

Журнал общей биологии, 2022, T. 83, № 5, стр. 346-357
Совместная динамика популяций брусники и черники в заповедных послепожарных сосняках: ретроспективный прогноз в рамках марковской модели
А. А. Маслов 1, *, Д. О. Логофет 1, 2
1 Институт лесоведения РАН
143030 Московская обл., п/о Успенское, Советская, 21, Россия
2 Институт физики атмосферы им. А.М. Обухова РАН
119017 Москва, Пыжевский пер., 3, Россия
* E-mail: amaslov@ilan.ras.ru
Поступила в редакцию 27.08.2022
После доработки 14.09.2022
Принята к публикации 19.09.2022
- EDN: DWTLQG
- DOI: 10.31857/S0044459622050062
Аннотация
Совместная динамика популяций брусники (Vaccinium vitis-idaea) и черники (V. myrtillus) после пожаров в сосняках двух типов: заболоченном сосняке-долгомошнике (Pinetum polytrichosum) и сухом сосняке-зеленомошнике (P. vacciniosum) – на бедных песчаных почвах в зоне смешанных лесов исследована с помощью математической модели. Полевые наблюдения провели в течение 25 лет (1980–2005 гг.) на постоянных трансектах, причем план эксперимента отличался большим размером выборки, 5-летним интервалом между учетами и регистрацией встречаемости видов на площадках 20 × 20 см. В качестве модели наблюдаемой динамики предложена дискретная марковская цепь из четырех состояний: отсутствие обоих видов на площадке, присутствие только брусники, присутствие только черники, совместное присутствие видов. Каждый 5-летний шаг выявляет переходы между любой парой состояний, и мы калибруем переходную матрицу цепи, усредняя частоты переходов за весь период наблюдений для каждого типа леса. Модель стандартным образом предсказывает стационарное распределение состояний площадок в будущем и впервые представляет ретроспективную картину того, как послепожарная сукцессия в ярусе кустарничков началась и продолжалась в прошлом. Показано, что в двух разных сообществах наблюдается одинаковый тренд изменений во времени: доля площадок только с V. vitis-idaea увеличивается на ранних стадиях послепожарной сукцессии, достигает локального максимума, а затем снижается. Наоборот, доли площадок только с V. myrtillus и площадок с обоими видами постоянно увеличиваются в ходе сукцессии. И в сосняке-долгомошнике, и в сосняке-зеленомошнике стационарное распределение состояний характеризуется максимальным процентом участков с двумя видами, преобладающими над участками только с черникой, которые, в свою очередь, преобладают над участками только с брусникой. Полученные результаты подтверждают возможность устойчивого сосуществования черники и брусники на заключительных стадиях сукцессии с заметным преобладанием черники. Различия между типами леса заключаются, главным образом, в скорости сукцессии: если в заболоченном сосняке-долгомошнике модельное время от начала послепожарной сукцессии в кустарничковом ярусе до выхода на устойчивое (финальное) распределение составляет 30–45 лет, то в сухом сосняке-зеленомошнике оно вдвое больше, 60–90 лет. Кроме того, брусника преобладает на ранних стадиях сукцессии в сосняке-зеленомошнике. Априори не известно, на какой именно стадии сукцессии был начат полевой эксперимент, а ретроспективный метод позволяет восстановить неочевидные ранние фазы сукцессии и привязать период сбора данных к определенному отрезку ее хода.
Настоящая публикация продолжает серию работ по изучению мелкомасштабной динамики популяций вересковых кустарничков – брусники (Vaccinium vitis-idaea) и черники (V. myrtillus) (Маслов, Логофет, 2016, 2020; Логофет, Маслов, 2018). В этих работах были построены марковские модели совместной динамики популяций брусники, черники в ходе послепожарной сукцессии в сосняках разными методами и, в частности, методом усреднения частот наблюдавшихся переходов между состояниями площадок (Logofet, Maslov, 2019; Maslov, Logofet, 2020). Построенные модели позволили рассчитать процент площадок с брусникой и черникой по отдельности, площадок с обоими видами и площадок, ими не занятых, а также процент площадок с брусникой, черникой в классическом понимании динамики популяций (Маслов, Логофет, 2020).
Эти результаты были получены в рамках однородных во времени марковских моделей, и их прогностическая ценность, с точки зрения биолога, заключается в том, что модель дает возможность рассчитать ход динамики или сукцессии вперед на многие годы/десятилетия по не слишком длинному ряду полевых наблюдений. Особую ценность при этом имеют характеристики равновесного (финального, климаксного) распределения состояний при гипотезе сохранения тех условий среды, которые реализовались за весь период наблюдений.
Вместе с тем любые графики хода сукцессии (или динамики популяций) как результаты расчета марковского процесса своей начальной точкой всегда имеют то состояние системы, которое зафиксировано в момент начала полевых наблюдений. От этой точки строится прогноз в будущее. Как следствие, если период наблюдений приходится на среднюю или позднюю фазу динамики, модельный прогноз не скажет нам ничего о характере более ранних стадий. Возникают два очевидных вопроса: 1) Можно ли сделать ретроспективный прогноз от точки начала фактических наблюдений, рассчитав модельное распределение состояний на более ранних фазах динамики? 2) Возможно ли определить в рамках модели ту стадию (фазу) динамики, на которой находится лес в период фактических наблюдений? Поиск ответов на эти вопросы, а также сравнение хода сукцессии в заболоченном и сухом типе леса составляет содержание настоящей работы.
С математической точки зрения вопрос о возможности прогноза вперед (из настоящего в будущее) или ретроспективного прогноза “из прошлого” в настоящее и будущее решается слишком просто, поскольку фундаментальное свойство однородной модели обеспечивает сходимость к единственному финальному распределению состояний из любого начального распределения – каким бы в прошлом оно ни было.
Поэтому ранее (Logofet, 2019) была предпринята попытка осуществить на основе наших полевых данных обратный прогноз, т.е. проекцию из настоящего назад в прошлое. Формально дело сводилось к обращению вспять оси времени, соответствующей инверсии данных наблюдений и осреднению полученных одношаговых переходных матриц марковской цепи (Logofet, 2019). Однако финальное распределение состояний, полученное в результате обратного прогноза и призванное установить начальное распределение в прошлом, лишь качественно подтвердило общее направление хода сукцессии в будущее при том, что количественные различия между прошлым и будущим распределениями оказались чрезвычайно малы (Logofet, 2019, Table 5). Такой результат не мог удовлетворить биолога, так как не соответствовал известным фактам об общем ходе послепожарных сукцессий.
Предлагаемый в настоящей работе ретроспективный метод основан на априорном предположении о распределении состояний в начальный момент сукцессии и неизменности переходной матрицы итоговой модели. Вполне логично предположить, что после пожара распределение состояний площадок исчерпывается 100% площадок, где оба вида отсутствуют. Подобный подход вполне оправдан не только для гарей, но и для любых сукцессий на месте обнаженной почвы или субстрата. Как оказалось, аналогичный прием использован ранее в работах по зарастанию обнаженной почвы в сосняке лишайниковом (Самойлов, Тархова, 1985) и шахтных отвалов (Baasch et al., 2010).
Положительный ответ на второй вопрос – возможно ли определить с помощью модели конкретную стадию (фазу) динамики, на время которой пришелся период фактических наблюдений, – поможет биологу в интерпретации наблюдаемой картины, понимании характеристик растительности до и после периода наблюдений в природе, а также добавит информации к принятию решения о продолжении или прекращении этих весьма трудоемких работ.
1. ОБЪЕКТЫ И МЕТОД НАБЛЮДЕНИЙ
1.1. Объекты
Сосновые леса на бедных песчаных почвах – широко распространенный тип сообществ зоны бореальных лесов (Hytteborn et al., 2005; Рысин, Савельева, 2008). В центре Русской равнины такие леса особенно часто встречаются на территории флювиогляциальных равнин (Огуреева и др., 1996). Полевые исследования проводили на севере Московской области – в заповедных лесных участках Клинского и Талдомского районов (Верхневолжская низменность, зона смешанных лесов). Постоянные пробные площади размером 50 × 40 м были заложены для изучения сукцессионной динамики в двух типах сосняков на бедной песчаной почве: “сухом” зеленомошнике и заболоченном долгомошнике. Время начала послепожарной сукцессии в сосняке-зеленомошнике – 1894 г. (год сплошного пожара). Время начала послепожарной сукцессии в сосняке-долгомошнике – 1954 г. (год низового пожара средней интенсивности). На момент начала наблюдений (1980 г.) возраст сосняка-зеленомошника составлял 70 лет, сосняка-долгомошника – 60 лет. И в том, и в другом сообществе абсолютными доминантами травяно-кустарничкового яруса были черника и брусника, однако суммарное покрытие двумя этими видами не превышало 30–40%. В мохово-лишайниковом ярусе в сосняке-зеленомошнике доминировали Pleurozium schreberi и Dicranum rugosum; отдельные пятна формировали лишайники Cladina arbuscula и C. rangiferina. В сосняке-долгомошнике преобладали Polytrichum commune, Pleurozium schreberi и Sphagnum girgensohnii. Подробное описание объектов приведено в недавних публикациях (Маслов, Логофет, 2016, 2020) и в монографии А.А. Маслова (1990).
1.2. Метод наблюдений, характер данных
Учеты видов нижних ярусов проводили на постоянных трансектах, состоящих из закрепленных на местности площадок размером 0.2 × 0.2 м; общее число площадок на пробной площади составляло 2000. На площадках отмечали факт присутствия укоренившихся побегов трав и кустарничков и присутствие видов мхов. В качестве счетных единиц у черники и брусники выступали парциальные побеги. Отдельно отмечали также наличие на поверхности почвы валежа – стволов и крупных ветвей. Подробнее полевая методика описана в первой публикации по теме (Маслов, Логофет, 2016) и в монографии (Маслов, 1990).
Продолжительность наблюдений на постоянных трансектах составила 25 лет: с 1980 по 2005 год с проведением перечетов каждые пять лет. Всего имеется шесть учетов, что позволяет отследить изменения на площадках за пять интервалов: 1980 → 1985, 1985 → 1990, 1990 → 1995, 1995 → 2000, 2000 → 2005. За весь период наблюдений общее число описаний площадок в лесу каждого типа составило 2000 × 6 = 12 000.
В ходе первичной обработки данных присутствие на площадке черники обозначали буквой V, а брусники – буквой W, и тогда состояние каждой площадки определялось в следующих терминах: 1) ∅ – оба вида отсутствуют (такие площадки для краткости называем далее “незанятыми”), 2) V – есть черника, 3) W – есть брусника, 4) VW – присутствуют оба вида. Это позволяет представить общую картину состояния объекта в год учета в виде частотного распределения всех учтенных площадок по четырем состояниям, а данные всех шести учетов – в виде таблицы из 4 строк и 6 столбцов (табл. 1).
Таблица 1.
Данные шести последовательных учетов в сосняке-долгомошнике и сосняке-зеленомошнике по годам в виде частотных распределений площадок по состояниями: ∅ – отсутствие видов, V – черника, W – брусника, VW – оба вида
| Состояния | 1980 | 1985 | 1990 | 1995 | 2000 | 2005 |
|---|---|---|---|---|---|---|
| Сосняк-долгомошник | ||||||
| ∅ | 401 | 350 | 505 | 482 | 321 | 280 |
| V | 580 | 451 | 516 | 524 | 646 | 601 |
| W | 273 | 278 | 246 | 245 | 157 | 175 |
| VW | 483 | 668 | 515 | 498 | 587 | 655 |
| Число площадок | 1737 | 1747 | 1782 | 1749 | 1711 | 1711 |
| Сосняк-зеленомошник | ||||||
| ∅ | 1188 | 917 | 660 | 530 | 401 | 344 |
| V | 133 | 138 | 208 | 280 | 297 | 440 |
| W | 359 | 454 | 511 | 508 | 515 | 469 |
| VW | 229 | 355 | 424 | 440 | 578 | 538 |
| Число площадок | 1909 | 1864 | 1803 | 1758 | 1791 | 1791 |
2. МАТЕМАТИЧЕСКАЯ МОДЕЛЬ И РЕТРОСПЕКТИВНЫЙ АНАЛИЗ
2.1. Марковская цепь как модель наблюдаемого процесса
Следуя способу представления данных (раздел 1.2), марковская цепь имеет четыре возможных состояния, обозначенных теми же буквами ∅, V, W и VW, а переменными состояния модели служат конечные распределения x(t) вероятностей состояний цепи в момент наблюдения t = 1980, 1985, … (Маслов, Логофет, 2016).
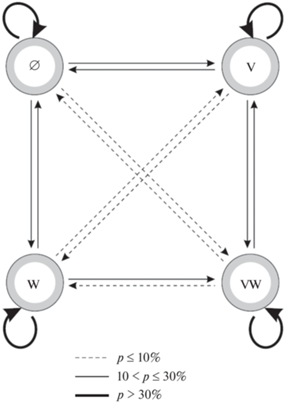
Постулат марковости означает, что распределение вероятностей переходов из любого состояния S (S = ∅, V, W, VW) во все остальные целиком определяется текущим состоянием и не зависит от того, каким путем цепь пришла в это состояние. Из априорных биологических представлений (подтвержденных нашими наблюдениями) следует, что за 5 лет в природе возможны все типы переходов между четырьмя перечисленными состояниями и, значит, схему переходов за один шаг модели можно изобразить в виде полного ориентированного графа (орграфа), в котором присутствуют дуги, ведущие из любого узла графа в любой другой, включая тот же самый. На рис. 1 показан полный орграф переходов в сосняке-долгомошнике, причем разными типами линий показан количественной уровень соответствующей частоты перехода из одного состояния в другое, но этот вопрос мы рассмотрим при обсуждении результатов. Аналогичный орграф для сосняка-зеленомошника опубликован ранее (Маслов, Логофет, 2020, рис. 1).
Рис. 1.
Полный граф переходов между четырьмя состояниями учетной площадки за один временной шаг Δt = 5 согласно данным учетов в сосняке-долгомошнике. Состояния: ∅ – оба вида отсутствуют, V– есть Vaccinium myrtillus, W – есть V. vitis-idaea, VW – присутствуют оба вида. Частоты переходов даны в процентах.
Формально, если распределение вероятностей перехода из некоторого состояния S (S = ∅, V, W, VW) в каждое из состояний представлено стохастическим вектором-столбцом [p∅S, pVS, pWS, pVW,S]T, а вероятности самих состояний в текущий момент времени t – стохастическим вектором-столбцом x(t) = [x∅, xV, xW, xVW]Т, то элементарная формула теории вероятностей дает вероятность состояния S в следующий момент, равную
(1)
$\begin{gathered} {{x}_{S}}(t + \Delta t) = {{x}_{\emptyset }}(t){{p}_{S}}_{\emptyset } + {{x}_{{\text{V}}}}(t){{p}_{S}}_{{\text{V}}} + \\ + \,\,{{x}_{{\text{W}}}}(t){{p}_{S}}_{{\text{W}}} + {{x}_{{{\text{VW}}}}}(t){{p}_{{S{\text{,VW}}}}}, \\ S = ~\,\,\emptyset ,{\text{V}},{\text{W}},{\text{VW}}. \\ \end{gathered} $Четыре скалярных уравнения (1) можно представить в векторно-матричной форме:
где P = [pij] – 4 × 4 матрица переходных вероятностей, или переходная матрица марковской цепи. Ее ненулевые элементы pij имеют смысл вероятностей перехода из состояния j в состояние i за один шаг Δt = 5 годам (i, j = 1, …, 4) и отражают относительную скорость процесса замещения видов на площадках. Из уравнения (2) следует, что если pij не изменяются во времени. Количественные значения pij определяются в результате калибровки переходной матрицы методом экспертной оценки или по данным наблюдений (см. ниже).Калиброванная переходная матрица позволяет вычислить такие характеристики модельного процесса, как стационарное распределение вероятностей для состояний цепи (x* = $[x_{1}^{*},x_{2}^{*},x_{3}^{*},x_{4}^{*}]$T), т.е. положительное и нормированное (условием $x_{1}^{*} + x_{2}^{*} + x_{3}^{*} + x_{4}^{*}$ = 1 либо 100%) решение уравнения
Общеизвестное следствие из классической теоремы Перрона–Фробениуса для неотрицательных матриц (Гантмахер, 1967) обеспечивает сходимость динамики x(t) к x* при достаточно больших значениях t из любого начального распределения x(0), т.е.
Для оценки длительности хода сукцессии было предложено (Маслов, Логофет, 2016) на каждом шаге по времени оценивать показатель несходства – меру Δ(x(t), x*) отличия текущего распределения x(t) от стационарного x* по Кейфицу (Keyfitz, 1968, p. 47),
где распределения выражены в процентах. Формальными критериями завершения сукцессии принимаем достижение показателя несходства на уровне ≤5% и (в более точном варианте) на уровне ≤1%.2.2. Метод калибровки
Характер имеющихся данных – наличие повторных учетов на постоянных трансектах в сочетании с большим размером выборки – предоставляет (довольно редкую в экологии) возможность вычислить все параметры модели однозначно и достоверно. Параметрами служат 16 переходных вероятностей pij из уравнения (2), точнее 12 переходных плюс 4 вероятности pjj сохранения текущего состояния за один шаг. О вероятностях мы судим по фактической картине переходов и в качестве их значений принимаем частоты состоявшихся событий перехода и сохранения состояния. Тогда четыре калибровочных уравнения (1) удовлетворялись бы автоматически для каждой из пяти пар последовательных учетов (табл. 1), если бы число учтенных площадок оставалась неизменным. Пять получаемых в результате такой калибровки переходных матриц закономерно отличались бы друг от друга, отражая временнóй тренд в изучаемом процессе и формируя тем самым неоднородную (во времени) цепь Маркова, или неавтономную модель в более общей терминологии. Однако в нашей ситуации не только число, но и сами учитываемые площадки меняются от учета к учету, и потому задача получить долгосрочный прогноз ставит вопрос об осреднении частот переходов в пяти интервалах между учетами. Временнóй тренд будет опосредовано учтен в значениях усредненных частот.
2.3. Метод осреднения
Наблюдая результаты процессов замещения или совмещения видов на площадках, мы не вникаем в причинные механизмы явления, а полагаем этот процесс случайным согласно известному тезису, что случайность есть непознанная закономерность. Тогда частота перехода из одного состояния в другое, зафиксированная в текущем учете, есть результат фактической реализации случайного процесса. Для каждого типа переходов частоты меняются по годам наблюдений. Логично предположить, что осреднение частот переходов, полученных в разных выборках (интервалах учетов) за весь период наблюдений позволит более точно оценить истинную вероятность перехода из одного состояния в другое (Маслов, Логофет, 2020).
Осреднение частот для каждого перехода из состояния j в состояние i проводили по стандартной формуле (Справочник…, 1989, с. 378):
(6)
$\overline {{{p}_{{ij}}}} = \frac{{\sum\limits_{l = 1}^k {{{n}_{l}}(j \to i)} }}{{\sum\limits_{l = 1}^k {{{N}_{l}}} }},$Все виды расчетов (как на этапе подготовки данных, так и в процессе моделирования) выполнены путем автоматизации вычислений в программе MS Excel (версия 2207), включая операции матричной алгебры.
2.4. Ретроспективный прогноз и временнáя локализация периода наблюдений
Ретроспективный прогноз основан на априорном предположении (или знании) о том, каким является распределение состояний в год начала сукцессии в кустарничковом ярусе – t0. В настоящей работе мы принимаем началом послепожарной сукцессии распределение состояний, где все 100% площадок относятся к состоянию ∅, т.е. “незанятых”.
Если начало отсчета времени поместить в год t0 начала послепожарной сукцессии в кустарничковом ярусе, t0 = 0, то год t1 первого учета окажется неопределенным: t1 = ?, а ретроспективный прогноз (из прошлого в будущее) будет означать вычисление модельных траекторий x(t) вплоть до достаточно малой окрестности финального состояния x* (см. (5)). Тогда вопрос, как “определить ту стадию (фазу) динамики, на которой находится лес в период фактических наблюдений”, формально сводится к задаче найти значение t1. Технически эта задача означает привязку временнóго периода наблюдений к модельному графику хода сукцессии. Поскольку распределение состояний в природе в конкретный год наблюдений вносит свой вклад случайности в итоговую переходную матрицу P, для более точной привязки мы предлагаем использовать полную картину распределений, включающую все шесть учетов.
Эта общая картина, т.е. матрица 6 × 4 из табл. 1, сравнивается с последовательными шестерками модельных распределений, т.е. с матрицами
Полученный для определенного набора из шести шагов модели наименьший квадрат отклонений от фактической матрицы позволяет определить, на какую фазу сукцессии приходится ряд полевых наблюдений.
3. РЕЗУЛЬТАТЫ
3.1. Фактическая картина переходов
Данные о фактических переходах между состояниями площадок за пять интервалов между учетами в сосняке-долгомошнике представлены в табл. 2 в абсолютных числах. Чтобы убрать влияние фактора пространственного исключения и фактора нарушения, в каждом интервале учетов из анализа исключались площадки с живыми и сухостойными стволами деревьев и площадки с наличием валежа. В результате размеры выборки в разных интервалах учетов варьировали от 1711 до 1782.
Таблица 2.
Данные о переходах между состояниями площадок по интервалам учетов t → t + 5 в сосняке-долгомошнике за 25 лет наблюдений
| Год t | Переходы в | Исходящие состояния в год t | Итого в год t + 5 | |||
|---|---|---|---|---|---|---|
| ∅ | V | W | VW | |||
| 1980 | ∅ | 250 | 83 | 19 | 5 | 357 |
| V | 51 | 351 | 14 | 48 | 464 | |
| W | 82 | 16 | 157 | 24 | 279 | |
| VW | 18 | 130 | 83 | 406 | 637 | |
| Итого в год t | 401 | 580 | 273 | 483 | 1737 | |
| 1985 | ∅ | 252 | 89 | 100 | 58 | 499 |
| V | 52 | 281 | 31 | 143 | 507 | |
| W | 34 | 14 | 94 | 93 | 235 | |
| VW | 12 | 67 | 53 | 374 | 506 | |
| Итого в год t | 350 | 451 | 278 | 668 | 1747 | |
| 1990 | ∅ | 356 | 78 | 46 | 20 | 500 |
| V | 79 | 324 | 18 | 107 | 528 | |
| W | 52 | 16 | 125 | 51 | 244 | |
| VW | 18 | 98 | 57 | 337 | 510 | |
| Итого в год t | 505 | 516 | 246 | 515 | 1782 | |
| 1995 | ∅ | 256 | 34 | 31 | 0 | 321 |
| V | 151 | 368 | 24 | 111 | 654 | |
| W | 48 | 6 | 90 | 18 | 162 | |
| VW | 27 | 116 | 100 | 369 | 612 | |
| Итого в год t | 482 | 524 | 245 | 498 | 1749 | |
| 2000 | ∅ | 194 | 70 | 10 | 6 | 280 |
| V | 71 | 430 | 11 | 89 | 601 | |
| W | 40 | 17 | 87 | 31 | 175 | |
| VW | 16 | 129 | 49 | 461 | 655 | |
| Итого в год t | 321 | 646 | 157 | 587 | 1711 | |
Из 80 ячеек таблицы только одна, отвечающая за переход VW → ∅ в интервале 1995 → 2000, оказалась нулевой, что подтверждает предположение о реализации всех теоретически возможных переходов между состояниями и сохранении текущих состояний за пятилетние интервалы между учетами. Одновременно это свидетельствует о правильном выборе длины временнóго шага и количества пробных площадок: за каждые 5 лет в сообществе реализовались практически все переходы, что подтверждает полноту орграфа переходов (рис. 1). Аналогичные данные о фактических переходах между состояниями площадок за пять интервалов учетов в сосняке-зеленомошнике опубликованы в предыдущей статье (Маслов, Логофет, 2020).
3.2. Усредненные частоты переходов
Переходная матрица P для сосняка-долгомошника, вычисленная по данным табл. 2, представлена в табл. 3. Ее элементы (переходные вероятности), найденные как средневзвешенные частоты соответствующих событий по формуле (6), показаны для наглядности в форме процентов. На рис. 1 градации этих же вероятностей показаны разными типами линий. Наибольшие значения (>60%) за пятилетний интервал характерны только для диагональных элементов матрицы – вероятностей сохранения каждого из четырех состояний. Это говорит об относительно медленном ходе сукцессии. Наиболее вероятными переходами в сосняке-долгомошнике являются W → → VW, V → VW и ∅ → V. Аналогичная матрица усредненных вероятностей переходов для сосняка-зеленомошника опубликована в предыдущей статье (Маслов, Логофет, 2020).
Таблица 3.
Матрица усредненных вероятностей переходов (за пять интервалов) в сосняке-долгомошнике и ее предельный вектор x*
| Куда переход | Исходящие состояния | Предельный вектор x*, % | |||
|---|---|---|---|---|---|
| ∅ | V | W | VW | ||
| ∅ | 0.6353 | 0.1303 | 0.1718 | 0.0324 | 20.1 |
| V | 0.1962 | 0.6456 | 0.0817 | 0.1810 | 32.2 |
| W | 0.1243 | 0.0254 | 0.4612 | 0.0789 | 11.5 |
| VW | 0.0442 | 0.1987 | 0.2853 | 0.7077 | 36.2 |
3.3. Стационарное распределение вероятностей состояний и графики сходимости
В последнем столбце табл. 3 для сосняка-долгомошника показан предельный (равновесный) вектор вероятностей x*, полученный как решение уравнения (4) с матрицей P из табл. 3 и выраженный в процентах. Иными словами, предельным результатом сукцессии – при гипотезе однородности во времени, т.е. сохранении тех условий среды, которые были реализованы за весь период наблюдений, – должно быть распределение состояний, где 11.5% площадок будет занято только брусникой, 32.2% – только черникой, на 36.2% площадок будут присутствовать оба вида, а 20.1% площадок будут существовать без кустарничков.
В табл. 4 для того же сообщества представлены показатели несходства между распределениями в конкретные годы после начала сукцессии и предельным распределением. Видно, что компоненты начального распределения достаточно быстро приближаются к предельным значениям (табл. 4; рис. 2а). Так, через 5 лет после начала послепожарной сукцессии распределение состояний отличается от стационарного на 44.4%, а через 20 лет сукцессии – на 11%. Отличие ≤5% достигается через 30 лет (4.3%), а через 45 лет оно составляет 1%. Важно отметить, что показатель несходства уменьшается на каждом шаге моделирования, что свидетельствует о направленности процесса в целом и в пользу состоятельности калиброванной матрицы из средневзвешенных частот.
Таблица 4.
Показатели несходства между равновесием x* и распределениями x(t) в конкретные годы после начала сукцессии в сосняке-долгомошнике
| Время с начала сукцессии, лет | 0 | 5 | 10 | 15 | 20 | 25 | 30 | 35 | 40 | 45 |
|---|---|---|---|---|---|---|---|---|---|---|
| Показатель, % | 79.9 | 44.4 | 28.1 | 17.6 | 11.0 | 6.8 | 4.3 | 2.7 | 1.7 | 1.0 |
Рис. 2.
Динамика распределения состояний площадок по пятилетним шагам в усредненной модели от условного начала сукцессии в кустарничковом ярусе до достижения равновесного состояния: а – сосняк-долгомошник, б – сосняк-зеленомошник. Серой заливкой показан расчетный период полевых наблюдений, вертикальными линиями – моменты времени, когда распределение состояний отличается от стационарного: ≤5% (штриховая линия) и ≤1% (штрих-пунктирная линия). Другие обозначения см. на рис. 1.
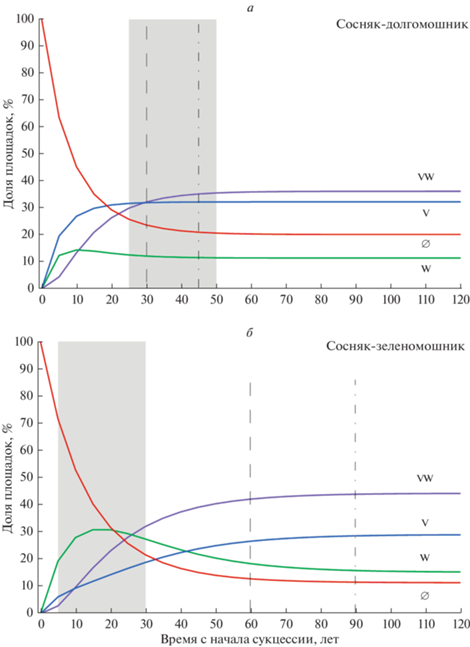
Равновесный предельный вектор вероятностей x* для сосняка-зеленомошника опубликован ранее (Маслов, Логофет, 2020). В процентах его значения составляют 11.3, 29.1, 15.2, 44.4%, что можно видеть из графика на рис. 2б. Как показал расчет показателей несходства между распределениями состояний в конкретные годы после начала сукцессии в кустарничковом ярусе и равновесием x*, отличие ≤5% достигается через 60 лет, а ≤1% – через 90 лет (рис. 2б).
Как следует из рис. 2, и в том, и в другом сообществе в ходе сукцессии возрастает доля площадок только с черникой (V) и доля площадок, где оба вида произрастают совместно (WV). Напротив, доля “незанятых” площадок (∅) быстро снижается. В отличие от монотонного возрастания/убывания для V, WV, ∅, число площадок только с брусникой (W) сначала растет, а затем снижается – кривая имеет локальный максимум.
3.4. Временнáя локализация периода наблюдений
Результаты сравнения матрицы фактических распределений состояний за шесть учетов с последовательными шестерками модельных распределений приведены в табл. 5.
Таблица 5.
Отклонения фактической картины шести распределений от последовательных наборов из шести модельных прогнозов в сосняке-долгомошнике в виде “наименьших квадратов” (жирным шрифтом выделено наименьшее значение)
| Время t1 с начала сукцессии, лет | 0 | 5 | 10 | 15 | 20 | 25 | 30 | 35 |
|---|---|---|---|---|---|---|---|---|
| Интервал сукцессии, лет | 0–25 | 5–30 | 10–35 | 15–40 | 20–45 | 25–50 | 30–55 | 35–60 |
| Сумма квадратов отклонений | 1.2192 | 0.3978 | 0.1449 | 0.0627 | 0.0395 | 0.0360 | 0.0383 | 0.0414 |
Наибольший квадрат отклонений фактической картины от модельного прогноза наблюдается в сосняке-долгомошнике в интервале сукцессии 0–25 лет, т.е. в самом начале. С увеличением времени от начала сукцессии сумма квадратов отклонений между фактической матрицей и модельной уменьшается, достигая минимума для времени t1 = 25 лет. При дальнейшем увеличении времени различия вновь возрастают. Таким образом, в сосняке-долгомошнике период фактических наблюдений наилучшим образом соответствует модельному распределению состояний в интервале времени 25–50 лет от начала сукцессии в кустарничковом ярусе. Для большей наглядности этот расчетный период полевых наблюдений показан серой заливкой на графиках хода сукцессии (рис. 2).
Аналогичным образом временнýю привязку периода наблюдений осуществили и в сосняке-зеленомошнике (рис. 2б).
3.5. Динамика популяций
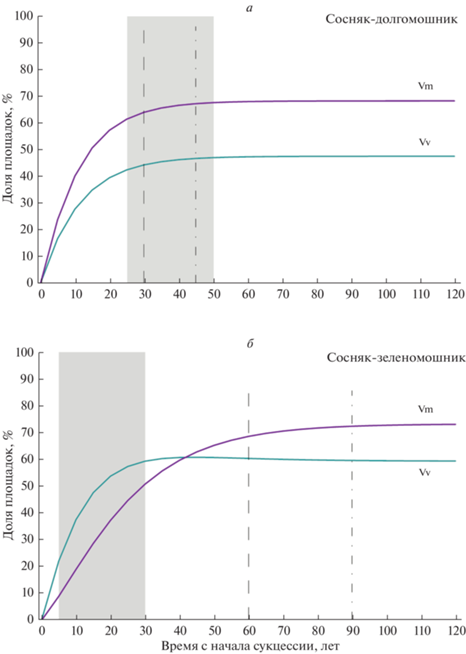
Расчет по шагам сходимости начального распределения состояний x(0) к равновесному состоянию x* (рис. 2) позволяет рассчитать и динамику популяций каждого вида отдельно в традиционной для популяционной теории форме. Действительно, если Vm обозначает число всех площадок с черникой, а Vv – число всех площадок с брусникой, то на каждом шаге Vm = V + WV, а Vv = W + WV.
На рис. 3 представлена динамика популяций брусники, черники по шагам усредненной модели – от условного начала сукцессии до достижения равновесного состояния. Как оказалось, и в сосняке-долгомошнике, и в сосняке-зеленомошнике в равновесном состоянии преобладают площадки с черникой – в долгомошнике они составляют 68%, в зеленомошнике – 73%. Между типами леса есть и различия. В заболоченном сосняке-долгомошнике роль черники увеличивается быстрее и в результате ее обилие всегда выше, чем у брусники (рис. 3а). В сухом сосняке-зеленомошнике на ранних стадиях быстрее растет встречаемость брусники, и в результате на протяжении первых 40 лет обилие брусники выше, чем у черники. На более поздних стадиях виды меняются местами (рис. 3б), в связи с чем были описаны несколько стадий сукцессии (Маслов, Логофет, 2020).
Рис. 3.
Динамика популяций Vaccinium myrtillus (Vm) и V. vitis-idaea (Vv) по шагам в усредненной модели от начала сукцессии в кустарничковом ярусе до достижения равновесного состояния: а – сосняк-долгомошник, б – сосняк-зеленомошник. Vm = V + VW, Vv = W + VW. Другие обозначения см. на рис. 2.
4. ОБСУЖДЕНИЕ
Результаты проведенной работы позволяют дать положительные ответы на оба поставленных вопроса: 1) ретроспективный прогноз динамики (или хода сукцессии) – из прошлого через период наблюдений в природе в будущее – возможен с помощью однородной марковской модели; 2) с помощью модели можно определить ту стадию (фазу) сукцессии, на время которой пришелся период фактических наблюдений.
4.1. Важность ретроспективы
Главным условием для прогноза “в прошлое” по данным марковской модели является биологически обоснованное определение начального состояния системы, т.е. распределения состояний в момент начала динамики. Такое распределение достаточно очевидно, когда речь идет о первичных сукцессиях на лишенных растительности участках (Денисенко и др., 1996; Baasch et al., 2010) или о вторичных сукцессиях на залежах (Korotkov et al., 2001) и сплошных гарях. Распределение состояний в этих случаях сводится к 100% площадок, свободных от изучаемых видов растений. Для других типов сукцессии, если распределение состояний в начальный момент не известно, применение ретроспективного анализа в рамках однородной марковской модели оказывается невозможным.
Ретроспективный прогноз динамики позволяет решить проблему оценки общей продолжительности сукцессии (или процесса динамики) – от начальной фазы до достижения равновесного состояния. Классический расчет модели – вперед от времени наблюдений в природе – не может дать такой оценки, так как зависит от фазы динамики. Наблюдения, проведенные в ранней фазе, дадут существенно бóльшую продолжительность сукцессии, чем наблюдения, проведенные в поздней фазе. В том случае, когда время начала сукцессии (например, год пожара) невозможно определить независимым способом, ретроспективный расчет, объединенный с расчетом “вперед”, дает реальный шанс оценить продолжительность всех фаз динамики и общее время сукцессии.
В сосняке-долгомошнике, как показал ретроспективный анализ, время от условного начала сукцессии до начала фактических наблюдений составило 25 лет, а общее время сукцессии от заданного “нуля” (все без кустарничков) до равновесного – 30–45 лет (рис. 2а). Как показала привязка периода наблюдений, учеты в природе пришлись на позднюю фазу сукцессии – последние годы учетов настолько близки к стационарному состоянию, что дальнейшее продолжение трудоемких полевых наблюдений вряд ли имело смысл. В этих условиях ретроспективный прогноз позволил выявить в сосняке-долгомошнике ранние фазы динамики, предсказать которые иными способами было бы невозможно. В частности, оказалось, что график для состояния “только брусника” имеет локальный максимум через 10 лет после начала сукцессии.
Иную картину мы наблюдаем в сосняке-зеленомошнике. Время от условного начала сукцессии (все без кустарничков) до начала фактических наблюдений составило всего 5 лет (рис. 2б), а общее время сукцессии до равновесного состояния – 60–90 лет. Учеты в природе пришлись на начальную фазу динамики популяций кустарничков и периоды существенных изменений в распределении состояний. Согласно модели, в случае продолжения полевых наблюдений еще 30 лет в распределении состояний наблюдались бы важные изменения.
4.2. Оценка общего времени послепожарной сукцессии
При анализе хода сукцессий прогностическая ценность марковских моделей определяется, в первую очередь, верной концептуальной схемой и правильным составлением переходной матрицы (Логофет, 2010). Очевидно, что всеобъемлющая модель сукцессии должна включать все состояния (фазы динамики) – от пионерных до завершающих, при этом разные представления авторов о начальных и конечных фазах сукцессии будут определять и результаты моделирования.
В настоящей работе за перечень состояний мы принимали наличие или отсутствие на учетных площадках побегов брусники и черники (рис. 1) в ходе послепожарной сукцессии. Это заведомо исключает из схемы первые стадии после пожара в незаболоченных сосняках. Согласно литературным данным, на ранних стадиях послепожарной сукцессии в сухих сосняках преобладают лишайники, затем им на смену приходят зеленые мхи (Pleurozium schreberi, виды р. Dicranum), и лишь на более поздних стадиях преобладают кустарнички (Горшков, 1995; Самойлов, Ипатов, 1995; Коротков, 2002; Маслов, 2002; Hautala et al., 2005).
В заболоченном сосняке-долгомошнике расчетное время от начала сукцессии кустарничкового яруса до начала фактических наблюдений составило 25 лет (рис. 2а). Наблюдения были начаты в 1980 г., а низовой пожар (по данным анализа пожарных шрамов на соснах) произошел в 1954 г. (Маслов, 1990). Началом послепожарной сукцессии можно считать 1955 г. Таким образом, интервал от фактического начала сукцессии до начала полевых наблюдений составил точно такие же 25 лет. Совпадение временны́х интервалов сукцессии (согласно модели и согласно фактическим данным) позволяет сделать важный вывод о том, что кустарнички появляются в заболоченном сосняке-долгомошнике сразу после низового пожара. Похожий эффект очень быстрого роста черники сразу после пожара описан для близкого типа леса в Литве (Marozas et al., 2011).
В сухом сосняке-зеленомошнике расчетное время от условного начала сукцессии кустарничкового яруса до начала фактических наблюдений составило только 5 лет (рис. 2б). Наблюдения были начаты в 1980 г., сплошной пожар произошел в 1894 г. Началом послепожарной сукцессии можно считать 1895 г. Таким образом, интервал от начала сукцессии до начала полевых наблюдений составил 85 лет. Существенное несовпадение временны́х интервалов сукцессии (согласно модели и согласно фактическим данным) позволяет сделать вывод о том, что в сухом боровом сосняке интервал от фактического начала послепожарной сукцессии до появления брусники, черники (за счет лишайниковых, зеленомошных стадий) составил восемь десятилетий. До достижения равновесного состояния, согласно модели, необходимо еще 60–90 лет. Таким образом, общее время послепожарной сукцессии составляет 140–170 лет (Маслов, Логофет, 2020), но это время рассчитано с привлечением данных, выходящих за рамки модели динамики популяций кустарничков. В лесах Севера, по данным В.В. Горшкова с соавт. (2009), доля черники достоверно увеличивается через 80–100 лет после пожара, тогда как на южной границе распространения изученных нами типов леса (Брянская обл.) кустарнички доминируют уже через 25 лет после пожара (Evstigneev, Korotkov, 2016).
4.3. Сравнение хода сукцессии в сосняках двух типов
Результаты моделирования позволяют подробнее описать этапы послепожарной сукцессии на разных уровнях детализации: а) динамика состояний площадок, отражающих раздельную и совместную встречаемость черники, брусники; б) динамика популяций черники и брусники (Маслов, Логофет, 2020).
По результатам динамики состояний площадок (рис. 2) можно выделить несколько стадий динамики. В заболоченном сосняке-долгомошнике на первой стадии преобладают площадки только с черникой, на втором месте – площадки только с брусникой, на последнем месте – площадки, где оба вида присутствуют вместе. Средняя стадия характеризуется быстрым увеличением роли площадок только с черникой и несколько медленнее – площадок с обоими видами. На заключительной стадии преобладают площадки с двумя видами и только с черникой. В сухом сосняке-зеленомошнике отличие первой стадии – преобладание площадок только с брусникой, тогда как тенденции средней и заключительных фаз такие же, как в долгомошнике.
Таким образом, результаты моделирования подтверждают тот факт, что между черникой и брусникой в ходе послепожарной сукцессии не происходит конкурентного исключения, хотя экологические ниши видов очень близки (Grime et al., 1988; Makipaa, 1999). Напротив, площадки, где оба вида произрастают совместно, преобладают на поздних стадиях сукцессии (рис. 2). На уровне популяций отдельных видов стационарные состояния (поздние фазы сукцессии) в заболоченном долгомошнике и сухом зеленомошнике практически совпадают: преобладает черника, однако брусники лишь ненамного меньше (рис. 3).
Время послепожарной сукцессии кустарничкового яруса в изученных типах леса существенно различается. В сухом сосняке-зеленомошнике оно составляет 60–90 лет, а в заболоченном долгомошнике только 30–45. Если же выйти за рамки двувидовой модели (с учетом начальных стадий без кустарничков), то общее время сукцессии составит в зеленомошнике 140–170 лет против 30–45 в долгомошнике. Различия в скорости изменений, по всей вероятности, связаны с более медленным накоплением гумуса в сухих условиях зеленомошника по сравнению с заболоченным долгомошником (Маслов, Логофет, 2020).
5. ЗАКЛЮЧЕНИЕ
Исследование совместной динамики популяций брусники (Vaccinium vitis-idaea) и черники (V. myrtillus) с помощью ретроспективного прогноза в рамках марковской модели позволило описать процесс послепожарной сукцессии в двух разных типах сосновых лесов.
Для заболоченного сосняка-долгомошника (Pinetum polytrichosum) и сухого сосняка-зеленомошника (P. vacciniosum) реализована однородная марковская модель с усредненными вероятностями перехода (Маслов, Логофет, 2020), причем модельные расчеты позволили не только получить прогноз стационарного состояния системы в будущем, но и выполнить ретроспективный анализ хода послепожарной сукцессии в прошлом и оценить длительность сукцессии на основании фактических и модельных данных. Ретроспективный метод основан на априорном предположении о распределении состояний в начальный момент сукцессии и неизменности матрицы переходов. Одновременно предложен оригинальный метод локализации периода сбора данных (полевых наблюдений) относительно хода сукцессии. Показано, что в двух разных сообществах наблюдается одинаковый тренд изменений во времени: на ранних этапах послепожарной сукцессии возрастает и достигает локального максимума доля площадок только с брусникой, в дальнейшем доля таких площадок снижается. Напротив, доли площадок только с черникой и площадок с присутствием двух видов постоянно увеличиваются в ходе сукцессии. И в сосняке-долгомошнике, и в сосняке-зеленомошнике стационарное распределение состояний характеризуется максимальным процентом площадок с присутствием обоих видов, на втором месте – площадки только с черникой, на третьем – только с брусникой. Результаты подтверждают возможность устойчивого сосуществования черники и брусники на заключительных стадиях сукцессии с заметным преобладанием черники. Установленные различия между типами леса заключаются, главным образом, в скоростях сукцессии. В заболоченном сосняке-долгомошнике модельное время от начала послепожарной сукцессии в ярусе кустарничков до достижения устойчивого (финального) состояния составляет 30–45 лет, а в сухом сосняке-зеленомошнике – вдвое больше (60–90). Кроме того, брусника преобладает на ранних стадиях сукцессии в сосняке-зеленомошнике. Локализация периода наблюдений в природе во времени хода сукцессии позволила восстановить неочевидные ранние фазы сукцессии, когда фактический ряд наблюдений в поле пришелся на средние или поздние стадии.
Список литературы
Гантмахер Ф.Р., 1967. Теория матриц. М.: Наука. 576 с.
Горшков В.В., 1995. Послепожарное восстановление мохово-лишайникового яруса в сосновых лесах Кольского полуострова // Экология. № 3. С. 179–183.
Горшков В.В., Ставрова Н.И., Баккал И.Ю., 2009. Основные этапы восстановительной динамики северотаежных лесов // Динамика лесных сообществ Северо-Запада России. СПб.: ВВМ. С. 228–236.
Денисенко Е.А., Каргополова У.Д., Логофет Д.О., 1996. Первичная сукцессия растительности в техногенном ландшафте лесостепной зоны (марковская модель) // Изв. РАН. Сер. Биол. № 5. С. 542–551.
Коротков В.Н., 2002. Биоразнообразие растительного покрова при разных типах пожарных нарушений: результаты наблюдений на стационарных пробных площадях в Окском заповеднике // Мониторинг сообществ на гарях и управление пожарами в заповедниках. М.: ВНИИ природы. С. 47–56.
Логофет Д.О., 2010. Марковские цепи как модели сукцессии: новые перспективы классической парадигмы // Лесоведение. № 2. С. 46–59.
Логофет Д.О., Маслов А.А., 2018. Анализ мелкомасштабной динамики двух видов-доминантов в сосняке чернично-бруснично-долгомошном. II. Неоднородная марковская цепь и осредненные показатели // Журн. общ. биологии. Т. 79. № 2. С. 135–147.
Маслов А.А., 1990. Количественный анализ горизонтальной структуры лесных сообществ. М.: Наука. 160 с.
Маслов А.А., 2002. Сукцессионная динамика древостоя и нижних ярусов в послепожарном 100-летнем сосняке лишайниково-зеленомошном // Лесоведение. № 2. С. 23–29.
Маслов А.А., Логофет Д.О., 2016. Анализ мелкомасштабной динамики двух видов-доминантов в сосняке чернично-бруснично-долгомошном. I. Однородная марковская цепь и показатели цикличности // Журн. общ. биологии. Т. 77. № 6. С. 423–433.
Маслов А.А., Логофет Д.О., 2020. Совместная динамика популяций черники и брусники в заповедном послепожарном сосняке-зеленомошнике. Модель с осредненными вероятностями перехода // Журн. общ. биологии. Т. 81. № 4. С. 243–256.
Огуреева Г.Н., Микляева И.М., Суслова Е.Г., Швергунова Л.В., 1996. Растительность Московской области. Пояснительный текст к карте (м-б 1:200 000). М.: Экор. 45 с.
Рысин Л.П., Савельева Л.И., 2008. Сосновые леса России. М.: Т-во науч. изд. КМК. 290 с.
Самойлов Ю.И., Ипатов В.С., 1995. Пирогенные сукцессии напочвенного покрова сухих сосняков на песках // Вестн. СПбГУ. Сер. 3. Биол. № 4. С. 58–67.
Самойлов Ю.И., Тархова Т.Н., 1985. Анализ сукцессионной мозаики напочвенного покрова с использованием марковских моделей // Бот. журн. Т. 70. № 1. C. 12–21.
Справочник по прикладной статистике, 1989 / Под ред. Ллойда Э., Ледермана У. Т. 1. М.: Финансы и статистика. 508 с.
Baasch A., Tischew S., Bruelheide H., 2010. Twelve years of succession on sandy substrates in a post-mining landscape: A Markov chain analysis // Ecol. Appl. V. 20. № 4. P. 1136–1147.
Evstigneev O.I., Korotkov V.N., 2016. Pine forest succession on sandy ridges within outwash plain (Sandur) in Nerussa-Desna Polesie // Russ. J. Ecosyst. Ecol. V. 1. № 3. https://doi.org/10.21685/2500-0578-2016-3-2
Grime J.P., Hodgson J.G., Hunt R., 1988. Comparative Plant Ecology: A Functional Approach to Common British Species. L.: Unwin Hyman. 742 p.
Hautala H., Kuuluvainen T., Hokkanen T.J., Tolvanen A., 2005. Long-term spatial organization of understorey vegetation in boreal Pinus sylvestris stands with different fire histories // Community Ecol. V. 6. № 2. P. 119–130.
Hytteborn H., Maslov A.A., Nazimova D.I., Rysin L.P., 2005. Boreal forests of Eurasia // Coniferous Forests (Ecosystems of the World, V. 6). Amsterdam: Elsevier. P. 23–99.
Keyfitz N., 1968. Introduction to the Mathematics of Population. Reading: Addison-Wesley. 490 p.
Korotkov V.N., Logofet D.O., Loreau M., 2001. Succession in mixed boreal forest of Russia: Markov models and non-Markov effects // Ecol. Model. V. 142. № 1–2. P. 25–38.
Logofet D.O., 2019. Bilberry vs. cowberry in a Scots pine boreal forest. II. Alternate modes of prediction // Ecol. Model. V. 405. P. 43–50.
Logofet D.O., Maslov A.A., 2019. Bilberry vs. cowberry in a Scots pine boreal forest: Exclusion or coexistence in a post-fire succession? // Ecol. Model. V. 401. P. 134–143.
Makipaa R., 1999. Response patterns of Vaccinium myrtillus and V. vitis-idaea along nutrient gradients in boreal forest // J. Veg. Sci. V. 10. № 1. P. 17–26.
Marozas V., Plausinyte E., Augustaitis A., Kaciulyte A., 2011. Changes of ground vegetation and tree-ring growth after surface fires in Scots pine forests // Acta Biol. Univ. Daugavp. V. 11. P. 156–162.
Maslov A.A., Logofet D.O., 2020. Bilberry vs. cowberry in a Scots pine boreal forest: III. Another forest, another method, and similar conclusions // Ecol. Model. V. 431. Art. 109193. https://doi.org/10.1016/j.ecolmodel.2020.109193
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии