

Палеонтологический журнал, 2021, № 1, стр. 70-76
Первые данные об остракодах келловея Центрального Дагестана
Л. А. Глинских a, *, Е. М. Тесакова b, c, **
a Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
b Московский государственный университет имени М.В. Ломоносова
Москва, Россия
c Геологический институт РАН
Москва, Россия
* E-mail: glor@mail.ru
** E-mail: ostracon@rambler.ru
Поступила в редакцию 12.08.2019
После доработки 20.08.2019
Принята к публикации 20.08.2020
Аннотация
Впервые в келловее (аммонитовые зоны Calloviense, Jason, Coronatum и Athleta) Центрального Дагестана выявлены и изучены остракоды. Комплекс насчитывает девять видов, один – Pleurocythere khapissovi sp. nov. – описан как новый. Для ранее известных форм приведены изображения.
ВВЕДЕНИЕ
Целью статьи является описание и изображение остракод из келловея разреза Цудахар (Центральный Дагестан) (рис. 1), перспективного для проведения бореально-тетической корреляции, так как в нем встречены комплексы аммонитов, содержащие представителей разного биогеографического происхождения (Гаврилов и др., 2015).
Обнажение у моста на правом берегу р. Казикумухское Койсу, в 500 м выше с. Цудахар, является классическим разрезом келловейских отложений, где полностью представлена армхинская свита (нижний келловей – низы верхнего келловея), включая ее несогласные контакты с подстилающей цудахарской и вышележащей иронской свитами (рис. 2). Этот разрез был мультидисциплинарно изучен в 2014–2015 гг. большим коллективом специалистов из Москвы, Ярославля и Новосибирска. Подробное описание литологии и комплексов ископаемых (аммониты, белемниты, двустворки), биостратиграфия, а также изображение характерных фораминифер и седиментологическая интерпретация приведены в “Путеводителе …” (Гаврилов и др., 2015).
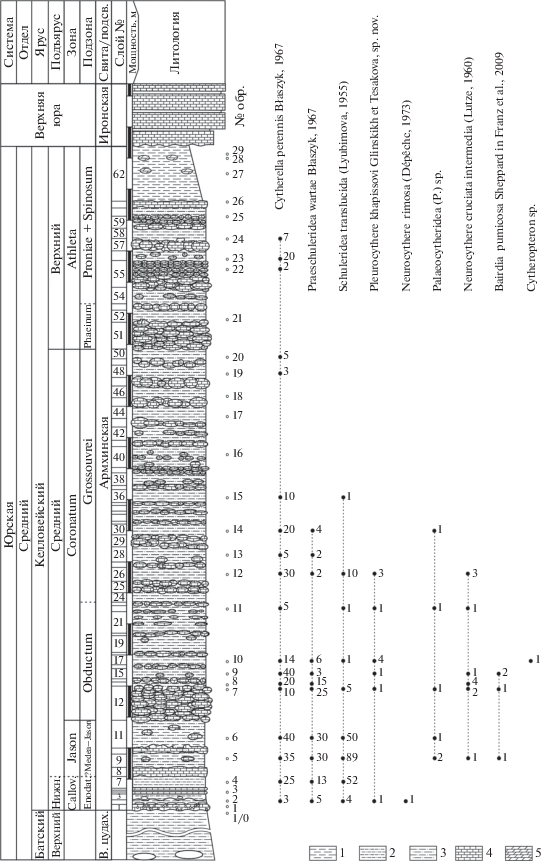
Рис. 2.
Распространение остракод в армхинской свите (келловей) разреза Цудахар (литологическая колонка и расчленение разреза по аммонитам приводится по: Гаврилов и др., 2015). Обозначения: 1 – глина; 2 – глина алевритовая; 3 – алевролит; 4 – известняк; 5 – мергель.
Необходимость исследования цудахарских остракод, встреченных в комплексе микрофауны совместно с фораминиферами, обусловлена тем, что юрские остракоды Дагестана до сих пор практически не изучались, за редким исключением (Касимова и др., 1956), где приведено определение (без описаний и изображений) трех видов из верхнего аалена и одного, в открытой номенклатуре, из байоса.
МАТЕРИАЛ И МЕТОДЫ
Материалом для исследования послужила коллекция остракод, полученная в результате обработки проб, отобранных Л.А. Глинских в 2015 г. в составе полевого отряда под руководством А.П. Ипполитова (ГИН РАН, Москва) из армхинской свиты (келловей) близ с. Цудахар.
Образцы были обработаны по стандартной методике. Относительно рыхлые разности пород (пробы навеской 200 г) отмачивались в воде в течение одной–двух недель, а затем дезинтегрировались простым кипячением в течение нескольких часов. Более твердые породы разрушались с использованием расплава гипосульфита натрия (Основы…, 1960). После этого осадок отмывался в проточной воде через сито с диаметром ячеи 56 мкм и сушился. После технической обработки образцов проводился отбор и изучение микрофауны под стереомикроскопом Stemi 2000-С (Carl Zeiss). Всего было обработано 29 образцов, ракушковые найдены в 18 из них. Выбрано порядка 680 экземпляров, принадлежащих девяти видам из восьми родов (рис. 2).
Подавляющее большинство изученных остракод Цудахара (сотни раковин и створок) принадлежит к широко распространенным и эврибионтным родам Cytherella Jones, 1849, Praeschuleridea Bate, 1963 и Schuleridea Swartz et Swain, 1946, не представляющим интерес с точки зрения биостратиграфии. Неоправданно большой стратиграфический диапазон ряда относящихся к ним видов (как и многих других фанерозойских остракод) может быть связан со сборными таксонами. Обычно к подобным видам ошибочно относят остракод с весьма ограниченным набором диагностических признаков (гладких или без ярко выраженной скульптуры, с простыми примитивными замками и т.п.). Правильный подход к выделению реальных палеонтологических видов внутри таких групп – это прослеживание в онтогенезе изменений замка, мускульных отпечатков, простых и/или ситовидных пор и морфометрических параметров отдельно левых и правых створок. Безусловно, подобные работы есть (напр., Błaszyk, 1959, 1967; Herrig, 1966 и др.), но они достаточно трудоемки и не всегда осуществимы, поскольку палеонтологический материал должен отличаться очень хорошей сохранностью и обилием разрозненных створок, иначе нельзя изучить их внутреннее строение. Кроме того, остракоды должны быть представлены экземплярами всех (или хотя бы большинства) возрастных стадий, притом статистически достоверными выборками. Перечисленные условия далеко не всегда отвечают реальному материалу (как в нашем случае), сохранность которого во многом зависит от фаций, его обилие – от трофности палеобассейна, а совместная встречаемость взрослых и ювенильных экземпляров – от особенностей строения ареалов этих видов и типов захоронений. Поэтому виды, определенные нами в этих родах [Cytherella perennis Błaszyk, 1967, Praeschuleridea wartae Błaszyk, 1967 и Schuleridea translucida (Lyubimova, 1955)], понимаются в широком смысле (s.l.).
Для лучшего представления развития остракод в этом регионе ниже приведен краткий обзор стратиграфического и географического распространения ранее известных видов.
Cytherella perennis s.l. (табл. VII , фиг. 1–3 ; см. вклейку) является доминирующим элементом комплекса в нижнем и среднем келловее Цудахара (зона Calloviense – нижняя половина зоны Coronatum), а в верхах среднего и в верхнем келловее (верхи зоны Coronatum – зона Athleta) выявлены монотаксонные комплексы, представленные этим видом (рис. 2). Распространение C. perennis известно в средней и верхней юре. Вид широко представлен в верхнем байосе и бате Польши (Błaszyk, 1967; Bielecka et al., 1988; Tesakova et al., 2008); встречен в верхнем байосе северо-востока Украины (Пяткова, Пермякова, 1978); в нижнем–среднем келловее севера Франции (Dépêche, 1969); в среднем келловее–среднем оксфорде Европейской России (Герасимов и др., 1996; Tesakova, 2003, 2008; Тесакова и др., 2017).
Представители Praeschuleridea wartae s.l. (табл. VII , фиг. 4, 5 ) в большом количестве встречены в нижнем – среднем келловее (зона Calloviense – нижняя половина зоны Coronatum) изученного разреза (рис. 2). Вид также известен из бата Польши (Błaszyk, 1967; Bielecka et al., 1988), нижнего–среднего келловея Европейской России (Tesakova, 2003, 2013; Тесакова и др., 2009).
Еще один среднеюрский вид, преобладающий в изученной коллекции, Schuleridea translucida s.l. (табл. VII , фиг. 7, 8 ), распространен в среднем–верхнем келловее Европейской России (Любимова, 1955, 1956; Герасимов и др., 1996; Tesakova, 2003, 2008; Тесакова и др., 2017) и нижнем келловее северо-востока Украины (Пяткова, Пермякова, 1978). В разрезе Цудахар он найден в нижнем–среднем келловее (зона Calloviense – нижняя половина зоны Coronatum) (рис. 2).
Представители Bairdia pumicosa Sheppard in Franz et al., 2009 (табл. VII , фиг. 6 ) в изученном разрезе встречены в среднем келловее (зона Jason и низы зоны Coronatum) (рис. 2) в виде трех створок и одной целой раковины удовлетворительной сохранности. Этот вид известен в бате северо-запада Франции (Sheppard, 1981a, b; Dépêche, 1984, 1985), нижнем бате юго-запада Англии (Sheppard, 1981a, b), бате и келловее Южной Германии (Buck, 1954; Schudack, Schudack, 2000; Franz et al., 2009, 2014; Beher et al., 2010), а также нижнем бате и нижнем келловее Польши (Tesakova, 2008).
Особый интерес представляет Neurocythere cruciata intermedia (Lutze, 1960) (табл. VII , фиг. 10, 11 ), являющийся зональным видом биостратиграфических шкал по остракодам, разработанных в разных странах Европы. Вид распространен в нижнем келловее – нижнем оксфорде Южной Германии (Buck, 1954; Huber et al., 1987; Franz et al., 2009, 2014; Beher et al., 2010; Dietze et al., 2017); в среднем келловее восточной части Нидерландов и северо-востока Украины (Пяткова, Пермякова, 1978; Herngreen et al., 1983); в среднем и верхнем келловее Северо-Восточной Германии (Lutze, 1960); в среднем–верхнем келловее и нижнем оксфорде Шотландии и Англии (Whatley, 1970; Kilenyi, 1978; Whatley et al., 2001; Whatley, Ballent, 2004; Wilkinson, Whatley, 2009); в верхнем келловее севера Франции и Польши (Oertli, 1963; Bielecka et al., 1988; Olempska, Błaszyk, 2001). В разрезе Цудахар представители данного вида встречены в среднем келловее (зона Jason и нижняя часть зоны Coronatum) в виде четырех раковин и восьми створок хорошей и удовлетворительной сохранности (рис. 2).
Neurocythere rimosa (Dépêche, 1973) (табл. VII , фиг. 9 ), известный из нижнего–среднего бата северо-запада Франции и всего бата южной части Англии (Dépêche, 1973, 1984, 1985; Sheppard, 1981b; Bate, 2009), а также нижнего–среднего бата Южной Германии (Franz et al., 2014), представлен в изученном разрезе в нижнем келловее (зона Calloviense) в виде одной раковины плохой сохранности (рис. 2).
Наряду с ранее известными формами, в нижнем–среднем келловее разреза Цудахар (зоны Calloviense – Coronatum) найдено 10 целых раковин и две створки плевроцитер хорошей и удовлетворительной сохранности, отнесенных авторами к новому виду Pleurocythere khapissovi sp. nov. (табл. VII , фиг. 13–15 ; рис. 2). Подробное описание таксона приведено ниже.
Неудовлетворительная сохранность единичных раковин и створок Palaeocytheridea (P.) sp. (табл. VII , фиг. 12 ) и Cytheropteron sp. из среднего келловея разреза Цудахар (рис. 2) не позволила определить видовую принадлежность найденных экземпляров.
Фотографирование остракод проводилось при помощи электронного сканирующего микроскопа в ИГМ СО РАН, а их изображения приведены на фототаблице.
Авторы используют систему надродовых таксонов, принятую в “Практическом руководстве…” (1999). При описании вида применялись следующие сокращения: Д – длина раковины, В – высота раковины, Т – толщина раковины.
Коллекция № DZ-19 хранится в лаб. микропалеонтологии Ин-та нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
ОТРЯД PODOCOPIDA
ПОДОТРЯД CYTHEROCOPINA
НАДСЕМЕЙСТВО PROGONOCYTHERACEA SYLVESTER-BRADLEY, 1948
СЕМЕЙСТВО PLEUROCYTHERIDAE MANDELSTAM, 1960
Род Pleurocythere Triebel, 1951
Pleurocythere khapissovi Glinskikh et Tesakova, sp. nov.
Табл. VII , фиг. 13–15
Название вида в честь энтузиаста палеонтологии Омара Хаписова.
Голотип – ИНГГ СО РАН, № DZ-19/13, целая раковина самки; Центр. Дагестан, с. Цудахар; средняя юра, нижний келловей, аммонитовая зона Calloviense (армхинская свита).
Описание. Раковина средних размеров, продолговато-овальная, умеренно выпуклая. Левая створка немного больше правой. Охват более выражен на заднеспинном крае. Передний конец раковины высокий, закругленный, с небольшим замочным ушком на левой створке, немного скошен со спинной стороны. Задний конец раковины ниже переднего, округло-треугольной формы, не симметричный (сильнее скошен сверху), на левых створках закруглен более плавно. Спинной край прямой, наклонен к заднему концу раковины. Соединяется с передним и задним концом плавно. Брюшной край прямой, не параллелен спинному, вогнут в передней трети. Наибольшая длина – в нижней трети раковины. Наибольшая высота – на переднем конце, а наибольшая толщина – в заднебрюшной части раковины. На поверхности створок развиты отчетливые ребра: четыре продольных и одно косое. Самое длинное из продольных ребер – верхнее брюшное – расположено в нижней трети раковины, пересекает оба конца и достигает переднего края (но не заднего). Ниже него на брюшной стороне наблюдается еще одно ребро, гораздо более короткое, не пересекающее концов. Оно нависает над брюшным краем, из-за чего создается ложное впечатление выпуклости брюшного края при виде сбоку. Своими концами, приподнятыми кверху, нижнее брюшное ребро сочленяется с верхним брюшным. Третье продольное ребро – срединное – расположено практически горизонтально в средней части створки. На заднем конце оно плавно отклоняется к спинной стороне, а в передней трети створки косо стремится книзу, но далеко не доходит до края переднего конца. От этой косой части срединного ребра отходят два коротких отростка, верхний – по направлению к переднему замочному ушку, нижний плавно отгибается к брюшной стороне, образуя маленькую открытую петлю. В задней части срединного ребра также наблюдается тонкий короткий отросток, отходящий от петли и наискось пересекающий задний конец по направлению к брюшной стороне. Четвертое и самое короткое из продольных ребер расположено вдоль спинного края в задней половине створки. На заднем конце оно плавно сопрягается со срединным ребром, образуя хорошо выраженную открытую петлю, а ближе к переднему концу истончается и распадается на 2–3 фрагмента. На левых створках спинное ребрышко короче, чем на правых. Последнее ребро, тонкое и косое, наблюдается на переднем конце – пересекает всю его верхнюю половину, достигая переднего края, и по большой дуге плавно переходит на спинную сторону. Срединное ребро может соединяться с косым посредством своего верхнего отростка. Вся межреберная поверхность створок, кроме уплощенной части переднего и заднего концов, покрыта крупными пяти- или шестигранными ячейками. Между срединным и верхним брюшным ребрами насчитывается четыре ряда ячеек. Толщина граней ячеек немногим меньше таковой ребер.
Размеры, мм:
| № экз. | Д | В | Т |
|---|---|---|---|
| Голотип № DZ-19/13 | 0.53 | 0.31 | |
| DZ-19/14 | 0.50 | 0.3 | |
| DZ-19/15 | 0.53 | 0.3 | |
| DZ-19/17 | 0.66 | 0.35 | 0.25 |
Изменчивость. Передние отростки срединного ребра могут быть более или менее развитыми, вследствие чего сплошная перемычка между косым и срединным ребрами имеется или отсутствует. Задний отросток срединного ребра может быть либо тонким и малозаметным, либо отчетливо выраженным. Слабо варьирует толщина граней ячеек.
Половой диморфизм выражен в большей длине раковин самцов при одинаковой высоте с раковинами самок.
Сравнение. Наиболее схожий по очертанию, размерам раковины, расположению ребер на поверхности створок и ячеистой межреберной скульптуре является вид P. caudata (uzbekistanensis) (Masumov, 1970) из нижнего келловея Узбекистана. Этот таксон был описан А.С. Масумовым из разреза Яккабаг (юго-западные отроги Гиссарского хребта) (1973, с. 55, табл. 5 , фиг. 2) и, по мнению авторов, ошибочно отнесен к роду Platylophocythere Oertli, 1959. От него новый вид отличается более коротким, тонким и распадающимся в передней части спинным ребром, которое не соединяется в первой трети створки ни с переднеспинным косым ребром (что отмечено в описании Масумова на с. 56, но не просматривается на табл. 5 , фиг. 2), ни со срединным (что наблюдается на табл. 5 , фиг. 2, но не отмечено в описании), но соединяется характерной плавной петлей со срединным ребром в задней трети створки (чего не наблюдается на табл. 5 , фиг. 2). Также важным отличием от P. caudata (uzbekistanensis) служит более короткое срединное ребро, которое никак не соединяется с верхним брюшным и имеет короткие отростки в передней части, отходящие от него под иными углами, чем у сравниваемого вида.
Кроме вышеупомянутого, известен еще один ранне- и среднекелловейский вид P. kurskensis Tesakova, 2009 из Курской обл. России (Тесакова и др., 2009, с. 31, табл. 3 , фиг. 1–3 , 5–8 ; Tesakova, 2013, табл. 6 , фиг. 11 ). От курских экземпляров описываемый вид достаточно сильно отличается меньшей длиной раковины, более тонкими ребрами, коротким спинным ребром, четырехрядной ячеистой (а не двурядной ямчатой) межреберной скульптурой, а также тем, что срединное и верхнее брюшные ребра не соединяются.
От P. elliptica Błaszyk, 1967 из верхнего байоса–среднего бата Польши (Błaszyk, 1967, с. 24, табл. 6 , фиг. 1–4 , рис. 5 ; Bielecka et al., 1988, с. 173, табл. 68 , фиг. 2), нижнего бата – нижнего келловея Германии (Brand, 1990, с. 202, табл. 12 , фиг. 17–20 ; Franz et al., 2009, с. 142, табл. 3 , фиг. 1; Beher et al., 2010, с. 50), весьма похожего ячеистой скульптурой и петлевидной формой сочленения срединного и спинного ребер, отличается меньшей длиной этих ребер, отсутствием соединения срединного и брюшного ребер, а также более длинным косым ребрышком.
От P. ohmerti Franz, Ebert et Stulpinaite, 2018 из среднего аалена – нижнего байоса Южной Германии (Franz et al., 2018, с. 76, табл. 5 , фиг. 9–12 ), сходного по ячеистой межреберной скульптуре, плавному петлевидному сочленению спинного ребра и срединного и по тонкому короткому спинному ребру левой створки, отличается продолговато-овальным (а не округло-прямоугольным) очертанием раковины сбоку, присутствием переднего замочного ушка, более тонким и продолжающимся на спинной стороне косым ребрышком, а также более низким и треугольным задним концом.
От всех остальных байосских и батских плевроцитер, сходных по ячеистой межреберной скульптуре, сравниваемый вид отличается характерной петлей в задней трети створки при плавном сочленении срединного ребра и спинного, а также субвертикальным положением переднего верхнего отростка срединного ребра.
Распространение. Средняя юра; нижний и средний келловей Центрального Дагестана.
Материал. 10 целых раковин и две створки хорошей и удовлетворительной сохранности из нижнего (зона Calloviense)–среднего (зона Coronatum) келловея разреза Цудахар (армхинская свита).
ЗАКЛЮЧЕНИЕ
Впервые в келловее (армхинская свита) Центрального Дагестана выявлен и изучен комплекс остракод, насчитывающий девять форм, семь из которых определены до вида, две – до рода; один вид – Pleurocythere khapissovi – описан как новый. Для ранее известных видов приведены изображения.
Анализ обзора хорологического и стратиграфического распространения цудахарских остракод показал, что в изученном комплексе встречены представители разного биогеографического происхождения, что является перспективным аспектом для проведения межрегиональной, а, возможно, и бореально-тетической корреляции по данной группе микрофауны.
* * *
Авторы выражают искреннюю признательность Л.М. Мельниковой (ПИН РАН, Москва), Б.Н. Шурыгину (ИНГГ СО РАН, Новосибирск) за советы и рекомендации при подготовке рукописи к печати, Н.Г. Изох (ИНГГ СО РАН) и И.В. Аборневой (ИГМ СО РАН, Новосибирск) за помощь в проведении съемки.
Работа выполнена в рамках тем госзадания №№ 0135-2019-0062 (ГИН РАН), АААА-А16-116033010096-8 (МГУ), а также является вкладом в проекты ФНИ № 0331-2019-0005 и РФФИ № 18-05-00501, № 19-05-00130.
Список литературы
Гаврилов Ю.О., Захаров В.А., Рогов М.А. и др. Цудахар // Юрские отложения центральной части Горного Дагестана. Путеводитель геологических экскурсий VI Всеросс. совещ. “Юрская система России: проблемы стратиграфии и палеогеографии”, 15–20 сентября 2015 г. / Ред. Черкашин В.И. Махачкала: “АЛЕФ”, 2015. С. 81–105.
Герасимов П.А., Митта В.В., Кочанова М.Д., Тесакова Е.М. Ископаемые келловейского яруса Центральной России. М., 1996. 126 с.
Касимова Г.К., Кузнецова З.В., Михеева З.Ф. Микрофауна юрских отложений разреза Уллучай (Центральный Дагестан) // Докл. АН АзССР. 1956. Т. 12. № 1. С. 9–13.
Любимова П.С. Остракоды мезозойских отложений Среднего Поволжья и Общего Сырта // Тр. ВНИГРИ. Нов. сер. 1955. Вып. 84. С. 3–190.
Любимова П.С. Триасовые и юрские остракоды восточных районов Украины // Тр. ВНИГРИ. Нов. сер. 1956. Вып. 98. С. 533–583.
Масумов А.С. Юрские остракоды Узбекистана. Ташкент: Фан, 1973. 197 с.
Основы палеонтологии. Членистоногие – трилобитообразные и ракообразные. М.: Изд-во АН СССР, 1960. 519 с.
Практическое руководство по микрофауне СССР. Том 7. Остракоды мезозоя / Ред. Соколов Б.С. СПб.: Изд-во ВСЕГЕИ, 1999. 244 с.
Пяткова Д.М., Пермякова М.Н. Фораминиферы и остракоды юры Украины. Киев: Наук. думка, 1978. 288 с.
Тесакова Е.М., Cтреж А.С., Гуляев Д.Б. Новые остракоды из нижнего келловея Курской области // Палеонтол. журн. 2009. № 3. С. 25–36.
Тесакова Е.М., Шурупова Я.А., Устинова М.А. Стратиграфия келловея и нижнего оксфорда разреза Михайловцемент (Рязанская обл.) по микрофауне и наннопланктону // Тр. ГИН РАН. 2017. Вып. 615. С. 264–300.
Bate R.H. Middle Jurassic (Aalenian–Bathonian) // Ostracods in British Stratigraphy / Eds. Whittaker J.E., Hart M.B. L., 2009. P. 199–223 (The Micropalaeontological Society, Spec. Publ. Geol. Soc. London).
Beher E., Brand E., Franz M. Bathonian and Lower Callovian ostracods of Albstadt-Pfeffingen (Middle Jurassic, Baden-Württemberg, Germany) // Palaeodiversity. 2010. V. 3. P. 43–57.
Bielecka W., Styk O., Błaszyk J., Kopik J. Middle Jurassic. Ostracoda Latreille, 1806 // Geology of Poland. Atlas of guide and characteristic fossils. Pt 2b. Mesozoic Jurassic / Ed. L. Malinowska. Warsaw, 1988. P. 165–179.
Błaszyk J. Two new bathonian ostracods of the genus Progonocythere // Acta Palaeontol. Pol. 1959. V. 4. № 4. P. 431–453.
Błaszyk J. Middle Jurassic ostracods of the Czestochowa region (Poland) // Acta Palaeontol. Pol. 1967. V. 12. № 1. P. 1–75.
Brand E. Biostratigraphische Untergliederung des Ober-Bathonium im Raum Hildesheim, Nordwestdeutschland mittels Ostracoden und Korrelation ihrer Vertikalreichweiten mit Ammoniten-Zonen // Geol. Jb. A. 1990. Bd 121. S. 119–273.
Buck E. Stratigraphisch wichtige Ostrakoden im Lias u. Dogger SW-Deutschlands; Freiburg i. Br. 1954 [unpublished table].
Dépêche F. Description de quelques ostracodes nouveaux de Bathonien Lorrain // Rev. Micropaléontol. 1969. V. 2. P. 107–118.
Dépêche F. Étude des Ostracodes du Dogger du Bassin Parisien. I. Ostracodes du Bathonien inférieur et de la base du Bathonien moyen de Port-en-Bessin // Rev. Micropaléontol. 1973. V. 4. P. 213–226.
Dépêche F. Les ostracodes d’une plate-forme continentale au Jurassique: recherches sur le Bathonien du Bassin Parisien // Mém. Sci. Terre. Univ. Pierre-et-Marie-Curie, Paris. Paris, 1984. No 84 38. P. 1–419 [unpublished thesis].
Dépêche F. Lias supérieur, Dogger, Malm // Atlas des ostracodes de France // Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine, Mém. 1985. № 9. P. 119–145.
Dietze V., Franz M., Kutz M., Waltschew A. Stratigraphy of the Middle Jurassic Sengenthal Formation of Polsingen-Ursheim (Nördlinger Ries, Bavaria, Southern Germany) // Palaeodiversity. 2017. V. 10. P. 49–95. https://doi.org/10.18476/pale.v10.a5 49
Franz M., Tesakova E., Beher E. Documentation and revision of the index ostracods from the Lower and Middle Jurassic in SW Germany according to BUCK (1954) // Palaeodiversity. 2009. V. 2. P. 119–167.
Franz M., Beher E., Dietl G. The Bathonian and Early Callovian Ostracoda of Baden-Wuerttemberg, southern Germany // N. Jb. Geol. Paläontol. Abh. 2014. V. 274. № 2/3. P. 149–185.
Franz M., Ebert M., Stulpinaite R. Aalenian – Lower Bajocian (Middle Jurassic) ostracods from the Geisingen clay pit (SW Germany) // Palaeodiversity. 2018. V. 11. P. 59–105.
Herngreen G.F.W., Lissenberg Th., de Boer K.F. et al. Middle Callovian beds in the Achterhoek, Eastern Netherlands // Med. Riks Geol. Dienst. 1983. V. 37. № 3. P. 1–29.
Herrig E. Ostracoden aus der Weisen Schreibkreide (Unter Maastricht) der Insel Rügen // Paläontol. Abh. A. 1966. Bd 2. № 4. S. 693–1069.
Huber B., Müller B., Luterbacher H. Mikropaläontologische Untersuchungen an der Callovien/Oxfordien-Grenze im Schweizer Jura und auf der Schwäbischen Alb (vorläufige Mitteilung) // Ecl. geol. Helv. 1987. V. 80. № 2. P. 449–459.
Kilenyi T. The Jurassic. Part III, Callovian–Portlandian // A stratigraphical index of British Ostracoda / Eds. Bate R., Robinson E. Liverpool, 1978. P. 259–298 (Geol. J. Spec. Iss. № 8).
Lutze G.F. Zur Stratigraphie und Palaontologie des Callovien und Oxfordien in Nordwest-Deutschland // Geol. Jg. 1960. Bd 77. P. 391–532.
Oertli H.J. Faunes d’Ostracodes du Mesozoique de France. Leiden: Brill, 1963. 57 p.
Olempska E., Błaszyk J. A boreal ostracod assemblage from the Callovian of the Lukow Area, Poland // Acta Palaeontol. Pol. 2001. V. 46. № 4. P. 553–582.
Schudack U., Schudack M.E. Ostracods from the Upper Jurassic (Oxfordian-Tithonian) of southern Germany // J. Micropalaeontol. 2000. V. 19. P. 97–112.
Sheppard L.M. Bathonian Ostracod Correlation North and South of the English Channel, with the description of two new Bathonian Ostracods // Microfossils of Recent and Fossil Shelf Seas / Eds. Neale J.W., Brasier M.D. Chichester: Ellis Horwood, 1981a. P. 71–83.
Sheppard L.M. Middle Jurassic Ostracoda from Southern England and Northern France. Thesis Ph.D. University College London. 1981b. 214 p. [unpubl.]
Tesakova E.M. Callovian and Oxfordian ostracodes from the central region of the Russian Plate // Paleontol. J. 2003. V. 37. Suppl. 2. P. 107–227.
Tesakova E. Late Callovian and Early Oxfordian ostracods from the Dubki section (Saratov area, Russia): implications for stratigraphy, paleoecology, eustatic cycles and palaeobiogeography // N. Jb. Geol. Paläontol. Abh. 2008. V. 249. № 1. P. 25–45.
Tesakova E.M. Ostracode-based reconstruction of paleodepths in the Early Callovian of the Kursk Region, Central Russia // Paleontol. J. 2013. V. 47. № 10. P. 1214–1229.
Tesakova E.M., Franz M., Baykina E., Beher E. A new view on Bathonian ostracods of Poland // Senckenb. Lethaea. 2008. V. 88. № 1. P. 55–65.
Whatley R.C. Scottish Callovian and Oxfordian Ostracoda // Bull. Brit. Mus. Natur. Hist. Geol. 1970. V. 19. № 6. P. 299–358.
Whatley R.C., Ballent S. A review of the Mesozoic ostracod genus Lophocythere and its close allies // Palaeontology. 2004. V. 47. P. 81–108.
Whatley R.C., Ballent S., Armitage J. Callovian Ostracoda from the Oxford Clay of southern England // Rev. Esp. Micropaleontol. 2001. V. 33. № 2. P. 135–162.
Wilkinson I.P., Whatley R.C. Upper Jurassic (Callovian–Portlandian) // Ostracods in British Stratigraphy / Eds. Whittaker J.E., Hart M.B. L., 2009. P. 241–287 (The Micropalaeontol. Soc.–Spec. Publ. Geol. Soc. London).
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал