

Журнал общей биологии, 2022, T. 83, № 3, стр. 170-182
Экспресс-перемещение частиц в столоне колониального гидроида Dynamena pumila (L., 1758)
В. С. Дементьев 1, Н. Н. Марфенин 1, *
1 Московский государственный университет им. М.В. Ломоносова, биологический факультет,
кафедра зоологии беспозвоночных
119234 Москва, Ленинские горы, 1, Россия
* E-mail: marf47@mail.ru
Поступила в редакцию 04.02.2022
После доработки 03.03.2022
Принята к публикации 12.03.2022
- EDN: DGDPHR
- DOI: 10.31857/S0044459622030046
Аннотация
Исследование выполнено по программе изучения механизмов самоуправления нецентрализованных биологических систем. Модельный объект – колониальный гидроид Dynamena pumila (L., 1758) – представитель модульных децентрализованных организмов. Распределительная система пульсаторно-реверсивного типа основана на пульсациях множества гидрантов и участков общего тела колонии – ценосарка. Их взаимодействие лежит в основе формирования время от времени протяженных течений жидкости (гидроплазмы), заполняющей внутреннюю трубковидную полость колониального организма. Особенность данной распределительной системы в том, что отдельных пульсаций недостаточно для перемещения частиц на значительное расстояние. Взаимодействие пульсаторов интерпретируется в научной литературе по-разному и поэтому нуждается в углубленном изучении. Это ключевой пункт любой концепции саморегуляции нецентрализованной системы. На колониальных гидроидах задача сводится к сравнению перемещения частиц в прямолинейном столоне двумя методами: макро- и микрорегистрацией. При макрорегистрации определяется время переноса частиц пищи к верхушке столона на максимально возможное расстояние от удаленного побега с гидрантами без учета отдельных пульсаций. При микрорегистрации учитываются пульсации с помощью цейтраферной микровидеосъемки, регистрирующей сжатия тела и локальное перемещение гидроплазмы. Нами впервые установлены протяженность и продолжительность перемещения не всего потока, а отдельных частиц в полости столона колонии D. pumila. Большинство частиц перемещались на расстояние одного-двух модулей столона, т.е. в среднем на 6–7 мм, что недостаточно для транспорта через всю колонию. Однако 18% из общего числа зарегистрированных частиц проходили безостановочно за 1–2 мин расстояние 23 мм – дистанцию от материнского побега, от которого выросла колония, до верхушки роста столона. Этот факт подтверждает гипотезу нецентрализованного согласования, в результате которого побеги, поочередно принимая в себя часть потока и сжимаясь после этого, принимают участие в однонаправленном перемещении гидроплазмы. Однако согласование пульсаторов неполное и непостоянное, что характерно для нецентрализованной саморегуляции. Всего прослежены траектории 372 частиц. Установленное соотношение траекторий по протяженности позволяет определить меры эффективности и хаотичности функционирования нецентрализованной распределительной системы пульсаторно-реверсивного типа.
Статья посвящена изучению механизмов функционирования нецентрализованных систем, в которых нет органов управления, а все части одинаковы по воздействию друг на друга. К таким системам относятся популяции, биоценозы, все модульные организмы и ткани многоклеточных организмов. Вероятно, и социумы в значительной мере подчиняются закономерностям нецентрализованной саморегуляции, хотя в них может быть значителен и фактор управления. Основная загадка нецентрализованных систем – это способ поддержания их целостности, интегрированности. Среди всех перечисленных объектов модульные организмы, пожалуй, наиболее интересны с позиции механизмов саморегуляции без участия управляющих органов. К ним относятся все растения, грибы, колониальные беспозвоночные, но не на всех этих объектах просто исследовать организменную интеграцию.
Колониальные гидроиды особо удобны для изучения нецентрализованных систем. Они просто устроены и состоят из множества равнозначных частей: зооидов, побегов, на которых они расположены, и столонов, от которых отходят побеги. Тело двухслойное и полупрозрачное, трубковидное, образует совмещенную пищеварительную и распределительную системы органов. Массивных органов нет. Нервная система развита настолько слабо, что в ней нет даже ганглиев. Других систем органов нет, кроме репродуктивной. Единственная разветвленная трубковидная полость трубчатого тела (ценосарка) объединяет все зооиды в единое целое и является основой интеграции. Все участки тела колониального гидроида пульсируют, сжимаясь и медленно расширяясь несколько раз в час, создавая внутри ценосарка течения заполняющей его жидкости (гидро-плазмы).
По сути, такой модульный организм представлен совокупностью пульсаторов, перемещающих жидкость в общей трубчатой разветвленной полости колониального организма. Интеграция столь необычного организма выражается в перемещении полупереваренной пищи из мест поступления в организм к зонам роста. Особый интерес вызывает способность столь несовершенного организма транспортировать пищевые частицы в удаленные от зооидов зоны роста – к верхушкам столонов. Учитывая “маломощность” отдельных пульсаторов, их множественность и несогласованность, а также протяженность пути доставки пищи и разветвленность ценосарка, сложно было представить, как возможна интеграция колониального организма.
Способ и дальность перемещения частиц пищи в колониальных гидроидах до сих пор вызывают споры (Бурыкин, 2010; Marfenin, Dementyev, 2017; Марфенин, Дементьев, 2019). У гидроидов гидроплазма (жидкость, заполняющая полость тела) приводится в движение множеством самостоятельных пульсаторов. У колониальных гидроидов нет главного пульсатора – сердца, а распределительная система разветвлена, причем ветви не замыкаются друг на друга, а заканчиваются слепо верхушками роста. Под микроскопом видно, что частицы движутся попеременно то в одну сторону, то в противоположную (Hale, 1960; Fulton, 1963; Карлсен, Марфенин, 1976, 1984). Создается впечатление, что перемещение пищи по колонии на значительные расстояния невозможно. Как тогда зоны роста, удаленные от мест получения пищи, могут быть обеспечены питательными веществами?
Благодаря подробным исследованиям функционирования распределительной системы (Марфенин, Дементьев, 2017, 2019, 2020) удалось установить, что протяженные гидроплазматические течения (ГПТ) образуются из цепочек коротких локальных токов. Однако оставался не до конца ясным вопрос о возможности непрерывного перемещения частиц в гидроплазме на дальние расстояния, сопоставимые с длиной основного столона, объединяющего весь колониальный организм. Косвенные данные позволяли предполагать, что такие перемещения возможны. Имелись и отдельные непосредственные наблюдения за частицами (Rees et al., 1970). Однако до сих пор не было специальных исследований, которые показали бы, насколько характерен безостановочный перенос частиц от одного конца колониального организма до противоположного.
Целью работы является определение дальности и траекторий движения взвешенных в гидроплазме частиц у модельного вида гидроидов Dynamena pumila (L., 1758), что необходимо не только для понимания механизма распределения пищи по колониальному организму, но и значительно шире – для определения интеграционных возможностей нецентрализованной биологической системы как таковой.
МАТЕРИАЛ И МЕТОДЫ
Исследование проводили на Беломорской биологической станции им. Н.А. Перцова Московского государственного университета им. М.В. Ломоносова, расположенной в проливе Великая Салма Кандалакшского залива Белого моря.
Объект исследования – гидроид Dynamena pumila (Sertulariidae, Leptothecata, Hydrozoa) – модельный вид, неоднократно использовавшийся ранее в различных экспериментальных исследованиях роста, морфогенеза и образа жизни. Обитает в зоне легкой доступности на границе литорали и сублиторали. Питается мелким зоопланктоном, особенно личинками беспозвоночных. С одного таллома фукуса собирали побеги D. pumila с растущими верхушками, хорошо различимыми по светлой окраске. В лаборатории побеги прикрепляли на предметные стекла, которые помещали в десятилитровые аквариумы с естественной морской водой и активной аэрацией. Воду в аквариумах обновляли два раза в неделю. Ежедневно кормили в течение 1 ч свежевылупившимися науплиями Artemia salina (L., 1758). Температура поддерживалась автоматически в пределах 15 ± 1°С. Подробнее методика культивирования описана ранее (Марфенин, Дементьев, 2018а, б; Dementyev, Marfenin, 2021).
Дальность перемещения частиц по столону определяли визуально, просматривая под микроскопом предметные стекла с колониями при увеличении ×100 на просвет и нанося на схему колонии (рис. 1) посегментно (последовательно по модулям столона) траекторию движения частиц в виде стрелочек. Эта процедура регистрации перемещения частиц в дальнейшем называется визуальным сканированием.
Рис. 1.
Траектории движения взвешенных в гидроплазме частиц в колонии D. pumila № 1–4 (2015). В левом столбце порядковый номер наблюдения; в крайнем правом – количество пройденных модулей столона (знак “‒” означает движение частицы от верхушки столона); во втором справа столбце – время, за которое частица проделала весь путь; сплошные и пунктирные стрелки – направление движения и модули столона, через которые частица прошла; сплошные стрелки – по направлению к верхушке столона, пунктирные – в обратном направлении; σ – отсутствие поступательного движения.

Длина стрелочки соответствует пройденному пути частицей.
Сплошная стрелка маркирует направление (+)ГПТ к верхушке роста столона, пунктирная – обратное направление (‒)ГПТ от верхушки столона. Протяженные ГПТ к верхушке столона называются магистральными, а от верхушки столона к материнскому побегу – компенсаторными. Кроме них могут наблюдаться и локальные короткие ГПТ. Обозначения (+)ГПТ и (–)ГПТ относятся ко всем течениям гидроплазмы: как локальным, так и протяженным.
Если частицы переставали двигаться направленно и начинали вращаться на месте, то подобное состояние мы обозначали символом “σ”.
Также указывали время начала и окончания движения для определения длительности пути. Мы не стали выдерживать постоянной шаг регистрации: начальное время зависело от того, когда обнаруживались четко различимые частицы, за движением которых можно было следить.
Перемещающимися частицами, чьи траектории мы исследовали, могли быть любые визуально отличающиеся от окружения структуры, за которыми можно было следить, не теряя их из виду. Сюда относились:
• частицы неправильной угловатой формы (заметные на фоне остальных округлых частиц);
• скопления частиц, где последние сохраняли постоянное расположение друг относительно друга, например, две мелкие, одна крупная и еще одна мелкая. Такие конгломераты не перемешиваются по ходу движения, благодаря чему можно отследить полную траекторию движения по столону;
• аномально пигментированные частицы (например, очень черные или рыжие).
Наблюдения проведены на трех колониях: № 1–4 (2015), № 2–8 (2015), № 10–8 (2015). Регистрацию дальности в каждой из них проводили неоднократно с перерывом в несколько дней. Колонии № 1–4 (2015) и № 2–8 (2015) активно росли: новые модули столона и побегов у них продолжали формироваться в течение всего периода исследований: 11–18 июля для № 1–4 (2015) и 14–23 июля для № 2–8 (2015). Третью колонию № 10–8 (2015) исследовали в течение трех дней (17–19 июля), однако за это время рост столона и побегов не происходил. Суммарно были зарегистрированы траектории движения 372 частиц (табл. 1).
Таблица 1.
Размер колонии, время последнего кормления и количество частиц, траектория движения которых была отслежена
| № колонии | Общая длина столона, мм | Дата | Последнее кормление | Количество зарегистрированных частиц |
|---|---|---|---|---|
| 1–4 (2015) | 23.4 | 11 июля 2015 г. | Непосредственно перед сканированием | 41 |
| 13 июля 2015 г. | За сутки до съемки | 43 | ||
| 15 июля 2015 г. | За 4 часа до съемки | 34 | ||
| 18 июля 2015 г. | За сутки до съемки | 40 | ||
| 2–8 (2015) | 21.7 | 14 июля 2015 г. | За 3 суток до съемки | 45 |
| 16 июля 2015 г. | За 20 часов до съемки | 33 | ||
| 22 июля 2015 г. | За сутки до съемки | 25 | ||
| 23 июля 2015 г. | За 2 суток до съемки | 18 | ||
| 10–8 (2015) | 13.8 | 17 июля 2015 г. | За 2 суток до съемки | 28 |
| 18 июля 2015 г. | За 15 часов до съемки | 40 | ||
| 19 июля 2015 г. | За 2 суток до съемки | 25 |
Результаты регистрации траектории движения частиц в одной колонии в течение одного дня заносили в отдельный бланк учета.
РЕЗУЛЬТАТЫ
Для трех колоний суммарно было составлено одиннадцать бланков учета траектории движения частиц. Вначале рассмотрим один бланк в качестве примера учета перемещения частиц в столоне колонии, а затем перейдем к обобщенным данным по всем полученным результатам.
18 июля 2015 г. была проведена регистрация перемещения частиц в колонии № 1–4 (2015) (рис. 1), столон которой состоял из шести модулей: пяти сформированных и одного растущего (так называемого нулевого). Колония последний раз получила пищу за сутки до регистрации движения частиц. За время наблюдения мы зарегистрировали 40 траекторий частиц, направленных примерно поровну к верхушке столона (+)ГПТ и от нее (‒)ГПТ. Частицы проходили дистанции длиной от одного до шести модулей, их время в пути иногда составляло менее минуты (0:00), иногда до 2 мин (0:02). Некоторые частицы переходили от ламинарного направленного движения к турбулентному, начиная вращаться на месте (например, наблюдения 2, 6, 9, 14, 16, 19 и т.п.). Другие заходили из столона в побеги, где пропадали из виду (наблюдения 1, 3, 4, 5, 7, 8 и т.п.). В продолжающем расти нулевом модуле столона направленно двигавшиеся частицы были обнаружены всего в пяти случаях из сорока (наблюдения 1, 2, 29, 31, 36).
В других бланках учета картина была сходной. Рассмотрим совокупные характеристики их траекторий.
Больше всего было зарегистрировано траекторий длиной в два модуля столона, как в направлении верхушки столона, так и в обратном (табл. 2, рис. 2). На втором месте по численности находятся траектории длиной в один модуль столона, реже – протяженностью в три модуля. Длинные траектории (четыре модуля и более) также встречались, хотя и в меньшем количестве. Тем не менее таких протяженных пробегов частиц среди магистральных ГПТ (направленных к верхушке столона) было 18%, а среди компенсаторных ГПТ (от верхушки столона) – 11%.
Таблица 2.
Дальность перемещения распознаваемых частиц в столоне при совместном учете всех исследованных колоний D. pumila, ранжированная по числу модулей столона и по направлению
| Число модулей столона, пройденных частицей | Число частиц с данной траекторией | Доля от общего числа, % |
|---|---|---|
| К верхушке столона, (+)ГПТ | ||
| Менее одного | 15 | 8 |
| Один | 47 | 25 |
| Два | 63 | 33 |
| Три | 32 | 17 |
| Четыре | 20 | 11 |
| Пять | 13 | 7 |
| Шесть | 0 | 0 |
| Сумма | 190 | 100 |
| От верхушки столона, (–)ГПТ | ||
| (–) один | 48 | 26 |
| (–) два | 78 | 43 |
| (–) три | 36 | 20 |
| (–) четыре | 15 | 8 |
| (–) пять | 4 | 2 |
| (–) шесть | 1 | 1 |
| Сумма | 182 | 100 |
Рис. 2.
Вариационные ряды дальности перемещения (а) и длительности пробегов (б) распознаваемой частицы в гидроплазме по столону всех исследованных колоний D. pumila.
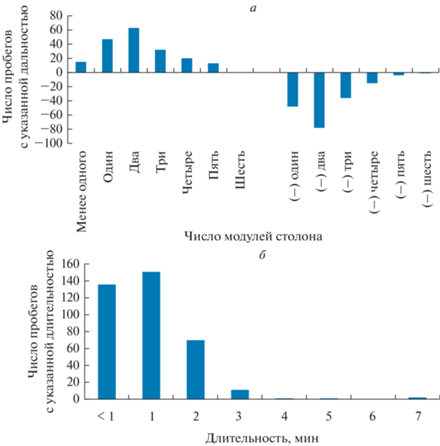
По длительности пробега в обоих направлениях: к верхушке столона и от нее – преобладали краткосрочные перемещения длительностью менее 1 мин (37%), 1 мин (41%) и 2 мин (19%). В совокупности они составляли 97% всех наблюдений (табл. 3, рис. 2). При этом быстрые перемещения были характерны как для частиц, проходивших короткие дистанции (один–два модуля столона), так и длинные, которые проходили всю дистанцию от материнского побега до дистального модуля столона с верхушкой роста всего за 1–2 мин (табл. 4).
Таблица 3.
Длительность пробега частиц по столону всех исследованных колоний D. pumila совместно, ранжированная поминутно
| Длительность пробега частицы, мин | Число пробегов с указанной длительностью | Доля от общего числа, % |
|---|---|---|
| <1 | 136 | 36.6 |
| 1 | 151 | 40.6 |
| 2 | 70 | 18.8 |
| 3 | 11 | 3 |
| 4 | 1 | 0.3 |
| 5 | 1 | 0.3 |
| 6 | 0 | 0 |
| 7 | 2 | 0.5 |
| Сумма | 372 | 100 |
Таблица 4.
Длительность протяженного (три модуля и более) пробега частиц по столону всех исследованных колоний D. pumila
| К верхушке столона, (+)ГПТ | От верхушки столона, (–)ГПТ | ||||
|---|---|---|---|---|---|
| длительность пробега частицы, мин | число пробегов с указанной длительностью | доля от общего числа, % | длительность пробега частицы, мин | число пробегов с указанной длительностью | доля от общего числа, % |
| <1 | 5 | 7.7 | <1 | 3 | 5.4 |
| 1 | 33 | 50.8 | 1 | 28 | 50 |
| 2 | 23 | 35.4 | 2 | 20 | 35.7 |
| 3 | 3 | 4.6 | 3 | 5 | 8.9 |
| 4 | 0 | 0 | 4 | 0 | 0 |
| 5 | 0 | 0 | 5 | 0 | 0 |
| 6 | 0 | 0 | 6 | 0 | 0 |
| 7 | 1 | 1.5 | 7 | 0 | 0 |
| Сумма | 65 | 100 | Сумма | 56 | 100 |
В среднем между очередными протяженными (четыре и более модулей) перемещениями частиц было зарегистрировано 6 ± 4 коротких перемещений (табл. 5). Колония № 10–8 (2015) была небольшой: три модуля столона. Поэтому редкие перемещения частиц более чем на три модуля означают их заход в короткий дистальный участок столона (длиной всего 0.8 мм) с верхушкой роста.
Таблица 5.
Последовательность смены протяженности траекторий частиц в каждой из исследованных колоний, представленных совместно
| № колонии | Номера бланков регистрации | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–4 (2015) | 2–8 (2015) | 10–8 (2015) | |||||||||
| дата | 11 июля 2015 г. | 13 июля 2015 г. | 15 июля 2015 г. | 18 июля 2015 г. | 14 июля 2015 г. | 16 июля 2015 г. | 22 июля 2015 г. | 23 июля 2015 г. | 17 июля 2015 г. | 18 июля 2015 г. | 19 июля 2015 г. |
| 1 | 1 | –4 | –1 | –6 | –3 | 3 | 5 | –1 | –2 | 2 | –1 |
| 2 | –4 | 3 | 2 | 2 | –4 | –3 | –2 | –2 | –2 | 1 | –2 |
| 3 | 4 | –1 | –3 | 5 | 1 | 2 | –3 | 2 | 1 | –1 | 1 |
| 4 | –2 | 0 | –2 | –2 | –2 | –1 | –2 | –2 | –1 | –3 | –1 |
| 5 | 1 | –2 | –1 | –1 | 2 | 2 | 2 | 3 | –4 | 3 | 1 |
| 6 | 3 | 3 | 1 | 2 | 2 | 3 | –3 | –2 | 1 | 2 | 3 |
| 7 | 3 | 2 | 1 | –5 | –3 | –3 | 5 | 1 | –1 | –2 | 2 |
| 8 | –3 | –3 | 5 | 1 | –2 | –3 | –3 | 5 | 1 | 3 | –3 |
| 9 | 2 | 0 | 3 | –4 | –2 | –2 | –2 | –2 | 1 | –3 | 1 |
| 10 | 5 | 1 | –2 | 3 | –2 | 1 | 1 | –3 | –4 | –1 | –2 |
| 11 | –2 | –2 | 0 | 5 | 4 | 1 | –1 | 4 | 2 | 2 | –3 |
| 12 | 2 | 2 | –2 | –4 | 2 | 3 | 1 | 1 | 3 | –2 | 2 |
| 13 | 5 | 2 | 1 | –2 | –2 | –2 | 3 | –2 | 3 | 3 | –1 |
| 14 | –1 | –2 | –2 | 2 | –2 | –2 | –2 | 4 | 1 | 3 | 2 |
| 15 | –3 | 3 | 2 | –5 | 2 | –2 | 2 | –2 | –1 | 2 | –3 |
| 16 | –1 | 2 | 4 | 3 | –3 | 1 | –2 | –2 | 1 | 1 | 2 |
| 17 | 2 | 0 | –4 | –2 | 3 | 4 | –3 | 4 | –3 | 1 | –1 |
| 18 | 3 | –3 | –2 | 3 | 4 | –2 | 2 | –2 | 4 | –3 | –3 |
| 19 | –2 | 0 | 1 | 1 | –3 | –3 | 1 | –2 | 0 | 4 | |
| 20 | 2 | 5 | 1 | 5 | 1 | –1 | –3 | –2 | 2 | –2 | |
| 21 | –1 | –4 | –1 | –1 | –1 | 2 | –1 | 2 | 0 | –3 | |
| 22 | –2 | –2 | –2 | 4 | –2 | –1 | 1 | 3 | –1 | 2 | |
| 23 | 5 | 0 | 2 | –4 | 2 | 4 | –2 | –4 | 1 | 2 | |
| 24 | –2 | 2 | 4 | –2 | –2 | –2 | 4 | 1 | 2 | –3 | |
| 25 | –1 | –1 | 0 | 1 | 3 | –1 | –3 | 2 | 2 | 3 | |
| 26 | 5 | 4 | –4 | –1 | 1 | 2 | –2 | 2 | |||
| 27 | –1 | 4 | –2 | 3 | –4 | –2 | –1 | –1 | |||
| 28 | –2 | –3 | 1 | 4 | 2 | 3 | –1 | –1 | |||
| 29 | 2 | –1 | –2 | –3 | –2 | –3 | 2 | ||||
| 30 | –3 | 0 | 2 | –2 | –2 | –2 | –2 | ||||
| 31 | 2 | –2 | –1 | 2 | 0 | –2 | –2 | ||||
| 32 | 4 | 0 | –2 | 2 | –1 | 1 | –2 | ||||
| 33 | –4 | 2 | 0 | –2 | –2 | 1 | 1 | ||||
| 34 | –1 | –1 | 1 | 3 | 3 | –1 | |||||
| 35 | 2 | 0 | –1 | –2 | –3 | ||||||
| 36 | 3 | 2 | –5 | –2 | –2 | ||||||
| 37 | –1 | –3 | 4 | 3 | 1 | ||||||
| 38 | 1 | –4 | –1 | –1 | 1 | ||||||
| 39 | –5 | 0 | 2 | –2 | –1 | ||||||
| 40 | 2 | 2 | 2 | 1 | 1 | ||||||
| 41 | 5 | 2 | 2 | ||||||||
| 42 | 4 | –1 | |||||||||
| 43 | –2 | 2 | |||||||||
| 44 | –1 | ||||||||||
| 45 | 2 | ||||||||||
Значительных различий в протяженности и продолжительности перемещения частиц к верхушке столона и от нее в сторону материнского побега не обнаружено (табл. 2, 4).
ОБСУЖДЕНИЕ
Хотя сами гидроиды не имеют прикладного значения, они, тем не менее, уже более века используются в качестве натурных моделей для фундаментальных исследований. Проведенная регистрация траекторий перемещения частиц в полости столона уникальна и представляет интерес для понимания принципов функционирования распределительной системы гидроидов, как примера нецентрализованной биологической системы.
О протяженности гидроплазматических течений (ГПТ) у колониальных гидроидов обычно получают представление косвенно несколькими способами.
Проще всего провести непосредственные наблюдения перемещения гидроплазмы под микроскопом. По перемещению взвешенных в ней частиц можно судить о направлении ГПТ, скорости течений и их постоянстве. Однако таким способом нельзя выяснить, как далеко перемещаются отдельные частицы, которые могут поступать из разных побегов, создавая впечатление непрерывного переноса пищи от одного конца колонии к другому.
На самом деле точные данные, полученные на основе цейтраферной микровидеосъемки, дают основание утверждать, что регистрируемые ГПТ непротяженны (Дементьев, Марфенин, 2018, 2019, 2021), и частицы переносятся не более чем на расстояние двух–трех модулей столона.
Третий способ связан с использованием изотопных меток, когда по меченным изотопами частицам можно судить о времени смещения местоположения метки. Этот метод неоднократно применяли при изучении обмена веществ у морских беспозвоночных (Fretter, 1953) и несколько раз при изучении транспорта пищи у разных стрекающих. Оказалось, что миграция изотопных меток может происходить за короткое время (около получаса) на значительное расстояние – на противоположный конец колонии (Rees et al., 1970). Как это происходит, оставалось неясно. Учитывая, что частицы в гастроваскулярной полости у гидроидов с пульсаторно-реверсивной распределительной системой движутся не поступательно, а колебательно, трудно было представить механизм быстрого переноса пищи на дальние расстояния, соразмерные с размерами колониального организма.
Тот же результат можно получить без авторадиографии – всего лишь наблюдая за положением в колонии естественно окрашенных частиц пищи, например, рачка Artemia. Оказалось, что пища за несколько часов, а может быть и быстрее, переносится через всю колонию (Dudgeon, Buss, 1996; Dudgeon et al., 1999). Аналогичные результаты были получены при скармливании гидрантам искусственных флюоресцирующих пищевых частиц (Buss et al., 2015). Также есть краткое описание использования программного обеспечения для определения смещения клеток в полости столона за единицу времени по материалам видеосъемки (Harmata et al., 2013).
Наконец, скорость ГПТ теоретически можно было бы определять лазерной доплеровской анемометрией (Briers, 2007), как и при измерении кровотока в сосудах, но этот метод до сих пор слишком громоздкий относительно миниатюрных гидроидов, требовательных к условиям содержания. Поэтому в исследованиях распределительной системы гидроидов он не нашел применения.
Учитывая изложенные особенности и трудности изучения протяженности переноса пищи у колониальных гидроидов, приходится признать, что до сих пор актуален самый простой метод непосредственного отслеживания движения конкретных частиц. К сожалению, никто раньше планомерно не использовал этот метод, ограничиваясь единичными наблюдениями.
Выяснилось, что прослеженные визуально отдельные частицы либо их скопления могли преодолевать безостановочно предельно большие расстояния (с учетом размеров используемых нами колоний). В нашем случае это расстояние покрывало пять модулей столона. При средней длине модуля столона D. pumila в 3 мм (Марфенин, Дементьев, 2017, 2019) это соответствует дистанции в 15 мм. В исследованных нами колониях расстояния между побегами были больше, и частицы непрерывно проходили до 23 мм – колония № 1–4 (2015).
Величина просвета столона составляет в среднем 0.1 мм, т.е. частицы преодолевают расстояние в 230 раз больше того места, в котором они исходно находились. Если сравнить с размерами самих частиц, то пройденное расстояние в 1–2 тыс. раз больше, чем они сами.
Установлена продолжительность таких единичных актов перемещения частиц. В подавляющем большинстве случаев частицы проходят через весь столон за 1–2 мин, а иногда и менее чем за минуту. Ранее, путем непосредственного визуального отслеживания перемещения отдельных частиц по столону, было показано, что средняя скорость ГПТ в столонах D. pumila равна 200 мкм/с, а максимальная – 800 мкм/с (Карлсен, Марфенин, 1984). При таких скоростях за 1 мин частицы могут пройти внутри ценосаркальной трубки от 12 до 48 мм. Менее чем за 1 мин фронт ГПТ может распространиться на расстояние, равное длине, по меньшей мере, трех модулей, т.е. в среднем на 8–9 мм. В таком случае скорость фронта ГПТ превышает 150 мкм/с (Марфенин, Дементьев, 2019). Столь высокие скорости позволяют чрезвычайно быстро распределять пищу по колониальному организму и кажутся удивительными для прикрепленного малоподвижного гидроида.
Следовательно, транспорт пищевых частиц в этих колониях максимально эффективен, если частицы за один акт перемещения в одну сторону могли быстро проходить расстояние от материнского побега до верхушки роста столона. Полученный нами результат противоречит достоверным данным о протяженности локальных ГПТ, полученным нами же ранее методом стационарной цейтраферной микровидеосъемки перемещения частиц в выбранном для этого участке ценосарка (Марфенин, Дементьев, 2017). Согласно этим данным, протяженность даже мощных ГПТ не превышает двух модулей столона, т.е. не более 7 мм. Как сочетается одно с другим?
На самом деле загадка была уже давно решена в виде гипотетической модели “рокировки объемов гидроплазмы” (Марфенин, 1985). Суть модели в том, что при движении в трубковидной полости столона столб жидкости (гидроплазмы) частично заходит в ближайший побег, а частично проходит дальше. Ближайший побег реагирует сжатием на расширение своей полости из-за притока гидроплазмы (рис. 3). Выходящая из него порция гидроплазмы оказывается сзади столба гидроплазмы, движущегося в столоне. Получается, что головная порция столба в результате захода в побег и затем выхода из него оказывается в хвосте того же столба гидроплазмы, т.е. происходит “рокировка”. Очередной побег начинает выполнять роль двигателя, выталкивая часть гидроплазмы из себя. Поочередное участие побегов в продвижении гидроплазмы в столоне может теоретически обеспечить транспортировку частиц на неограниченное расстояние.
Рис. 3.
Схема “рокировки объемов гидроплазмы” по ходу продвижения порции частиц (№ 1 – выделено темным) в столоне колониального гидроида. Обозначения: римскими цифрами – последовательные этапы продвижения порции частиц; арабскими цифрами – условное обозначение положения порций гидроплазмы. На этапе I все порции гидроплазмы в последовательном порядке от 1 до 6. На этапе VIII – в порядке 2–3–4–5–1–6.
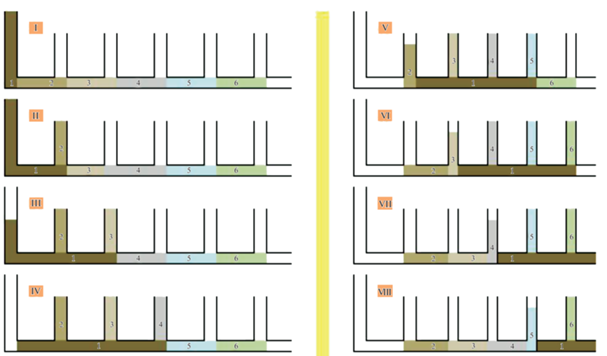
Однако работа этого механизма не столь упорядочена. Передняя порция столба гидроплазмы может не зайти в очередной побег, если его ценосарк сопротивляется расширению, а это происходит в тех случаях, когда ценосарк недавно сжимался, и требуется некоторое время на его релаксацию. Возможно, при получении порции гидроплазмы ценосарк побега не сразу сжимается, а со значительной задержкой. Или же в момент прохождения столба жидкости под побегом этот побег сам начинает выдавливать избыточную гидроплазму, накопившуюся в нем – например, после переваривания пищи.
Все это вносит значительные искажения в теоретически простую и четкую модель транспортировки пищи по колонии. Получается, что модель работает плохо, неупорядоченно, хаотично. Об этом писал Ю.Б. Бурыкин (2010, 2013, 2015). Признав “эстафетный” механизм перемещения частиц от одного побега к следующему, он все же настаивал на том, что подобная распределительная система не способна интегрировать колониальный организм, т.е. неэффективна. На наш взгляд, он переоценил неупорядоченность функционирования нецентрализованного организма (Марфенин, 2016).
Понять, насколько транспорт пищевых частиц не случаен и эффективен, помогают проведенные нами наблюдения. Из обобщенных данных по всем колониям следует, что подавляющее большинство траекторий частиц короткие – один–два модуля столона (табл. 2, рис. 2). Более протяженных траекторий от материнского побега к верхушке столона немного (всего 18%), но они есть, причем довольно быстрые. Частицы преодолевают всю длину столона за 1–2 мин (табл. 4). Следовательно, отдельные ГПТ, переносящие частицы, проходят безостановочно расстояние, в 2–3 раза превышающее возможности локального течения. Этот факт прямо подтверждает гипотезу о механизме “рокировки” порций гидроплазмы в ГПТ. Принципиально важно, что для осуществления такого транспорта частицам вовсе не обязательно заходить в промежуточные побеги, как утверждал Ю.Б. Бурыкин (2013).
Опубликованные нами данные (Марфенин, Дементьев, 2017, 2019) о ежеминутном мониторинге направления перемещения гидроплазмы в колониях D. pumila позволили определить одномоментную протяженность ГПТ и обнаружить, что не только передний конец ГПТ продвигается вперед, но и задний его конец также смещается. Это означает, что протяженное ГПТ существует не только за счет сжатия пульсаторов в породившем его материнском побеге, но и за счет пульсаций ценосарка промежуточных побегов. Возможно два объяснения этого процесса:
1) протяженное ГПТ генерируется исключительно одним побегом, который лишь иногда порождает очень длинное ГПТ, а в большинстве случаев – короткие;
2) протяженное ГПТ генерируется не одним, а множеством побегов, пульсации ценосарка которых в какой-то из циклов способствуют безостановочному прохождению порции гидроплазмы на значительное расстояние.
Подтверждение получает вторая гипотеза. Поскольку в распределительной системе колониальных гидроидов пульсаторы в значительной мере самостоятельны, их взаимодействие слабо упорядочено. Вся система нецентрализованная – без главного пульсатора (сердца) и без нервной регуляции. Поэтому часто она работает как будто хаотично, но протяженные ГПТ, тем не менее, возникают, причем регулярно и с определенным ритмом (Марфенин, Дементьев, 2020).
Следовательно, рассмотренная нами распределительная система пульсаторно-реверсивного типа функционирует достаточно эффективно, несмотря на отсутствие управляющей инфраструктуры. Она обеспечивает время от времени транспортировку частиц в полости колониального гидроида безостановочно от одного конца разветвленного организма к противоположному. Так пища из зоны питания с наиболее крупными побегами и множеством гидрантов поступает быстро к зоне роста, где гидрантов меньше. Продолжительность протяженной транспортировки не превышает двух минут. Для обеспечения зон роста питательными веществами достаточно нерегулярной доставки частиц по столону, тем более экспресс-доставки, что важно при распределении пищи по телу разветвленного колониального организма, поскольку она усваивается клетками гастродермы в течение двух–трех часов (Dudgeon, Buss, 1996; Dudgeon et al., 1999).
Таким образом, распределительная система пульсаторно-реверсивного типа способна далеко и быстро перемещать частицы пищи по столону колонии D. pumila.
Очевидно, что архитектоника колониального организма предопределяет возможность и эффективность дальнего переноса пищи. От размеров пульсаторов (гидрантов, терминальных участков побегов и столонов), а также от диаметра трубки ценосарка и расстояния между побегами зависит протяженность траектории, которую может преодолеть порция гидроплазмы за один акт сжатия пульсаторов. Весьма вероятно, что свойственное многим видам колониальных гидроидов из отряда Leptothecata определенное расстояние между побегами имеет ограничивающее функциональное значение. Благодаря небольшим дистанциям между побегами возможен механизм “рокировки” порций гидроплазмы, за счет которого частицы пищи могут безостановочно проходить значительные расстояния от зон потребления пищи к зонам роста, т.е. экспресс-транспорт.
Механизм нецентрализованного взаимодействия пульсаторов еще не вполне ясен. Однако мы уже знаем, что ведущую роль играют пульсации ценосарка ствола побега, а не гидрантов, что кормление активизирует распределительную функцию, а протяженный транспорт гидроплазмы возможен и при голодании гидроида (Марфенин, Дементьев, 2017, 2019, 2020).
Исследование колониальных гидроидов позволяет постепенно разбираться в фундаментальных особенностях функционирования нецентрализованных биологических систем, широко представленных в природе и до сих пор слабо изученных.
ВЫВОДЫ
1. Больше всего траекторий длиной один–два модуля столона как в направлении верхушки столона, так и в обратном.
2. Протяженные траектории частиц (четыре модуля столона и более) составляют 18% от общего числа наблюдений к верхушке столона (магистральные течения) и 11% от нее (компенсаторные течения). Протяженные перемещения частиц происходят с нестрогой периодичностью.
3. Преобладали краткосрочные перемещения длительностью менее 1 мин (37%), 1 мин (41%) и 2 мин (19%). В совокупности они образуют абсолютное большинство всех наблюдений.
4. Не обнаружено различий в протяженности и продолжительности перемещения частиц к верхушке столона (магистральные течения) и от нее (компенсаторные течения).
5. Полученные результаты подтверждают гипотезу последовательного участия побегов в перемещении гидроплазмы, т.е. возможности хотя бы временного согласования пульсаторов, достаточного для обеспечения экспресс-доставки пищи из зон кормления в зоны роста.
Список литературы
Бурыкин Ю.Б., 2010. Основы функционирования распределительной системы колониальных гидроидов // Онтогенез. Т. 41. № 4. С. 300–311.
Бурыкин Ю.Б., 2013. Эстафетный способ перемещения гидроплазмы в колониях гидроидных полипов // Онтогенез. Т. 44. № 2. С. 115–125.
Бурыкин Ю.Б., 2015. Функционирование распределительной системы у колониального гидроида Dynamena pumila (L., 1758) // Вестн. МГУ. Сер. 16. Биол. Вып. 3. С. 44–48.
Дементьев В.С., Марфенин Н.Н., 2018. Влияние опреснения на рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 79. № 5. С. 376–392. https://doi.org/10.1134/S0044459618050044
Дементьев В.С., Марфенин Н.Н., 2019. Воздействие температуры на рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 80. № 1. С. 22–42. https://doi.org/10.1134/S0044459619010032
Дементьев В.С., Марфенин Н.Н., 2021. Эффективность распределительной системы гидроида Dynamena pumila (L., 1758) при различных абиотических воздействиях // Журн. общ. биологии. Т. 82. № 5. С. 323–336. https://doi.org/10.31857/S0044459621050031
Карлсен А.Г., Марфенин Н.Н., 1976. Упорядоченность перемещения гидроплазмы в колониальном гидроиде Dynamena pumila (L.) (Thecaphora, Sertulariidae) // Журн. общ. биологии. Т. 37. № 6. С. 917–923.
Карлсен А.Г., Марфенин Н.Н., 1984. Перемещение гидроплазмы в колонии у гидроидов на примере Dynamena pumila (L.) и некоторых других видов гидроидов // Журн. общ. биологии. Т. 45. № 5. С. 670–680.
Марфенин Н.Н., 1985. Функционирование распределительной системы пульсаторно-перистальтического типа у колониальных гидроидов // Журн. общ. биологии. Т. 46. № 2. С. 153–164.
Марфенин Н.Н., 2016. Децентрализованный организм на примере колониальных гидроидов // Биосфера. Т. 8. № 3. С. 315–337.
Марфенин Н.Н., Дементьев В.С., 2017. Парадокс протяженных течений гидроплазмы в колониальном гидроиде Dynamena pumila (Linnaeus, 1758) // Журн. общ. биологии. Т. 78. № 4. С. 3–20.
Марфенин Н.Н., Дементьев В.С., 2018а. Продольные пульсации столона у колониального гидроида Dynamena pumila (Linnaeus, 1758) // Журн. общ. биологии. Т. 79. № 2. С. 85–96.
Марфенин Н.Н., Дементьев В.С., 2018б. Рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) в проточной и непроточной кюветах // Журн. общ. биологии. Т. 79. № 2. С. 97–107.
Марфенин Н.Н., Дементьев В.С., 2019. К вопросу о протяженности гидроплазматических течений у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 80. № 5. С. 348–363. https://doi.org/10.1134/S0044459619050051
Марфенин Н.Н., Дементьев В.С., 2020. Побеги как генераторы гидроплазматических течений в колониальном гидроиде Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 81. № 6. С. 421–443. https://doi.org/10.31857/S0044459620060032
Briers D.J., 2007. Laser speckle contrast imaging for measuring blood flow // Optica Applicata. V. 37. P. 328–332.
Buss L.W., Anderson C.P., Perry E.K., Buss E.D., Bolton E.W., 2015. Nutrient distribution and absorption in the colonial hydroid Podocoryna carnea is sequentially diffusive and directional // PLoS One. V. 10. № 9. https://doi.org/10.1371/journal.pone.0136814
Dementyev V.S., Marfenin N.N., 2021. Effect of air exposure on the growth and distribution system in the colonial hydroid Dynamena pumila (L., 1758) // Invert. Zool. V. 18. № 2. P. 69–79. https://doi.org/10.15298/invertzool.18.2.01
Dudgeon S.R., Buss L.W., 1996. Growing with the flow: on the maintenance and malleability of colony form in the hydroid Hydractinia // Am. Nat. V. 147. № 5. P. 667–691.
Dudgeon S.R., Wagner A., Vaisnys J.R., Buss J.W., 1999. Dynamics of gastrovascular circulation in the hydrozoan Podocoryne carnea: the one-polyp case // Biol. Bull. V. 196. P. 1–17.
Fretter V., 1953. Experiments with radioactive strontium (90Sr) on certain molluscs and polychaetes // J. Mar. Biol. Assoc. UK. V. 32. № 2. P. 367–384. https://doi.org/10.1017/S0025315400014636
Fulton C., 1963. Rhythmic movements in Cordylophora // J. Cell. Comp. Physiol. V. 61. № 1. P. 39–51.
Hale L.J., 1960. Contractility and hydroplasmic movements in the hydroid Clytia johnstoni // Quart. J. Microscop. Sci. V. 101. P. 339–350.
Harmata K.L., Parrin A.P., Morrison P., Bross L.S., Blackstone N.W., 2013. Quantitative measures of gastrovascular flow in octocorals and hydroids: Towards a comparative biology of transport systems in cnidarians // Invertebr. Biol. V. 132. P. 291–304.
Marfenin N.N., Dementyev V.S., 2017. Functional morphology of hydrozoan stolons: Stolonal growth, contractility, and hydroplasmic movement in Gonothyraea loveni (Allman, 1859) // Mar. Biol. Res. V. 13. № 5. P. 521–537. https://doi.org/10.1080/17451000.2016.1276292
Rees J., Davis L.V., Lenhoff H.M., 1970. Paths and rates of food distribution in the colonial hydroid Pennaria // Comp. Biochem. Physiol. V. 34. P. 309–316.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии