

Журнал общей биологии, 2021, T. 82, № 5, стр. 368-381
Биоморфология в таксономии растений, на примере кавказских Minuartia s.l. (Caryophyllaceae)
С. Г. Зайченко 1, *, А. С. Зернов 1, **
1 Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, стр. 12, Россия
* E-mail: zaychenko_so@mail.ru
** E-mail: zernov72@yandex.ru
Поступила в редакцию 18.02.2021
После доработки 20.04.2021
Принята к публикации 26.04.2021
Аннотация
Систематика растений требует комплексного подхода к выполнению поставленных задач. Преобладание молекулярно-генетических методов в систематике в настоящее время не отменяет необходимости поиска новых таксономически значимых признаков. Источником принципиально новых признаков у растений может служить биоморфология. Исследование посвящено изучению возможностей применения биоморфологических подходов в систематике и филогении растений. На примере рода Minuartia s.l. (Caryophyllaceae), который является полифилетической группой, рассмотрены основные особенности структурной организации высокогорных растений применительно к определению их таксономического статуса и возможных способов возникновения биоморф в процессе эволюции. Для анализа побегообразования растений выбран один из современных подходов – модульная организация. Понятие “универсальный модуль” использовано для сравнения участков побеговых систем у разных видов. Материал для исследования собран на Северном Кавказе в 2016–2019 гг. Модельные виды для исследования отбирались по принципу систематического положения внутри рода, а также наличию существенных экологических различий представителей и, как следствие, отличающимся габитусом. Часть видов являются высокогорными растениями и распределены по скалам от среднелесного до альпийского пояса. Выбранные для исследования представители рода обладают разной степенью выраженности розеточности: полурозеточные моноподиальные оси с боковыми генеративными структурами (M. imbricata), полурозеточные оси с вегетативно-генеративными побегами (M. aizoides) и розеточные оси с дициклическими вегетативно-генеративными побегами (M. circassica); универсальным модулем у вышеперечисленных видов является полициклические поликарпические оси. Еще два вида – травы с симподиальным нарастанием, но уже полурозеточные (M. hirsuta subsp. oreina) и длиннопобеговые (M. setacea) с универсальными модулями – типичными монокарпическими побегами. Особое место занимает однолетник M. glomerata со скал среднелесного пояса. По результатам проведенного исследования выявлены три различные стратегии роста у модельных видов, а также описаны разные тенденции моноподиально и симподиально нарастающих трав. Благодаря выявлению универсальных модулей и других биоморфологических характеристик модельных видов, появляется возможность использовать в дальнейшем полученные данные в таксономических целях.
Главное, на что обращает внимание человек при взгляде на живой объект, – это его внешний облик. Самые первые таксономические системы основывались именно на чертах внешнего сходства объектов. Позже системы усложнялись и дополнялись другими признаками, присущими организмам, однако морфологические характеристики всегда оставались в лидирующей роли на пути становления таксономии до последнего времени. С возникновением молекулярно-генетических методов в научном сообществе произошел некоторый “переворот” в понимании таксономии. Морфологические признаки стали менее значимыми в филогенетических системах, элементарным признаком в которых служит нуклеотид (Schuh, Brower, 2009). Несмотря на ряд противоречий между морфологическим и филогенетическим подходами в систематике, о чем написано немало работ (Judd et al., 1999; Soltis et al., 2005; Лухтанов, Кузнецова, 2009), хотелось бы отметить, что систематика как наука является лишь упрощенной и неполной моделью, неким частичным отражением, созданным человеком, реально существующего биологического разнообразия (Павлинов, 2001). Так или иначе, таксономия в ее современном понимании строится на эволюционном учении и непременно должна отражать суть эволюционных преобразований (Hennig, 1966). Если принять во внимание, что реально существующие живые системы, скорее всего, имеют значительно более сложные взаимосвязи внутри себя, нежели мы себе их представляем, то отвергать какие-либо методы в пути познания живых систем нельзя. Систематика едина, и использование разных подходов для ее усовершенствования крайне эффективно (Лухтанов, Кузнецова, 2009). Одним из таких подходов может служить биоморфологический анализ. В настоящее время выделяется несколько направлений биоморфологических исследований: структурное, онтогенетическое, экологическое, географическое, эволюционное (Савиных, Черемушкина, 2015). К сожалению, биоморфологические характеристики растений не обрели должной популярности среди большинства зарубежных исследователей и не используются в качестве таксономически значимых. Чаще всего биоморфологические характеристики растений рассматриваются в экологическом или эволюционном ключе (Серебрякова, 1977; Барыкина, 1999; Цвелев, 2005; Савиных, 2019) и редко с точки зрения “помощницы” систематики. Однако анализ биоморфологических данных может служить мощным комплементарным методом к филогенетической системе. Филогенетика в настоящее время позволяет успешно выявлять родственные связи между организмами и реконструировать пути эволюционного развития (Лухтанов, 2013), но не указывает непосредственно на приспособления, которые организмы получили в результате адаптации к условиям внешней среды. Иными словами, филогенетика не дает прямого ответа на вопрос о том, какие структуры вырабатывают организмы в процессе эволюции. При этом современная филогенетика строится на кладистических принципах, которые диктуют непременное требование – включение в монофилетическую группу, выделяемую на основе апоморфных признаков, всех потомков одного общего предка. Зачастую подобное требование к вновь образованным таксонам трудноисполнимо применительно к морфологическим признакам тех групп организмов, которые ранее относились к одному таксону. Примером подобной ситуации служит р. Minuartia L. из сем. Caryoрhyllaceae Juss. После публикации в 2014 г. филогенетического исследования рода (Dillenberger, Kadereit, 2014) было установлено, что Minuartia s.l. является полифилетической (в рамках кладистического подхода) группой. Отсюда возникает вопрос о поиске морфологических признаков для разделения этой полифилетической группы.
Выбор объектов для исследования был сделан в пользу кавказских представителей р. Minuartia. Интерес к этим растениям вызван существенными экологическими различиями представителей и, как следствие, отличающимся габитусом, а также их систематическим положением внутри рода.
На сегодняшний день существует несколько монографических работ, посвященных систематике и географии рода (Mattfeld, 1922; McNeill, 1962, 1963), несколько работ было посвящено описанию кариотипов (Çelebioğlu, Favarger, 1982; Çelebioğlu et al., 1983; Favarger, Recoder, 1987; Galland, 1988), а также карпологии рода, в частности исследованию структуры семян (Кожанчиков, 1975; Yildiz, 2002; Minuto et al., 2006, 2011; Amini et al., 2011; Zaychenko, Zernov, 2017). Из перечисленных наиболее важных опубликованных ранее работ, касающихся р. Minuartia, нет ни одной работы, посвященной биоморфологии его представителей.
МАТЕРИАЛЫ И МЕТОДЫ
Род Minuartia s.l. включает в себя около 175 видов (McNeill, 1962; Rechinger, 1988; Halliday, 1993; Kamari, 1997; Rabeler et al., 2005). Ареал представителей в основном приурочен к аркто-альпийской зоне Северного полушария: в Европе, прежде всего в Средиземноморье, и в Кавказско-Малоазиатском (ирано-турецком) регионе. На территории Кавказа род представлен 22 видами (Лазьков, 2012).
Модельные виды для изучения биоморфологических характеристик и моделей побегообразования были отобраны по следующим критериям: 1) положение вида в системе рода, 2) экологические особенности представителей, 3) доступность репрезентативного материала. На основании этих критериев в качестве модельных были выбраны шесть видов, обитающих на Кавказе, представленные в табл. 1 (Лазьков, 2012).
Таблица 1.
Модельные виды Minuartia s.l. и их характеристика
| Номенклатура по системе McNeil, 1962 | Принадлежность к секциям по системе McNeil, 1962 | Номенклатура по системе Dillenberger & Kadereit, 2014 | Условия обитания на Кавказе |
|---|---|---|---|
| M. imbricata (Bieb.) Woronow | Spectabiles (Fezl) Hayek | Pseudocherleria imbricata (Bieb.) Dillenb. & Kadereit | Скалы, ледниковые морены, мелкокаменистые осыпи и россыпи альпийского пояса, 2500 м над ур. м. и более |
| M. aizoides (Boiss.) Borm. | Spectabiles (Fezl) Hayek | Pseudocherleria aizoides (Boiss.) Dillenb. & Kadereit | Скалы, мелкокаменистые осыпи и россыпи субальпийского и альпийского поясов, 2000 м над ур. м. и более |
| M. circassica (Albov) Woronow | Spectabiles (Fezl) Hayek | Cherleria circassica (Albov) A.J. Moore & Dillenb. | Скалы, мелкокаменистые осыпи и россыпи субальпийского и альпийского поясов, 2000 м над ур. м. и более |
| M. oreina (Mattf.) Schischk. | Plurinerviae McNeill | Minuartia hirsuta Hand.-Mazz. subsp. oreina Mattf. | Скалы субальпийского, верхне- и среднелесного поясов, высоты 1500–2500 м над ур. м. |
| M. setacea (Thuill.) Hayek | Minuartia | Minuartia setacea (Thuill.) Hayek | Скалы среднелесного пояса, высоты не более 1500 м над ур. м. |
| M. glomerata (Bieb.) Degen. | Minuartia | Minuartia glomerata (Bieb.) Degen. | Скалы среднелесного пояса, высоты не более 1500 м над ур. м. |
Для анализа биоморфологии модельных видов проводили натурные наблюдения, использовали гербарные образцы собственных сборов, а также проанализировали несколько сотен гербарных листов из фондов Московского государственного университета (MW), Гербария Главного ботанического сада им. Н.В. Цицина (MHA). Полевой материал был собран на территории Тебердинского государственного природного биосферного заповедника в августе 2015 и 2016 гг. и в ущельях Цей и Фиагдон Северо-Осетинской Республики в августе 2017 и 2019 гг.
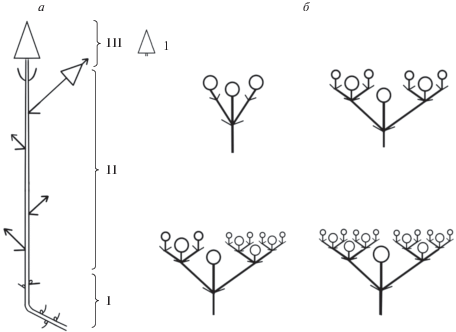
При описании биоморфологии модельных видов в основном использовали сравнительно-морфологический и структурно-функциональный методы. Для описания жизненных форм взрослых растений использовали принятую в отечественной биоморфологии терминологию (Современные подходы…, 2008; Актуальные проблемы…, 2012). Растения анализировали с позиции моделей побегообразования (Серебрякова, 1977, 1981, 1987), модульной организации (Шафранова, 1980, 1981, 1993; Бигон и др., 1989; Шафранова, Гатцук, 1994; Гатцук, 1994, 1995; Савиных, 2008). Для удобства сравнения модельных видов использовали терминологию Л.Е. Гатцук и Н.П. Савиных (Гатцук, 1994, 1995; Савиных, 2008), в частности понятие об универсальном модуле как об одноосном побеге, образованном в результате деятельности одной апикальной меристемы. Изучали ритм сезонного развития по методике И.Г. Серебрякова (1954, 1964). Соцветия были охарактеризованы по Паркину (Parkin, 1914) и Троллю (Troll, 1964). Растения схематично зарисовывали, данные по строению побегов фиксировали на схемах.
РЕЗУЛЬТАТЫ
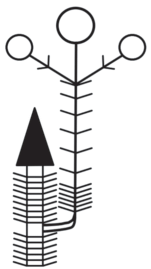
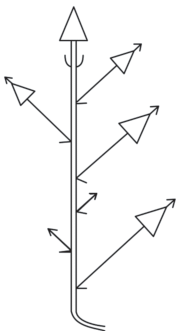
Модель побегообразования Pseudocherleria imbricata (M. imbricata) – моноподиальная полурозеточная (подобная модель побегообразования у высокогорных трав была описана в работе Н.П. Савиных (2006)). Растение заселяет территорию при помощи моноподиального нарастания (рис. 1). Разрастание осуществляется благодаря развитию пазушных побегов из почек без состояния покоя (силлепсис). Растение образует густую дерновинку, которая закреплена в почве за счет хорошо развитого главного корня, достигающего в длину у крупных особей нескольких десятков сантиметров. Придаточных корней на надземной системе анизотропных побегов практически не образуется. Цветки обычно одиночные. Используя подходы к описанию строения побегов Тролля (Troll, 1964), с дополнениями Н.П. Савиных (2006), у побегов P. imbricata можно выделить следующие зоны: 1) вегетативную (вегетативный участок побега); 2) вегетативно-генеративную (от метамера с первым до метамера с последним пазушным цветком или соцветием); 3) зону вторичного вегетативного нарастания – вегетативный участок побега, сформированный после заложения последнего пазушного цветка или соцветия.
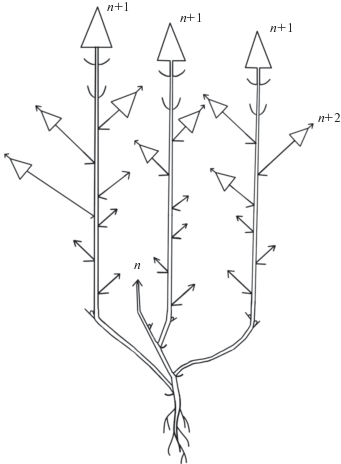
Рис. 1.
Схема строения побегов P. imbricata. 1 – цветок; 2 – вегетативный облиственный побег; 3 – укороченный побег; 4 – лист; 5 – плод; 6 – многолетняя часть побега; 7 – граница годового прироста; n, n + 1 – порядки ветвления осей.
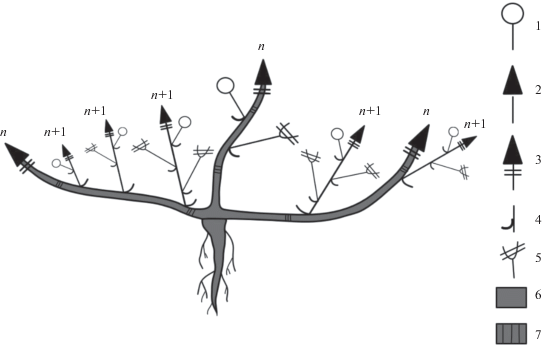
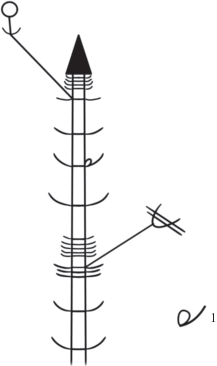
Развитие генеративных органов происходит одновременно с ростом побегов. Это возможно за счет того, что поликарпические побеги начинают свое развитие и растут вертикально до тех пор, пока не произойдет заложение одиночного цветка, затем побег продолжает вертикальный рост, однако формирует до начала плодоношения только короткие междоузлия. Тем временем цветоножка удлиняется, вынося цветок над поверхностью всей дерновинки, и, таким образом, пазушный цветок занимает псевдотерминальное положение на оси побега. После формирования коробочки побег снова начинает рост с длинными междоузлиями до образования нового цветка. Таким образом, можно говорить о том, что универсальным модулем (Савиных, 2008), т.е. тем, что формируется в течение моноподиального нарастания побега P. imbricata, являются полициклические поликарпические моноподиальные оси с редуцированными до одного цветка боковыми генеративными структурами (рис. 2).
У взрослого растения можно наблюдать очень активное ветвление за счет развития пазушных побегов, которые со временем входят в состав многолетней части дерновинки (рис. 1). В конце вегетационного периода многочисленные побеги, которые входили в вегетативно-генеративную зону, полегают. После зимовки, весной следующего года, входят в состав вегетативной зоны растения, а из почек возобновления на этих побегах начинают свое развитие побеги возобновления. Таким образом, каждый побег прошлого года входит в многолетнюю часть всего растения. Это позволяет максимально эффективно осваивать пространство.
P. aizoides (M. aizoides) имеет моноподиальную полурозеточную модель побегообразования. Захват пространства осуществляется путем роста многолетних горизонтально растущих побегов с развивающимися на них многочисленными розеточными побегами. Розеточные побеги многолетние, на протяжении нескольких лет сохраняющие отмершие прошлогодние листья. В начале вегетационного периода из прошлогодних почек на многолетних побегах начинают развиваться плагиотропные побеги разрастания с относительно длинными междоузлиями (рис. 3). Получившиеся побеги продолжают моноподиально нарастать, и одновременно в пазухах листьев закладываются почки, которые без периода покоя дают начало розеточным побегам (рис. 4). Такие побеги имеют короткие междоузлия и в фазе вегетативного ассимилирующего побега могут провести от одного года до нескольких лет, но в любом случае такие побеги никогда не переходят к цветению в первый вегетационный период после начала развития, что характеризует их как полициклические. Следует отметить, что оси P. aizoides по функциональному назначению производных апикальной и пазушной меристем соответствуют моноподиальной розеточной модели Т.И. Серебряковой, однако по длине междоузлий такие оси следует относить к моноподиальной полурозеточной модели побегообразования, описанной Н.П. Савиных (2006). В большинстве случаев генеративный пазушный побег закладывается на второй или последующие года развития. Таким образом, универсальным модулем P. aizoides можно считать полициклические поликарпические моноподиальные оси со специализированными вегетативно-генеративными побегами.
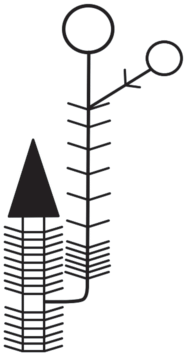
Генеративные побеги обладают анизотропным ростом и в базальной части имеют короткие междоузлия, в средней части – длинные. В апикальной зоне развивается терминальный цветок или малоцветковый дихазий из двух, максимум четырех цветков. После цветения и плодоношения генеративный пазушный побег полностью отмирает.
За счет активного ветвления, благодаря развитию моноподиальных плагиотропных побегов, со временем растение образует густую радиально-плоскую подушку. Пространство между многолетними осями плагиотропных моноподиев и развивающихся на них розеточных побегах плотно заполнено отмершими остатками листьев.
Модель побегообразования Cherleria circassica (M. circassica) моноподиальная розеточная. Укороченные многолетние побеги нарастают моноподиально в течение многих лет. В результате развития пазушных побегов образуется очень плотная радиально-шаровидная подушка, заполненная внутри остатками листьев. Растение имеет хорошо развитую систему главного корня, благодаря которому крепко закреплено в почве. Придаточных корней на побегах не образуется.
На многолетних розеточных побегах за один вегетационный период могут развиваться несколько вегетативно-генеративных облиственных побегов. Таким образом, розеточные побеги являются поликарпическими. Вегетативно-генеративные побеги всегда развиваются из почек, переживших состояние покоя. Такие вегетативно-генеративные почки могут развиваться на втором и последующих годах развития розеточного побега, из этого следует, что такие побеги дициклические. Универсальным модулем C. circassica можно считать полициклические поликарпические моноподиальные оси с дициклическими вегетативно-генеративными побегами (рис. 5).
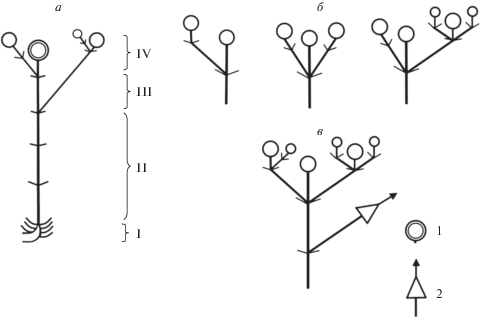
В строении вегетативно-генеративного облиственного побега выделяют несколько структурно-функциональных зон: нижнюю зону торможения, представленную участком побега с короткими междоузлиями; среднюю зону торможения – участок побега с длинными междоузлиями; зону обогащения и зону соцветия (рис. 6). Зона обогащения, если она имеется, обычно включает в себя от одного до трех паракладиев. Зона терминального соцветия представлена дихазием, степень разветвления которого может быть различна.
Рис. 6.
Схема строения: а – генеративного побега С. circassica, б – вариантов терминального соцветия генеративного побега, в – терминального соцветия с паракладием в зоне обогащения. I – нижняя зона торможения, II – средняя зона торможения, III – зона обогащения, IV – зона соцветия. 1 – терминальное соцветие, 2 – паракладий.
Биоморфология C. circassica в целом очень схожа с таковой P. aizoides, однако важной отличительной чертой является то, что C. circassica не образует плагиотропных побегов разрастания, выполняющих функцию захвата близлежащего к особи пространства. Вегетативные побеги C. circassica нарастают скорее вертикально вверх, образуя не более 4–5 порядков ветвления. Вся многолетняя часть подушки представлена безлистными стеблями с короткими междоузлиями (рис. 7).
Рис. 7.
Схема строения дерновинки С. circassica. Порядки ветвления осей указаны только на левой части схемы.
Модель побегообразования Minuartia hirsuta subsp. oreina симподиальная полурозеточная. В побеговой системе M. hirsuta subsp. oreina четко выделяются монокарпические побеги возобновления, обладающие анизотропным ростом и берущие свое начало из почек, переживших состояние покоя, при этом проводя один вегетационный сезон в стадии вегетативного ассимилирующего побега, и лишь на следующий год начинают свое развитие генеративные облиственные побеги с длинными междоузлиями. Такие побеги носят название дициклических. Монокарпический дициклический побег (рис. 8) обладает типичными структурно-функциональными зонами. Зона возобновления представлена базальным участком монокарпического побега с короткими междоузлиями и отмершими прошлогодними листьями, а также почками возобновления и уже сформированными на момент цветения вегетативными побегами замещения, которые на будущий год перейдут в фазу цветения и плодоношения, а на данный момент находятся в состоянии вегетативного ассимилирующего побега. Средняя зона торможения представлена верхним участком розеточного побега с вышерасположенными участками стебля с длинными междоузлиями. В зоне обогащения, как и в зоне соцветия, можно наблюдать широкую морфологическую поливариантность. В зоне обогащения, в случае ее наличия, могут располагаться от одного до пяти паракладиев. В зоне соцветия можно наблюдать от одного терминального цветка до дихазия с 15 и более цветками.
Рис. 8.
Схема строения: а – дикциклического монокарпического побега M. hirsuta subsp. oreina, б – вариантов дихазиев и расположения паракладиев на монокарпическом побеге. I – зона возобновления, II – средняя зона торможения, III – зона обогащения, IV – зона соцветия.
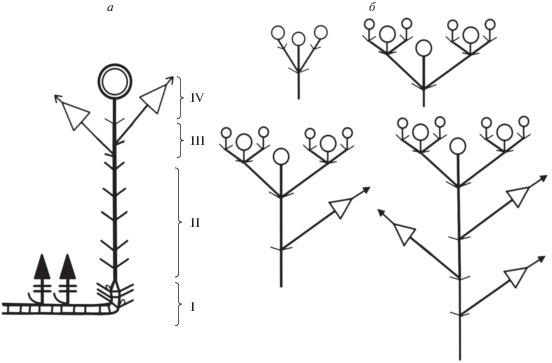
После окончания фазы цветения и плодоношения монокарпический дициклический побег переходит к фазе вторичной деятельности, в процессе которой все части побега выше зоны возобновления отмирают, и остается только нижняя плагиотропная часть, которая входит в состав многолетних осей подушковидной дерновинки.
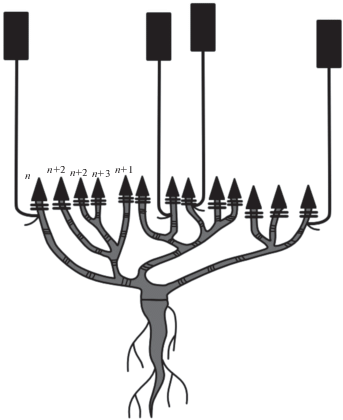
Активное ветвление в процессе побегообразования связано с наличием большого количества почек и побегов возобновления. В среднем на каждом цветущем монокарпическом побеге развивается от одного до четырех побегов возобновления (рис. 9). Универсальным модулем для M. hirsuta subsp. oreina можно считать дициклический монокарпический побег.
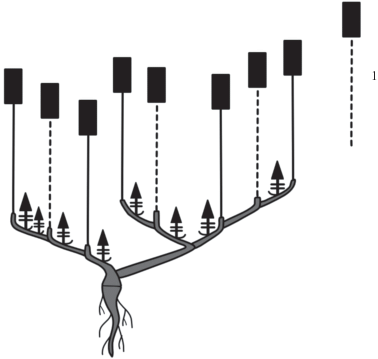
Побеговые системы M. setacea нарастают по симподиальной длиннопобеговой модели. Вертикально растущие монокарпические побеги имеют типичное разделение на структурно-функциональные зоны (рис. 10). Самая нижняя часть монокарпического побега представлена зоной возобновления, в которой сосредоточено большое количество почек возобновления. Зоны торможения нет, и зона возобновления сразу переходит в зону обогащения, в нижней части которой можно наблюдать большое количество вегетативных ассимилирующих побегов, а в верхней – паракладиев. Паракладии имеют сходное строение с монокарпическим побегом, только в разной степени сложности повторяют его структуры: от нескольких междоузлий с пазушными вегетативными побегами и терминальным соцветием в верхней части до большого количества паракладиев, расположенных на исходном паракладии. Зона соцветия в редких случаях представлена либо одним терминальным цветком, либо дихазием, степень разветвления которого может быть различна.
Рис. 10.
Схема строения: а – монокарпического побега M. setacea, б – вариантов терминального соцветия. I – зона возобновления, II – зона обогащения, III – зона соцветия. 1 – соцветие.
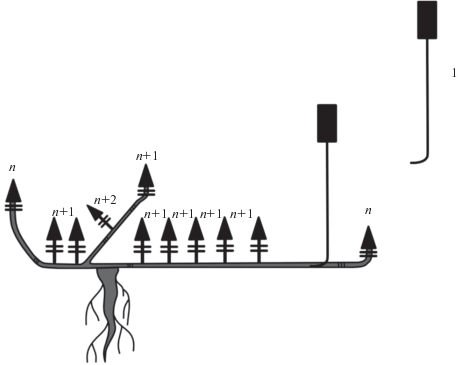
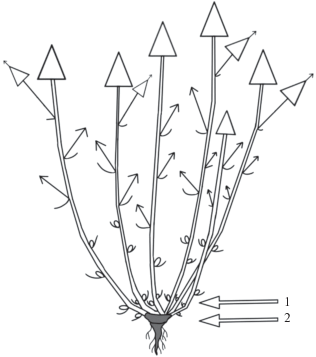
Монокарпические побеги каждый год развиваются из почек возобновления в прикорневой зоне растения. После вегетационного периода монокарпический побег выше зоны возобновления полностью отмирает. Зона возобновления после окончания жизнедеятельности монокарпического побега входит в состав многолетних скелетных осей. Таким образом, можно наблюдать активное базитонное ветвление, в результате которого формируется рыхлая подушковидная дерновинка (рис. 11). Универсальный модуль M. setacea – моноциклический монокарпический побег.
Рис. 11.
Схема строения дерновинки M. setacea. 1 – зона возобновления, 2 – многолетняя часть растения.
Модель побегообразования однолетнего растения M. glomerata длиннопобеговая с ортотропными побегами. В целом побеги M. glomerata имеют схожее с M. setacea строение. На начальных этапах онтогенеза формируется хорошо развитый главный корень, который сохраняется в течение всей жизни растения. Универсальным модулем M. glomerata является синфлоресценция (рис. 12), которая формируется в течение моноподиального нарастания оси, в данном случае – первичного побега. Синфлоресценция представлена большим количеством хорошо развитых паракладиев, в верхней своей части представлена терминальным соцветием – дихазием из 5–15 цветков с очень короткими цветоножками. При нарастании дерновинки можно наблюдать мезотонное ветвление, благодаря активному развитию паракладиев (рис. 13).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Соотношение жизненных форм и моделей побегообразования с условиями обитания. Все изученные модельные виды, представленные в данной работе, являются обитателями гор. Так или иначе, их развитие сопряжено с определенными условиями среды и влиянием на них различных абиотических факторов. В горных местах обитания, особенно на больших высотах, наблюдаются крайне неблагоприятные условия для роста надземных органов растений. Ограничивающими факторами роста можно считать низкую температуру воздуха и почвы, воздействие сильных холодных ветров в безлесной альпийской зоне, различные эдафические и солярные факторы. Для выживания в подобных условиях, безусловно, растениям необходимо обладать определенными адаптациями. Одним из таких приспособлений является подушковидная жизненная форма. Однако следует отметить, что изученные подушковидные представители р. Minuartia s.l. имеют разные стратегии максимально эффективного для подобных условий побегообразования. Этим объясняются их биоморфологические различия.
P. imbricata является самым высокогорным из рассмотренных видов, его можно обнаружить исключительно в альпийском поясе, на ледниковых моренах, мелкокаменистых осыпях и россыпях. Стратегия этого подушковидного растения напрямую связана с очень коротким вегетационным периодом, свойственным высокогорью. Только P. imbricata из всех изученных видов обладает полициклическими поликарпическими моноподиальными осями с редуцированными до одного цветка боковыми генеративными структурами. Такие побеги после окончания вегетационного периода не отмирают, а входят в состав многолетних частей побеговой системы. К тому же P. imbricata не обладает сложными, продолжительное время развивающимися генеративными побегами, а вместо этого формирует силлептические, очень просто устроенные генеративные побеги, как правило, представленные одним цветком. Такой цветок, в силу полурозеточной модели побегообразования, занимает псевдотерминальное положение и лишь после формирования семян переходит в боковое. Таким образом, цветение и плодоношение может продолжаться в течение всего вегетационного периода, при этом одновременно происходит разрастание особи сразу во многих направлениях вследствие развития множества силлептических побегов. К тому же время формирования каждого флорального элемента существенно сокращается в силу простоты его строения. Другими словами, P. imbricata активно наращивает биомассу, при этом захватывая пространство вокруг себя, проявляя стратегию стресс-толерант-рудерала (Онипченко и др., 2020), одновременно выполняя функцию генеративного размножения. Отсутствие сложных многоцветковых дихазиев, а также паракладиев на генеративных побегах полностью компенсируется большим количеством силлептических побегов, непрерывно формирующихся в процессе вегетации.
Другие высокогорные обитатели – P. aizoides, C. circassica и M. hirsuta subsp. oreina – также представляют собой подушковидные растения, однако, в отличие от P. imbricata, формируют специализированные вегетативно-генеративные облиственные побеги, которые после завершения плодоношения отмирают и не входят в состав многолетней части побеговой системы. Эти три вида имеют принципиально отличную от P. imbricata стратегию роста, при которой наращивание многолетних осей происходит отдельно от функционирования генеративных побегов. При этом развитие генеративных и вегетативных структур также происходит параллельно, но дезинтегрировано в пространстве. Необходимость существования вегетативных побегов в розеточной форме, возможно, продиктована суровыми условиями обитания. Розеточные побеги позволяют иметь большое количество ассимилирующих листьев, при этом не сильно возвышаться над поверхностью почвы, что в условиях открытых высокогорий представляется жизненно важным. Однако все так же остро существует проблема короткого вегетационного сезона, и, как следствие, в высокогорье, впрочем, как и в других сообществах, возникает конкуренция за опылителей среди растений. Для более удачного экспонирования своих цветков для опылителей P. aizoides, C. circassica и M. hirsuta subsp. oreina формируют генеративные побеги с длинными междоузлиями, выносящие цветки над поверхностью подушек. Подобное разнесение в пространстве функционально различных структур является примером адаптивного компромисса, в котором многолетнее тело растения существует в розеточной форме, что позволяет наиболее “комфортно” развиваться в высокогорных условия, а генеративные структуры развиваются иначе, по типу побега с длинными междоузлиями, что решает проблему с экспонированием цветка для опылителя. Помимо этого, генеративный побег с длинными междоузлиями позволяет создать несколько структурно-функциональных зон, включая зону обогащения с паракладиями. Именно так зачастую решается проблема немногочисленности развивающихся побегов. За счет создания большого количества паракладиев, как у M. hirsuta subsp. oreina, общее количество цветков существенно увеличивается.
Принципиально иная стратегия роста наблюдается у M. glomerata и M. setacea. Эти два вида обитают в более благоприятных условиях. Вегетационный период существенно длиннее, а также отсутствуют такие неблагоприятные факторы, как холодные штормовые ветра и критически низкие для роста температуры. Вследствие этого наличие розеточных побегов и подушковидной жизненной формы перестает быть столь актуальным, как в случае с высокогорными представителями р. Minuartia s.l. Хотя эти растения различаются по сроку жизни (M. glomerata – однолетник, M. setacea – многолетник), модель побегообразования и стратегия роста у них сходные. Развитие моноциклических монокарпических побегов в начале вегетационного периода позволяет за короткий срок сформировать генеративные побеги, которые, в свою очередь, обладая хорошо развитой зоной обогащения и очень короткой зоной торможения, несут на себе многочисленные паракладии. Развитие паракладиев базипетальное, что позволяет пролонгировать цветение каждой особи. Во время плодоношения монокарпического побега паракладий, расположенный ближе к терминальному соцветию монокарпического побега, начинает зацветать, на смену ему после завершения цветения доходит до завершающей стадии развития паракладий, расположенный ниже него, и так продолжается до конца вегетационного периода. Таким образом, цветение каждого растения существенно пролонгировано во времени.
Основываясь на вышесказанном, можно говорить о трех стратегиях побегообразования для изученных видов р. Minuartia s.l.: 1) по типу M. imbricata с формированием полициклических поликарпических моноподиальных осей с редуцированными до одного цветка боковыми генеративными структурами; 2) по типу M. circassica с развитием розеточных вегетативных побегов и пазушных вегетативно-генеративных побегов с длинными междоузлиями; 3) по типу M. setacea с образованием моноциклических монокарпических побегов.
Биоморфологические характеристики в таксономии рода Minuartia s.l. и возможные пути эволюции жизненных форм. В результате биоморфологического анализа модельных видов стало очевидно, что существует несколько моделей побегообразования (табл. 2), характерных для р. Minuartia s.l. По молекулярно-генетическим данным (Dillenberger, Kadereit, 2014), P. imbricata и P. aizoides являются близкородственными видами, расположенными в одной кладе, что послужило основанием для обособления этих видов в отдельный род. Можно отметить, что для этих представителей характерна одинаковая модель побегообразования – моноподиальная полурозеточная. Для C. circassica, которая по филогенетическим данным имеет обособленное положение на филогенетическом древе, но ранее принадлежала к той же секции, что и представители р. Pseudocherleria, свойственна моноподиальная розеточная модель побегообразования. Модельные виды, которые относятся непосредственно к р. Minuartia s. str.: M. hirsuta subsp. oreina, M. setacea – обладают симподиальными моделями побегообразования и монокарпическими побегами с типичным разделением на структурно-функциональные зоны. Таким образом, можно говорить о том, что новые сепаратные группы, описанные в ранге родов, обладают иными способами нарастания и моделями побегообразования.
Таблица 2.
Биоморфология модельных видов
| Вид | Модель побегообразования | Универсальный модуль |
|---|---|---|
| Pseudocherleria imbricata | Моноподиальная полурозеточная |
Полициклические поликарпические моноподиальные оси с редуцированными до одного цветка боковыми генеративными структурами |
| P. aizoides | Моноподиальная полурозеточная |
Полициклические поликарпические моноподиальные оси со специализированными вегетативно-генеративными побегами |
| Cherleria circassica | Моноподиальная розеточная | Полициклические поликарпические моноподиальные оси с дициклическими вегетативно-генеративными побегами |
| Minuartia hirsuta subsp. oreina | Симподиальная полурозеточная | Дициклический монокарпический побег |
| M. setacea | Симподиальная длиннопобеговая | Моноциклический монокарпический побег |
| M. glomerata | Длиннопобеговая с ортотропными побегами | Синфлоресценция |
Появление в побеговой системе монокарпического побега в р. Minuartia s.l. можно ассоциировать с переходом растений к симподиальному способу нарастания, что является более эволюционно продвинутой чертой (Недолужко, 1997). Нарастание за счет боковых почек сопровождается омоложением вновь возникающей оси по сравнению с осью предыдущего порядка, и чем ниже на побеге располагается замещающая боковая почка, тем сильнее омоложение побеговой системы, а это, в свою очередь, приводит к усилению вегетативного размножения, увеличению общей продолжительности жизни растения, а также ускорению его развития (Серебряков, 1954; Хохряков, 1975). Таким образом, можно предполагать, что существует эволюционная тенденция от более примитивной моноподиальной модели побегообразования с поликарпическими побегами к симподиальной и формированию эволюционно продвинутых монокарпических побегов.
ВЫВОДЫ
По результатам проведенного исследования описаны три модели побегообразования для модельных видов:
1) Для P. imbricata, P. aizoides характерна моноподиальная полурозеточная модель;
2) Представители вида C. сircassica имеют моноподиальную розеточную модель побегообразования;
3) Представители видов M. hirsuta subsp. oreina, M. setacea обладают симподиальными моделями побегообразования с монокарпическими побегами.
Выявлены три стратегии побегообразования у модельных видов:
1) по типу M. imbricata с формированием полициклических поликарпических моноподиальных осей с редуцированными до одного цветка боковыми генеративными структурами;
2) по типу M. circassica с развитием розеточных вегетативных побегов и пазушных вегетативно-генеративных побегов с длинными междоузлиями;
3) по типу M. setacea с образованием моноциклических монокарпических побегов.
В р. Minuartia s.l. отмечен эволюционный тренд к переходу от более примитивной моноподиальной модели побегообразования к симподиальной, а также формированию монокарпических побегов.
Использование биоморфологии в классификации Minuartia s.l. позволит сделать более репрезентативные заключения о систематике рода в целом.
Работа выполнена в рамках темы “Анализ структурного и хорологического разнообразия высших растений в связи с проблемами их филогении и таксономии; проблемы экологии города и устойчивого развития” (№12-2-21, Номер ЦИТИС: 121032500084-6).
Список литературы
Актуальные проблемы современной биоморфологии, 2012 / Под ред. Савиных Н.П. Киров: Радуга-ПРЕСС. 610 с.
Барыкина Р.П., 1999. О некоторых модусах преобразований онто-, органо- и гистогенеза в соматической эволюции сем. Ranunculaceae // Бюлл. МОИП. Отд. биол. Т. 104. № 1. С. 49–52.
Бигон М., Харпер Дж., Таунсенд К., 1989. Экология: Особи, популяции и сообщества. Т. 1. М.: МИР. 667 с.
Гатцук Л.Е., 1994. Иерархическая система структурно-биологических единиц растительного организма, выделенных на макроморфологическом уровне // Успехи экологической морфологии растений и ее влияние на смежные науки. М.: Прометей. С. 18–19.
Гатцук Л.Е., 1995. Комплементарные модели побега и их синтез // Бот. журн. Т. 80. № 6. С. 1–4.
Кожанчиков В.И., 1975. Морфологические признаки семян Caryophyllaceae и возможные пути их эволюции // Бот. журн. Т. 52. № 7. С. 1277–1286.
Лазьков Г.А., 2012. Конспект флоры Кавказа. Т. 3(2). М.; СПб. Т-во науч. изд. КМК. С. 165–170.
Лухтанов В.А., 2013. Принципы реконструкции филогенезов: признаки, модели эволюции и методы филогенетического анализа // Тр. Зоол. ин-та РАН. № 2. С. 39–52.
Лухтанов В.А., Кузнецова В.Г., 2009. Молекулярно-генетические и цитогенетические подходы к проблемам видовой диагностики, систематики и филогенетики // Журн. общ. биологии. Т. 70. № 5. С. 415–437.
Недолужко В.А., 1997. Древесные растения: проблема эволюции жизненных форм. Владивосток: Дальнаука. 120 с.
Онипченко В.Г., Дудова К.В., Ахметжанова А.А. и др., 2020. Какие стратегии растений способствуют их доминированию в альпийских сообществах? // Журн. общ. биологии. Т. 81. № 1. С. 37–46.
Павлинов И.Я., 2001. Концепции систематики и концепции биоразнообразия: проблема взаимодействия // Журн. общ. биологии. Т. 62. № 4. С. 362–366.
Савиных Н.П., 2006. Род вероника: морфология и эволюция жизненных форм. Киров: Изд-во ВятГГУ. 324 с.
Савиных Н.П., 2008. Применение концепции модульной организации к описанию структуры растения // Современные подходы к описанию структуры растений. Киров: ООО “Лобань”. С. 47–69.
Савиных Н.П., 2019. Эволюция жизненных форм цветковых растений в формировании биологического разнообразия // Изв. РАН. Сер. биол. № 1. С. 72–80.
Савиных Н.П., Черемушкина В.А., 2015. Биоморфология: современное состояние и перспективы // Сиб. экол. журн. Т. 22. № 5. С. 659–670.
Серебряков И.Г., 1954. О методах изучения ритмики сезонного развития растений в геоботанических стационарах // Доклады совещания по стационарным геоботаническим исследованиям. Л.: Изд-во АН СССР. С. 145–159.
Серебряков И.Г., 1964. Жизненные формы высших растений и их изучение // Полевая геоботаника. Т. 3. М.; Л.: Наука. С. 146–208.
Серебрякова Т.И., 1977. Об основных “архитектурных моделях” травянистых многолетников и модусах их преобразования // Бюлл. МОИП. Отд. биол. Т. 82. № 5. С. 112–128.
Серебрякова Т.И., 1981. Жизненные формы и модели побегообразования наземно-ползучих многолетних трав // Жизненные формы: Структура, спектры и эволюция. М.: Наука. С. 161–179.
Серебрякова Т.И., 1987. О вариантах моделей побегообразования у многолетних трав // Морфогенез и ритмы развития высших растений: Межвуз. сб. науч. тр. М.: МГПИ им. В.И. Ленина. С. 3–19.
Современные подходы к описанию структуры растений, 2008 / Под ред. Савиных Н.П., Бобровова Ю.А. Киров: ООО “Лобань”. 355 с.
Хохряков А.П., 1975. Закономерности эволюции растений. Новосибирск: Наука. 202 с.
Цвелев Н.Н., 2005. Проблемы теоретической морфологии и эволюции высших растений. Сб. избр. тр. М.; СПб.: Т-во науч. изд. КМК. 407 с.
Шафранова Л.М., 1980. О метамерности и метамерах у растений // Журн. общ. биологии. Т. 41. № 3. С. 437–447.
Шафранова Л.М., 1981. Ветвление растений: процесс и результат // Жизненные формы: структура, спектры и эволюция. М.: Наука. С. 179–212.
Шафранова Л.М., 1993. Растение как объект гомологизации // Жизненные формы: онтогенез и архитектура. М.: Прометей. С. 219–222.
Шафранова Л.М., Гатцук Л.Е., 1994. Растение как пространственно- временная метамерная (модульная) система // Успехи экологической морфологии растений и ее влияние на смежные науки: Межвуз. сб. науч. тр. М.: Прометей. С. 6–7.
Amini E., Zarre S.H., Assadi M., 2011. Seed micromorphology and its systematic significance in Gypsophila (Caryophyllaceae) and allied genera // Nord. J. Bot. V. 29. P. 660–669.
Çelebioğlu T., Favarger C., 1982. Contribution à la cytotaxonomie du genre Minuartia L. (Caryophyllacées) en Turquie et dans quelques régions voisines // Biol. Ecol. Medit. V. 9. P. 139–160.
Çelebioğlu T., Favarger C., Huynh K.L., 1983. Contribution a la micromorphologie de la testa des graines du genre Minuartia L. (Caryophyllaceae) sect. Minuartia // Adansonia. V. 4. P. 415–435.
Dillenberger M.S., Kadereit J.W., 2014. Maximum polyphyly: Multiple origins and delimitation with plesiomorphic characters require a new circumscription of Minuartia (Caryophyllaceae) // Taxon. V. 63. № 1. P. 64–88.
Favarger C., Recoder P.M., 1987. Commentaires sur la caryologie des espèces de Minuartia L. de la Péninsule Ibérique // Anales Jard. Bot. Madrid. V. 44. P. 558–564.
Galland N., 1988. Recherche sur l’origine de la flore orophile du Maroc étude caryologique et cytogéographique / Trav. Inst. Sci. Sér. Bot. V. 35. Rabat: Univ. Mohammed V, Institut scientifique. 168 р.
Halliday G., 1993. Minuartia // Flora Europaea. V. 1. 2nd ed. / Eds Tutin T.G., Burges N.A., Chater A.O., Edmondson J.R., Heywood V.H., et al. Cambridge: Cambridge Univ. Press. P. 152–160.
Hennig W., 1966. Phylogenetic Systematics. Urbana: Univ. Illinois Press. 263 p.
Judd W.S., Campbell C.S., Kellogg E.A., Stevens P.F., 1999. Plant Systematics: A Phylogenetic Approach. Sunderland: Sinauer. 464 p.
Kamari G., 1997. Minuartia L. // Flora Hellenica. V. 1 / Eds Strid A., Tan K. Konigstein: Koeltz Scientific Books. P. 170–191.
Mattfeld J., 1922. Geographisch-genetische Untersuchungeil iiber die Gattung Minuartia (L.) Hiern // Feddes Repert. Beih. V. 15. P. 1–228.
McNeill J., 1962. Taxonomic studies in the Alsinoideae I: Genetic and infrageneric groups // Notes Roy. Bot. Gard. Edinburgh. V. 24. P. 79–155.
McNeill J., 1963. Taxonomic studies in the Alsinoideae II: A revision of the species in the Orient // Notes Roy. Bot. Gard. Edinburgh. V. 24. P. 241–404.
Minuto L., Roccotiello E., Casazza G., 2011. New seed morphological features in Moehringia L. (Caryophyllaceae) and their taxonomic and ecological significance // Plant Syst. Evol. V. 145. P. 60–67.
Minuto L., Fior S., Roccotiello E., Casazza G., 2006. Seed morphology in Moehringia L. and its taxonomic significance in comparative studies within the Caryophyllaceae // Plant Syst. Evol. V. 262. P. 189–208.
Parkin I., 1914. The evolution of the inflorescence // J. Linn. Soc. V. 42. № 287. P. 511–563.
Rabeler R.K., Hartman R.L., Utech F.H., 2005. Minuartia // Flora of North America. V. 5 / Ed. Flora of North America Editorial Committee. Oxford: Oxford Univ. Press. P. 16–136.
Rechinger K.H., 1988. Minuartia L. // Flora Iranica. V. 163 / Ed. Rechinger K.H. Graz: Akademische Druck- und Verlagsanstalt. P. 28–53.
Schuh R.T., Brower A.V.Z., 2009. Biological Systematics: Principles and Applications. 2nd ed. Ithaca, NY: Cornell Univ. Press. 328 p.
Soltis D.E., Soltis P.S., Endress P.K., Chase M.W., 2005. Phylogeny and Evolution of Angiosperms. Sunderland: Sinauer. 370 p.
Troll W., 1964. Die Infloreszenzen. Bd. 1. Jena: Fischer Verlag. 615 s.
Yıldız K., 2002. Seed morphology of Caryophyllaceae species from Turkey (North Anatolia) // Pak. J. Bot. V. 34. P. 161–171.
Zaychenko S.G., Zernov A.S., 2017. Structural features of the seed coat in Caucasian representatives of Minuartia L. (Caryophyllaceae) // Wulfenia. V. 24. P. 205–220.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии