

Журнал общей биологии, 2022, T. 83, № 3, стр. 151-169
Структура изменчивости палиноморфологических признаков внутри рода Galeopsis L. Hjl. (Lamiaceae) и за его пределами в связи с идеей дивергентной морфологической эволюции
А. Е. Пожидаев 1, *, Н. В. Петрова 1, **
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: pae62@mail.ru
** E-mail: NPetrova@binran.ru
Поступила в редакцию 01.02.2022
После доработки 11.03.2022
Принята к публикации 07.04.2022
- EDN: VKKZEA
- DOI: 10.31857/S004445962203006X
Аннотация
С помощью светового и сканирующего электронного микроскопов исследована индивидуальная и межвидовая изменчивость палиноморфологических признаков девяти видов рода Galeopsis L. Hjl. Типичная форма пыльцы всех видов – 3-кольпатная, с двумя широкими и одним узким мезокольпиумом и биретикулятной макролакунарной микроперфорированной скульптурой. Род палиноморфологически однотипен (отсутствуют существенные различия между видами) и единообразен (в образцах преобладают типичные формы); подроды по морфологии пыльцы не различаются. Для признаков числа и расположения апертур, чередования ширины мезокольпиумов и скульптуры поверхности пыльцы описаны размах и структура изменчивости на видовом и надвидовом (род, семейство) уровне. Изменчивость отсутствует на уровне типичных форм разных видов (там, где ее возникновение можно объяснить механизмами наследования, приспособления и дивергенции); но обнаруживается на индивидуальном уровне (с минимальной генетической изменчивостью – число и расположения апертур, ширина мезокольпиумов), либо на уровне разных родов (генетически несовместимых – скульптура поверхности, ширина мезокольпиумов). Наблюдаемая индивидуальная изменчивость разных признаков и таксономических групп разного ранга может быть без остатка (с точностью до живого тела) сведена в непрерывные транзитивно упорядоченные (таксон- и мерон-неспецифично и рангово-независимо) геометрические ряды. Структура изменчивости палинологических признаков внутри р. Galeopsis и за его пределами рассмотрена с точки зрения нетипологической концепции морфологического многообразия – как система параллелизмов, в которой гомологические ряды не могут быть отделены от гетерологических (случаи негомологического сходства), а типичные формы от отклоняющихся.
Пыльца, содержащаяся в одном пыльнике или бутоне – это возникающая в результате бесполого размножения популяция близкородственных мужских гаметофитов, генетическая изменчивость которых ограничена результатами рекомбинации родительских генов одного вида; это максимально полное поколение живых тел, которое полнее быть не может (близкое к теоретически возможному, на уровне онтологически целого, где смерти нет; Пожидаев, 2015); это естественная порция и целостный образец (в отличие от спорофитов, поколение которых предельно изрежено катастрофической элиминацией и доступно для наблюдения на случайных выборках). Это результат мейоза (с кроссинговером, как механизмом рекомбинации генов и характерным соотношением частот родительских аллелей в одной тетраде 1 : 1 или 2 : 2), а не митоза (как механизма комбинирования хромосом с менделевским соотношением 1 : 3 частот генотипов первого поколения в простейшем случае моногибридного скрещивания двух гетерозигот). Исследование расположения отклоняющихся форм пыльцы у видов с не распадающимися тетрадами показало, что соотношение форм в постмейотической тетраде может быть произвольным, любым (Пожидаев, 2015, табл. 2).
Бесполое размножение – без полового процесса и всего, что с ним связано. То есть до и без явлений, связанных с механизмами комбинирования признаков (диплоидность, рецессивность и доминирование, скрещивание, расщепление, отбор, вымирание) – вне коннотаций (смыслов), составляющих немалую часть содержания, которое мы вкладываем в такие широкие идеи, как наследственность и генетика. Поэтому многообразие палиноморфологических признаков конкретных таксонов представляет интерес как модельный объект для исследования механизмов возникновения изменчивости самой по себе, в собственном значении и почти абсолютном виде, не смазанная явлениями, связанными с комбинированием признаков (скрещивание, расщепление, доминирование, элиминация, отбор).
Обсуждая изменчивость любого таксономического уровня, следует помнить, что существует предметно и доступно наблюдению (объективно) только многообразие и изменчивость индивидуальных живых тел (единичные предметы, натуралии). Любая другая изменчивость – это всегда часть, аспект одной и той же генеральной совокупности – индивидуальной изменчивости. Другой изменчивости не существует – это единственная объективная реальность, эволюцию которой мы пытаемся объяснить (или для начала верно описать). Поэтому мы можем на равных основаниях сравнить друг с другом изменчивость внутри вида, на уровне родов и в любых других таксонах и рангах.
Исследование индивидуальной изменчивости – это не описание повторяющихся явлений (универсалии, общие понятия) – продукт умозрения, который требует обоснования и допускает фальсификацию (и во многом определяется познавательной ситуацией; Любарский, 2018; Павлинов, 2019), а неопровержимое свидетельство уникальных событий (натуралии, единичные предметы), результат наблюдения, который может быть подтвержден предметно и документально (коллекционный образец, фотография), существование которого невозможно оспаривать, даже если оно противоречит результатам логической дедукции или здравому смыслу.
Содержимое пыльника (результат бесполого размножения; генетически сходная популяция) может служить экстремальной моделью (максимальная полнота, объем, экстенсионал, при минимальной сложности, содержание, интенсионал) индивидуальной изменчивости полного синхронного поколения однородственной генеалогии (род, клада; близкородственные виды) – все без исключения формы и все без исключения потомки одного предка.
ТАКСОНОМИЧЕСКИЕ УРОВНИ И РАЗМАХ ИЗМЕНЧИВОСТИ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ ПЫЛЬЦЫ
Изучение признаков пыльцы различных таксономических групп показало (Pozhidaev, 1993, 1995, 2000а, b, 2002; Пожидаев, 2001, 2015; Шелудякова и др., 2017; Gavrilova et al., 2018; Григорьева и др., 2019), что изменчивость пыльцы отличается от изменчивости морфологических признаков спорофитов (по которым строится система таксонов) значительным своеобразием. Для признаков пыльцы характерна особая стабильность и единообразность. Большинство пыльцевых зерен в пыльнике в одном образце имеют типичные (нормальные, обычные) признаки. На внутривидовом уровне (в разных образцах одного вида, в панмиктических популяциях, у скрещивающихся разновидностей) существенная изменчивость обычно отсутствует.
По признакам пыльцы виды внутри рода (близко-родственные виды, состоящие в кровном родстве, но обычно генетически изолированные) различаются незначительно и редко, либо четкая разница между разными видами может отсутствовать вовсе. Таксономически значимые различия подтаксонов внутриродового ранга по форме пыльцы встречаются гораздо реже. Например, в р. Sideritis (сем. Lamiaceae) 6-руговая тетраэдр-форма является типичной для подрода Empedoclea, распространенного в западном средиземноморье; 3-кольпатная форма – для подрода Hesiodia; а 4-кольпатная форма – для подродов Burgsdorfia, Siderites, Empedocliopsi, Marrubiasrtum из восточного средиземноморья (Huynh, 1972).
Для признаков пыльцы более характерна макротаксономическая, надродовая изменчивость (между разными родами, в разных семействах, порядках и классах – у заведомо неблизкородственных групп, не состоящих в прямом родстве и генетически полностью несовместимых). Например, форма пыльцы (число и расположение апертур) характеризует группы высокого систематического ранга. Так, 3-кольпатная форма с меридионально ориентированными апертурами встречается как типичная у большинства таксонов подкласса двудольных; а моно-сулькатные и сулькулятные формы с полярным и суб-полярным расположением апертуры типичны для подкласса однодольных и класса голосеменных растений (Куприянова, Алешина, 1967; Walker, Doyle, 1975; Campo, 1976; Blackmore, Crane, 1998).
Практика определения таксономической принадлежности пыльцы по ее морфологии (пыльцевой анализ; Сладков, 1967) свидетельствует, что по признакам пыльцы разные виды в большинстве родов мало различимы. До уровня вида пыльца может быть определена, когда выбор не велик – в некоторых родах, обычно с малым числом видов, или для узко-региональных флор с небольшим числом видов во флоре; иногда удается уверенно определять род, чаще и достовернее удается определить семейство.
Более подробное изучение свойств изменчивости в далеких родах (Acer – Pozhidaev, 1993; Aesculus – Pozhidaev, 1995; Krameria – Pozhidaev, 2002; Scrophularia –Шелудякова и др., 2017; Artemisia – Григорьева и др., 2009, 2018; Еuonymus – Gavrilova et al., 2018; Nicotiana – Григорьева и др., 2019; Physalis, Cestrum, Rubus, Euphorbia – готовятся к публикации) показало, что признаки пыльцы, рассмотренные на большом числе видов и их образцов, очень быстро теряют свою таксономическую специфичность. Чем полнее представлен в исследовании состав систематической группы растений (чем полнее число исследованных видов и больше образцов), тем чаще вместо специфичности признаков пыльцы приходится сталкиваться с непрерывной неразличимостью их переходных состояний, комбинативностью и параллелизмом изменчивости. А чем менее подробно изучен таксон, тем проще представить эту изменчивость дискретной и специфической, тем удобнее оказывается типологический способ описания – классификация.
Так, в образцах 45% видов исследованных родов в пределах одного бутона или пыльника была обнаружена индивидуальная изменчивость формы пыльцевого зерна (числа и расположения апертур; Пожидаев, 2015). В индивидуальной изменчивости популяции генетически сходных гаметофитов возникают типичные формы далеких, неродственных таксонов, одинаковые отклонения и их непрерывные и геометрически закономерные ряды. В этой изменчивости выявляется сходство форм и чрезвычайно высокая упорядоченность, транзитивность параллелизмов (In Memoriam, 2007), необъяснимые с типологической и генетической точек зрения, выходящие далеко за рамки представлений о родстве таксонов и гомологии структур (ряды таксон- и мерон-неспецифичны и рангово-независимы; Pozhidaev, 1998, 2002).
Учитывая биологические особенности пыльцы как модельного объекта для описания свойств биологической изменчивости в чистом виде, свободной от явлений скрещивания, расщепления, доминирования, вымирания и отбора признаков, возникает задача сравнения свойств палиноморфологических признаков (структура, размах) в конкретных таксонах семенных растений равного и разного ранга.
Объектом такого исследования послужила пыльца видов небольшого р. Galeopsis L. (пикульник) типового подсемейства Lamioideae сем. Lamiaceae. Несмотря на небольшой объем, р. Galeopsis является сравнительно сложным в таксономическом и номенклатурном отношении родом. Сложность его систематики во многом определяется активными процессами межвидовой гибридизации, затрудняющими понимание границ видов и подродов. Виды пикульника распространены в Европе и большей части Азии (Юзепчук, 1954; Townsend, 1972; Гладкова, 1978; Harley et al., 2004). Занимая достаточно широкий ареал, этот род вошел в большинство крупных флористических сводок, однако система р. Galeopsis, его объем и родственные связи остаются дискуссионными и продолжают исследоваться.
До сих пор спорными остаются вопросы внутриродового подразделения, их ранг и входящие в них виды. В разное время предпринимались попытки разделить р. Galeopsis на группы разного ранга. Работа Грэя (Gray, 1821) обозначила первую попытку выделения надвидовых таксонов Galeopsis. В составе рода он описал две группы неопределенного ранга: Tetrahit, в которую вошли виды G. nodosum S.F. Gray (= G. tetrahit L.), G. versicolor Gurt. (= G. speciosa Mill.), и Ladanum с видами G. parviflora Lam. (= G. ladanum L.), G. angustifolia Ehrh. ex Hoffm., G. segetum S.F. Gray (= G. ladanum L.), G. ochroleuca Lam. (= G. segetum Necker). Виды Tetrahit отличаются стеблем, имеющим утолщения под узлами, наличием в опушении щетинковидных волосков и шиловидными зубцами чашечки; виды Ladanum не имеют стеблевых утолщений под узлами и щетинковидных волосков и отличаются треугольными зубцами чашечки.
Райхенбах (Reichenbach, 1830) выделенные Грэем группы рассматривал в ранге подродов, отмечая, что для видов подрода Tetrahit (S.F. Gray) Reichenb. характерна слегка изогнутая трубка венчика и стебель, утолщенный под узлами, а для представителей подрода Ladanum (S.F. Gray) Reichenb. – стебель без утолщений под узлами и прямая трубка венчика. В подрод Ladanum Райхенбах отнес G. angustifolia Ehrh. ex Hoffm., G. ladanum L., G. ochroleuca Lam. и G. intermedia Vill. (последние два вида являются синонимами G. ladanum L.). В подрод Tetrahit он поместил G. pubescens Bess., G. bifida Boenn., G. tetrahit L., G. versicolor Curt. (= G. speciosa Mill.) и G. acuminate Reichenb. (= G. tetrahit L.). Такое разделение рода принял Брике в первой и, пожалуй, единственной монографии по р. Galeopsis (Briquet, 1893). Виды обоих подродов Брике различал по наличию или отсутствию утолщений стеблей в узлах, а также по особенностям опушения. С.В. Юзепчук во “Флоре СССР” (1954) и Таунсенд во “Flora Europaea” (Townsend, 1972) также придерживаются разделения на два подрода.
Славикова (Slavíková, 1963), изучив строение нектарников, утолщений под узлами, особенности опушения, предложила рассматривать р. Galeopsis s.l. как два самостоятельных рода: Galeopsis и Ladanum; далее в совместной статье с Пужаром изменила неподходящее в номенклатурном отношении название выделенного р. Ladanum Gilib. на Ladanella Pouzar et Slavíková (Pouzar, Slavíková, 2000). Таким образом, основными диагностическими признаками р. Ladanella являются более короткие по сравнению с видами Galeopsis стебли, отсутствие утолщений под узлами побегов, отсутствие в опушении щетинковидных волосков, более узкие листья, значительно более короткие (по сравнению с центральной долей) боковые доли нижней губы венчика.
Однако объем и структура отдельных подродов Galeopsis, как и их подразделение на два самостоятельных рода остаются спорными, поскольку вышеперечисленные морфологические признаки спорофитов, на основе которых осуществляется деление на роды, не являются достоверно постоянными и имеют собственную изменчивость (Тюнникова, 2006). В настоящее время, благодаря современным молекулярным методам филогенетических исследований, прояснились многие вопросы межродовых связей в подсемействе Lamioideae (Bendiksby et al., 2011; Olmstead, 2016). Однако положение р. Galeopsis до сих пор остается не до конца ясным. Недавние исследования хоть и подтверждают родственную связь между родами Galeopsis и Betonica L., но степень их родства низкая, что стало основным аргументом в пользу создания отдельной монотипной трибы Galeopseae. В подсемействе она расположена между трибами Betoniceae и Stachydeae (Zhao et al., 2021).
Пыльца видов р. Galeopsis не исследовалась в полном объеме. В литературе имеются описания и изображения пыльцы лишь небольшого числа видов: палиноморфологические данные были получены А.Е. Пожидаевым (1989), Абу-Асабом и Кантино (Abu-Asab, Cantino, 1994), исследованы размеры пыльцевых зерен (Müntzing, 1928). Фрагментарность палиноморфологических данных о видах р. Galeopsis не позволяет в полной мере оценить степень морфологической изменчивости и уровень таксономической специфичности (таксономическую значимость) морфологических признаков пыльцы.
Цели данной работы – описание свойств индивидуальной изменчивости морфологических признаков пыльцы конкретных таксонов, на примере р. Galeopsis; сравнение размаха изменчивости исследуемых признаков пыльцы в группах разного таксономического объема и ранга как модельного объекта для изучения предполагаемых механизмов ее эволюционного возникновения.
МАТЕРИАЛ И МЕТОДИКА
Материалом для исследования послужила пыльца девяти видов р. Galeopsis, собранная с гербарных экземпляров, хранящихся в Гербарии БИН РАН (LE), Гербарии Удмуртского Государственного университета (UDU), а также из личных сборов (табл. 1). Для исследования пыльцы отбирались зрелые нераскрывшиеся бутоны. Созревание пыльцы происходит задолго до окончания бутонизации, поэтому в бутонах, готовых открыться, вся пыльца зрелая, на одной финальной стадии – готовая к высеванию.
Таблица 1.
Исследованные образцы и состав подродов р. Galeopsis
| Исследованный образец | Иллюстрации |
|---|---|
| Subgen.Ladanum: | |
| G. segetum Necker. Schleswig-Holstein, Lauenburg. K. Larsen, L. Holm-Nielsen, S. Jeppesen. 136. (LE) | рис. 1а–е |
| G. pyrenaica Bartl. Pyrenees-Orientalis, Banyuls-Sur-Mer. C. Ballot. 831. (LE) | рис. 1ж–н |
| G. angustifolia Ehrh. ex Hoffm. Калининградская обл., Гурьевский р-н. Петрова Н., Губарева И., Халаим А. (личные сборы) |
рис. 1о–у |
| G. angustifolia. Bohemia centr., silvatica Křivolátsko, opp. Beroun, pagus Žloukovice. Štĕpánek J. (personal collection) |
рис. 1ф |
| G. ladanum L. Псковская обл., Великие Луки, Конечная Г.Ю. (LE) | рис. 2а–е |
| Subgen.Galeopsis: | |
| G. bifida Boenn. Костромская обл, Галичский р-н, с. Михайловское. Леострин А. (личные сборы) |
рис. 2ж–н |
| G. bifida. Cанкт-Петербург, парк БИН. Петрова Н.В. (личные сборы) | рис. 2о |
| G. speciosa Mill. Новгородская обл. Боровичский р-н, окр. д. Ровное. Петрова Н.В. (личные сборы) |
рис. 2н–с |
| G. speciosa Mill. Ленинградская обл., пос. Юкки. Петрова Н.В. (личные сборы) | рис. 2т |
| G. pernhofferi Wettst. in Kerner. Удм. АССР, Болезино Мельников Д.Г. (UDU) | рис. 3а–е |
| G. tetrahit L. Санкт-Петербург, парк БИН. Петрова Н.В. (личные сборы) | рис. 3ж–л |
| G. tetrahit. Новгородская обл., г. Валдай. Петрова Н.В. (личные сборы) | рис. 3м |
| G. pubescens Bess. Cанкт-Петербург, ст. Ораниенбаум-2. Цвелев Н.Н. (LE) | рис. 3н–с |
| G. pubescens. Псковская обл., Себеж. Конечная Г.Ю. (LE) | рис. 3т |
Для исследования под световым микроскопом (СМ) пыльцу предварительно подвергали ацетолизной обработке по стандартной методике (Erdtman, 1952). Для исследования деталей строения поверхности пыльцевых зерен и влияния обработки на его форму как ацетолизированную, так и необработанную пыльцу исследовали с помощью сканирующего электронного микроскопа (СЭМ) JEOLJSM-6390 в центре коллективного пользования Ботанического института им. В.Л. Комарова РАН. Размер пыльцевых зерен измеряли на ацетолизированных препаратах с помощью окулярмикрометра. В каждом образце измеряли 15–20 пыльцевых зерен. Влияние влажности, различие в форме и размерах сухой и набухшей пыльцы у яснотковых описывал еще Моль (Mohl, 1834).
РЕЗУЛЬТАТЫ
Исследование морфологии пыльцы девяти видов р. Galeopsis показало (рис. 1–3), что род палиноморфологически однотипен (отсутствуют существенные различия между видами) и чрезвычайно единообразен (в образцах преобладают типичные формы).
Рис. 1.
Морфология пыльцы видов Galeopsis; типичные формы: а–е – G. segetum; ж–н – G. pyrenaica; о–ф – G. angustifolia. а, в, ж, н, о, р, с – вид с полюса; б, г, з, к, л, т – вид с экватора; д, м, н, у – скульптура необработанной пыльцы; е, ф – скульптура ацетализированной пыльцы.
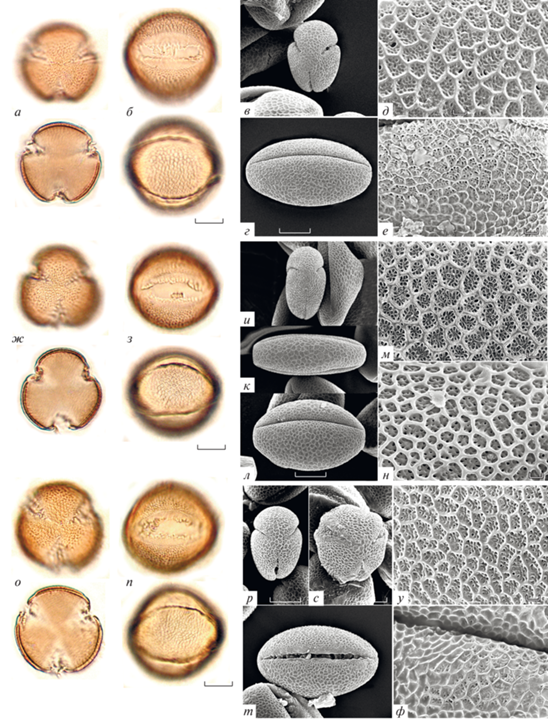
Рис. 2.
Морфология пыльцы видов Galeopsis; типичные формы: а–е – G. ladanum; ж–о – G. bifida; п–ф – G. speciosa. а, в, ж, и–л, п, с – вид с полюса; б, г, з, м, р, т – вид с экватора; д, н, у – скульптура необработанной пыльцы; е, о, ф – скульптура ацетализированной пыльцы.
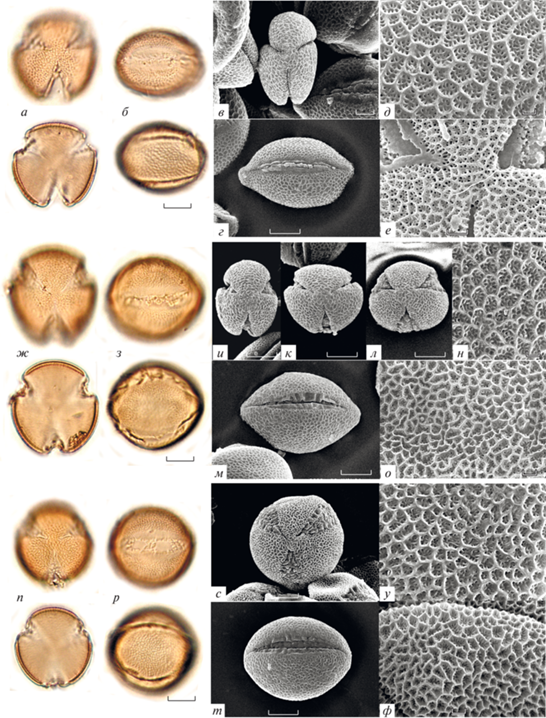
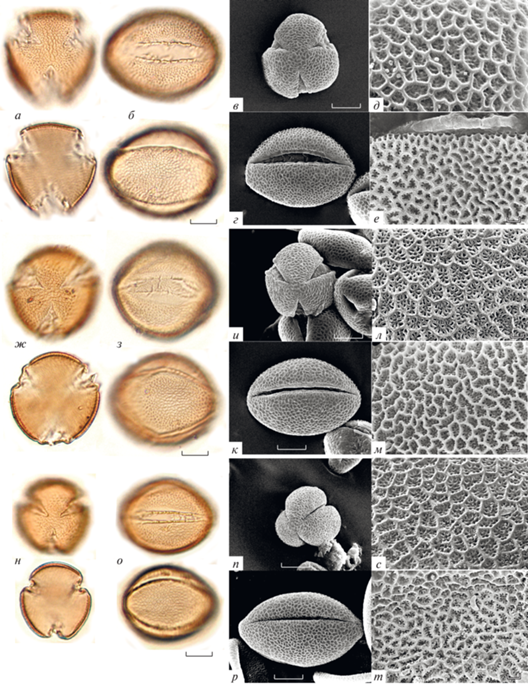
Рис. 3.
Морфология пыльцы видов Galeopsis; типичные формы: а–е – G. penthofery; ж–м – G. tetrachit; н–т – G. pubescens. а, в, ж, и, н, п – вид с полюса; б, г, з, к, о, р – вид с экватора; д, л, с – скульптура необработанной пыльцы; е, м, т – скульптура ацетализированной пыльцы.
Апертуры
Роды и подсемейства сем. Lamiaceae резко распадаются на две группы: с типичной 3- и 6-кольпатной пыльцой (Wunderlich, 1967). Род Galeopsis принадлежит к подтрибе Lamiinea подсем. Lamioideae, типичная форма пыльцы всех видов в которой 3-кольпатная. Апертуры – простые кольпы, неширокие, с заостренными, слегка оттянутыми концами; край кольпы волнистый, неровный. Мембрана покрыта разноразмерными бугорками неправильной формы, в дегидратированном состоянии глубоко западает между мезокольпиумами и может быть не видна.
Форма пыльцевого зерна
Разные признаки формы пыльцевого зерна могут быть охарактеризованы разными способами. Форму описывают по очертанию пыльцевого зерна с полюса и с экватора (овальные, округлые, округло-треугольные, ангулеапертурные, планапертурные (Куприянова, Алешина, 1967; Punt et al., 1994)); по объемной форме (сфероидальные, эллипсоидальные, тетраэдрические); по соотношению длин полярной оси и экваториального диаметра у биполярных форм (P/E индекс; облятная, пролятная), по числу и расположению апертур (3-кольпатная, 6-руговая, 1-сулькатная).
Практика применения существующей терминологии такова, что словосочетание “3-кольпатная пыльца” может обозначать и вариант формы пыльцевого зерна, и тип расположения апертур. Далее, при обсуждении формы пыльцевого зерна будем иметь в виду число и расположение апертур. При описании мы не используем понятие “тип”, подразумевающий (исторически) исходную дискретность изменчивости, еще до начала ее изучения.
Очертание пыльцевого зерна
Очертание пыльцевых зерен всех изученных видов с полюса округло-треугольное, ангулеапертурное; очертание с экватора широко-овальное, полюса иногда слегка приподняты.
На очертание сильно влияет степень гидратации пыльцевой стенки и содержимого пыльцевого зерна и ацетолизная обработка, уничтожающие это содержимое. При гидратации и в результате обработки происходит раздувание пыльцевого зерна за счет раскрывания кольп и уменьшения кривизны мезокольпиумов. Очертание с полюса может меняться от сильно сплюснутого с боков, билательно-симметричного (в слабо гидратированном состоянии; рис. 1в, и, р, 2в, и), до почти округлого (в гидратированном или в ацетолизированном состоянии; рис. 1с, 2к, л, с).
Чередование ширины мезокольпиумов
У всех видов Galeopsis в очертании пыльцевых зерен с полюса, особенно в слабогидратированном состоянии, бывает видно, что пыльцевое зерно сильно сплющено; мезокольпиумы немного различаются по ширине – два широких мезокольпиума чередуются с одним более узким. Такая билатеральная симметрия пыльцевых зерен (рис. 4а) характерна для представителей всей подтрибы Lamiinea подсем. Lamioideae (Wunderlich, 1967; Abu-Asab, Cantino, 1994).
Рис. 4.
Типичные и отклоняющиеся формы расположения апертур пыльцы Galeopsis, некоторых родов сем. Lamiaceae и других семейств цветковых: а – Wiedemannia orientalis; б – Scutelaria nepetoides; в – Salvia splendens; г – Kudrjachevia jacub; д, е – Galeopsis bifida; ж – G. speciosa; з – G. angustifolia; и – G. segetum; к – G. bifida; л, м – G. penhoffery; н, о – Lamium intermedium; п – Sideritis vulgare; р – Stachys turcomanica. с–х – отклоняющаяся пыльца формы B из разных семейств: с – Filipendula ulmaria (сем. Rosaceae); т – Solanum tuberosum (сем. Solanaceae); у – Medicago sativa (сем. Fabaceae); ф – Veronica spicata (сем. Scrophulariaceae); х – Valeriana officinalis (сем. Valerianaceae). Буквенные обозначения форм см. на рис. 6. а, б, в, г, н, п, с–х – вид с полюса; е–л – вид с экватора; а, б – типичная форма; д–з, л–п, с–х – форма С; и, к – форма D; м, р – форма Е; д, е, с–х – верхний и нижний фокус; а–в – индивидуальная изменчивость по чередованию ширины мезокольпиумов; г – отклоняющееся 4-кольпатное и типичное 6-кольпатное пыльцевые зерна; н, о – отклоняющаяся форма В и типичная 3-кольпатная форма.
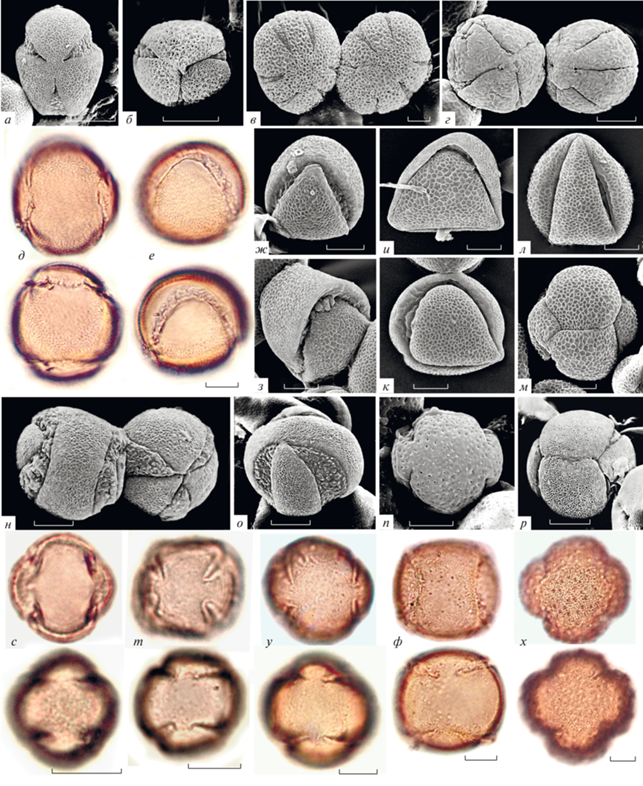
Среди групп с 3-кольпатной пыльцой в сем. Lamiaceae встречается альтернативный вариант чередования ширины мезокольпиумов (два узких, один широкий) – эта форма является характерным отличием пыльцы р. Scutellaria и подсем. Scutellarioidea (рис. 4б). Таким образом, в группах губоцветных с типичной 3-кольпатной пыльцой признак чередования ширины мезокольпиумов разделяет надродовые таксономические группы (подсемейства). Тот же признак чередования ширины мезокольпиумов в группах губоцветных с 6‑кольпатной типичной пыльцой (подсем. Nepetoidea, Salvioidea) характеризует разные роды (Pozhidaev, 1992), но может иметь изменчивость на индивидуальном уровне (рис. 4в). Ранее (Pozhidaev, 2000а), на примере рядов отклоняющихся форм поликольпатной пыльцы (с числом кольп 4–8 и более) были описаны более сложные закономерности в чередовании ширины мезокольпиумов, наборы возможных и существующих изомеров и их зеркально-симметричные варианты.
Условия вакуума при исследовании пыльцы под сканирующим микроскопом и ацетолизная обработка влияют на пыльцевое зерно по-разному (диаметрально противоположно: в вакууме зерно съеживается, при ацетолизе раздувается). В условиях вакуума и в сухом состоянии характерные признаки формы пыльцевого зерна (сплюснутые очертания в полярной проекции, чередование ширины мезокольпиумов) лучше различимы. Технически ширина мезокольпиумов измеряется не по дуге (поверхность пыльцевого зерна), а по хорде (соединяющей края мезокольпиума). Поэтому в дегидратированном (съежившемся, высушенным вакуумом, пусть в разной степени) состоянии признак соотношения ширины мезокольпиумов пыльцевого зерна виден лучше (сильнее сказывается на очертании в полярной проекции). При раздувании пыльцевого зерна, вызванном обработкой или гидратацией, происходит уменьшение кривизны мезокольпиумов (длина хорды увеличивается) и признак становится менее заметным.
Число и расположение апертур на пыльцевом зерне
Далее, говоря о форме пыльцевого зерна, будем иметь в виду число и расположение его апертур.
Для всего р. Galeopsis типичной формой пыльцы по числу и расположению апертур является 3-кольпатная форма. Кроме типичной 3-кольпатной формы пыльцы в пяти образцах (G. speciosa, G. angustifolia, G. segetum, G. bifida, G. penhoffery) из 15 изученных обнаружены единичные пыльцевые зерна, отклоняющиеся от типичной по числу и расположению апертур (рис. 4д–м). Отклоняющиеся формы имеют нормальную стратификацию слоев оболочки, скульптуру, строение апертур и размеры, близкие к типичным.
Все обнаруженные формы расположения апертур (типичные и отклоняющиеся) являются членами единого непрерывного и ненаправленного геометрического ряда (рис. 6а), описанного ранее (Pozhidaev, 1993, 1998, 2000а), в котором наиболее простая форма с циклической апертурой, лежащей в одной плоскости (рис. 6а, форма А), может изгибаться асимметрично в сторону одного из полюсов (рис. 6а, А–F), и за счет возникновения дополнительной апертуры постепенно превращаться в 3-бороздную форму. Либо циклическая апертура той же формы постепенно симметрично изгибается, как шов на теннисном мячике (рис. 6а, формы В и С; рис. 4д–з), и за счет возникновения двух дополнительных апертур, накрест лежащих на противоположных сторонах (рис. 6а, форма D; рис. 4и, к), постепенно преобразуется в 6-руговую форму с апертурами, расположенными глобально по ребрам тетраэдра (рис. 6а, форма Е; рис. 4м).
Из отклонений в изученных образцах Galeopsis чаще всего встречаются форма с единой слитной циклической апертурой, закрученной как шов на теннисном мячике (рис. 4д–з), форма с четырьмя W-образно попарно скошенными кольпами (рис. 4л), а также форма с шестью ругами, расположенными по ребрам тетраэдра (рис. 4м). Остальные промежуточные варианты более редки и единичны (рис. 4и, к).
Следует отметить, что во всех исследованных образцах р. Galeopsis, как и в сем. Lamiaceae в целом, признаки формы пыльцы являются достаточно стабильными, отклоняющиеся формы крайне малочисленны (Pozhidaev, 1992). Тем не менее расположение апертур даже этих единичных отклонений, с частотами встречаемости, стремящимися к нулю, не является случайным, а в точности следует описанной геометрической закономерности (рис. 6а, А–G). Хотя некоторые формы этого ряда в образцах изученных видов Galeopsis не обнаружены, они зарегистрированы во многих других родах подсем. Lamioideae (рис. 4н–р).
Скульптура мезокольпиумов
Все изученные виды Galeopsis имеют биретикулятную макролакунарную микроперфорированную скульптуру, широко распространенную в сем. Lamiaceae (Пожидаев, 1989). У всех видов Galeopsis перфорации сетки первого порядка мелкие, иногда слегка вытянутые и радиально-ориентированные, расположены в лакунах неправильной формы сетки второго порядка, образованных гладкими и неширокими перегородками (мури). В лакунах насчитывается от 7 до 25 перфораций. При исследовании в СЭМ видно, что ближе к краю мезокольпиумов ячеи второй сетки становятся мельче. По краю вдоль кольп поверхность мезокольпиумов гладкая, образует широкую или более узкую кайму, иногда волнистую (в зависимости от степени гидратации оболочки пыльцевого зерна и его содержимого).
Ацетолизная обработка оказывает существенное влияние на размер элементов скульптуры – размер ячей сетки второго порядка уменьшается, рисунок оболочки заметно съеживается (рис. 1е, ф, 2е, о, ф, 3е, м, т), вероятно, из-за удаления при обработке частей пыльцевой оболочки, не содержащих спорополленин. Уменьшение размеров элементов скульптуры может показаться неожиданным, учитывая, что ацетолиз приводит к “раздуванию” пыльцевого зерна (за счет раскрывания кольп и уменьшения кривизны мезокольпиумов).
Размеры пыльцевых зерен
Пыльца изученных видов Galeopsis имеет средние размеры (табл. 2). Самая крупная пыльца у G. segetum (полярная ось 39.0–49.0 мкм, экваториальный диаметр 29.8–39.3 мкм; рис. 1ф–е); самые мелкие пыльцевые зерна обнаружены у G. pyrenaica (полярная ось 34.7–39.3 мкм, экваториальный диаметр 24.8–32.2 мкм; рис. 1ж–н) и G. pubescens (рис. 3н–т). Размеры в пределах вида варьируют незначительно и не превышают 10 мкм; максимальный размах изменчивости этого признака составляет 9.5 мкм у пыльцевых зерен G. pernhofferi (рис. 3а–е) и G. pubescens. Разделить изученные виды на какие-либо группы по размерам не удается из-за варьирования признака внутри видов и сильного перекрывания значений у разных видов.
Таблица 2.
Размеры пыльцы исследованных видов р. Galeopsis и число хромосом в их диплоидном наборе
| Таксон | Диплоидный набор хромосом (2n) | Полярная ось (мкм); среднее значение и стандартное отклонение | Экваториальный диаметр (мкм); среднее значение и стандартное отклонение |
|---|---|---|---|
| Subgen. Ladanum | |||
| G. pyrenaica | 16 | 39.32–34.704; 36.88 ± 1.88 | 32.22–24.84; 28.78 ± 2.93 |
| G. ladanum | 16 | 43.43–38.08; 35.60 ± 5.53 | 36.56–26.82; 30.40 ± 4.06 |
| G. segetum | 16 | 49.06–39.05; 43.99 ± 8.12 | 39.24–29.79; 33.32 ± 5.83 |
| G. angustifolia | 16 | 48.94–45.06; 47.81 ± 1.48 | 28.30–23.18; 27.11 ± 5.77 |
| Subgen.Tetrachit | |||
| G. pernhofferi | 32 | 47.52–31.50; 41.71 ± 7.93 | 30.21–20.90; 26.64 ± 4.49 |
| G. pubescens | 16 | 46.55–34.50; 40.67 ± 4.86 | 39.50–27.88; 33.74 ± 3.22 |
| G. speciose | 16 | 44.72–36.95; 40.97 ± 3.55 | 35.11–26.90; 30.29 ± 3.74 |
| G. tetrahit | 32 | 44.72–38.79; 46.91 ± 2.61 | 35.11–26.90; 34.21 ± 3.88 |
| G. bifida | 32 | 44.11–40.89; 30.58 ± 2.87 | 38.95–33.00; 30.68 ± 2.89 |
Сравнение размеров пыльцы диплоидных видов (2n = 16) с размерами пыльцы тетраплоидных видов (2n = 32) показало отсутствие корреляции между плоидностью и размерами пыльцы. Для других цветковых растений отмечалось, что у диплоидных видов пыльца может быть несколько мельче, чем у тетраплоидных (Григорьева и др., 2009; Zlesak, 2009).
ОБСУЖДЕНИЕ
Сравнительное исследование морфологии пыльцевых зерен девяти видов р. Galeopsis показало, что пыльца всех видов, описанных в р. Galeopsis, крайне единообразна, значительной морфологической изменчивости внутри рода не выявлено. Сопоставление полученных данных с систематическим делением р. Galeopsis, предложенным Пужаром и Славиковой (Pouzar, Slavíková, 2000), показало, что по комплексу морфологических признаков пыльцы не удается выделить надвидовые группы, соответствующие таксономическим подразделениям, установленными по макроморфологическим признакам спорофита (Reichenbach, 1830; Briquet, 1893; Slavíková, 1963).
Скульптура поверхности пыльцы
Признаки скульптуры в роде Galeopsis чрезвычайно стабильны, виды по признакам скульптуры пыльцы отчетливо не различаются (рис. 1–3). Отсутствует индивидуальная изменчивость – все пыльцевые зерна в одном образце имеют одинаковую скульптуру; каких-либо отклонений по признакам скульптуры в образцах исследованных видов Galeopsis не выявлено. Сходную скульптуру имеют пыльцевые зерна в разных образцах одного вида и в образцах разных видов, размах изменчивости крайне невелик. Максимальная изменчивость, обнаруженная внутри одного вида (G. pyrenaica; рис. 1м, н), выглядит незначительной. В других изученных таксонах отклонения по признакам скульптуры у нормальной пыльцы также встречаются очень редко (Pozhidaev, 1995, Fig. 5, V–Z), обычно связаны с дефектами и приостановкой развития и встречаются чаще у стерильных пыльцевых зерен.
Рис. 5.
Многообразие вариантов скульптур р. Lamium и в сем. Lamiaceae, р. Euphorbia (подрод Esula) и р. Artemisia. a – Варианты скульптуры р. Lamium: 1 – L. galactophyllum; 2 – L. album; 3 – L. ovata; 4 – L. galeobdolon spp. vulgare; 5 – L. galeobdolon spp. montanum; 6 – L. galeobdolon spp. galeobdolon; 7 – L. bifidum; 8 – L. purpureum. б – Варианты ретикулятных скульптур в сем. Lamiaceae: 9 – Orthosiphon wulfenioides; 10 – Bystropogon tomentosum; 11 – Sphacele caluciana; 12 – Ziziphora bungeana; 13 – Dicerandra frutescens; 14 – Blephilia hirsute; 15 – Brittoniastrum micranthum; 16 – Plectranthus glaucocalyx; 17 – Galeopsis ladanum; 18 – Scutellaria calericulata; 19 – Dysophylla yatabeana; 20 – Physostegia virginiana; 21 – Eriope latifolia; 22 – Coleus igmaricus; 23 – Marrubium peregrinum; 24 – Sideritis montana; 25 – Tinnea eathiopica; 26 – Pseudomarrubium eremostachyoides; 27 – Geniosporum paludosum; 28 – Moshosms polystachum; 29 – Acrocephalus zambesiacus; 30 – Pogostemon velatus; 31 – Mosla grosseserrata; 32 – M. japonica; 33 – Elscholtzia densa; 34 – Collinsonia anisata; 35 – Mesona chinensis; 36 – Keiskea japonica. в – Варианты скульптуры р. Euphorbia подрода Esula: 37 – E. portlandica; 38 – E. mellifera; 39 – E. mongolica; 40 – E. broteri; 41 – E. tauriensis. г – Варианты скульптуры р. Artemisia: 42 – A. kelleri; 43 – A. leontopodioides; 44 – A. flava; 45 – A. badhysi.
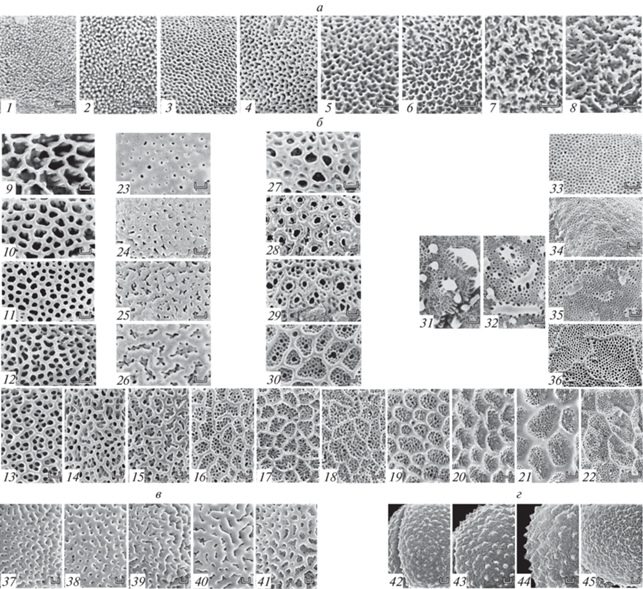
Объем таксона и размах изменчивости признаков скульптуры пыльцы
Интерес представляет сравнение полученных данных по р. Galeopsis с имеющимися данными по изменчивости признаков скульптуры в других родах сем. Lamiaceae, на уровне семейства в целом (Пожидаев, 1989) и за его пределами. Полученные данные показали, что нет однозначной зависимости между размахом изменчивости признаков скульптуры и объемом таксона (рода). Крупные и очень крупные роды по признакам скульптуры пыльцы могут быть удивительно однообразны. Так, например, в одном из крупнейших родов цветковых Euphorbia (около 2000 видов, чрезвычайно разнообразных по жизненным формам спорофитов) размах изменчивости по морфологическим признакам пыльцы крайне невелик (рис. 5в); в крупном р. Artemisia (от 200 до 400 видов; рис. 5г) виды мало различимы (Григорьева и др., 2018). Нередко средние и мелкие роды обладают не меньшей, а иногда и более широкой изменчивостью признаков скульптуры – например, сравнительно небольшой р. Lamium (40–50 видов; 1–3, 7, 8 на рис. 5а) вместе с выделенным из него р. Galeobdolon (1 вид; 4–6 на рис. 5а). Таким образом, изменчивость признаков скульптуры пыльцы видов внутри родов обычно невелика, либо отсутствует. Если изменчивость есть, варианты скульптуры обычно удается упорядочить в непрерывные ряды (рис. 5; Pozhidaev, 1995, Plate. 5, K–R).
Ранг таксона и размах изменчивости признаков скульптуры пыльцы
Заметная изменчивость по признакам скульптуры внутри сем. Lamiaceae разворачивается на уровне надродовых таксонов (Пожидаев, 1989) – в пределах подтриб и триб (в неблизко-родственных и генетически изолированных группах). На уровне всего сем. Lamiaceae варианты ретикулятных скульптур, встречающихся в разных подтаксонах, также удается выстроить в непрерывный (плавный, постепенный) и упорядоченный ряд (рис. 5а, б).
Отметим, что в случае признаков скульптуры непрерывную изменчивость (ряды, рис. 5) имеет многообразие ее типичных вариантов (индивидуальной изменчивости и отклонений признаков скульптуры в исследованных образцах не обнаружено). Все состояния признака скульптуры могут встречаться как типичные: и терминальные, хорошо различимые (простая сетка, двойная сетка), и варианты с промежуточными состояниями признака (рис. 5).
Форма пыльцевого зерна
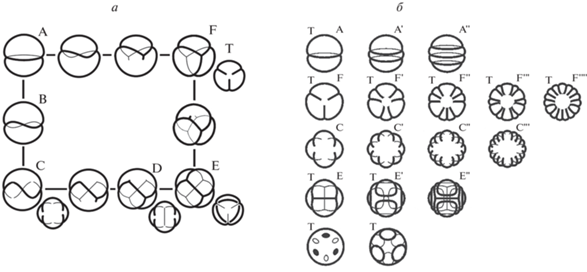
В отличие от рядов форм пыльцы, где в качестве типичных в разных таксонах встречаются наиболее симметричные формы (цикло-апертурная, 3-кольпатная и 6-руговая; рис. 6, формы А, F и E), все формы пыльцы р. Galeopsis, и типичная, и отклоняющиеся (вся индивидуальная изменчивость, даже самые редкие формы), одинаково упорядочены и в равной степени подчиняются одной и той же геометрической закономерности. Никаких других форм, кроме принадлежащих описанному ряду, в изученных образцах не обнаружено. Типичная 3-кольпатная форма (более симметричная; рис. 6, F), наравне с другими формами (менее симметричными, но не менее упорядоченными) является гармоничной и равноценной частью упорядоченного целого, никак не выделенная онтологически как отдельная сущность (родительская, предковая, первичная). Описанное многообразие форм пыльцы едино – непрерывно (все формы существуют; рис. 6а) и периодично упорядочено (рис. 6б), т.е. имеет собственную целостную и симметричную форму (Пожидаев, 2009, рис. 1).
Рис. 6.
Универсальная структура многообразия форм пыльцы: а – схематическое изображение морфологического ряда форм пыльцы; б – система параллелизмов форм пыльцы; периодическая упорядоченность гомологических и гетерологических рядов. Буквой Т обозначены формы, встречающиеся в таксонах цветковых растений как типичные; изображения меньшего размера рядом с формами С, D, E, F показывают ту же форму в другой проекции.
Это дает основания рассматривать отклонения не как патологическое состояние (дефект), имеющее собственные причины, а как вариант нормы, возникающий вследствие тех же причин, что и типичная форма. Формы пыльцы, которые действительно стоит считать уродливыми, чаще всего возникают в результате нарушений при расхождении в мейозе (Шелудякова и др., 2017, рис. VII , 25) и обычно приводят к приостановке развития.
Любопытно, что упорядоченность многообразия форм пыльцы проявляется именно при учете отклоняющихся форм с “неправильным” расположением апертур, которые нередко рассматриваются как уродства, случайные отклонения от типа, результат нарушения развития (Clarke, 1975). Их существование чаще игнорируется в морфологических и в систематических исследованиях и не учитывается при эволюционной интерпретации данных.
Объем таксона и размах изменчивости признаков формы пыльцы
Индивидуальная изменчивость формы пыльцевых зерен (расположение апертур и чередование ширины мезокольпиумов) в единичном образце (содержимое одного бутона или пыльника) может быть сопоставима по размаху с изменчивостью таксонов высших, надродовых рангов независимо от их объема. Размах изменчивости форм пыльцы не зависит от объема таксона.
Ранг таксона и размах изменчивости признаков формы пыльцы
Одна и та же изменчивость (одни и те же формы и их ряды) повторяется и в отклоняющихся формах индивидуальной изменчивости генетически сходных пыльцевых зерен одного пыльника, и в макротаксономической изменчивости типичных форм далеких, неродственных, надродовых таксонов независимо от их ранга. Других форм не бывает; изменчивость транзитивна (In Memoriam, 2007), упорядочена поверх границ генетического родства таксонов и гомологии структур (Pozhidaev, 2002). Размах изменчивости форм пыльцы не зависит от ранга таксона.
В качестве примера, подтверждающего универсальность описанного ряда форм пыльцы (рис. 6а) для групп цветковых растений, приведены фотографии одной и той же 4-кольпатной W‑формы (рис. 6а, форма С) у представителей сем. Rosaceae, Solanaceae, Fabaceae, Scrophulariaceae, Valerianaceae (рис. 4с–х). Полные наборы форм пыльцы рассматриваемого ряда были описаны ранее для многих неродственных таксонов цветковых растений (Acer – Pozhidaev, 1993; Aesculus – Pozhidaev, 1995; Krameria – Pozhidaev, 2002; Scrophularia – Шелудякова и др., 2017; Artemisia – Григорьева и др., 2018; Еuonymus – Gavrilova et al., 2018; Nicotiana – Григорьева и др., 2019; Physalis, Cestrum, Rubus, Euphorbia – готовятся к публикации). Одна и та же упорядоченная изменчивость рефреном (In Memoriam, 2007) повторяется в неродственных, далеких таксонах семенных растений.
Структура изменчивости морфологических признаков пыльцы
Изменчивость признаков скульптуры поверхности и формы расположения апертур разворачивается на разном таксономическом уровне. Признаки скульптуры поверхности пыльцы зависят от ранга (обычно не имеют индивидуальной изменчивости и изменчивы на уровне высших таксонов – родов и семейств). Хотя изменчивость формы пыльцы и вариантов чередования ширины мезокольпиумов не зависят от ранга, тем не менее, изменчивость обeих групп признаков имеет сходную структуру. Многообразие их вариантов таково, что может быть также сведено к непрерывным, рационально упорядоченным и логически завершенным рядам.
Если ряды вариантов скульптуры (рис. 5) еще можно попытаться “списать” на результат скорее логического упорядочивания и достаточно произвольных формальных построений (упорядочивание по принципу больше–меньше; распределение случайных уклонений от среднего или типичного), то в рядах форм пыльцы (рис. 4, 6a) очевидна чисто геометрическая упорядоченность, не допускающая никакой произвольности. Естественная изменчивость рассмотренных признаков пыльцы оказывается не типологичной (In Memoriam, 2007; Шаталкин, 2012; Любарский, 2018; Павлинов, 2019). Равно не удается ни мерономическое деление непрерывного многообразия признаков на части (сведение к дискретным типам) – непрерывное бесконечно делимо, непрерывность самая мощная бесконечность; ни таксономическое объединение живых тел в группы (таксоны, классы) – остаются живые тела промежуточной формы (индивиды – неделимые по условиям задачи). Мерономические отношения (часть–целое: сравнительный метод; Раутиан, 2001) оказываются сложнее и разнообразнее типологической задачи и не могут быть сведены к классификационным процедурам (дискретизация, классифицирование; Пожидаев, 2009). Типологическая редукция (сведение всего естественного многообразия к типичным формам, как к более существенным, таксоноспецифическим, диагностическим, предковым) необратимо искажает естественную структуру упорядоченности, наблюдаемую при другом подходе, и приводит к потере части наблюдаемого многообразия (переходные формы; Пожидаев, 2009, 2015).
Возможно, разница в таксономическом уровне разворачивания многообразий разных признаков может быть связана со временем их закладки в развитии пыльцевого зерна. Форма пыльцы (расположение апертур) детерминируется в раннем пост-мейозе, сразу после завершения цитокинеза и возникновения клеточной стенки разделившихся микроспороцитов – это одно из первых событий индивидуального развития мужского гаметофита (возникновение, детерминация дефинитивной формы). В форме зрелой пыльцы могут фиксироваться результаты самых ранних событий поляризации и конформации цитоплазмы в раннем пост-мейозе (Pozhidaev, 2000b, Fig. 72 ; Пожидаев, 2001, табл. II ). Скульптура поверхности (эктекзина) формируется на более поздних стадиях, в ранней тетраде и в течение всего тетрадного периода, и ее изменчивость может быть определена более жестко.
ТЕОРЕТИЧЕСКОЕ ЗАКЛЮЧЕНИЕ
Проведенное исследование морфологических признаков пыльцы Galeopsis, учитывая биологические особенности пыльцы, которые делают ее почти идеальным модельным объектом для исследования свойств изменчивости, приводит к некоторым теоретическим заключениям.
На примере конкретного таксона показано, что изменчивость по исследованным морфологическим признакам пыльцы отсутствует и на внутривидовом уровне (особи одного вида, разновидности, панмиктические популяции), и на внутриродовом уровне (генетически изолированные, но близкородственные виды одного рода) – т.е. на уровне тех таксономических рангов, где возникновение изменчивости могло бы быть объяснено действием механизмов филетической эволюции (дивергенция, отбор, вымирание, наследование, родство).
Широкая изменчивость признаков формы пыльцы и скульптуры поверхности обнаруживается либо в генетически сходном поколении (т.е. не объясняется наследованием), либо на уровне более крупных таксонов, где мы теряем критерии и не можем достоверно различать унаследованное сходство от случаев параллелизмов (независимое сходство, для которого гипотеза родства не подтверждается) – не можем объяснить (фило-)генетически, разделить на монофилетические дискретные группы, построить генеалогическое древо, схему дивергенции.
Многообразие современных форм непрерывно – промежуточные формы признаков (и скульптур, и расположения апертур) не вымерли, все формы существуют одновременно у современных растений в полном ассортименте (пусть даже некоторые из них редки и никогда не бывают типичными, т.е. не специфичны ни для одного отдельного таксона). Изменчивость не дивергентна – нет следов расхождения, дивергенции признаков за счет селективного вымирания промежуточных форм, как механизма возникновения изменчивости. Изменчивости либо нет вовсе, либо везде, где она есть (индивидуальная изменчивость или изменчивость на уровне неблизкородственных родов), многообразие наблюдаемых форм укладывается не в дискретно-иерархические клады (дивергенция, дискретные формы, генеалогический порядок следования от предка к потомку), а в непрерывные клины (упорядоченные и ненаправленные ряды). Проблема и интерес обсуждаемых данных по изменчивости признаков пыльцы р. Galeopsis не в том, что на внутривидовом и внутриродовом уровне ничего не вымерло (нет дивергенции признаков), а в том, что ничего не возникло (нет изменчивости). А там, где изменчивость есть – нет дивергенции.
Морфоз – эволюция каждый день
На пыльце, как на модельном объекте, можно наблюдать как в результате индивидуального развития в полном и генетически сходном поколении отдельной генеалогической линии предков и потомков происходит не наследование родового сходства, индивидуальной специфичной формы (предковой, типичной, нормальной или отклоняющейся), не случайное мутирование и не расхождение признаков (дивергенция) за счет вымирания промежуточных форм в процессе отбора адаптаций (Дарвин, 1939). Выглядит, будто в ходе развития пыльцы механизмы рекомбинации признаков в мейозе могут привести к детерминации форм, типичных для других таксонов. Происходит не наследование типичной формы, а воспроизведение фрагментов непрерывной и упорядоченной изменчивости (типичные формы других таксонов, одинаковые отклонения и ряды форм, и многообразие в целом). Происходит не генезис (понятый ли как творение или как порождение и развитие “по роду их”), а морфоз – перерождение формы в рамках закономерности, изменение схемы общей организации живого тела (макротаксономическая изменчивость, возникновение признаков формы другого рода, без изменения видовых – по: Создатели…, 2012; массовая перечеканка формы, эволюция на основе закономерностей – по: Берг, 1922).
Многообразие форм пыльцы в одном образце (пыльнике), генетическая изменчивость которого исчерпывается результатами рекомбинации родительских генов спорофита, оказывается сопоставимым с изменчивостью форм пыльцы высших (родовых и надродовых) таксонов – далеких, заведомо неродственных и генетически несовместимых. Полное морфологическое сходство форм пыльцы, таксономическая (генетическая) неспецифичность их признаков может быть показана для таксонов на уровне классов цветковых и отделов семенных растений (Pozhidaev, 2000a; Шелудякова и др., 2017).
На индивидуальном уровне (содержимое одного пыльника) наблюдается макропотенциальная изменчивость – возникновение типичных (и отклоняющихся) признаков другого таксона (другого архетипа) – изменение признаков рода (макротаксономическая изменчивость). Индивидуальное развитие имеет альтернативы, многообразие которых подчинено общим (геометрическим и транзитивным) закономерностям. На примере полного поколения пыльцы удается наблюдать, как признак другого типа организации схемы живого тела (макротаксономический признак) может возникнуть сразу, за одно поколение, скачком. Индивидуальная форма воспроизводится каждый раз заново, из ничего, с нуля, следуя собственным закономерностям (номогенез; Берг, 1922). Отклонения (промежуточные формы) не бывают типичными (не характеризуют никакого таксона), всегда редки, никогда не вымирали (встречаются у современных растений в полном ассортименте).
В полном поколении пыльцы наблюдается возникновение массовой и политопной параллельной макропотенциальной изменчивости без гипотетических макромутаций, чрезвычайно редких, уникальных эволюционных событий с эпохальными последствиями (возникновение нового макротаксона, специфичной формы, нового типа организации, плана строения живого тела).
Логическая неопределенность дискретной формы в непрерывном многообразии может служить обоснованием как возможности перерождения индивидуальной формы (текучесть формы, метаморфоз, по: Гете, 2014) при ее возникновении (размножении), так и способность такого многообразия к “автоматическому” самовоспроизведению (не только численности поголовья, но и самой изменчивости), без каких-либо дополнительных причин, с точностью, феноменальной для биологических объектов (упорядочена вся индивидуальная изменчивость, до долей процента), и независимо от обстоятельств персональной истории (родство, борьба за существование). Непрерывное многообразие меняется само, в силу собственных свойств, и не нуждается в дополнительной причинности для объяснения возникновения параллелизмов и упорядоченности (наблюдаемой изменчивости). Единый принцип может объяснить и самовоспроизведение (устойчивость), и изменчивость (эволюционирование).
Нетипологическая концепция морфологического многообразия форм пыльцы. Морфопластика
Естественную систему морфологической изменчивости образуют не таксоны (монофилетические группы родства; отдельные сущности, дискретные классы, типы организации, общие понятия, результаты умозрения – универсалии), а формы индивидуальных живых тел (единичные предметы, доступные наблюдению – натуралии). Типичные формы таксонов входят в эту систему, составляя ее малую часть (рис. 1а, б).
В силу единой упорядоченности ряда невозможно в сходстве одних форм (типичных) видеть родство, и объяснять их возникновение отбором приспособлений и наследованием (селективно и генеалогически), а другие (отклоняющиеся) формы того же ряда рассматривать как результат некоей морфологической закономерности или случайность.
Естественное многообразие форм пыльцы описывается системой параллелизмов – непрерывных, упорядоченных и ненаправленных рядов (клина): не дивергентных и не генеалогических последовательностей (не клада). События произошедшей морфологической эволюции привели к совсем иным наблюдаемым последствиям, нежели те свойства естественной изменчивости, которые прогнозирует теория (дивергенция). Современное многообразие оказывается непрерывным (и ненаправленным) рядом, а последовательность форм в череде предков–потомков (генеалогической линии) оказывается прерывной, дискретной (не клина, так как допускает морфоз, скачкообразное перерождение формы, возникновение типичной формы иного рода, таксона).
Ряды отражают не динамический аспект формы (морфодинамика – пути преобразований, направление трансформации и ее необратимость, эволюционные связи типов, последовательность во времени, траектория, первичный–вторичный, примитивный–продвинутый, анцестральный–адвентивный; In Memoriam, 2007), а ее пластический аспект (морфопластика – потенциал, эволюционные возможности морфологической структуры); закономерности как собственный, геометрический аспект формы – свойства пустого вмещающего пространства (свойством непрерывности обладают форма, пустота и время).
Представление о смене типичных форм во времени (эволюция), как о непрерывном, направленном и необратимом трансформационном ряде (семогенез), оказывается наивным (упрощенным, вследствие исходной редукции данных в ходе их типологизации, классифицирования). Свойства наблюдаемой индивидуальной изменчивости оказываются иными до противоположности (не дискретная, а непрерывная; не случайная, а закономерная; не таксон-специфичная, а транзитивная; не иерархичная, а рангово-независимая).
Многообразие современных форм пыльцы описывается как надорганизменная (над схемой тела) целостная (непрерывная и упорядоченная) система параллелизмов, имеющая собственную симметричную и периодическую форму. В этой системе параллелизмов гомологические и гетерологические ряды составляют единое целое (рис. 1б), в котором гомологии не могут быть оторваны от гетерологий и объяснены отдельно, например родством; ряды обладают собственной симметрией и выделенного направления (предок–потомок, первичный–вторичный, примитивный–продвинутый) не имеют.
ВЫВОДЫ
На примере девяти видов р. Galeopsis и других исследованных таксонов показано следующее.
1. Отсутствует изменчивость, а значит и само явление дивергенции по основным признакам пыльцы там, где она должна возникать согласно теории филогенеза и представлениям классической генетики (теоретическое ожидание): в панмиктических популяциях одного вида; между близкими видами; между видами одного рода, связанных кровным родством (род, как группа родства, монофилетическая клада).
2. Макротаксономическая изменчивость признаков пыльцы наблюдается либо на уровне с минимальной генетической изменчивостью (пыльца одного пыльника, результат рекомбинации генов одного спорофита), либо на уровне далеких, неродственных групп. Там, где возникновение изменчивости не может быть обосновано представлениями о дивергенции, механизмом естественного отбора и наследованием случайных (неупорядоченных, неопределенных и ненаправленных) мутационных изменений (а другого теоретического обоснования не имеет). Там, где должны наблюдаться следы дивергенции – нет изменчивости; а там, где изменчивость есть – она не дивергентна.
3. Наблюдаемая полная индивидуальная изменчивость основных признаков пыльцы (форма пыльцы, скульптура поверхности, чередования ширины мезокольпиумов), описанная на примере полного поколения пыльцы девяти видов р. Galeopsis, имеет структуру рядов – непрерывных (делимых только на дискретные живые тела и не делимые без остатка ни на отдельные группы живых тел, ни на дискретные формы признаков) и упорядоченных транзитивно (таксон-неспецифично и рангово-независимо). Одинаковая структура изменчивости (непрерывный и упорядоченный ряд) наблюдается у разных признаков и независимо от ранга рассматриваемых групп (так как во всех случаях рассматривается одна и та же индивидуальная изменчивость).
4. В структуре изменчивости морфологических признаков пыльцы изученных таксонов не наблюдается ни следов вымирания промежуточных вариантов (наблюдаемые отклоняющиеся формы), ни случайной и неопределенной изменчивости. Существуют все формы – ряды форм непрерывны. Отклонения – формы промежуточные между типичными – теряются при типологической редукции (в процедурах классифицирования); не наследуются, не бывают типичными, всегда редки, никогда не вымирали. Эволюционная новизна (современное состояние) оказывается всегда упорядочена.
Список литературы
Берг Л.С., 1922. Номогенез или эволюция на основе закономерностей. Петроград: Гос. изд-во. 306 с.
Гете И.В., 2014. Научные сочинения. Т. 1. Образование и преобразование органических существ (морфология). М.: КМК. 696 с.
Гладкова В.Н., 1978. Род Пикульник – Galeopsis L. // Флора Европейской части СССР. Т. 3. Л.: Наука. С. 157–160.
Григорьева В.В., Коробков А.А., Токарев П.И., 2009. Палиноморфология рода Artemisia L. (Asteraceae) // Бот. журн. Т. 94. № 3. С. 328–351.
Григорьева В.В., Брицкий Д.А., Коробков А.А., 2018. Морфология пыльцевых зерен видов рода Artemisia (Asteraceae) Дальнего Востока России // Бот. журн. Т. 103. № 10. С. 1255–1264. https://doi.org/10.7868/S0006813618100046
Григорьева В.В., Пожидаев А.Е., Семенов А.Н., Брицкий Д.А., 2019. Морфологическая изменчивость пыльцы представителей рода Nicotiana (Solanaceae) // Бот. журн. Т. 104. № 6. С. 900–917. https://doi.org/10.1134/S0006813619060061
Дарвин Ч., 1939. Сочинения. Т. 3. М.; Л.: Изд-во АН СССР. С. 253–667.
Куприянова Л.А., Алешина Л.А., 1967. Палинологическая терминология покрытосеменных растений. Л.: Наука. 84 с.
Любарский Г.Ю., 2018. Происхождение иерархии: история таксономического ранга. М.: КМК. 659 с.
Павлинов И.Я., 2019. Биологическая систематика: в поисках естественной системы. М.: КМК. 246 с.
Пожидаев A.E., 1989. Структура экзины пыльцевых зерен представителей сем. Lamiaceae // Бот. журн. Т. 74. № 10. С. 1410–1422.
Пожидаев А.Е., 2001. Естественный полиморфизм в расположении апертур пыльцы цветковых растений // Мат-лы I Междунар. Семинара “Пыльца как индикатор состояния окружающей среды и палеогеографические реконструкции” / Отв. ред. Белонин М.Д., Киричкова А.И. СПб.: ВНИГРИ. С. 141–144.
Пожидаев А.Е., 2009. Структура многообразия морфологического признака на примере расположения апертур пыльцы цветковых и естественная упорядоченность биологического многообразия. Или – что такое многообразие (описание и интерпретация) // Вид и видообразование. Анализ новых взглядов и тенденций. Тр. ЗИН РАН. Приложение № 1. М: Т-во науч. изд. КМК. С. 151–182.
Пожидаев А.Е., 2015. Рефренная структура биологического многообразия и теория филогенеза // Палеоботанический временник. Вып. 2. М.: ГЕОС. С. 115–127.
Раутиан А.С., 2001. Апология сравнительного метода: о природе типологического знания // Гомологии в ботанике: опыт и рефлексия. Тр. IX школы по теоретической морфологии растений “Типы сходства и принципы гомологизации в морфологии растений” (Санкт-Петербург, 31 января – 3 февраля 2001 г.) / Ред. Оскольский А.А., Соколов Д.Д., Тимонин А.К. СПб.: Санкт-Петербургский союз ученых. С. 73–80.
Сладков А.Н., 1967. Введение в спорово-пыльцевой анализ. М.: Наука. 275 с.
Создатели современного эволюционного синтеза: коллективная монография, 2012 / Отв. ред.-сост. Колчинский Э.И. СПб.: Нестор-История. 996 с.
Тюнникова Н.В., 2006. Об объеме рода Galeopsis L. (Lamiaceae) // Бот. журн. Т. 91. № 2. С. 290–296.
Шаталкин А.И., 2012. Таксономия. Основания, принципы и правила. М.: Т-во науч. изд. КМК. 600 с.
Шелудякова М.Б., Григорьева В.В., Пожидаев А.Е., 2017. Морфология пыльцевых зерен представителей рода Scrophularia (Scrophulariaceae) // Бот. журн. Т. 102. № 3. С. 361–379.
Юзепчук С.В., 1954. Род Пикульник – Galeopsis L. // Флора СССР. Т. 21. М.; Л.: АН СССР. С. 111–124.
Abu-Asab M.S., Cantino P.D., 1994. Systematic implications of pollen morphology in subfamilies Lamioideae and Pogostemonoideae (Labiatae) // Ann. Miss. Bot. Gard. V. 81. № 4. P. 653–686.
Bendiksby M., Thorbek L., Scheen A.-C., Lindqvist C., Ryding O., 2011a. An updated phylogeny and classification of Lamiaceae subfamily Lamioideae // Taxon. V. 60. № 2. P. 471–484. https://doi.org/10.1002/tax.602015
Bendiksby M., Tribsch A., Borgen L., Trávníček P., Brysting A., 2011b. Allopolyploid origins of the Galeopsis tetraploids – revisiting Müntzing’s classical textbook example using molecular tools // New Phytol. V. 191. № 4. P. 1150–1167. https://doi.org/10.1111/j.1469-8137.2011.03753.x
Blackmore S., Crane P.R., 1998. The evolution of apertures in the spores and pollen grains of embryophytes // Reproductive Biology / Eds Owens S.J., Rudall P.J. Kew: Royal Botanic Gardens. P. 159–218.
Briquet J., 1893. Monographie du genre Galeopsis. Bruxelles: Kessinger Publishing, LLC. 319 p.
Campo M., van, 1976. Patterns of pollen morphological variation within taxa // The Evolutionary Significance of the Exine / Eds Ferguson I.K., Muller J. L.: Academic Press. P. 125–137.
Clarke G.C.S., 1975. Irregular pollen grains in some Hypericum species // Grana. V. 15. P. 117–125.
Erdtman G., 1952. Pollen Morphology and Taxonomy. Angiosperms. Stockholm: Almquist and Wiksell. 539 p.
Gavrilova O., Britski D., Grigorieva V., Tarasevich V., Pozhidaev A., Leunova V., 2018. Pollen morphology of the genus Euonymus (Celastraceae) // Turczaninowia. V. 21. № 4. P. 188–206. https://doi.org/10.14258/turczaninowia.21.4.20
Gray S.F., 1821. A Natural Arrangement of British Plants. V. 2. L.: Baldwin, Cradock, and Joy. 760 p.
Harley R.M., Atkins S., Budantsev A.L., Cantino P.D., Conn B.J., et al., 2004. Labiatae // The Families and Genera of Vascular Plants. V. 6 / Ed. Kadereit J.W. Berlin: Springer-Verlag. P. 167–275.
Huynh K., 1972. Le pollen et la systematique du genere Sideritis L. (Labiatae) // Bull. Mus. Nat. Hist. Nat. 3 ser. № 45. Bot. 1. P. 1–28.
In Memoriam. С.В. Мейен: палеоботаник, эволюционист, мыслитель, 2007. М.: ГЕОС. 348 с.
Mohl H., 1834. Ueber den Bau und die Formen der Polenkörner. Bern.: Chr. Fisher u. Comp. 130 p.
Müntzing A., 1928. Chromosome number, nuclear volume and pollen grain size in Galeopsis // Hereditas. V. 10. № 3. P. 241–260. https://doi.org/10.1111/j.1601-5223.1928.tb02473.x
Olmstead R., 2016. F synoptical classification of the Lamiaceae, version 2.6.2. http://depts.washington.edu/phylo/Classification.pdf
Pouzar Z., Slavíková Z., 2000. The problem of correct name for Ladanum (Lamiaceae) // Čas. Nár. Muz. Řada Přir. V. 169. № 1–4. P. 41–43.
Pozhidaev A.E., 1992. The origin of three- and sixcolpate pollen grains in the Lamiaceae // Grana. V. 31. P. 49–52.
Pozhidaev A.E., 1993. Polymorphism of pollen in the genus Acer (Aceraceae). Isomorphism of deviant forms of Angiosperm pollen // Grana. V. 32. № 1. P. 79–85. https://doi.org/10.1080/00173139509429028
Pozhidaev A.E., 1995. Pollen morphology of the genus Aesculus (Hippocastanaceae). Patterns in the variety of morphological characteristics // Grana. V. 34. № 1. P. 10–20. https://doi.org/10.1080/00173139509429028
Pozhidaev A.E., 1998. Hypothetical way of pollen aperture patterning. 1. Formation of 3-colpate patterns and endoaperture geometry // Rev. Palaeobot. Palynol. V. 104. № 1. P. 67–83.
Pozhidaev A.E., 2000a. Hypothetical way of pollen aperture patterning. 2. Formation of polycolpate patterns and pseudoaperture geometry // Rev. Palaeobot. Palynol. V. 109. № 3–4. P. 235–254. https://doi.org/10.1016/s0034-6667(99)00057-3
Pozhidaev A.E., 2000b. Pollen variety and aperture patterning // Pollen and Spores: Morphology and Biology / Eds Harley M.M., Morton C.M., Blackmore S. Kew: Royal Botanic Gardens. P. 205–225.
Pozhidaev A.E., 2002. Hypothetical way of pollen aperture patterning. 3. A family-based study of Krameriaceae // Rev. Palaeobot. Palynol. V. 127. № 1–2. P. 1–23. https://doi.org/10.1016/S0034-6667(02)00251-8
Punt W., Blackmore S., Nilsson S., Le Tomas A., 1994. Glossary of Pollen and Spore Terminology. LPP Contributions Series № 1. Utrecht: LPP Foundation. 71 p.
Reichenbach L., 1830. Flora Germanica Excursoria. Lipsiae: Carolum Cnobloch. 878 p.
Slavíková Z., 1963. Bemerkungen zur taxonomie der gattung Galeopsis L. // Novit. Botan. Horti Bot. Univ. Carolinae Pragensis. P. 39–43.
Townsend C.C., 1972. Gen. Galeopsis L. // Flora Europaea. V. 3. Cambridge: Cambridge Univ. Press. P. 145–147.
Walker J.W., Doyle J.A., 1975. The bases of angeosperms phylogeny: Palynology // Ann. Miss. Bot. Gard. V. 62. № 3. P. 664–723. https://doi.org/10.2307/2395271
Wunderlich R., 1967. Ein Vorschlag zu einer natürlichen Gliederung der Labiaten auf Grund der Pollenkörner, des Samenentwicklung und des reifen Samens // Oesterr. Bot. Z. Bd. 114. № 4–5. S. 383–483.
Zhao F., Chen Y.P., Salmaki Y., Drew B.T., Wilson T.C. et al., 2021. An updated tribal classification of Lamiaceae based on plastome phylogenomics // BMC Biol. V. 19. № 2. https://doi.org/10.1186/s12915-020-00931-z
Zlesak D.C., 2009. Pollen diameter and guard cell length as predictors of ploidy in diverse Rose cultivars, species, and breeding lines // Floricult. Ornam. Biotechnol. V. 3. P. 53–70.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии