

Палеонтологический журнал, 2021, № 5, стр. 79-85
Древнейший представитель надсемейства Hagloidea (Orthoptera) из верхней перми России и замечания о кузнечикоподобных формах из палеозоя
А. В. Горохов *
Зоологический институт РАН
Санкт-Петербург, Россия
* E-mail: orthopt@zin.ru
Поступила в редакцию 17.02.2021
После доработки 07.03.2021
Принята к публикации 17.03.2021
Аннотация
Описан древнейший представитель надсемейства Hagloidea из верхней перми Кемеровской области России (Permohagla tomica gen. et sp. nov.). Это первое указание на формирование данного надсемейства еще в палеозое и косвенное подтверждение гипотезы о происхождении всех современных длинноусых прямокрылых с элитральным стридуляционным аппаратом (сверчковых и кузнечиковых) от ранних представителей Hagloidea. Предположение о происхождении Hagloidea, кузнечиковых и сверчковых от пермского “семейства” Raphoglidae syn. nov., изначально лишенного элитрального стридуляционного аппарата, опровергнуто, а само это “семейство” сведено в синонимы к подсемейству Kamiinae, принадлежащему пермскому семейству Pruvostitidae из инфраотряда Oedischiidea. Кратко рассмотрены также некоторые другие кузнечикоподобные формы из палеозоя, ошибочно отнесенные к позднемезозойскому и кайнозойскому надсемейству Tettigonioidea.
ВВЕДЕНИЕ
В монографии, посвященной разработке высшей классификации подотряда длинноусых прямокрылых (Ensifera) на основе обобщения всех доступных к тому времени данных по систематике современных и ископаемых прямокрылых (Горохов, 1995), этот подотряд был разбит на четыре инфраотряда и девять надсемейств, но для двух ископаемых семейств – Phasmomimidae из юры и Vitimiidae из мела – систематическое положение не было установлено, хотя их принадлежность к Ensifera поддерживалась. Такая классификация подотряда до сих пор представляется наиболее обоснованной и дана ниже с включением всех данных по геологическому распространению, известных до настоящего момента:
I. Инфраотряд Oedischiidea (верхний карбон–нижняя юра)
1) Надсемейство Oedischioidea (верхний карбон–нижняя юра)
2) Надсемейство Xenopteroidea (нижняя пермь–верхний триас)
II. Инфраотряд Elcanidea (нижняя пермь–палеоцен)
3) Надсемейство Permorhaphidioidea (нижняя пермь, ?пермь Мадагаскара)
4) Надсемейство Elcanoidea (нижняя пермь–палеоцен)
III. Инфраотряд Tettigoniidea (средний триас–современность)
5) Надсемейство Hagloidea (средний триас–современность)
6) Надсемейство Tettigonioidea (нижний мел–современность)
7) Надсемейство Stenopelmatoidea (нижний мел–современность)
IV. Инфраотряд Gryllidea (средний триас–современность)
7) Надсемейство Gryllavoidea (средний–верхний триас)
8) Надсемейство Grylloidea (верхний триас–современность).
Надсемейство Hagloidea рассматривается как предковое для остальных надсемейств инфраотряда Tettigoniidea и для инфраотряда Gryllidea, а его предками считаются представители надсемейства Oedischioidea из семейства Pruvostitidae, в частности, из его ранне- и среднепермского подсемейства Sylvoedischiinae. Надкрылья Sylvoedischiinae были очень похожи по строению на таковые большинства самок Hagloidea, поскольку они были лишены элитрального стридуляционного аппарата у обоих полов (рис. 1, а), тогда как надкрылья самцов Hagloidea всегда снабжены в той или иной степени развитым стридуляционным аппаратом, в котором стридуляционной жилкой стала изогнутая CuP (рис. 1, б, в). Последний признак был унаследован возможными потомками этого надсемейства, перечисленными выше.
Рис. 1.
Надкрылье представителей надсемейств Oedischioidea (а) и Hagloidea (б–г): а – Sylvoedischia crassa Gor., Sylvoedischiinae (Pruvostitidae), нижняя пермь Пермского края (по: Горохов, 1987); б – Haglomorpha martynovi Gor., Haglinae (Haglidae), самец, триас Кыргызстана (по: Горохов, 1986); в, г – Paracyrtophyllites undulatus Shar., Paracyrtophyllitinae (Tuphellidae), самец (в) и самка (г), верхняя юра Казахстана (по: Шаров, 1968). Без масштаба.
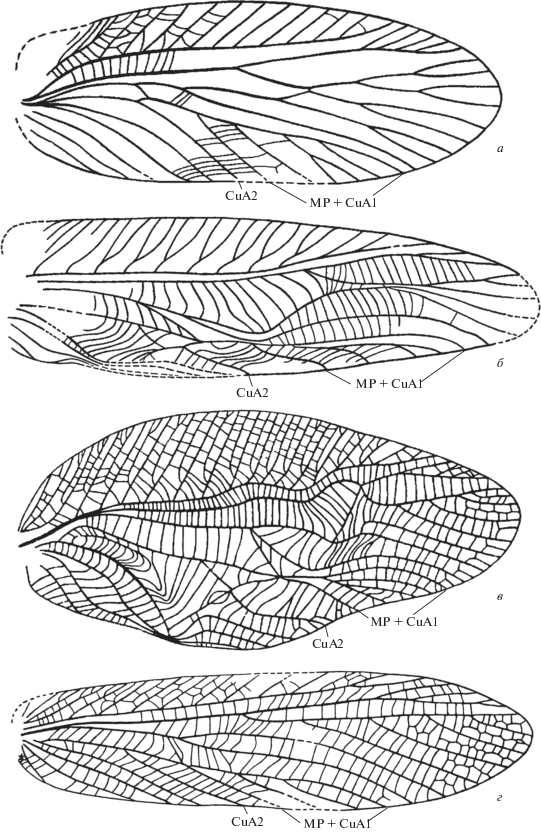
В качестве единственного признака, якобы позволяющего отличить надкрылья обоих полов общего предка Hagloidea, кузнечиковых и сверчковых, от надкрылий их более ранних предков (Bethoux et al., 2002), были названы некоторые особенности строения области MP + CuA1, а именно: нарушение правильной гребенчатости этих ветвей, характерной для Sylvoedischiinae (рис. 1, а), за счет смещения основания передней ветви MP + CuA1 проксимальнее оснований ряда последующих ветвей MP + CuA1 (рис. 1, б, в). Это преобразование, согласно А.В. Горохову (1995), могло быть вызвано переходом от более или менее геофильного образа жизни, при котором надкрылья складываются коробчато с резким сгибом вдоль ствола MP + CuA1 (рис. 1, а), к более фитофильному, для которого коробчатое складывание надкрылий не характерно (т.е., надкрылье в дистальной половине теряет такой сгиб и располагается почти в вертикальной плоскости), а затем у самцов в связи с развитием крупного стридуляционного аппарата надкрылья вновь начинают складываться коробчато, но на этот раз основной сгиб начинает проходить не вдоль ствола MP + CuA1 (т.е., не вдоль его передней ветви), а вдоль следующей ветви, которая в этом случае формирует новый ствол для остальных ветвей MP + CuA1 (рис. 1, б, в).
Последняя гипотеза предполагает, что у самок такое преобразование области MP + CuA1 могло отсутствовать (рис. 1, г) или проходить как частичная маскулинизация их жилкования (рис. 2, в) при усиленном развитии стридуляционного аппарата у самцов. Подтверждениями этой гипотезы являются наличие у всех самцов Hagloidea с развитым стридуляционным аппаратом и неукороченными крыльями смещения основания передней ветви MP + CuA1 в надкрыльях проксимальнее оснований других ветвей MP + CuA1 (рис. 1, б, в), а также сохранение в надкрыльях самок некоторых Hagloidea примитивного строения MP + CuA1, сходного с таковым Sylvoedischiinae (рис. 1, г). Обнаружение описываемого ниже древнейшего представителя Наgloidea в верхней перми является дополнительным подтверждением этой гипотезы, поскольку делает предположение о происхождении этой группы от форм почти того же возраста, но лишенных стридуляционного аппарата (Bethoux et al., 2002), как минимум маловероятным.
Рис. 2.
Надкрылье Hagloidea (а–в) и Oedischioidea (г): а – Permohagla tomica sp. nov. (Haglidae), голотип ПИН, № 4887/363, проксимальная половина надкрылья самца, верхняя пермь Кемеровской обл.; б – Hagloedischia primitiva Gor. (Hagloedischiidae), триас Кыргызстана (по: Горохов, 1986); в – Voliopinae gen. et sp. indet. (Haglidae), анальная часть надкрылья самки, триас Кыргызстана (по: Горохов, 1986); г – Kamia angustovenosa Mart., Kamiinae (Pruvostitidae), средняя пермь Татарстана (по: Шаров, 1968). Масштабная линейка относится лишь к рис. 2, а.

Изученный материал хранится в коллекции Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН) в Москве. За предоставленную возможность изучения этого материала автор благодарен коллективу лаборатории членистоногих ПИН. Данная работа выполнена в рамках Государственного исследовательского проекта № AAAA-A19-119020690101-6 (Российская Федерация).
НАДСЕМЕЙСТВО HAGLOIDEA HANDLIRSCH, 1906
Это надсемейство было разделено на четыре семейства с многочисленными подсемействами (Горохов, 1995): Hagloedischiidae (средний–верхний триас), Haglidae и Tuphellidae (средний триас–нижний мел), Prophalangopsidae (нижняя юра–современность). Описываемый ниже новый род из верхней перми не может быть включен в последнее семейство, поскольку надкрылья Prophalangopsidae обычно с хорошо развитой вторичной С, образованной из участков ветвей Sc и поперечных жилок между ними, а также со слегка ослабленным стридуляционным аппаратом у самца, в котором некоторые из его структур (расширенное ланцетовидное поле позади основания MA2, диагональная жилка или ее веер) в той или иной мере редуцированы, тогда как у нового рода в надкрылье самца нет даже следов вторичной C, а стридуляционный аппарат и все его структуры хорошо развиты (рис. 2, а). Признаки остальных семейств не противоречат тем, что обнаружены у нового рода; но его принадлежность к Tuphellidae все же кажется менее вероятной, так как у лучше сохранившихся самцов Tuphellidae поперечные жилки между CuA и CuP значительно сильнее изогнуты, а поле между МP + CuA1 и CuA2 более расширенное (рис. 1, в). Таким образом, новый род здесь отнесен к одному из двух оставшихся семейств, а именно, к Haglidae; причины такого выбора будут рассмотрены в замечаниях, приведенных после описания этого рода.
СЕМЕЙСТВО HAGLIDAE HANDLIRSCH, 1906
Род Permohagla Gorochov, gen. nov.
Название рода – от пермского периода и мезозойского рода Hagla.
Типовой вид – P. tomica sp. nov.
Диагноз. Надкрылье среднего для семейства размера. У самца оно со следующими особенностями (рис. 2, a): участки полей Sc-R и R‑M, расположенные проксимальнее MP, значительно более узкие, чем поле М-CuA в той же части надкрылья; поперечные жилки в основании последнего поля сильно S-образные и косые, а в его средней части почти прямые и поперечные; ланцетовидное поле между R и проксимальной частью MA1 сильно расширенное, с косыми поперечными жилками и, возможно, с MA2, ответвляющейся от MA недалеко от проксимального участка MP; поле между CuA и CuP заметно (но не очень сильно) расширено в проксимальной половине надкрылья, с более или менее дуговидными поперечными жилками и хорошо развитой диагональной жилкой, несущей небольшой веер у CuP и отделяющей большую часть CuA2 от его проксимального участка; проксимальный участок поля CuA1–CuA2 кпереди от диагональной жилки сравнительно крупный (широкий и длинный) и с довольно редким и несколько нарушенным жилкованием; более дистальная часть этого поля (между MP + CuA1 и CuA2) c густым и более правильным жилкованием, но слегка сужена в проксимальной части за счет дуговидного изгиба и значительного смещения основания дистального участка CuA2 к MP + CuA1 вдоль диагональной жилки; CuP умеренно изогнутая и формирует довольно пологую стридуляционную жилку в проксимальной половине, а также отчетливо дуговидную переднюю струну в дистальной части; 1А еще менее изогнутая, но также формирует дуговидную струну в дистальной части; 2А с двумя ветвями, более анальная из которых участвует в формировании примитивного плектрума.
Видовой состав. Типовой вид.
Сравнение. Подсемейственная принадлежность этого рода неясна, поскольку неизвестно строение дистальной части ланцетовидного поля и образующих эту часть жилок, но от родов, входящих в состав разных подсемейств Haglidae, новый род отличается следующими признаками: от Hagloptera Gorochov, 1986 (единственного рода триасового подсемейства Haglopterinae) – более пологой стридуляционной жилкой, значительно меньшим числом ветвей в веере диагональной жилки, значительно более коротким полем между CuA2 и передней струной, а также более узким полем между MP + CuA1 и CuA2 около диагональной жилки, но кзади от нее; от родов подсемейства Haglinae (триас–нижний мел) – более широким проксимальным участком поля CuA1–CuA2 с более беспорядочным жилкованием, явно более пологой стридуляционной жилкой и часто отсутствием дужки (характерной сильно дуговидной поперечной жилки) в поле между MP + CuA1 и CuA2 у диагональной жилки; от родов Triassaginae (триас) – теми же признаками поля CuA1–CuA2 и значительно более широким ланцетовидным полем; от родов Voliopinae (триас–юра) – основанием дистальной части CuA2, значительно сдвинутым относительно проксимальной части этой жилки вдоль диагональной жилки и теми же признаками, как от Haglinae; от Isfaropterinae, Bachariinae, Cyrtophyllitinae и Angarohaglinae (юра и/или нижний мел) – отсутствием следов даже зачатка вторичной C и теми же признаками, как от Voliopinae (кроме дужки в поле между MP + CuA1 и CuA2, которая, видимо, отсутствует у всех этих таксонов, но развита у Voliopinae).
Замечания. Если допустить, что семейство Hagloedischiidae c двумя родами из триаса (Hagloedischia Gorochov, 1986 и Voltziahagla Marchal-Papier, Nel et Grauvogel-Stamm, 2000) было описано не по самцу с первично слаборазвитым элитральным стридуляционным аппаратом, а по значительно маскулинизированному надкрылью самки, то новый род вполне может оказаться близким к Hagloedischia, a Hagloedischiidae могут быть понижены в ранге до подсемейства в Haglidae. Однако, едва ли Hagloedischiidae можно синонимизировать с каким-либо уже известным подсемейством Haglidae, поскольку перестройка жилкования области ветвей MP + CuA1 у Hagloedischia находится лишь в начальной стадии (рис. 2, б), тогда как у обоих полов в других группах Haglidae эта перестройка выражена значительно лучше (рис. 1, б; 2, в).
Permohagla tomica Gorochov, sp. nov.
Название вида – от реки Томь.
Голотип – ПИН, 4887/363, прямой и обратный отпечатки передней половины надкрылья самца; Россия, Кемеровская обл., Новокузнецкий р-н, правый берег р. Томь в 10 км ниже пос. Усть-Булак, местонахождение Бабий Камень; верхняя пермь, чансиньский ярус (Yan et al., 2020), мальцевская свита, слой 87–90; сб. А. Башкуева и Д. Василенко, 2013 г.
Описание (рис. 2, а; 3). Окраска не сохранилась. Костальное поле примерно в 1.5 раза уже остальной части надкрылья; ветви Sc в проксимальной половине надкрылья отчетливо косые и почти не изогнуты; поля Sc-R и R-M в этой же части надкрылья узкие, но приблизительно равные по ширине; поле M-CuA в наиболее широкой части примерно вдвое шире сохранившихся частей полей Sc-R и R-M; поле CuA-CuP в наиболее широкой части почти вдвое шире наиболее широкой части предыдущего поля, с довольно густыми поперечными жилками и трехветвистым веером диагональной жилки между CuA2 и CuP.
Рис. 3.
Permohagla tomica sp. nov. (Haglidae), голотип ПИН, № 4887/363, отпечаток проксимальной половины надкрылья самца; Кемеровская обл., местонахождение Бабий Камень; верхняя пермь. Масштабная линейка соответствует 5 мм.

Размеры в мм: длина отпечатка 13.5; предполагаемая длина надкрылья 20–25.
Материал. Голотип.
НАДСЕМЕЙСТВО OEDISCHIOIDEA HANDLIRSCH, 1906
Это надсемейство включает в себя древнейших (позднекарбоновых, пермских, триасовых и раннеюрских) длинноусых прямокрылых, изначально лишенных элитрального стридуляционного аппарата у обоих полов, но габитуально похожих на представителей Hagloidea и современных кузнечиковых и лжекузнечиковых. Возможно, представители Oedischioidea являлись их экологическими предшественниками. Надсемейство было подразделено на пять семейств и 17 подсемейств (Горохов, 1995), среди которых были триасовое семейство Mesoedischiidae, начавшее независимо от Hagloidea формировать элитральный стридуляционный аппарат, негомологичный таковому инфраотрядов Tettigoniidae и Gryllidea, а также пермское семейство Pruvostitidae, к которому принадлежит подсемейство Sylvoedischiinae – наиболее вероятная предковая группа для инфраотрядов Tettigoniidea и Gryllidea.
К последнему семейству принадлежит также подсемейство Kamiinae (нижняя и средняя пермь), в строении надкрылий которого наблюдаются элементы имитации древесных или кустарниковых листьев, что делает их отдаленно похожими на многих современных кузнечиковых – имитаторов листьев. К таким элементам относятся некоторое расширение надкрылья и появление легкой беспорядочности в его жилковании (продольное складывание надкрылий почти исчезает – они становятся расположенными в основном в вертикальной плоскости и имитируют листья, вследствие чего функциональный смысл ряда жилок как границ между сгибающимися частями надкрылий утрачивается), включая появление некоторой беспорядочности в строении области ветвей MP + CuA1, при которой эти ветви утрачивают гребенчатое расположение, и эта область по строению начинает несколько напоминать таковую большинства Hagloidea (рис. 2, г).
Примерно такие же особенности характеризуют пермский род Raphogla Bethoux, Nel, Lapeyrie, Gand et Galtier, 2002, трактуемый его авторами как древнейший представитель “clade of modern Ensifera” (Bethoux et al., 2002). Для этого рода ими создано отдельное семейство, которое они, видимо, считают предковым для инфраотрядов Tettigoniidea и Gryllidea. Но значительно более вероятно, что этот род следует поместить в подсемейство Kamiinae, a установленное для него “семейство” целесообразно свести в синонимы к этому подсемейству (Kamiinae Sharov, 1968 = Raphoglidae Bethoux, Nel, Lapeyrie, Gand et Galtier, 2002, syn. nov.).
Ряд других палеозойских форм были включены или предположительно включены их авторами в надсемейство Tettigonioidea, достоверно известное лишь из раннего мела и кайнозоя, но у этих форм не найдено ни одного признака, который бы позволил отнести их к этому надсемейству, а не к инфраотряду Oedischiidea. Они явно не принадлежат Tettigonioidea, а могут быть представителями Kamiinae или какой-либо иной группы их возможных экологических аналогов из Oedischiidea. К ним относятся загадочный род Permotettigonia Nel et Garrouste, 2016 из средней перми, описанный по недостаточно сохранившемуся фрагменту надкрылья, и созданное для этого рода семейство Permotettigoniidae Nel et Garrouste, 2016 (Garrouste et al., 2016), а также еще более загадочный пермский род Permophyllum Prokop, Szwedo, Lapeyrie, Garrouste et Nel, 2015, который был помещен в Orthoptera без уточнения его систематического положения, но, судя по подписям к иллюстрациям этого рода, с указанием на его возможную принадлежность к Tettigoniidae (Prokop et al., 2015).
Список литературы
Горохов А.В. Триасовые прямокрылые надсемейства Hagloidea (Orthoptera) // Тр. Зоол. ин-та АН СССР. 1986. Т. 143. С. 65–100.
Горохов А.В. Пермские прямокрылые инфраотряда Oedischiidea (Orthoptera, Ensifera) // Палеонтол. журн. 1987. № 1. С. 62–75.
Горохов А.В. Система и эволюция прямокрылых подотряда Ensifera (Orthoptera). Части 1 и 2 // Труды Зоологического института РАН. 1995. Т. 260. С. 1–224; 1–213.
Шаров А.Г. Филогения ортоптероидных насекомых // Тр. Палеонтол. ин-та АН СССР. 1968. Т. 118. С. 1–218.
Bethoux O., Nel A., Lapeyrie J. et al. Raphogla rubra gen. n., sp. n., the oldest representative of the clade of modern Ensifera (Orthoptera: Tettigoniidea, Gryllidea) // Eur. J. Entomol. 2002. V. 99. P. 111–116.
Garrouste R., Hugel S., Jasquelin L. et al. Insect mimicry of plants dates back to the Permian // Nature Communications. 2016. V. 7. 13735. https://doi.org/10.1038/ncomms13735
Prokop J., Szwedo J., Lapeyrie J. et al. New Middle Permian insects from Salagou Formation of the Lodève Basin in southern France (Insecta: Pterygota) // Ann. Soc. Entomol. Fr. (Nouv. Sér.). 2015. V. 51. № 1. P. 14–51. https://doi.org/10.1080/00379271.2015.1054645
Yan E.V., Beutel R.G., Lawrence J.F. et al. Archaeomalthus (Coleoptera, Archostemata), a ‘ghost adult’ of Micromalthidae from Upper Permian deposits of Siberia? // Histor. Biol. 2020. V. 32. № 8. P. 1019–1027. https://doi.org/10.1080/08912963.2018.1561672
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал