 | 1.4 | 2.0 | 0.23 |
| 15 | M2 L | 3 | 1.42 | 1.15 | 1.6 | 0.236 |
| 16 | M2 W | 3 | 2.53 | 2.0 | 3.0 | 0.503 |
| 17 | M2 hyp | 3 | 1.65 | 1.45 | 1.9 | 0.229 |
Таблица 3.
Промеры нижних зубов Bellatona kazakhstanica Erbajeva
| Промеры | n | m | min | max | sd | |
|---|---|---|---|---|---|---|
| 1 | p4-m2 | 1 | 5.6 | |||
| 2 | p3 L | 3 | 1.4 | 1.35 | 1.45 | 0.05 |
| 3 | p3 W | 3 | 1.85 | 1.75 | 1.9 | 0.087 |
| 4 | p4 L | 1 | 1.75 | |||
| 5 | trig L | 1 | 0.85 | |||
| 6 | trig W | 1 | 2.0 | |||
| 7 | tal L | 1 | 0.9 | |||
| 8 | tal W | 1 | 1.65 | |||
| 9 | m1 L | 2 | 2.0 | 2.0 | 2.0 | |
| 10 | trig L | 2 | 1.0 | 1.0 | 1.0 | |
| 11 | trig W | 2 | 1.95 | 1.9 | 2.0 | |
| 12 | tal L | 2 | 0.95 | 0.9 | 1.0 | |
| 13 | tal W | 2 | 1.87 | 1.75 | 2.0 | |
| 14 | m2 L | 4 | 2.05 | 2.0 | 2.2 | 0.1 |
| 15 | trig L | 4 | 1.05 | 1.0 | 1.1 | 0.05 |
| 16 | trig W | 4 | 1.8 | 1.6 | 1.9 | 0.126 |
| 17 | tal L | 4 | 0.9 | 0.8 | 1.0 | 0.082 |
| 18 | tal W | 4 | 1.7 | 1.5 | 1.9 | 0.165 |
| 19 | m3 L | 2 | 0.8 | 0.7 | 0.9 | |
| 20 | m3 W | 2 | 0.8 | 0.7 | 0.9 |
Сравнение. По размерам B. kazakhstanica из Монголии близка к материалу из Казахстана, однако, у нее в среднем p3 незначительно крупнее, а P3 чуть мельче. От B. forsythmajori отличается несколько большими размерами зубов и закругленными краями p3. От совместно встречающегося вида B. yanghuensis рассматриваемая форма отличается более крупными размерами, строением p3, наличием передней входящей складки на P2 и несколько меньшей глубиной гипострии на P4–M2.
Распространение. Центральная Монголия, Долина Озер, свита Лу, ранний миоцен, зона D (Унхэльцэг, UNCH-A/3; Хотулиин Тээг, HTE*, HTE-005, HTE-12/7) (Daxner- Höck et al., 2017); Казахстан, Зайсанская впадина, ранний миоцен (Ербаева, 1988, 1994).
Материал. Унхэльцэг, UNCH-A/3: 1 P2-M2 (NHMW 2013/0188/0007), 4 P3 (NHMW 2013/0188/0001, /0005, /0008, /0009), 2 M1 (NHMW 2013/0188/0002, /0010), 1 m1-m3 (NHMW 2013/0188/0014), 4 p3 (NHMW 2013/0188/0003, /0004, /0006, /0011), 2 m2 (NHMW 2013/0188/0012, /0013); Хотулиин Тээг, HTE*: 1 P3-M2 (NHMW 2013/0237/0001), 1 p4-m3 (NHMW 2013/0237/0002); HTE-005: 1 M1 (NHMW 2013/0377/0001), 1 M2 (NHMW 2013/0377/0002); HTE-12/7: 3 M1 (NHMW 2013/0393/0001-0003).
Bellatona yanghuensis Zhou, 1988
Bellatona yanghuensis: Zhou, 1988, c. 140–142, рис. 1–4; Erbajeva, Daxner-Höck, с. 239, рис. 17-1, 2 .
Голотип – IVPP, № V5247, фрагменты левой верхнечелюстной кости с P2–M2 и левой нижнечелюстной кости с p3–m3; Китай, Шанси, Синьчжо; средний миоцен.
Описание (рис. 3, 4). Пищуха мелких размеров. P2 овальной формы, входящие складки на переднем крае зуба отсутствуют. На P3 антеролоф короткий, меньше половины ширины зуба, гипострия мелкая, заполнена небольшим количеством цемента (рис. 3, а). На верхних зубах глубина гипострии варьирует: на P4 немного превышает ширину зуба, на M1 достигает 2/3 ширины зуба (рис. 3, а), а на M2 проходит до наружного края зуба. Наружные края зубов выпрямленные, внутренние заостренные; задний край M2 гладкий. Эмаль развита на передней и внутренней сторонах зубов и отсутствует на наружной и задней сторонах.
Рис. 3.
Bellatona yanghuensis Zhou, 1988, зубы, вид со стороны жевательной поверхности: а – левый P3-M1 (NHMW 2011/0189/0001); б – левый p3 (NHMW 2013/0381/0001); в – левый p4 (NHMW 2013/0381/0002); г – левый p3-m2 (NHMW 2011/0189/0008); д – правый p3 (NHMW 2011/0189/0002); е – правый p3 (NHMW 2011/0189/0006); а, г–е – Унхэльцэг (UNCH-A/3), б, в – Олон Овооны Хурэм (ODO-B/1).
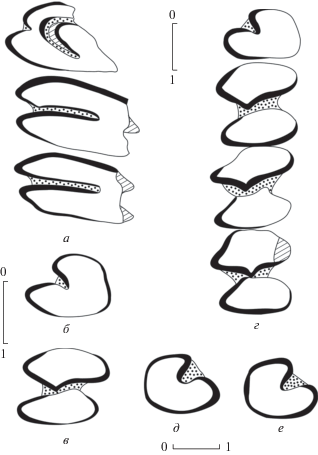
Рис. 4.
Bellatona yanghuensis Zhou, 1988, зубы, вид со стороны жевательной поверхности: а – левый p3–m3 (NHMW 2011/0190/0001); б – левый p3–m3 (NHMW 2011/0190/0002); Бугат.
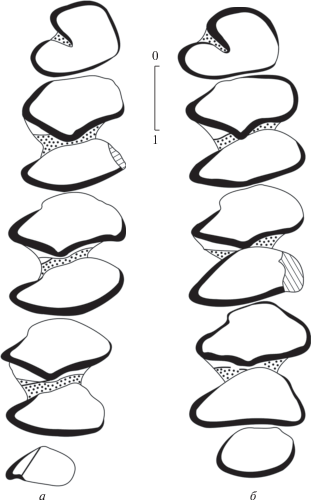
Нижнечелюстная кость сравнительно массивная и высокая, резец проходит до уровня талонида m1, образуя выпуклость на внутренней стороне челюсти и утолщение на внешней стороне под p4. Переднее ментальное отверстие расположено перед p3 на середине высоты челюсти, второе – под m1.
На p3 одна передненаружная складка, заполненная цементом (рис. 3, б–е; 4, а, б). Передний край зуба варьирует от слабовыпуклого к выпрямленному; эмаль развита по периметру зуба, кроме заднего края. На p4–m2 (рис. 3, г; 4, а, б) тригонид немного длиннее и шире талонида, m3 состоит из одного сравнительно крупного конида овально-округлой формы.
Размеры. См. табл. 4, 5.
Таблица 4.
Промеры верхних зубов Bellatona yanghuensis Zhou
| Промеры | n | m | min | max | sd | |
|---|---|---|---|---|---|---|
| 1 | P3-M1 | 1 | 3.8 | |||
| 2 | P3-P4 | 4 | 2.88 | 2.45 | 3.7 | 0.620 |
| 3 | P2 L | 2 | 0.65 | 0.55 | 0.75 | 0.141 |
| 4 | P2 W | 2 | 1.2 | |||
| 5 | P3 L | 4 | 1.38 | 1.1 | 1.8 | 0.295 |
| 6 | P3 W | 4 | 2.52 | 2.0 | 3.25 | 0.703 |
| 7 | P3a-lop | 4 | 1.25 | 0.9 | 1.8 | 0.404 |
| 8 | P4 L | 6 | 1.39 | 1.2 | 1.8 | 0.225 |
| 9 | P4 W | 6 | 2.73 | 2.25 | 3.8 | 0.640 |
| 10 | P4 hyp | 6 | 1.65 | 1.35 | 2.6 | 0.475 |
| 11 | M1 L | 14 | 1.3 | 1.0 | 1.5 | 0.160 |
| 12 | M1 W | 14 | 2.3 | 1.9 | 3.15 | 0.373 |
| 13 | M1 hyp | 14 | 1.42 | 1.15 | 1.7 | 0.214 |
| 14 | M2 L | 10 | 1.25 | 1.1 | 1.5 | 0.104 |
| 15 | M2 W | 10 | 2.17 | 2.0 | 2.75 | 0.337 |
| 16 | M2 hyp | 10 | 1.59 | 1.5 | 1.7 | 0.074 |
Таблица 5.
Промеры нижних зубов Bellatona yanghuensis Zhou
| Промеры | n | m | min | max | sd | |
|---|---|---|---|---|---|---|
| 1 | p3-m3 | 2 | 7.5 | 7.3 | 7.7 | 0.283 |
| 2 | p3-m2 | 4 | 6.55 | 5.8 | 7.1 | 0.557 |
| 3 | p3-m1 | 4 | 4.7 | 4.25 | 5.1 | 0.371 |
| 4 | p3-p4 | 4 | 2.9 | 2.75 | 3.1 | 0.165 |
| 5 | p4-m2 | 4 | 5.3 | 4.8 | 5.6 | 0.297 |
| 6 | p4-m1 | 6 | 3.7 | 3.2 | 4.5 | 0.451 |
| 7 | p3 L | 6 | 1.0 | 0.85 | 1.1 | 0.111 |
| 8 | p3 W | 6 | 1.4 | 1.25 | 1.5 | 0.174 |
| 9 | p4 L | 8 | 1.68 | 1.4 | 1.8 | 0.133 |
| 10 | trig L | 8 | 0.9 | 0.65 | 1.0 | 0.12 |
| 11 | trig W | 8 | 1.67 | 1.55 | 1.7 | 0.08 |
| 12 | tal L | 8 | 0.77 | 0.6 | 0.9 | 0.143 |
| 13 | tal W | 8 | 1.59 | 1.35 | 1.75 | 0.146 |
| 14 | m1 L | 8 | 1.77 | 1.55 | 1.95 | 0.06 |
| 15 | trig L | 8 | 0.92 | 0.85 | 1.0 | 0.078 |
| 16 | trig W | 8 | 1.7 | 1.55 | 1.75 | 0.068 |
| 17 | tal L | 8 | 0.8 | 0.7 | 0.85 | 0.132 |
| 18 | tal W | 8 | 1.65 | 1.5 | 1.8 | 0.14 |
| 19 | m2 L | 8 | 1.84 | 1.65 | 2.0 | 0.074 |
| 20 | trig L | 8 | 0.96 | 0.85 | 1.05 | 0.1 |
| 21 | trig W | 8 | 1.67 | 1.5 | 1.8 | 0.08 |
| 22 | tal L | 7 | 0.86 | 0.75 | 1.0 | 0.178 |
| 23 | tal W | 7 | 1.62 | 1.4 | 1.8 | 0.058 |
| 24 | m3 L | 3 | 0.68 | 0.65 | 0.75 | 0.153 |
| 25 | m3 W | 3 | 0.96 | 0.8 | 1.1 | 0.153 |
Сравнение. B. yanghuensis отличается от известных видов более мелкими размерами, отсутствием передней складки на P2, а от B. forsythmajori также отсутствием дополнительных входящих складок на p3 и дополнительного выступа (талона) на M2.
Распространение. Центральная Монголия, Долина Озер, свита Лу, ранний миоцен, зона D (Унхэльцэг, UNCH-A/3; Хотулиин Тээг, HTE-001, HTE-003, HTE-008), зона D1/1 (Олон Овооны Хурэм, ODO-B/1) (Daxner-Höck et al., 2017); Ара Хангайский аймак, Бугат; Китай, Шанси, Синьчжо, ранний миоцен (Zhou, 1988).
Материал. Долина Озер, Унхэльцэг, UNCH-A/3: 1 P3-M1 (NHMW 2011/0189/0001), 1 P3-P4 (NHMW 2011/0189/0009),1 P4 (NHMW 2011/0189/0013), 12 M1 (NHMW 2011/0189/0003, /0007, /0010, /0014-0016, /0018-0024), 5 M2 (NHMW 2011/0189/0011, /0017, /0025-0028),1 p3-m2 (NHMW 2011/0189/0008), 1 p4-m2 (NHMW 2011/0189/0012),3 p3 (NHMW 2011/0189/0002, /0005, /0006), 1 m2 (NHMW 2011/0189/0004); Хотулиин Тээг, HTE-001: 1 P3 (NHMW 2018/0220/ 0001); HTE-003:1 p3 (NHMW 2018/0222/0001); 1 p4 (NHMW 2018/0222/0002); 1 M2 (NHMW 2018/0222/0003); HTE-008: 1 M1 (NHMW 2019/0112/0003), 1 M2 (NHMW 2019/0112/0004, /0005); 1 p4 (NHMW 2019/0112/0006); Олон Овооны Хурэм, ODO-B/1: 1 p3 (NHMW 2013/0381/ 0001); 1 p4 (NHMW 2018/0381/0002); 1 M1 (NHMW 2013/0381/0003); Бугат: 2 p3-m3 (NHMW 2011/0190/0001, /0002), 1 p3-m2 (NHMW 2011/ 0190/0003), 1 p4-m2 (NHMW 2011/0190/0004), 1 m2 (NHMW 2011/0190/0005); 1 m1-m2 (NHMW 2011/0190/0006).
ОБСУЖДЕНИЕ
Сравнительно-морфологический анализ рассмотренных таксонов позволяет проследить эволюционное развитие рода Bellatona. Доусон, несомненно, была права, предположив, что род Sinolagomys с его прогрессивными чертами в строении зубов может быть в основании линии рода Bellatona. Так, из имеющихся видов рода Sinolagomys наибольшее сходство с Bellatona обнаруживает вид S. badamae Erbajeva (Erbajeva et al., 2017). Для него характерно удлинение тригонида p3, имеющего закругленные края, cходное с таковым у Bellatona kazakhstanica. У Sinolagomys pachygnathus Li et Qiu, 1980 парафлекса P3 намного глубже, в отличие от других видов синолагомиса, что сближает этот вид с Bellatona.
В связи с прогрессивным похолоданием климата, усилением аридизации в растительном покрове увеличилась доля ксерофитов, которые становятся превалирующими в рационе пищуховых. Переход к питанию жесткой растительностью отразился на строении зубной системы этой группы. Сглаженные боковые края зубов становятся более угловатыми, что приводит к существенным изменениям в строении жевательной поверхности зубов. Так, p3 Bellatona yanghuensis (рис. 3, 4) и B. kazakhstanica (рис. 2) имеют выпрямленный и закругленный передние края, у B. forsythmajori – заостренный; позднее на внешнем крае тригонида зуба появляются углубления от мелких до глубоких и дополнительные углубления на передневнутреннем крае p3. Постепенно увеличивается ширина талонида нижних зубов, появляются небольшие вырезки на передненаружном крае тригонидов m1 и m2 (рис. 4). На верхних зубах увеличивается глубина гипострии. Все это отражает приспособление к настриганию и нарезанию растительной пищи острыми эмалевыми краями коронок зубов и способствует увеличению площади жевательной поверхности, в связи с переходом к питанию ксерофитной растительностью. Эти морфологические изменения прослежены в линии рода Bellatona. В его составе наиболее архаичный вид B. kazakhstanica обладает сглаженными краями зубов, гипострии на верхних зубах проходят до середины ширины зуба. В дальнейшем происходит увеличение глубины гипострии, наблюдается тенденция к выпрямлению переднего края p3, появление вырезок на передненаружном крае тригонидов m1 и m2, что характерно для B. yanghuensis. Среднемиоценовый вид B. forsythmajori, обильно представленный в ряде местонахождений Китая, демонстрирует многочисленные прогрессивные черты, такие как значительное углубление гипострии на верхних зубах, появление зачатков талона на M2 и большая вариабельность в строении p3. Появление дополнительных углублений различной величины, как на наружном, так и на внутреннем крае p3 в дальнейшем привели к формированию рода Bellatonoides, отдельные морфотаксоны которого становятся, возможно, предковыми формами рода Ochotona.
Присутствие Bellatona forsythmajori, B. yanghuensis и B. kazakhstanica в фаунистических сообществах Долины Озер позволяет коррелировать одновозрастные фауны и природную среду Центральной Монголии с таковыми Казахстана и Китая.
Список литературы
Ербаева М.А. Миоценовые пищухи Монголии // Ископаемые позвоночные Монголии. М.: Наука, 1981. С. 86–95 (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 15).
Ербаева М.А. Пищухи кайнозоя (таксономия, систематика, филогения). М.: Наука, 1988. 224 с.
Ербаева М.А. Стратиграфическое распространение зайцеобразных (Lagomorpha, Mammalia) в третичных отложениях Зайсанской впадины (Восточный Казахстан) // Палеотериология / Ред. Соколов В.Е. М.: Наука, 1994. С. 65–78.
Синицын В.М. Древние климаты Евразии. Ч. 1. Л.: Изд‑во ЛГУ, 1965. 167 с.
Dawson M.R. On two ochotonids (Mammalia, Lagomorpha) from the Later Tertiary of Inner Mongolia // Amer. Museum Novit. 1961. № 2061. 15 p.
Daxner-Höck G., Badamgarav D., Barsbold R. et al. Oligocene stratigraphy across the Eocene and Miocene boundaries in the Valley of Lakes (Mongolia) // The Valley of Lakes in Mongolia, a key area of Cenozoic mammal evolution and stratigraphy / Eds. Daxner-Höck G., Göhlich U.B. Palaeodiv. and Palaeoenvir. 2017. V. 97. № 1. P. 111–218. https://doi.org/10.1007/s12549-016-0257-9
Erbajeva M.A., Bayarmaa B., Daxner-Höck G., Flynn L.J. Occurrences of Sinolagomys (Lagomorpha) from the Valley of Lakes (Mongolia) // The Valley of Lakes in Mongolia, a key area of Cenozoic mammal evolution and stratigraphy / Eds. Daxner-Höck G., Göhlich U.B. Palaeodiv. and Palaeoenvir. 2017. V. 97. № 1. P. 11–24.
Erbajeva M.A., Daxner-Höck G. The most prominent Lagomorpha from the Oligocene and Early Miocene of Mongolia // Ann. Naturhist. Museum Wien, Ser. A. 2014. Bd 116. P. 215–245.
Harzhauser M., Daxner-Höck G., Erbajeva M. et al. Oligocene and early Miocene mammal biostratigraphy of the Valley of Lakes in Mongolia // The Valley of Lakes in Mongolia, a key area of Cenozoic mammal evolution and stratigraphy / Eds. Daxner-Höck G., Göhlich U.B. Palaeodiv. and Palaeoenvir. 2017. V. 97. № 1. P. 219–231.
Harzhauser M., Daxner-Höck G, Lopez-Guerro P. et al. Stepwise onset of the icehouse world and its impact on Oligo-Miocene Central Asian mammals // Sci. Reports. 2016. 6:36169. https://doi.org/10.1038/srep36169
Lopez Martinez N. Revision sistematica y biostratigrafica de los Lagomorpha (Mammalia) del Terciario y Cuaternario de España // Mem. Museo Paleontol. Univ. Zaragoza. 1989. № 3(3). P. 1–350.
Qiu Z.D. Middle Miocene micromammalian fauna from Tunggur, Nei Mongol. Beijing: Sci. Press, 1996. P. 1–213 (in Chinese).
Sen S. Lagomorpha // Geology and paleontology of the Miocene Sinap formation, Turkey. N.Y.: Columbia Univ. Press, 2003. P. 163–178.
Young C.C. On a new ochotonid from north Suiyuan // Bull. Geol. Soc. China. 1932. V. 11. P. 255–258.
Zhang Z.Q., Kaakinen A., Wang L.H. et al. Middle Miocene ochotonids (Ochotonidae, Lagomorpha) from Damiao Pliopeticid locality // Vertebr. PalAsiat. 2012. V. 50. № 3. P. 281–292.
Zhou X. Miocene ochotonid (Mammalia, Lagomorpha) from Xinzhou, Shanxi // Vertebr. PalAsiat. 1988. V. 26. № 2. P. 139–148.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал