

Палеонтологический журнал, 2021, № 5, стр. 86-97
Палеобиогеографический анализ комплексов сеноманских эласмобранхий (Chondrichthyes, Elasmobranchii)
a Саратовский государственный университет им. Н.Г. Чернышевского
Саратов, Россия
b Саратовский областной музей краеведения
Саратов, Россия
* E-mail: palaeoanacorax@gmail.com
Поступила в редакцию 25.09.2020
После доработки 13.11.2020
Принята к публикации 11.12.2020
Аннотация
Проведена оценка степени сходства на уровне родов 23 комплексов сеноманских эласмобранхий из разных регионов мира (Поволжье, Мангышлак, Англо-Парижский бассейн, Литва, северо-запад Германии, юго-запад Франции, север Испании, Северная Америка, Венесуэла, Ливан, Египет, Алжир, Нигерия, Ангола, Индия, Австралия) по коэффициенту Жаккара. На основе кластерного анализа и палеогеографической карты высказаны предположения о возможных связях одновозрастных сообществ эласмобранхий и палеогеографических факторах, влияющих на них. Выделено пять фаунистических группировок: европейская, атлантическая, южно-тетическая, индийская, австралийская. Выявлена их зависимость от конфигурации акваторий и суши, температурного режима. Зависимость от значений глубин менее явная, хотя заметна внутри самих группировок.
ВВЕДЕНИЕ
Исследования позднемеловых, в частности, сеноманских эласмобранхий сегодня идут практически по всему миру. Накоплен огромный фактический материал. Но недостаточно освещены вопросы палеобиогеографии группы. Неравномерна степень изученности комплексов в разных регионах. На фоне большого числа публикаций по европейским, американским, австралийским сообществам, эласмобранхиям из сеномана России и сопредельных государств уделяется сравнительно мало внимания.
Остатки эласмобранхий (акулы, скаты) из сеноманских отложений бывшего СССР известны с середины XIX в. (Kiprianoff, 1852, 1854, 1859, 1881; Рогович, 1860; Синцов, 1872). Внимание исследователей в XX в. привлекали, в основном, таксономический состав и возможное биостратиграфическое значение, как отдельных таксонов, так и комплексов эласмобранхий (Гликман, 1955, 1957, 1958; Гликман, Шважайте, 1971; Мертинене, 1982; Железко, 1997). Палеобиогеографическому аспекту уделялось явно недостаточное внимание. Особенности распространения эласмобранхий рассматривались Л.С. Гликманом (1980) и Л.А. Несовым (1997), в более поздних работах данный вопрос освещается также эпизодически (Бирюков, 2013, 2014; Попов, Бирюков, 2014). Зарубежные авторы вопросы палеобиогеографии чаще рассматривают среди экологических (Underwood, Cumbaa, 2010; Guinot, 2013), но иногда и отдельно (Vullo et al., 2009; Underwood et al., 2011; Verma, 2015).
В последние годы комплекс сеноманских эласмобранхий Поволжья был ревизован, благодаря чему его разнообразие выросло; его удалось разделить на четыре ассоциации, предположительно соответствующие подъярусным подразделениям: нижнесеноманскому подъярусу, верхней и нижней частям среднесеноманского подъяруса и верхнесеноманскому подъярусу (Бирюков, 2018). Появилась возможность сравнения фауны эласмобранхий различных регионов мира с поволжскими сообществами и встраивания данных по поволжскому комплексу в мировой контекст.
В предлагаемой работе на основе современных данных о таксономическом составе комплексов эласмобранхий из сеномана различных регионов мира проведено сравнение между одновозрастными ассоциациями ряда регионов путем оценки их сходства при помощи кластерного анализа. В том числе, это позволяет понять и место поволжского комплекса в общей картине распределения группы в указанное время.
Автор выражает благодарность Е.В. Попову и Е.М. Первушову (СГУ) за ценные замечания и пожелания, высказанные в ходе обсуждения рукописи, Г. Гуино (Ун-т Монпелье, Франция) за консультации по комплексам эласмобранхий Англо-Парижского бассейна. Работа выполнена при финансовой поддержке РФФИ (проект № 14-05-00828).
МАТЕРИАЛ И МЕТОДИКА
Для характеристики сеноманского сообщества эласмобранхий Поволжья использовался ископаемый материал – зубы эласмобранхий, собиравшиеся с конца 1980-х гг. (колл. СГУ № 155), как геологами и палеонтологами СГУ, так и любителями. Обработан материал из 15 местонахождений из пределов Саратовской, Волгоградской, Пензенской, Самарской областей (Бирюков, 2018). Основная масса зубов происходит из уровней концентрации – фосфоритовых горизонтов, которые часты в отложениях сеноманского яруса Поволжья (Геология СССР, 1949, 1967, 1970). Ископаемый материал отбирался преимущественно по методу объемных проб (средний объем 0.1 м3) из выбранных интервалов с отсевом или отмывкой проб на ситах с размером ячеи 2.5, 1 и (при возможности) 0.6 мм с разделением на размерные фракции и разбором концентрата в камеральных условиях. Часть материала получена при поверхностном сборе.
Комплексом эласмобранхий охарактеризованы уровни, соответствующие трем подсвитам меловатской свиты Саратовского, Волгоградского и Пензенского Поволжья, а также пограничный интервал никольской толщи и гулюшевской свиты в Самарском Предволжье (Бирюков и др., 2018), т.е., всем трем подъярусам сеноманского яруса. По Поволжью было обработано более 100 проб с общим числом зубов более 36 тыс. Сеноманский комплекс эласмобранхий в Поволжье включает в себя представителей 10 отрядов, 24 семейств, 34 родов и 40 видов. Присутствие представителей 14 родов и 22 видов в регионе установлено впервые (Бирюков, 2018). Все таксоны даются в соответствии с систематическими взглядами А. Каппетты (Cappetta, 2012), кроме признания валидности рода Palaeoanacorax (sensu Glickman, 1971) и рассмотрения в составе одонтаспидид недавно установленного рода Pseudomegachasma Shimada et al., 2015.
Для сравнения эласмобранхий были использованы данные по следующим комплексам: 1) для раннего сеномана – Мангышлак (разрез Кольбай) (Kennedy et al., 2008), Англо-Парижский бассейн (Guinot et al., 2013), северо-запад Германии (Тевтобургский Лес) (Müller, Diedrich, 1991), юго-запад Франции (Шаранта) (Vullo et al., 2007), Алжир (Фортасса) (Mennad et al., 2020) и Индия (бассейн Кавери) (Underwood et al., 2011) (табл. 1); для среднего сеномана – Англо-Парижский бассейн (Guinot et al., 2013), Северная Америка (Западный Внутренний пролив) (Cumbaa et al., 2010), Ливан (Намура) (Forey et al., 2003) и Австралия (бассейн Карнарвон) (Berrell et al., 2020) (табл. 1); для позднего сеномана – Англо-Парижский бассейн (Guinot et al., 2013), Литва (Dalinkevičius, 1935; Adnet et al., 2008), юго-запад Франции (Шаранта) (Vullo et al., 2007), север Испании (Астурия) (Vullo et al., 2009), Техас (Welton, Farish, 1993; Cappetta, Case, 1999; Hamm, 2008), Египет (Бахария) (Werner, 1989), Нигерия (грабен Бенуэ) (Vullo, Courville, 2014), Ангола (бассейн Бенгуэла) (Antunes, Cappetta, 2002), Венесуэла (бассейн Маракайбо) (Guinot, Carrillo-Briceño, 2018) и Австралия (бассейн Карнарвон) (Berrell et al., 2020) (табл. 1). Перечень эласмобранхий Литвы из работы Й. Далинкявичюса (Dalinkevičius, 1935) приведен по современным таксономическим представлениям (Cappetta, 2012). Вследствие расхождения точек зрения разных авторов на видовой состав комплексов, сравнение произведено на уровне родов, как это сделано ранее для Англо-Парижского бассейна (Guinot, 2013) (табл. 1). В работу не вошли комплексы из сеномана Ливии (Rage, Cappetta, 2002) и Японии (Goto et al., 1996) из-за недостаточно определенной стратиграфической принадлежности материала (только до уровня яруса или шире). Комплексы из Нигерии и Анголы следует оценивать осторожно вследствие не очень точной стратиграфической привязки и бедности материала, вероятно, проистекающей из относительно слабой изученности территории. Для Венесуэлы известны данные только по ламнообразным.
Таблица 1.
Наличие (1) родов эласмобранхий в сеноманских отложениях мира
| Meristodon | Meristodonoides | Hybodus | Tribodus | Acrodus | Polyacrodus | Lissodus | Asteracanthus | Distobatus | Aegyptobatus | Ptychodus | Hexanchus | Gladioserratus | ?Notidanodon | Paraorthacodus | Synechodus | Pseudospinax | Squatina | Cretascyllium | Squalus | Protosqualus | Squalodalatias | Protocentrophorus | Heterodontus | Chiloscyllium | Hemiscyllium | Almascyllium | Mesiteia | Adnetoscyllium | Cantioscyllium | Pararhincodon | cf. Pararhincodon | Orectoloboides | Cretorectolobus | ?Cederstroemia | Cederstroemia | Annea | Palaeoanacorax | Squalicorax | Pseudocorax | Nanocorax | Microcorax | Archaeolamna | Cretolamna | Kenolamna | Cretoxyrhina | Dallasiella | Plicatolamna | Dwardius | Cardabiodon | Protolamna | Pseudoscapanorhynchus | |
| РАННИЙ СЕНОМАН | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Поволжье1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Мангышлак | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||
| АПБ1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||
| СЗГ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||
| ЮЗФ1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||
| Алжир | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Индия | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||
| СРЕДНИЙ СЕНОМАН | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Поволжье2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||
| АПБ2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||
| СА | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Ливан | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||
| Австралия2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||
| ПОЗДНИЙ СЕНОМАН | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Поволжье3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||
| Литва | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||
| АПБ3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||
| ЮЗФ3 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||
| СИ | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||
| Техас | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||
| Венесуэла | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Египет | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||
| Нигерия | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Австралия3 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||
| Ангола | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||
| Paraisurus | Scapanorhynchus | Anomotodon | Eostriatolamia | Carcharoides | Roulettia | Carcharias | ?Carcharias | Microcarcharias | Telodontaspis | Cenocarcharias | Johnlongia | Pseudomegachasma | Hispidaspis | Cretodus | Paranomotodon | Acutalamna | Pteroscyllium | Scyliorhinus | Pseudoscyliorhinus | Galeorhinus | Protoscyliorhinus | Cretascyliorhinus | `Rhinobatos` | Rhinobatos | Squatirhina | Pseudohypolophus | Libanopristis | Micropristis | Onchopristis | Baharipristis | Marckgraphia | Ptychotrigonoides | Ptychotrigon | Sechmetia | Renpetia | Peyeria | Isidobatus | Mafdentia | Archingeayia | Engolismaia | Hamrabatis | Turoniabatis | Germanobatos | Rhombopterygia | Pararaja | Cyclobatis | Cretomanta | Gymnura | Rhinoptera | Enantiobatis | ||
| РАННИЙ СЕНОМАН | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Поволжье1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||
| Мангышлак | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||
| АПБ1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||
| СЗГ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||
| ЮЗФ1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||
| Алжир | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Индия | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| СРЕДНИЙ СЕНОМАН | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Поволжье2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||
| АПБ2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| СА | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||
| Ливан | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||
| Австралия2 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ПОЗДНИЙ СЕНОМАН | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Поволжье3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||
| Литва | 1 | 1 | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||
| АПБ3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| ЮЗФ3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||
| СИ | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| Техас | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||
| Венесуэла | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Египет | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||
| Нигерия | 1 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||
| Австралия3 | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ангола | 1 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
Сокращения здесь и далее: АПБ1 – Англо-Парижский бассейн, ранний сеноман; АПБ2 – Англо-Парижский бассейн, средний сеноман; АПБ3 – Англо-Парижский бассейн, поздний сеноман; СЗГ – северо-запад Германии; ЮЗФ1 – юго-запад Франции, ранний сеноман; ЮЗФ3 – юго-запад Франции, поздний сеноман; СА – Северная Америка; СИ – север Испании.
Для более объективной оценки сходства между комплексами эласмобранхий Поволжья и иных регионов был применен кластерный анализ, ранее показавший хорошие результаты при анализе верхнемеловых комплексов эласмобранхий Англо-Парижского бассейна (Guinot, 2013). На основе наличия или отсутствия родов в регионах произведено вычисление коэффициента сходства Жаккара по формуле Kj = c/a + b – c (где a – число таксонов в первом сравниваемом комплексе, b – число таксонов во втором сравниваемом комплексе, с – число таксонов, общих для 1-го и 2-го комплексов) методом одиночной связи с последующим построением дендрограмм при помощи программы Past 3 (Hammer, 2018). Анализ произведен на основе подъярусных комплексов (табл. 2). Коэффициенты сходства Отиаи, Кульчинского, Дайса имеют близкие значения.
Таблица 2.
Значения коэффициента Жаккара для комплексов сеноманских эласмобранхий мира
| Поволжье1 | Поволжье2 | Поволжье3 | Мангышлак | Литва | АПБ1 | АПБ2 | АПБ3 | СЗГ | ЮЗФ1 | ЮЗФ3 | СИ | СА | Техас | Венесуэла | Ливан | Алжир | Индия | Египет | Нигерия | Австралия2 | Австралия3 | Ангола | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Повол-жье1 | 1 | 0.81481 | 0.69697 | 0.4 | 0.28125 | 0.39474 | 0.425 | 0.4 | 0.26316 | 0.15909 | 0.125 | 0.10811 | 0.2619 | 0.10811 | 0.11111 | 0.03125 | 0 | 0.15385 | 0.02381 | 0.037037 | 0.26923 | 0.11111 | 0.11538 |
| Повол-жье2 | 0.81481 | 1 | 0.78125 | 0.38889 | 0.3125 | 0.42105 | 0.48718 | 0.46154 | 0.25641 | 0.18182 | 0.12121 | 0.10526 | 0.28571 | 0.16667 | 0.10714 | 0.030303 | 0 | 0.14815 | 0.047619 | 0.035714 | 0.25926 | 0.14815 | 0.11111 |
| Повол-жье3 | 0.69697 | 0.78125 | 1 | 0.39024 | 0.32432 | 0.4186 | 0.47727 | 0.45455 | 0.24444 | 0.20408 | 0.15789 | 0.13953 | 0.29787 | 0.19512 | 0.085714 | 0.025 | 0 | 0.11765 | 0.0625 | 0.028571 | 0.20588 | 0.11765 | 0.088235 |
| Мангышлак | 0.4 | 0.38889 | 0.39024 | 1 | 0.27273 | 0.31707 | 0.31818 | 0.35714 | 0.19512 | 0.2381 | 0.23333 | 0.16667 | 0.25581 | 0.23529 | 0.10714 | 0.030303 | 0 | 0.10714 | 0.073171 | 0.074074 | 0.21429 | 0.068966 | 0.2 |
| Литва | 0.28125 | 0.3125 | 0.32432 | 0.27273 | 1 | 0.27778 | 0.31579 | 0.36111 | 0.28125 | 0.073171 | 0.16 | 0.13333 | 0.17949 | 0.17241 | 0.15 | 0.04 | 0 | 0.15 | 0.028571 | 0.05 | 0.083333 | 0.21053 | 0.22222 |
| АПБ1 | 0.39474 | 0.42105 | 0.4186 | 0.31707 | 0.27778 | 1 | 0.87879 | 0.79412 | 0.47222 | 0.21739 | 0.13889 | 0.21053 | 0.16 | 0.12195 | 0.09375 | 0.027027 | 0 | 0.060606 | 0.021277 | 0.03125 | 0.1875 | 0.09375 | 0.096774 |
| АПБ2 | 0.425 | 0.48718 | 0.47727 | 0.31818 | 0.31579 | 0.87879 | 1 | 0.85714 | 0.46154 | 0.2 | 0.125 | 0.19048 | 0.19231 | 0.13636 | 0.11429 | 0.02439 | 0 | 0.11429 | 0.019608 | 0.027778 | 0.2 | 0.11429 | 0.085714 |
| АПБ3 | 0.4 | 0.46154 | 0.45455 | 0.35714 | 0.36111 | 0.79412 | 0.85714 | 1 | 0.4359 | 0.20408 | 0.12821 | 0.19512 | 0.19608 | 0.13953 | 0.11765 | 0.025 | 0 | 0.085714 | 0.02 | 0.028571 | 0.17143 | 0.11765 | 0.088235 |
| СЗГ | 0.26316 | 0.25641 | 0.24444 | 0.19512 | 0.28125 | 0.47222 | 0.46154 | 0.4359 | 1 | 0.13333 | 0.090909 | 0.17143 | 0.10417 | 0.078947 | 0.071429 | 0.03125 | 0 | 0.11111 | 0 | 0.037037 | 0.1 | 0.11111 | 0.074074 |
| ЮЗФ1 | 0.15909 | 0.18182 | 0.20408 | 0.2381 | 0.073171 | 0.21739 | 0.2 | 0.20408 | 0.13333 | 1 | 0.34483 | 0.33333 | 0.19149 | 0.25714 | 0 | 0 | 0 | 0.03125 | 0.069767 | 0.10714 | 0.090909 | 0 | 0.066667 |
| ЮЗФ3 | 0.125 | 0.12121 | 0.15789 | 0.23333 | 0.16 | 0.13889 | 0.125 | 0.12821 | 0.090909 | 0.34483 | 1 | 0.61111 | 0.24242 | 0.38095 | 0.125 | 0.05 | 0 | 0.125 | 0.068966 | 0.23077 | 0.23529 | 0.058824 | 0.30769 |
| СИ | 0.10811 | 0.10526 | 0.13953 | 0.16667 | 0.13333 | 0.21053 | 0.19048 | 0.19512 | 0.17143 | 0.33333 | 0.61111 | 1 | 0.21053 | 0.25926 | 0.095238 | 0.083333 | 0 | 0.095238 | 0.058824 | 0.10526 | 0.18182 | 0.045455 | 0.22222 |
| СА | 0.2619 | 0.28571 | 0.29787 | 0.25581 | 0.17949 | 0.16 | 0.19231 | 0.19608 | 0.10417 | 0.19149 | 0.24242 | 0.21053 | 1 | 0.39394 | 0.09375 | 0.055556 | 0 | 0.09375 | 0.14286 | 0.064516 | 0.11765 | 0.12903 | 0.17241 |
| Техас | 0.10811 | 0.16667 | 0.19512 | 0.23529 | 0.17241 | 0.12195 | 0.13636 | 0.13953 | 0.078947 | 0.25714 | 0.38095 | 0.25926 | 0.39394 | 1 | 0.095238 | 0.04 | 0 | 0.045455 | 0.125 | 0.10526 | 0.083333 | 0.095238 | 0.22222 |
| Венесуэла | 0.11111 | 0.10714 | 0.085714 | 0.10714 | 0.15 | 0.09375 | 0.11429 | 0.11765 | 0.071429 | 0 | 0.125 | 0.095238 | 0.09375 | 0.095238 | 1 | 0.071429 | 0 | 0.090909 | 0.041667 | 0 | 0.15385 | 0.2 | 0.375 |
| Ливан | 0.03125 | 0.030303 | 0.025 | 0.030303 | 0.04 | 0.027027 | 0.02439 | 0.025 | 0.03125 | 0 | 0.05 | 0.083333 | 0.055556 | 0.04 | 0.071429 | 1 | 0 | 0 | 0.037037 | 0.083333 | 0.058824 | 0 | 0.076923 |
| Алжир | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0.10526 | 0 | 0 | 0 | 0 |
| Индия | 0.15385 | 0.14815 | 0.11765 | 0.10714 | 0.15 | 0.060606 | 0.11429 | 0.085714 | 0.11111 | 0.03125 | 0.125 | 0.095238 | 0.09375 | 0.045455 | 0.090909 | 0 | 0 | 1 | 0.041667 | 0 | 0.15385 | 0.2 | 0.22222 |
| Египет | 0.02381 | 0.047619 | 0.0625 | 0.073171 | 0.028571 | 0.021277 | 0.019608 | 0.02 | 0 | 0.069767 | 0.068966 | 0.058824 | 0.14286 | 0.125 | 0.041667 | 0.037037 | 0.10526 | 0.041667 | 1 | 0 | 0.037037 | 0 | 0.090909 |
| Нигерия | 0.037037 | 0.035714 | 0.028571 | 0.074074 | 0.05 | 0.03125 | 0.027778 | 0.028571 | 0.037037 | 0.10714 | 0.23077 | 0.10526 | 0.064516 | 0.10526 | 0 | 0.083333 | 0 | 0 | 0 | 1 | 0.083333 | 0 | 0.125 |
| Австралия2 | 0.26923 | 0.25926 | 0.20588 | 0.21429 | 0.083333 | 0.1875 | 0.2 | 0.17143 | 0.1 | 0.090909 | 0.23529 | 0.18182 | 0.11765 | 0.083333 | 0.15385 | 0.058824 | 0 | 0.15385 | 0.037037 | 0.083333 | 1 | 0.15385 | 0.16667 |
| Австралия3 | 0.11111 | 0.14815 | 0.11765 | 0.068966 | 0.21053 | 0.09375 | 0.11429 | 0.11765 | 0.11111 | 0 | 0.058824 | 0.045455 | 0.12903 | 0.095238 | 0.2 | 0 | 0 | 0.2 | 0 | 0 | 0.15385 | 1 | 0.22222 |
| Ангола | 0.11538 | 0.11111 | 0.088235 | 0.2 | 0.22222 | 0.096774 | 0.085714 | 0.088235 | 0.074074 | 0.066667 | 0.30769 | 0.22222 | 0.17241 | 0.22222 | 0.375 | 0.076923 | 0 | 0.22222 | 0.090909 | 0.125 | 0.16667 | 0.22222 | 1 |
Результаты кластерного анализа были наложены на палеогеографические карты для сеноманского века, предложенные К. Скотизом (Scotese, 2014). Подобный метод был использован Ю. Криветом и Ш. Клуг для анализа палеобиогеографии юрских эласмобранхий Европы (Kriwet, Klug, 2008). Сочетание значения коэффициента сходства и карты позволяет оценить расстояния между регионами, откуда происходят сравниваемые сообщества, и господствующие там условия (например, возможные глубины, размер разделяющих их массивов суши и т.п.). Это дало возможность рассмотреть вероятность связи между комплексами эласмобранхий различных регионов.
ОЦЕНКА СТЕПЕНИ СХОДСТВА МЕЖДУ СЕНОМАНСКИМИ КОМПЛЕКСАМИ ЭЛАСМОБРАНХИЙ
Среди раннесеноманских комплексов наибольшее сходство с поволжским демонстрирует комплекс из Мангышлака [коэффициент Жаккара (далее – КЖ) – 0.4]. Несколько дальше отстоит от Поволжского комплекс Англо-Парижского бассейна (КЖ 0.39), гораздо выше его сходство с комплексом из Северо-Западной Германии (КЖ 0.47). Одинаково отдален от всех перечисленных комплекс с юго-запада Франции (КЖ с Поволжским комплексом, например, 0.16). Очень сильно обособлено индийское сообщество эласмобранхий (КЖ с сообществом из Поволжья 0.15). Нулевым КЖ ко всем остальным комплексам обладает сообщество из Алжира (рис. 1, а; табл. 2). Таким образом, один кластер формируют комплексы из Поволжья и Мангышлака, в составе другого кластера – комплексы из Англо-Парижского бассейна и с северо-запада Германии. Остальные сообщества демонстрируют большую (Алжир) или меньшую (юго-запад Франции, Индия) степень обособленности.
Рис. 1.
Дендрограммы сходства комплексов эласмобранхий сеномана: а – ранний сеноман; б – средний сеноман; в – поздний сеноман. Обозначения: 1 – терригенные отложения; 2 – карбонатные отложения; 3 – терригенно-карбонатные отложения.
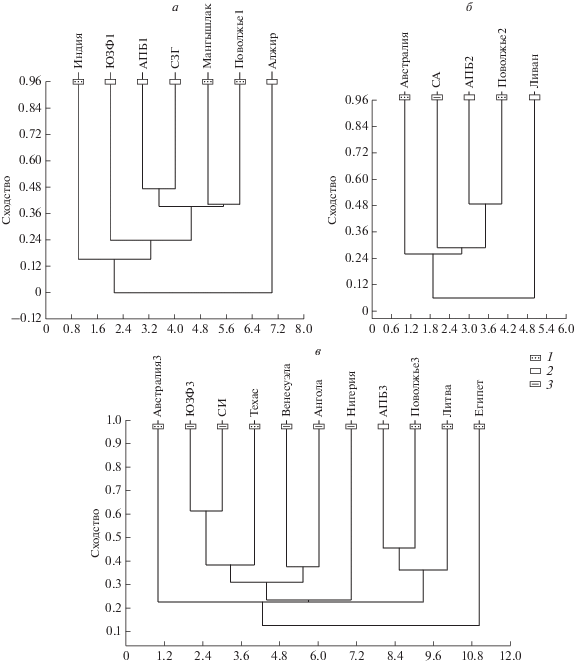
Из комплексов среднего сеномана в отдельный кластер объединяются комплексы Поволжья и Англо-Парижского бассейна (КЖ 0.49). Значительно дальше от них отстоят североамериканский (КЖ с поволжским комплексом 0.29) и австралийский (КЖ с поволжским 0.26) комплексы. Наименее схож со всеми ними комплекс из Ливана (КЖ с поволжским 0.03), выделяющийся в отдельную ветвь (рис. 1, б; табл. 2).
Для позднего сеномана сохраняется значительное сходство между поволжским и англо-парижским комплексами (КЖ 0.45), литовское сообщество эласмобранхий отличается от поволжского несколько сильнее (КЖ 0.32). Большее сходство между собой демонстрируют сообщества юго-запада Франции и севера Испании (КЖ 0.61), которым, в свою очередь, близка фауна Техаса (КЖ с французским комплексом 0.38). Комплексы из Венесуэлы и Анголы близки между собой (КЖ 0.38), но далеко отстоят от поволжского (КЖ с ангольским комплексом 0.09). Нигерийское сообщество близко к атлантическим сообществам (КЖ с французским комплексом 0.23), но имеет мало сходства с поволжским и англо-парижским (КЖ с поволжским комплексом 0.03). Австралийский комплекс демонстрирует большее сходство с ними (КЖ с поволжским комплексом 0.12). Египетское сообщество одинаково отдалено ото всех данных кластеров (КЖ с поволжским комплексом 0.06) (рис. 1, в; табл. 2).
ОБСУЖДЕНИЕ
Таким образом, при помощи коэффициентов сходства и кластерного анализа все комплексы сеноманских эласмобранхий мира можно объединить в несколько фаунистических группировок, тяготеющих к определенному интервалу широт. Условно группировки можно назвать: группировка Европейской палеобиогеографической области (Беньямовский, 2008) (комплексы Поволжья, Мангышлака, Англо-Парижского бассейна, северо-запада Германии, Литвы), атлантическая группировка (комплексы юго-запада Франции, севера Испании, Северной Америки, Венесуэлы, Анголы, Нигерии), южно-тетическая группировка (комплексы Алжира, Ливана, Египта). Отдельные группировки составляют комплексы из Индии и Австралии.
На основе анализа сходства между комплексами эламобранхий из различных регионов можно сделать предположения о возможных связях между ними, привлекая данные палеогеографии. Для этого были задействованы реконструкции Скотиза (Scotese, 2014).
Высокое значение коэффициента Жаккара для раннесеноманских комплексов Поволжья и Мангышлака хорошо объясняется тем, что эти сообщества находились в соседних бассейнах [Поволжско-Донской и Мангышлак-Восточно-Прикаспийский по В.Н. Беньямовскому (2008)] и не разделялись сколько-нибудь значимыми преградами (рис. 2, а), хотя для Мангышлака и характерны более теплолюбивые формы, например, роды Cretodus и Hispidaspis (Kennedy et al., 2008). Последний на основании характера распространения был отнесен к теплолюбивым Гликманом (1980) в противоположность роду Eostriatolamia. Точно так же нахождение в рамках смежных бассейнов можно назвать причиной еще большего сходства сообществ Англо-Парижского бассейна и северо-запада Германии (рис. 2, а). Комплексы бассейнов позднемелового Русского моря и Англо-Парижского бассейна также довольно близки между собой, вероятно, вследствие их относительно свободного сообщения друг с другом (рис. 2, а).
Рис. 2.
Диаграмма связей между сеноманскими сообществами эласмобранхий: а – ранний сеноман (1 – Поволжье, 2 – Мангышлак, 3 – Англо-Парижский бассейн, 4 – северо-запад Германии, 5 – юго-запад Франции, 6 – Алжир, 7 – Индия); б – средний сеноман (1 – Поволжье, 2 – Англо-Парижский бассейн, 3 – Северная Америка, 4 – Ливан, 5 – Австралия); в – поздний сеноман (1 – Поволжье, 2 – Литва, 3 – Англо-Парижский бассейн, 4 – юго-запад Франции, 5 – север Испании, 6 – Техас, 7 – Венесуэла, 8 – Египет, 9 – Нигерия, 10 – Австралия, 11 – Ангола). В прямоугольниках приведено значение коэффициента Жаккара. Палеогеографическая основа – по Scotese (2014).

Фауна юго-запада Франции одинаково мало схожа с фаунами названных акваторий. Вероятно, это явилось следствием того, что Аквитанский бассейн на востоке был почти изолирован крупным островом и фактически являлся частью восточной Атлантики.
Еще более слабое сходство комплекса эласмобранхий из Индии со всеми ранее упомянутыми сообществами можно объяснить ее отдаленностью ото всех остальных – она единственная находилась в то время в южном полушарии (рис. 2, а). Алжирское же сообщество продемонстрировало полное отсутствие сходства с остальными. Это, возможно, продиктовано его нахождением на южной периферии океана Тетис и приуроченностью к заливу, глубоко вдающемуся в массив суши.
В середине сеноманского века продолжает оставаться актуальным сходство комплексов Поволжья и Англо-Парижского бассейна, несколько сильнее от них отличается комплекс из Северной Америки и наименее схож комплекс из Ливана. Подобная ситуация также объяснима отсутствием преград и близостью по расстояниям между Поволжьем и Англо-Парижским бассейном и, напротив, некоторой изолированностью Внутреннего Западного пролива Северной Америки крупными массивами суши, а Ливана – глубоководной частью океана Тетис и, возможно, сочетанием специфических местных условий (температуры, глубины, очертания береговой линии), характерных для южной периферии океана Тетис (рис. 2, б). Австралийский комплекс схож с европейскими и североамериканским комплексами, возможно, из-за приуроченности к относительно открытым акваториям.
Сходство между поволжским и англо-парижским, а также между комплексами с юго-запада Франции, севера Испании, из Техаса и, в меньшей степени, из Венесуэлы, Анголы и Нигерии в позднем сеномане также можно интерпретировать через отсутствие между ними значимых преград в виде суши (юго-запад Франции и север Испании разделяет крайне небольшое расстояние в пределах Аквитанского бассейна) (рис. 2, в). А территория Литвы отделена от Поволжья крупным массивом суши и представляет собой почти замкнутый залив. Восточная Атлантика также изолирована островом значительной площади от остальных европейских бассейнов. Комплекс из Австралии, вследствие отделения ото всех остальных огромными акваториями, сильно обособлен, хотя, вероятно, сохранил с ними связь. Египетская фауна занимает южную периферию океана Тетис, которая обособлена от Европы глубоководными участками, а от Америки – обширной сушей. Также, вероятно, сыграла роль близость обширного залива (рис. 2, в).
Различия между фаунами эласмобранхий также могут быть проинтерпретированы при помощи значений широтной приуроченности и глубин, господствовавших на тот момент в регионах. Так, комплексы из Поволжья, Англо-Парижского бассейна, Германии, Литвы носят явно бореальный характер за счет обилия ламнообразных, синеходонтиформных, катранообразных, многожаберных акул (Guinot, 2013). При этом в Поволжье преобладают ламнообразные акулы, что указывает, вероятно, на меньшие глубины в Русском море. А род Eostriatolamia, наиболее многочисленный в среднем и позднем сеномане Поволжья, еще Гликманом (1980) рассматривался как типично бореальный. Мелководность и относительная холодноводность этой акватории подтверждаются и данными по микрофауне (Vishnevskaya, Kopaevich, 2020). Хотя мангышлакский комплекс и характеризуется наличием более теплолюбивых форм (роды Cretodus и Hispidaspis, гитарниковые скаты Rhinobatos) (Kennedy et al., 2008), общность его с поволжским чрезвычайно велика.
Гораздо более теплолюбивыми выглядят фауны юго-запада Франции (Vullo et al., 2007), севера Испании (Vullo et al., 2009), Ливана (Forey et al., 2003), Египта (Werner, 1989), вследствие преобладания мелких кархариновых, оректолобиморфов, скатов (в т.ч. различных склероринхид), большого числа родов гибодонтов (Guinot, 2013) (табл. 1). По данным признакам к ним примыкают фауны Северной Америки (Welton, Farish, 1993; Cappetta, Case, 1999; Hamm, 2008; Cumbaa et al., 2010), но количество и разнообразие ламнообразных в них гораздо выше. Однако значение КЖ (чуть менее 0.2) между техасским и поволжским сообществами выше, чем между поволжским и гораздо более близким к нему территориально сообществом юго-запада Франции (0.16). Вероятно, в данном случае сыграл роль фактор глубин; судя по фациальному составу, комплексы Техаса и Поволжья более мелководные, что и сказалось на их сходстве.
Венесуэльский и африканские комплексы также схожи с техасским и комплексами Аквитанского бассейна. Черты тепловодности (наличие рода Cretodus) несет в себе и индийский комплекс. Это обеспечивает его сходство с мангышлакским, аквитанскими, североамериканскими, египетским и ангольским комплексами и одновременно отделяет от австралийских, чьи места обитания располагались не так уж и далеко, но сходство с бореальными сообществами гораздо выше. Можно предположить, что здесь влияние оказал характер движения водных масс. Присутствие же шестижаберниковых рода ?Notidanodon обособляет индийский комплекс ото всех остальных.
Любопытно, что комплекс из Венесуэлы гораздо более схож с поволжским (КЖ 0.09), чем с более близким широтно египетским (КЖ 0.04), также сильно отличающимся от поволжского (КЖ 0.06), вероятно, за счет большего количества ламнообразных акул в поволжском и венесуэльском комплексах, причиной чего, возможно, были местные условия – более открытое море, в отличие от лагунных условий Египта.
Средне- и познесеноманский комплексы Австралии приурочены, как и европейские, к умеренным широтам, только нотальной зоны. Это, очевидно, и сказалось на их составе, схожем с таковым в европейских комплексах (особенно для среднего сеномана).
Фауна эласмобранхий Поволжья выглядит не только как бедная и мелководная, но и как одна из наиболее консервативных (КЖ между ранне- и среднесеноманским комплексами составляет 0.81, между средне- и позднесеноманским 0.78, между ранне- и позднесеноманским почти 0.7). Большим коэффициентом обладают только сообщества Англо-Парижского бассейна (между ранне- и среднесеноманским комплексами 0.88, между средне- и позднесеноманским 0.86, между ранне- и позднесеноманским 0.79). Фауны из других регионов гораздо менее консервативны. Например, КЖ между ранне- и позднесеноманскими комплексами юго-запада Франции достигает 0.34, а между австралийскими средне- и позднесеноманским лишь 0.15. Разнообразие поволжского комплекса в среднем и позднем сеномане растет за счет появления лишь нескольких родов (Protoscyliorhinus в первой половине среднего сеномана, Ptychodus и Squalicorax в его второй половине, Galeorhinus и Cantioscyllium в позднем сеномане). Возможно, подобный консерватизм – общая черта бореальных сообществ. Несмотря на консерватизм и бедность Поволжского комплекса, он практически полностью состоит из космополитных форм. Такой же чертой обладает и мангышлакское сообщество. В противоположность этому, комплексы Англо-Парижского бассейна включают в себя несколько эндемичных на тот отрезок времени родов – Annea, Adnetoscyllium, Cretascyliorhinus и дидемиков (sensu Lebedev, Zakharenko, 2010), встреченных еще и на северо-западе Германии – Pseudospinax и Pseudoscyliorhinus. Вероятно, данную ситуацию можно проинтерпретировать, опираясь на реконструкцию условий, господствовавших в акваториях. Поволжские и мангышлакское сообщества мелководны (местонахождения приурочены к терригенным отложениям), а сообщества Англо-Парижского бассейна и северо-запада Германии обитали при более значительных глубинах (местонахождения приурочены к карбонатным отложениям). Поскольку эндемики и дидемики представлены относительно глубоководными формами, можно предположить затрудненность их миграции в акватории с меньшими глубинами. Наличие эндемиков в позднем сеномане Литвы (Squalodalatias, Protocentrophorus) логично было бы объяснить некоторой изолированностью данного бассейна крупным массивом суши. Любопытно, что оба рода – катранообразные. Парадоксально, что данный отряд преимущественно глубоководный, а в случае с литовским сообществом названные формы известны из терригенных фаций.
В фаунах Аквитанского бассейна в раннем и позднем сеномане юго-запада Франции и позднем сеномане севера Испании заметно присутствие эндемичных скатов Ptychotrigonoides, Archingeayia и Engolismaia (Vullo et al., 2007, 2009). На южной периферии океана Тетис в среднем сеномане Ливана значительную часть комплекса составляют эндемичные скаты Libanopristis, Pararaja и Cyclobatis, а в позднем сеномане Египта такие эндемики, как гибодонты Distobatus, Aegyptobatus и скаты Baharipristis, Marckgrafia, Sechmetia, Renpetia, Isidobatus и Mafdentia фактически формируют облик сообщества (Werner, 1989; Forey et al., 2003). Еще более это заметно в раннесеноманском алжирском комплексе, где эласмобранхии представлены исключительно скатами-эндемиками Baharipristis и Mafdentia (Mennad et al., 2020). Все перечисленные формы можно рассматривать как региональные эндемики (Lebedev, Zakharenko, 2010).
Помимо географической принадлежности (бореальная, тетическая, нотальная области) на состав сообществ оказывает влияние и фактор глубин. Примерные значения глубин могут быть реконструированы по фациальной принадлежности вмещающих пород (рис. 1) и по таксономическому составу сообществ. Наиболее глубоководными предстают комплексы Англо-Парижского бассейна и северо-запада Германии, на что указывает как состав (представители родов Protosqualus, Squalus, Notorynchus, Cretascyliorhinus, Pteroscyllium, Scyliorhinus и Hexanchus), так и карбонатные отложения в качестве вмещающих, соответствующие условиям внешнего шельфа. Также глубоководным, судя по вмещающим породам (мел и известняк), являлось среднесеноманское ливанское сообщество. Несколько менее глубоководны комплексы из Аквитанского бассейна (отсутствие палеоспинацид, катранов, сцилиоринид), доля карбонатов там значительно меньше. Остальные сообщества гораздо более мелководные, вплоть до скорее лагунных комплексов Египта и Алжира. Как видно из дендрограмм сходства (рис. 1), фактор глубин оказывает влияние на состав сообществ, но оно, вероятно, не такое прямое, как у широтного фактора – в одном кластере могут находиться комплексы из акваторий с разными глубинами (как поволжский и англо-парижский комплексы в среднем и позднем сеномане). Наиболее заметно влияние значений глубин внутри каждой группировки в отдельности (сходство англо-парижского и северогерманского комплексов в европейской группировке или комплексов юго-запада Франции и севера Испании в атлантической).
ВЫВОДЫ
На основании анализа коэффициента сходства Жаккара комплексы сеноманских эласмобранхий мира были разделены на пять фаунистических группировок: европейскую, атлантическую, южно-тетическую, индийскую и австралийскую.
Применение палеогеографической карты позволило выявить зависимость группировок от широтной приуроченности и конфигурации акваторий и массивов суши, работающих как фактор изоляции. Очертания береговой линии также оказывают влияние – обширные заливы (как в Алжире и Египте) становятся местами обитания изолированных, в значительной мере эндемичных сообществ. При наличии открытых вод сходство даже географически отдаленных сообществ может быть высоким, как, например, между поволжским и англо-парижским, или венесуэльским и ангольским комплексами.
Фаунистические группировки явно зависимы от температурного режима. Преимущественно холодноводными являются европейская и австралийская группировки, обитавшие в бореальной и нотальной зонах, соответственно. Более тепловодны атлантическая, южно-тетическая и индийская группировки. Бореальные комплексы Англо-Парижского бассейна и Поволжья в сеномане наиболее консервативны. Влияние значения глубин является не столь прямым, но его роль также бесспорна, что видно внутри каждой отдельной группировки. Факторы оказывают комплексное влияние, благодаря сочетанию местных условий даже географически отдаленные сообщества (например, поволжское и техасское) могут обладать заметным сходством.
Палеогеографические выводы по эласмобранхиям могут подтверждаться данными по другим группам. Относительная холодноводность и мелководность акваторий Поволжья отражена и в характере микрофауны.
Оценка степени сходства между комплексами сеноманских эласмобранхий с привлечением палеогеографической карты и данных по фациальному составу местонахождений позволяет выявить вероятные взаимосвязи между сообществами, оценить факторы, воздействующие на них. В дальнейшем это, возможно, будет способствовать разрешению вопросов о динамике разнообразия сеноманских эласмобранхий в различных регионах.
Список литературы
Беньямовский В.Н. Схема инфразонального биостратиграфического расчленения верхнего мела Восточно-Европейской провинции по бентосным фораминиферам. Статья 1. Сеноман–коньяк // Стратигр. Геол. корреляция. 2008. Т. 16. № 3. С. 36–46.
Бирюков А.В. Первая находка остатков гитарниковых скатов (Elasmobranchii: Rhinobatidae) в верхнем мелу России // Современная палеонтология: классические и новейшие методы. Тез. докл. десятой всеросс. науч. школы молодых ученых-палеонтологов. М.: ПИН РАН, 2013. С. 2–3.
Бирюков А.В. Комплекс эласмобранхий (Chondrichthyes) из нижнего сеномана Поволжья // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии / Ред. Барабошкин Е.Ю., Маркевич В.С., Бугдаева Е.В. и др. Владивосток: Дальнаука, 2014. С. 55–58.
Бирюков А.В. О стратиграфическом значении эласмобранхий (Chondrichthyes, Elasmobranchii) в сеномане Правобережного Поволжья // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2018. Т. 18. Вып. 1. С. 27–40.
Бирюков А.В., Попов Е.В., Морова А.А., Моров В.П. Новый комплекс эласмобранхий (Chondrichthyes) из базального горизонта турона Самарского Предволжья // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Матер. IX Всеросс. совещ. 17–21 сентября 2018 г. / Ред. Барабошкин Е.Ю., Липницкая Т.А., Гужиков А.Ю. Белгород: ПОЛИТЕРРА, 2018. С. 58–61.
Геология СССР. Т. VI. Воронежская и смежные области. Ч. I. Геологическое описание / Ред. Дубянский А.А. и др. М.: Гос. изд-во геол. лит-ры, 1949. 340 с.
Геология СССР. Том XI. Поволжье и Прикамье. Ч. I. Геологическое описание / Ред. Чепиков К.Р. и др. М.: Недра, 1967. 872 с.
Геология СССР. Т. XLVI. Ростовская, Волгоградская, Астраханская области и Калмыцкая АССР. Ч. I. Геологическое описание / Ред. Белов Ф.А. и др. М.: Недра, 1970. 668 с.
Гликман Л.С. О возрасте фосфоритового горизонта в кровле сеномана окрестностей Саратова по находкам зубов рыб // Уч. зап. СГУ. 1955. Т. 45. Вып. геол. С. 83–84.
Гликман Л.С. О возрасте нижнего фосфоритового горизонта в окрестностях Красного Яра Сталинградской области // Тр. Геол. музея им. А.П.Карпинского. 1957. Вып. 1. С. 118–120.
Гликман Л.С. О темпах эволюции ламноидных акул // Докл. АН СССР. 1958. Т. 123. № 3. С. 568–571.
Гликман Л.С. Эволюция меловых и кайнозойских ламноидных акул. М.: Наука, 1980. 248 с.
Гликман Л.С., Шважайте Р.А. Акулы семейства Anacoracidae из сеномана и турона Литвы, Поволжья и Средней Азии // Палеонтология и стратиграфия Прибалтики и Белоруссии. Вильнюс, 1971. С. 185–193.
Железко В.И. Зональная хроностратиграфическая шкала верхнего мела Евразии по акуловым рыбам // Ежегодник-96. Екатеринбург, 1997. С. 9–13.
Мертинене Р.А. Зональное расчленение альбских и сеноманских отложений западной части Восточно-Европейской платформы по зубам акул // Современное значение палеонтологии для стратиграфии. Л., 1982. С. 110–117.
Несов Л.А. Неморские позвоночные мелового периода Северной Евразии. СПб.: СПб. ун-т, 1997. 218 с.
Попов Е.В., Бирюков А.В. Кархаринообразные акулы (Elasmobranchi: Carcharhiniformes) в сеномане Поволжья // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии / Ред. Барабошкин Е.Ю., Маркевич В.С., Бугдаева Е.В. и др. Владивосток: Дальнаука, 2014. С. 262–263.
Рогович А.С. Об ископаемых рыбах губерний Киевского учебного округа. Киев,1860. 105 с.
Синцов И.Ф. Об юрских и меловых окаменелостях Саратовской губернии // Матер. для геологии России. Т. 4. СПб.: Имп. Акад. наук, 1872. С. 1–127.
Adnet S., Cappetta H., Mertiniene R.A. Re-evaluation of squaloid shark records from the Albian and Cenomanian of Lithuania // Cret. Res. 2008. V. 29. № 4. P. 711–722.
Antunes M.T., Cappetta H. Sélaciens du Crétacé (Albien-Maastrichtien) d’Angola // Palaeontogr. Abt. A. 2002. V. 264. № 5–6. P. 85–146.
Berrell R.W., Boisvert C., Trinajstic K. et al. A review of Australia’s Mesozoic fishes // Alcheringa. 2020. V. 44. № 2. P. 286–311. https://doi.org/10.1080/03115518.2019.1701078
Cappetta H. Handbook of Paleoichthyology. Vol. 3E: Chondrichthyes Mesozoic and Cenozoic Elasmobranchii: Teeth. München: Verlag Dr. F. Pfeil, 2012. 512 p.
Cappetta H., Case G.R. Additions aux faunes de sélaciens du Crétacé du Texas (Albien supérieur–Campanien) // Palaeo Ichthyol. 1999. V. 9. P. 5–111.
Cumbaa S.L., Shimada K., Cook T.D. Mid-Cenomanian vertebrate faunas of the Western Interior Seaway of North America and their evolutionary, paleobiogeographical, and paleoecological implications // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2010. V. 295. № 1–2. P. 199–214.
Dalinkevičius J.A. On the fossil fishes of the Lithuanian chalk. I. Selachii // Mém. Fac. Sci. Univ. Vytautas le Grand. 1935. V. 9. P. 243–305.
Forey P.L., Yi L., Patterson C., Davies C.E. Fossil fishes from the Cenomanian (Upper Cretaceous) of Namoura, Lebanon // J. Syst. Palaeontol. 2003. V. 1. № 4. P. 227–330.
Goto M., Uyeno T., Yabumoto Y. Summary of Mesozoic elasmobranch remains from Japan // Mesozoic Fishes – Systematics and Paleoecology / Eds. Arratia G., Viohl G. München: Verlag Dr. F. Pfeil, 1996. P. 73–82.
Guinot G. Late Cretaceous elasmobranch palaeoecology in NW Europe // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2013. V. 388. P. 23–41.
Guinot G., Underwood C.J., Cappetta H., Ward D.J. Sharks (Elasmobranchii: Euselachii) from the Late Cretaceous of France and the UK // J. Syst. Palaeontol. 2013. V. 11. № 6. P. 589–671.
Guinot G., Carrillo-Briceño J.D. Lamniform sharks from the Cenomanian (Upper Cretaceous) of Venezuela // Cret. Res. 2018. V. 82. P. 1–20.
Hamm S.A. Systematic, stratigraphic, geographic, and paleoecological distribution of the Late Cretaceous shark genus Ptychodus within the Western Interior Seaway. Unpubl. MS thesis. Dallas, 2008. 434 p.
Hammer Ø. PAST. Paleontological Statistics. Version 3.21. Reference manual. Oslo: Univ. of Oslo, 2018. 264 p.
Kennedy W.J., King C., Ward D.J. The upper Albian and lower Cenomanian succession at Kolbay, eastern Mangyshlak (southwest Kazakhstan) // Bull. Inst. Roy. Sci. Natur. Belg. Sci. Terre. 2008. V. 78. P. 117–147.
Kiprijanoff V. Ueberreste von Fischen im Kurskischen eisenhaltigen Sandsteine (caмopoдъ) // Bull. Soc. imp. natur. Moscou. 1852. V. 25. № 2. P. 221–226.
Kiprijanoff V. Fisch-Ueberreste im Kurskischen eisenhaltigen Sandsteine (caмopoдъ) // Bull. Soc. imp. natur. Moscou. 1854. V. 27. № 4. P. 373–397.
Kiprijanoff V. Fisch-Ueberreste im Kurskischen eisenhaltigen Sandsteine (caмopoдъ) // Bull. Soc. imp. natur. Moscou. 1859. V. 32. № 1. P. 1–12.
Kiprijanoff V. Fish-Ueberreste im kurskischen eisenhaltigen Sandsteine oder Siwerischen Osteolith // Bull. Soc. imp. natur. Moscou. 1881. V. 55. № 3. P. 1–30.
Kriwet J., Klug S. Diversity and biogeography patterns of Late Jurassic neoselachians (Chondrichthys: Elasmobranchii) // Geol. Soc. London Spec. Publ. 2008. V. 295. P. 55–70.
Lebedev O.A., Zakharenko G.V. Global vertebrate-based palaeozoogeographical subdivision for the Givetian-Famennian (Middle-Late Devonian): Endemism-cosmopolitanism spectrum as an indicator of interprovincial faunal exchanges // Palaeoworld. 2010. V. 19. P. 186–205.
Mennad A., Tabuce R., Guinot G. et al. Découverte d’une nouvelle faune d’âge cénomanien dans la région de Forthassa (Atlas saharien occidental, Algérie): Implications biostratigraphiques et paléoenvironnementales // Ann. Paléontol. 2020. V. 106. № 1. 102355. https://doi.org/10.1016/j.annpal.2019.05.005
Müller A., Diedrich C.G. Selachier (Pisces, Chondrichthyes) aus dem Cenomanium von Ascheloh am Teutoburger Wald (Nordrhein-Westfalen, NW-Deutschland) // Geol. Paläontol. Westfalen. 1991. V. 20. P. 3–105.
Rage J.-C., Cappetta H. Vertebrates from the Cenomanian, and the geological age of the Draa Ubari fauna (Libya) // Ann. Paléontol. 2002. V. 88. P. 79–84.
Shimada K., Popov E.V., Siversson M. et al. A new clade of putative plankton-feeding sharks from the Upper Cretaceous of Russia and the United States // J. Vertebr. Paleontol. 2015. V. 35. № 5. https://doi.org/10.1080/02724634.2015.981335
Scotese C.R. Atlas of Late Cretaceous Paleogeographic Maps // PALEOMAP Atlas for ArcGIS. V. 2. The Cretaceous. Mollweide Projection. PALEOMAP Project. Evanston. IL. 2014. Maps 16–22.
Underwood C.J., Cumbaa S.L. Chondrichthyans from the Cenomanian (Late Cretaceous) bonebed, Saskatchewan, Canada // Palaeontology. 2010. V. 53. № 4. P. 903–944.
Underwood C.J., Goswami A., Prasad G.V.R. et al. Marine vertebrates from the ‘middle’ Cretaceous (early Cenomanian) of South India // J. Vertebr. Paleontol. 2011. V. 31. № 3. P. 539–552.
Verma O. Cretaceous vertebrate fauna of the Cauvery Basin, southern India: Palaeodiversity and palaeobiogeographic implications // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2015. V. 431. P. 53–67.
Vishnevskaya V.S., Kopaevich L.F. Microfossil assemblages as key to reconstruct sea-level fluctuations, cooling episodes and palaeogeography: The Albian to Maastrichtian of Boreal and Peri-Tethyan Russia // Geol. Soc. London Spec. Publ. 2020. V. 498. P. 165–187.
Vullo R., Cappetta H., Néraudeau D. New sharks and rays from the Cenomanian and Turonian of Charentes, France // Acta Palaeontol. Pol. 2007. V. 52. № 1. P. 99–116.
Vullo R., Bernárdez E., Buscalioni A.D. Vertebrates from the middle?–late Cenomanian La Cabaña Formation (Asturias, northern Spain): Palaeoenvironmental and palaeobiogeographic implications // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2009. V. 276. P. 120–129.
Vullo R., Courville P. Fish remains (Elasmobranchii, Actinopterygii) from the Late Cretaceous of the Benue Trough, Nigeria // J. Afr. Earth Sci. 2014. V. 97. P. 194–206.
Welton B.J., Farish R.F. The collector’s guide to fossil sharks and rays from the Cretaceous of Texas. Lewisville: Before Time, 1993. 204 p.
Werner C. Die Elasmobranchier-Fauna des Gebel Dist Member der Bahariya Formation (Obercenoman) der Oase Bahariya, Ägypten // Palaeo Ichthyol. 1989. V. 5. P. 1–112.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал