

Журнал высшей нервной деятельности им. И.П. Павлова, 2021, T. 71, № 4, стр. 529-546
Отражение процесса созревания ЦНС у детей и подростков Северного региона РФ в динамике интегральных параметров ЭЭГ
В. П. Рожков 1, *, М. И. Трифонов 1, С. И. Сороко 1
1 Институт эволюционной физиологии и биохимии им. И.М. Сеченова Российской академии наук
г. Санкт-Петербург, Россия
* E-mail: vlrozhkov@mail.ru
Поступила в редакцию 07.12.2020
После доработки 25.01.2021
Принята к публикации 02.03.2021
Аннотация
Представлены результаты анализа формирования биоэлектрической активности мозга у детей и подростков, проживающих в северном регионе России, на основе оценки интегральных параметров многоканальной ЭЭГ. В исследованиях приняли участие 33 мальчика и 32 девочки в возрасте от 7 до 18 лет, учащихся сельской школы. Работа проведена в 2 этапа, каждый из которых состоял из осенней и весенней сессии. Регистрировали ЭЭГ, по которой строили структурную функцию и рассчитывали обобщенный спектр, а также интегральные параметры, характеризующие меру временно́й (0 ≤ Pt ≤ 1) и пространственной (0 ≤ Ps ≤ 1) связности ЭЭГ-процессов. Предельные значения параметров (0, 1) соответствуют случаю либо полностью упорядоченной, либо совершенно случайной пространственной или временно́й организации ЭЭГ. Выявлено уменьшение с возрастом значения параметра Ps, свидетельствующее о повышении пространственной связности ЭЭГ-процессов у учащихся от младших к старшим классам. Напротив, величина Pt с возрастом увеличивалась, что характеризовало уменьшение временного масштаба связности, инерционности ЭЭГ-процессов и могло отражать повышение уровня функциональной подвижности (лабильности) ЦНС по мере взросления. Показана зависимость оценок возрастной динамики интегральных параметров ЭЭГ у школьников-северян от сезона (осень-весна) и выборочного контингента обследуемых.
Важной задачей применения ЭЭГ в медицинской и исследовательской практике является индивидуальная диагностика уровня зрелости мозга. Период жизни от 6–7 до 17–18 лет (“школьный возраст”) характеризуется интенсивным физическим, психическим и социальным развитием ребенка. Происходят существенные изменения когнитивных функций с совершенствованием процессов восприятия, внимания, памяти (Безруких и др., 2009; Anderson, Perone, 2018). Процессы когнитивного развития, с одной стороны, связаны с морфофункциональным созреванием ЦНС, с другой – сами являются драйвером такого созревания.
Доказательства структурного и функционального созревания мозга на протяжении не только детского, но и всего подросткового возрастного периода получены в нейроанатомических и функциональных МРТ- и ЭЭГ-исследованиях. На протяжении второго десятилетия жизни значительно меняются соотношения объемов серого и белого вещества головного мозга (Gedd, 2004). Объем белого вещества непрерывно, хотя и с замедлением, увеличивается на протяжении возрастного периода от 10 до 30 лет (Whitford et al., 2007). В фундаментальных трудах С.Б. Дзугаевой (Дзугаева, 1975) содержатся сведения о последовательности роста и созревания проекционных и ассоциационных путей головного мозга. К числу наиболее поздно дифференцирующихся (вплоть до зрелого возраста) относятся структуры верхнего продольного пучка, соединяющего кору лобной доли с затылочной и рядом полей теменной и височной долей. Заметно увеличивается в объеме в возрасте 7 лет и позднее важнейший комплекс комиссуральных связей – мозолистое тело, обеспечивающее сопряженную деятельность обоих полушарий.
Уровень функциональной зрелости мозга отражается в характере ЭЭГ покоя, регистрируемой в состоянии спокойного бодрствования. Показано, что с возрастом относительная мощность высокочастотных составляющих альфа-ритма растет, а относительная мощность низкочастотных составляющих ЭЭГ падает (Gasser et al., 1988; Алферова, Фарбер, 1990; Горбачевская, Кожушко, 1990). По изменениям соотношений активности в основных диапазонах частот ЭЭГ установлен принцип гетерохронного функционального созревания различных областей коры головного мозга (Алферова, Фарбер, 1990; Горбачевская, Кожушко, 1990, Vijayakumar et al., 2018). Важные сведения о направленности возрастных изменений, свидетельствующих о созревании ЦНС, получены с использованием оценок статистической связи между локальными колебаниями потенциалов в различных областях мозга методами кросс-корреляционного (в широком диапазоне частот) и когерентного (в ряду гармонических составляющих) анализа. Несмотря на серьезные замечания относительно корректности использования анализа когерентности в ЭЭГ (Кулаичев, 2009), получены воспроизводимые различными исследовательскими группами результаты, свидетельствующие о повышении с возрастом у детей и подростков когерентности отдельных гармонических составляющих в ЭЭГ (Barry et al., 2004; Мачинская и др., 2007; Thatcher et al., 2008; Gmehlin et al., 2011).
Описаны возрастная динамика и топические особенности формирования межполушарных и межрегиональных взаимосвязей ЭЭГ-процессов в левом и правом полушариях мозга (Цицерошин, Шеповальников, 2009; Gmehlin et al., 2011), которые в целом указывают на генеральную тенденцию к повышению “функциональной связности” или уровня пространственной (дистантной) синхронизации биопотенциалов мозга ЭЭГ-процессов.
Развитие количественного подхода к определению возрастных ЭЭГ-нормативов идет по пути уточнения характера зависимости отдельных показателей от возраста, а также поиска интегральных ЭЭГ-критериев созревания ЦНС с учетом гетерохронии формирования различных корковых регионов и волоконных структур связей между ними. Здесь речь идет как о выборе обобщенных характеристик многоканальной ЭЭГ, так и об определении набора показателей, полномерно отражающих процесс формирования ЦНС в заданном временно́м периоде. К числу таких показателей могут быть отнесены обобщенные характеристики частотной, пространственной и временно́й организации электрической активности мозга, оцениваемые по всей совокупности ЭЭГ-процессов.
Примерами интегральных характеристик пространственной связности ЭЭГ-процессов могут служить оценки коротко-дистантных и длинно-дистантных (Barry et al., 2004; Thatcher et al., 2008), “локальных” и “глобальных” (Gmehlin et al., 2011), в зависимости от межэлектродных расстояний, уровней когерентности, а также параметр VOL, характеризующий обобщенный уровень пространственной синхронизации биопотенциалов мозга. Параметр VOL рассчитывается на основе матрицы коэффициентов кросс-корреляции ЭЭГ (Барвинок, Рожков, 1992), которая в своей геометрической интерпретации позволяет представить совокупность ЭЭГ-процессов как пучок векторов, занимающий определенный “объем” в многомерном пространстве.
Показано, что при переходе учащихся от младшего школьного возраста к старшему происходит постепенное увеличение как величин “локальных” и “глобальных” уровней когерентности (Gmehlin et al., 2011; Snyder et al., 2015), величин коротко-дистантных (Thatcher et al., 2008), длинно-дистантных (Barry et al., 2004) когерентных связей, так и обобщенных уровней пространственной синхронизации ЭЭГ (Сороко и др., 2012; Кручинина и др., 2020).
Эти показатели характеризуют пространственную структуру ЭЭГ, без учета ее временно́й организации. Между тем процесс созревания ЦНС ярко проявляется в изменениях временно́й организации паттерна ЭЭГ, структуры взаимодействия волновых компонентов основных ритмов ЭЭГ (Сороко и др., 2012). Анализ вероятностных переходов волн ЭЭГ из одних частотных диапазонов к другим и использование подходов, разработанных в рамках теории графов и потоков в сетях, позволил выявить маркеры и определить “критические” возрасты, когда меняется “алгоритм” взаимодействия волновых компонентов ЭЭГ (Сороко и др., 2012, Сороко и др., 2015). Однако в большинстве случаев анализ во временно́й области (time domain) переводится в плоскость изучения временны́х или фазовых отношений колебаний потенциалов в различных отведениях ЭЭГ (Jiruška, 2005). Используют также и мультивариативные статистические методы, основанные на анализе причинности Грейнджера (Granger causality) (Cekic et al., 2015; Stokes, Purdon, 2017) или на построении авторегрессионной модели прогнозирования, что позволяет выявлять направленные функциональные (“причинные”) взаимодействия активности в различных отведениях ЭЭГ (Seth, 2015; Мачинская и др., 2019).
В нашем исследовании были использованы оценки интегральных пространственных и временны́х параметров, которые были получены на основе построения и анализа структурной функции (СФ1) многоканальной ЭЭГ. Структурная функция, характеризующая многоканальные процессы, позволяет количественно оценивать масштабы их временно́й и пространственной связности (Trifonov, 2016; Rozhkov et al., 2018) и описывать общие, системные реакции мозга без подробного описания локальных изменений. При этом интегральный временно́й параметр может характеризовать изменения физиологической лабильности ЦНС (по А.А. Ухтомскому, 1951), а интегральный пространственный – обобщенный уровень пространственной синхронизации биопотенциалов мозга (по М.Н. Ливанову, 1972).
Применение обобщенных (интегральных) показателей облегчает решение сложной проблемы межиндивидуальной вариабельности физиологических реакций и процессов, обеспечивающих деятельность человека и его адаптацию к среде. Оценка индивидуальных или индивидуально-типологических особенностей функционирования нейрофизиологических механизмов является одним из ведущих трендов современных исследований организации нервных сетей покоя (resting-state networks) (Гаврон и др., 2019), которые могут определять специфику эмоционального и мотивационного контроля и реализации исполнительных функций (Kurgansky et al., 2020), а также изучения динамических перестроек электрической активности мозга при воздействии факторов внешней среды (Рожков и др., 2019) и в процессе решения когнитивных заданий (Seghier, Price, 2018; Лебедева, Каримова, 2020).
Формирование физиологических функций и развитие организма происходят на фоне адаптации к условиям внешней среды. Неблагоприятные природно-климатические, сложные социально-бытовые условия жизни на Севере предъявляют повышенные требования к физиологическим системам растущего организма (Грибанов и др., 2016а). В течение года организм проходит естественные циклы сезонной акклиматизации к холодному и теплому периоду с контрастными изменениями фотопериодизма (Евдокимов и др., 2007).
Эти сезонные перестройки у жителей Севера обнаруживаются на уровне метаболических процессов (Бойко, 2005), выявляются в цирканнуальных ритмах нейроэндокринных функций (Кубасов и др., 2006). Изменение продолжительности светового дня является одним из наиболее значимых факторов, влияющих на формирование биоэлектрической активности головного мозга у школьников-северян (Грибанов и др., 2016б). Процесс адаптации определяется функциональными возможностями организма, ее “ценой” может быть истощение физиологических резервов, что проявляется в замедлении темпов созревания организма и формирования ЦНС (Сороко и др., 2005).
Целью работы являлось изучение возрастной динамики формирования биоэлектрической активности мозга у детей и подростков-северян школьного возраста на основе анализа интегральных параметров многоканальной ЭЭГ, характеризующих временно́й и пространственный масштабы связности колебаний ЭЭГ-процессов, а также обобщенных по совокупности отведений ЭЭГ левого и правого полушария показателей частоты основного ритма и “глобальной мощности поля” (многомерного аналога дисперсии колебаний потенциалов). В числе отдельных задач было сопоставление оценок возрастной динамики показателей при обследовании одного и того же контингента учащихся в осенний и весенний период, а также двух различающихся по списочному составу групп учащихся, в разное время проживающих на одной и той же территории.
МЕТОДИКА
В исследованиях приняли участие 33 мальчика и 32 девочки в возрасте от 7.6 года до 18 лет, учащихся сельской школы Архангельской обл. (61° с.ш., 41° в.д.). Работа проведена в два этапа, каждый из которых включал осеннюю и весеннюю сессии. На первом этапе в ходе сессии 1 (октябрь 2008 г.) обследованы 50 школьников, сессии 2 (апрель 2009 г.) – 51 школьник, на втором этапе в ходе сессии 3 (октябрь 2012 г.) – 31 школьник, сессии 4 (март 2013 г.) – 31 школьник. Сведения о количестве мальчиков и девочек, разбитых на 3 возрастные группы (младшая, средняя, старшая) для каждого сезона обследования, даны в табл. 1. Младшую группу составляли школьники 7–11.5 лет, среднюю – школьники 11.6–14.5 лет и старшую – школьники 14.6–18 лет. Все 50 школьников, участвовавших в обследованиях на 1-м этапе, приняли участие в работе на втором этапе. В числе обследуемых на 2-м этапе были 23 школьника, участвовавших в работе и в осеннюю, и в весеннюю сессии. Всего в течение двух этапов работы и 4 сессий выполнено 163 регистрации ЭЭГ. Все исследования проводились с соблюдением требований Хельсинкской Декларации. Протокол исследований утвержден Комиссией по этике Ученого совета ИЭФБ РАН. Родители учащихся были информированы о целях обследования и дали согласие на его проведение. Все испытуемые участвовали в исследованиях добровольно.
Таблица 1.
Количество мальчиков и девочек в трех возрастных группах, обследованных на каждом из этапов Table 1. Number of males and females in three age groups surveyed at each of the four sessions
| Возрастная группа | Этап 1, сессия 1 | Этап 1, сессия 2 | Этап 2, сессия 3 | Этап 2, сессия 4 | ||||
|---|---|---|---|---|---|---|---|---|
| мальчики | девочки | мальчики | девочки | мальчики | девочки | мальчики | девочки | |
| Младшая | 8 | 7 | 9 | 7 | 6 | 4 | 6 | 6 |
| Средняя | 8 | 10 | 8 | 10 | 6 | 3 | 7 | 4 |
| Старшая | 9 | 8 | 9 | 8 | 5 | 7 | 5 | 3 |
| Итого | 25 | 25 | 26 | 25 | 17 | 14 | 18 | 13 |
ЭЭГ регистрировали на электроэнцефалографе ЭЭГА-21/26 “Энцефалан-131–03” (фирмы “Медиком МТД”, Россия) в 21 отведении по международной системе “10–20” в полосе 0.5–70 Гц с частотой дискретизации 250 Гц. Использовали монополярные отведения с объединенными ушными референтными электродами. Представленные в статье результаты относятся к 16 отведениям: Fp1, Fp2, F7, F3, F4, F8, T3, C3, C4, T4, T5, P3, P4, T6, O1, O2.
Регистрацию ЭЭГ проводили по схеме стандартного клинико-диагностического обследования с записью ЭЭГ в состоянии спокойного бодрствования с закрытыми и открытыми глазами с применением функциональных нагрузок (фотостимуляция, гипервентиляция). Испытуемые располагались в удобном кресле с подлокотниками. Представленные результаты характеризуют состояние спокойного бодрствования с закрытыми глазами. Участки записей, содержащие двигательные, электроокулографические, электродные артефакты, исключали из обработки. Для дальнейшего анализа частотный диапазон ЭЭГ ограничивали пределами от 1.6 до 30 Гц.
Согласно предложенному нами ранее методу анализа ЭЭГ-данных (Trifonov, 2016; Рожков и др., 2019), каждая запись ЭЭГ, представленная в виде векторного временно́го ряда последовательных значений амплитуд в n отведениях (n = 8 для отведений левого или правого полушария, n = 16 для всей совокупности отведений), предварительно центрировалась путем вычитания соответствующего среднего в каждом отведении и нормировалась на величину определителя выборочной ковариационной матрицы в степени 1/(2n). В результате получали новый векторный временно́й ряд, который являлся безразмерным и характеризовался одинаковой выборочной обобщенной дисперсией для всех испытуемых. Полученный векторный временно́й ряд анализировали на основе СФ1 (Trifonov, 2016), определяемой как среднее приращение амплитуды вектора в 8- или 16-мерном (по числу отведений ЭЭГ) пространстве на интервалах, кратных интервалу дискретизации ЭЭГ (4 мс).
Определенная выше СФ1 характеризует ЭЭГ-запись в целом, учитывая сигналы от всех 16 (или 8) отведений, и представляет собой кривую, имеющую начальную восходящую ветвь, переходящую или стремящуюся в пределе к горизонтальной асимптоте Asf, как показано на рис. 1.
Рис. 1.
Структурная функция многоканальной ЭЭГ (CФ1) и характеристики, используемые для расчета интегральных параметров ЭЭГ. По горизонтальной оси (Δt) – временной интервал, мс, по вертикальной оси (СФ1) – значение структурной функции в отн. ед. Asf – горизонтальная асимптота реальной CФ1, СФ1ш – горизонтальная асимптота для гипотетической случайной (“шумовой”) ЭЭГш, СФ1(Δt) – значение СФ1 при минимальном временно́м интервале (лаге) Δt = 4 мс.
Fig. 1. Structure function of multichannel EEG (SF1) and features used to calculate integral EEG parameters. Along the horizontal axis (Δt) – time lag, ms, along the vertical axis (SF1) – the value of the structure function in rel. units. Asf – horizontal asymptote of real SF1, SF1sh – horizontal asymptote for a hypothetical random (“noise”) EEGsh, SF1 (Δt) – value of SF1 at the minimal time lag Δt = 4 ms.
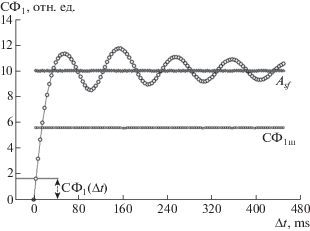
Оценивали значение СФ1(Δt) при минимальном временно́м масштабе Δt, величину Asf, а также значение горизонтальной асимптоты СФ1ш для гипотетической случайной (“шумовой”) ЭЭГш. Для получения оценки Asf произ-водился расчет СФ1п путем случайного перемешивания порядка следования значений амплитуд реальной ЭЭГ одновременно по всем отведениям. Такое перемешивание производилось в среде MATLAB с использованием функции randperm и приводило к разрушению корреляционных связей между значениями амплитуд внутри каждого отведения, но сохраняло неизменной выборочную корреляционную матрицу. Значение СФ1ш вычислялось по аналитической формуле (Вадзинский, 2001). На основе СФ1(Δt), Asf и СФ1ш рассчитывали величины Ps = СФ1ш/Asf (0 ≤ ≤ Ps ≤ 1) и Pt = СФ1(Δt)/Asf (0 ≤ Pt ≤ 1).
Важно подчеркнуть, что временна́я упорядоченность относится здесь к минимальному временно́му масштабу Δt. Крайние значения (0 и 1) параметров Ps и Pt соответствуют полностью детерминированной и “случайной” пространственной и, соответственно, временно́й организации ЭЭГ в целом. Отметим, что параметр Pt служит для оценки масштаба интегральной временно́й корреляции многоканальной ЭЭГ, тогда как значение Ps характеризует уровень пространственной взаимосвязи ЭЭГ между всеми отведениями, поскольку величина Asf связана с определителем выборочной корреляционной матрицы ЭЭГ (Trifonov, 2016).
Оценки Ps, Pt были получены как для 4-секундных, так и для более длительных (порядка 30–60 с) чистых от артефактов эпох анализа ЭЭГ. Расчет СФ1, определение параметров Ps, Pt, а также спектральный анализ СФ1 проводился в среде MATLAB, причем для оценки спектральной плотности мощности (спектра) СФ1 использовалась функция fft. Прямое дискретное преобразование Фурье выполнялось на интервале 8.192 с (2048 отсчетов) за пределами начальной восходящей ветви СФ1.
К числу анализируемых интегральных показателей было также добавлено значение обобщенной (генеральной) дисперсии GV, рассчитываемой по матрице ковариации 16-канальной ЭЭГ, а также по 8 отведениям отдельно для левого (Fp1, F7, F3, T3, C3, T5, P3, O1) и правого (Fp2, F4, F8, C4, T4, P4, T6, O2) полушария мозга. Обобщенная дисперсия является многомерным аналогом одномерной дисперсии, которая характеризует диапазон колебаний случайной величины, и является оценкой абсолютных размеров вариации амплитуды ЭЭГ-сигналов в совокупности отведений.
Статистическую связь между возрастом школьников и величиной параметров СФ1 и GV оценивали на основе коэффициентов корреляции по Пирсону. С целью изучения влияния факторов “Сессия” (4 градации), “Пол” (2 градации) и “Возраст” (3 градации: 7–11.5 лет, 11.6–14.5 лет и 14.5–18 лет) на показатели ЭЭГ использовали трехфакторный дисперсионный анализ (ANOVA). Применяли процедуру апостериорных сравнений с использованием критериев Фишера и Тьюки. Коэффициенты корреляции, а также различия между средними величинами параметров считали значимыми при p < 0.05. Статистическая обработка полученных данных производилась при помощи пакета программ Statistica-10.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Данные дисперсионного анализа, в котором в качестве зависимой переменной взята возрастная группа, а предикторами служили пол ребенка и сессия, показывают, что возрастные группы были относительно сбалансированы по числу мальчиков и девочек на каждом этапе исследований. Об этом свидетельствует то, что главные эффекты взаимодействия “возрастная группа – пол”: F(1, 155) = = 0.23, p = 0.64 и “возрастная группа – сессия”: F(3, 155) = 0.40, p = 0.75 – статистически незначимы.
Частоту доминирующего ритма ЭЭГ индивидуально для каждого школьника определяли по положению максимума амплитудного спектра СФ1, рассчитанной по 16 отведениям ЭЭГ. Поскольку СФ1 рассчитывали на основе многоканальной ЭЭГ, этот показатель служил обобщенной характеристикой индивидуальной частоты доминирующего ритма для ЭЭГ в целом. Как показал трехфакторный дисперсионный анализ, на численное значение частоты доминирующего ритма не оказывали значимого влияния факторы Сессия и Пол (F(3, 139) = 0.64, p = 0.59; F(1, 139) = = 0.71, p = 0.39 соответственно), слабая статистическая связь обнаружилась с фактором Возраст (F(2, 139) = 3.14, p = 0.046). Возрастная динамика показателя частоты с аппроксимацией методом наименьших квадратов показана на рис. 2.
Рис. 2.
Оценка возрастной динамики частоты основного ритма ЭЭГ по 4 выборкам школьников в 4 сессиях исследования. По горизонтальной оси – возраст в годах, по вертикальной оси – частота основного ритма, в Гц. Каждый символ соответствует одному ребенку; школьники, участвовавшие в исследованиях в одной и той же сессии, обозначены одинаковыми символами. Сплошные и штриховые линии аппроксимируют изменения параметра с возрастом методом наименьших квадратов.
Fig. 2. Assessment of age-related dynamics of the frequency of the main EEG rhythm based on 4 samples of the schoolchildren at 4 sessions of the study. The horizontal axis is an age, in years, the vertical axis is the frequency of the main rhythm, in Hz. Each symbol corresponds to one child; the schoolchildren who participated in the research at the same session are marked with the same symbols. Solid and dashed lines approximate changes in parameter with age using the least squares method.
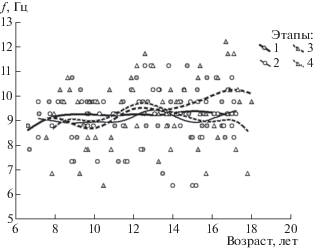
Коэффициенты корреляции между частотой доминирующего ритма и возрастом школьников составили на 1-й сессии r = 0.09, на 2-й сессии r = 0.07, на 3-й сессии r = 0.36 (p = 0.046), на 4-й сессии r = –0.001, по совокупности наблюдений r = 0.13. То есть только для одной (третьей) сессии обследования выявлена значимая статистическая связь между переменными, характеризующая слабо выраженную тенденцию к увеличению частоты доминирующего ритма ЭЭГ с возрастом у школьников.
С учетом отсутствия значимого влияния на величину данного показателя факторов Сессия и Пол и минимального влияния фактора Возраст можно дать совокупную оценку распределения доминирующей частоты СФ1 ЭЭГ по всему контингенту школьников: размах от 6.3 до 12.2 Гц, среднее 9.2 ± 1.18 Гц (m ± sd), медиана 9.3 Гц, 1-я квартиль 8.8 Гц, 3-я квартиль 9.9 Гц. При этом у 8 школьников доминирующая частота ЭЭГ на том или ином этапе обследования была менее 7 Гц.
Значения трех других интегральных параметров оценивали отдельно для отведений левого и правого полушария мозга.
В табл. 2 приведены данные дисперсионного анализа, характеризующие зависимость величин трех интегральных показателей многоканальной ЭЭГ: Pt, Ps и GV – от влияния трех факторов: Возраста и Пола школьников, а также Сессии обследования. Величина каждого из трех анализируемых показателей зависела от влияния факторов Сессия исследования и Возраст и не зависела от фактора Пол. Принимая во внимание отсутствие значимых различий у мальчиков и девочек в величинах Pt, Ps и GV в каждой из трех возрастных групп во всех сессиях исследования, дальнейший анализ был проведен без учета пола детей и подростков (данные мальчиков и девочек были объединены).
Таблица 2.
Оценка влияния факторов “Сессия”, “Возраст” и “Пол” на значения трех интегральных показателей ЭЭГ для левого и правого полушария мозга (данные дисперсионного анализа, n = 163) Table 2. Estimates of the influence of the factors “Session”, “Age” and “Sex” on the values of three integral EEG parameters for the left and right hemispheres of the brain (analysis of variance, n = 163)
| Показатель, фактор | Левое полушарие | Правое полушарие | ||||
|---|---|---|---|---|---|---|
| Pt | Ps | GV | Pt | Ps | GV | |
| Сессия | 23.05** | 6.35** | 9.76** | 28.68** | 3.31* | 7.70** |
| Пол | 2.31 | 0.01 | 0.02 | 1.14 | 1.16 | 0.01 |
| Возраст | 16.02** | 10.70** | 24.07** | 14.55** | 12.75** | 21.74** |
Диаграммы рассеяния на рис. 3 отображают зависимость параметра Pt от возраста учащегося. Логарифмическая аппроксимация динамики Pt показывает тенденцию к увеличению с возрастом у школьников значения Pt, рассчитанного по СФ1 как для левого, так и для правого полушария мозга. Эта тенденция более выражена для 3-ей и 4-ой сессий исследования, чем для 1-ой и 2-ой. Это находит отражение в величинах коэффициентов корреляции между значением Pt и возрастом (табл. 3). Значимые различия в величинах коэффициентов корреляции выявлены в сессиях 1 и 3 для правого полушария мозга (р = 0.024). Возрастная динамика параметра Pt характеризует тенденцию к уменьшению масштаба временно́й связности в организации биоэлектрической активности мозга у школьников от младших к старшим классам. Численные значения параметра Pt для ЭЭГ левого и правого полушария мозга значимо не различались в каждой из 4 сессий исследования (величина t-критерия варьировала от 0.41 до 1.02, p > 0.05).
Рис. 3.
Оценка возрастной динамики параметра Pt, рассчитанного по СФ1 ЭЭГ для левого (а) и правого (б) полушария мозга по 4 выборкам школьников в 4 сессиях исследования. По горизонтальной оси – возраст в годах, по вертикальной оси – значение Pt в отн. ед. Каждый символ соответствует одному ребенку; школьники, участвовавшие в исследованиях в одной и той же сессии, обозначены одинаковыми символами. Черные линии: толстая – сессия 1, тонкая – сессия 2; серые линии: толстая – сессия 3, тонкая – сессия 4. Аппроксимация динамики логарифмической функцией.
Fig. 3. Assessment of the age dynamics of the Pt parameter, calculated from the EEG SF1 for the left (a) and the right (б) cerebral hemispheres, based on 4 samples of the schoolchildren at 4 sessions of the study. The horizontal axis is the age, in years, the vertical axis is the Pt value in relative units. Each symbol corresponds to one child, the schoolchildren who participated in the research at the same session are marked with the same symbols. Black lines: thick – session 1, thin – session 2; gray lines: thick – session 3, thin – session 4. Approximation of the dynamics is done by a logarithmic function.
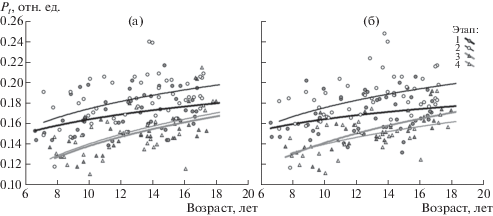
Таблица 3.
Коэффициенты корреляции между изменениями интегральных параметров ЭЭГ и возрастом школьников Table 3. Correlation coefficients between changes in the integral parameters of the EEG and the age of schoolchildren
| Параметр | Левое полушарие | Правое полушарие | n | ||||
|---|---|---|---|---|---|---|---|
| Pt | Ps | GV | Pt | Ps | GV | ||
| Сессия 1 | 0.34* | –0.60*** | –0.50*** | 0.29* | –0.62*** | –0.45*** | 50 |
| Сессия 2 | 0.43** | –0.47*** | –0.51*** | 0.42** | –0.51*** | –0.48*** | 51 |
| Сессия 3 | 0.61*** | –0.12 | –0.62*** | 0.69*** | –0.08 | –0.61*** | 31 |
| Сессия 4 | 0.58*** | –0.12 | –0.66*** | 0.52** | –0.20 | –0.66*** | 31 |
| Сессия 1–4 | 0.38*** | –0.36*** | –0.52*** | 0.35*** | –0.41*** | –0.51*** | 163 |
Изменения в зависимости от возраста у учащихся параметра Ps показаны на рис. 4. Аппроксимация динамики Ps логарифмической функцией демонстрирует тенденцию к уменьшению с возрастом у школьников значения Ps, более выраженную в сессиях 1 и 2, чем в сессиях 3 и 4, что подтверждается оценками коэффициентов корреляции между Ps и возрастом (табл. 3). Эти изменения характеризуют повышение уровня пространственной связности в ЭЭГ у учащихся от младшего к старшему возрасту. Возрастная динамика параметра Ps в каждой сессии исследования для левого полушария мозга соответствует таковой для правого полушария.
Рис. 4.
Оценка возрастной динамики параметра Ps, рассчитанного по СФ1 ЭЭГ для левого (а) и правого (б) полушария мозга по 4 выборкам школьников в 4 сессиях исследования. По вертикальной оси – значение Ps в отн. ед. Обозначения как на рис. 3.
Fig. 4. Evaluation of the age dynamics of the Ps parameter, calculated by the EEG SF1 for the left (a) and the right (б) hemispheres of the brain according to 4 samples of the schoolchildren at 4 sessions stages of the study. Vertical axis – Ps value in relative units. Designations are the same as for fig. 3.
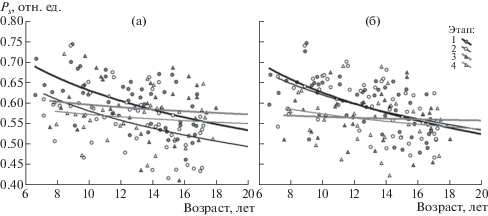
Численные значения параметра Ps для ЭЭГ левого и правого полушария мозга значимо не различались в первой и четвертой сессии исследований. Во второй сессии величина параметра Ps для левого полушария в среднем была значимо меньше, чем для правого (t(50) = –6.82, р < 0.001), в третьей, напротив, величина параметра Ps для левого полушария в среднем была значимо больше, чем для правого (t(30) = 2.55, р < 0.02).
Величина показателя GV также демонстрировала тенденцию к уменьшению с возрастом у учащихся. Судя по численным значениям коэффициентов корреляции, такое снижение было относительно более выражено для 3 и 4 сессий, чем для 1 и 2 сессий исследования (табл. 3).
Численные значения показателя GV в первой и второй сессии исследования для ЭЭГ левого полушария мозга были меньше, чем для правого (t(49) = –2.62, р = 0.012; t(50) = –9.19, р < 0.001) соответственно). В третьей сессии величина GV для ЭЭГ в левом полушарии мозга была выше, чем в правом (t(30) = –2.55, р = 0.019), тогда как в четвертой сессии билатеральные различия величины GV были статистически незначимы (t(30) = 1.90).
Поскольку исследования в сессиях 1 и 3 проведены в октябре месяце, а в сессиях 2 и 4 – в апреле и марте соответственно, и практически на одном и том же контингенте с разницей в 6 (сессии 1 и 2) и 5 (сессии 3 и 4) месяцев, мы оценили эффект влияния сезона (осень-весна) на 4 анализируемых интегральных параметрах. Эффект оценивали методом сравнения зависимых переменных, т.е. сопоставляли значения интегральных параметров для тех школьников, которые приняли участие и в осеннем, и в весеннем исследовании. На этапе 1 такое сопоставление было проведено у 50 школьников, на этапе 2 – у 23 школьников.
В целом по всей группе учащихся значимых сезонных изменений частоты основного ритма ЭЭГ, оцениваемой по данным спектрального анализа СФ, не выявлено: t(49) = = 1.07, р = 0.29 для сессий 1 и 2; t(22) = –0.39, р = 0.69 для сессий 3 и 4.
В табл. 4 даны оценки сезонных изменений трех других интегральных параметров ЭЭГ. Величина параметра Pt в целом по группе учащихся в сессии 1 была меньше, чем в сессии 2, что характеризовало более высокий уровень временнóй связности ЭЭГ-процессов во время осеннего сезона в сравнении с последующим весенним, как для левого, так и для правого полушария мозга. Величина параметра GV, напротив, в сессии 1 была больше, чем в сессии 2 для ЭЭГ в обоих полушариях мозга, что отражало более высокие значения интегральной амплитуды ЭЭГ во время осеннего сезона в сравнении с последующим весенним, как для левого, так и для правого полушария мозга. Величина параметра Ps для ЭЭГ-процессов в правом полушарии мозга в обеих сессиях исследования значимо не различалась. Для ЭЭГ-процессов в отведениях левого полушария величина параметра Ps была выше в сессии 1 в сравнении с сессией 2, что характеризовало больший уровень пространственной связности колебаний потенциалов мозга в период весеннего сезона в сравнении с осенним сезоном.
Таблица 4.
Значение t-критерия (зависимые выборки) и оценки значимости различий при сравнении интегральных параметров ЭЭГ, рассчитанных для левого и правого полушария мозга, в осеннем (сессия 1, 3) и весеннем (сессия 2, 4) сезонах Table 4. The value of the t-test (dependent samples) and the assessment of the significance of differences when comparing the integral EEG parameters calculated for the left and the right hemispheres of the brain in the autumn (session 1, 3) and spring (session 2, 4) seasons
| Сессии | Левое полушарие, параметр | Правое полушарие, параметр | n | ||||
|---|---|---|---|---|---|---|---|
| Pt | Ps | GV | Pt | Ps | GV | ||
| 1 и 2 | –5.43* | 7.11* | 11.05* | –6.38* | 0.58 | 7.77* | 50 |
| 3 и 4 | –0.54 | 1.98 | 1.17 | 0.39 | 0.22 | 0.98 | 23 |
Сопоставление значений интегральных параметров ЭЭГ у 23 школьников, которые участвовали в исследованиях в сессиях 3 и 4 на этапе 2, не выявило значимых различий в величинах параметров при первой (осенней) и второй (весенней) регистрации ЭЭГ как для левого, так и для правого полушария мозга.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Мы хотим отметить две важные особенности нашего подхода к анализу биоэлектрической активности мозга. Первая из них состоит в использовании интегральных показателей, расчет которых основан на построении СФ1 многоканальной ЭЭГ и которые представляют собой обобщенные оценки по всей совокупности ЭЭГ-процессов, регистрируемых в различных отведениях. Это позволяло оценивать динамику функционального созревания мозга как целого, без подробного описания локальных изменений, которые могут протекать гетерохронно в различных областях мозга.
Вторая особенность нашего подхода состояла в одновременной раздельной оценке пространственного и временнóго параметра, расчет которых базировался на одной и той же основе – СФ1 многоканальной ЭЭГ.
Широко используемые в онтогенетических исследованиях созревания ЦНС и становления когнитивных функций оценки значений взаимной когерентности или коэффициентов кросс-корреляции, характеризующие определенные стороны пространственной организации ЭЭГ, не содержат информации о временн${\text{ы}}'$х параметрах организации ЭЭГ-процессов, кроме ссылки на длительность эпохи анализа. Временнáя составляющая появляется только при сопоставлении этих оценок в последовательных эпохах анализа.
Отчасти дефицит подходов к изучению организации временнóй упорядоченности ЭЭГ восполняется методами анализа вероятностей переходов волновых компонентов одних диапазонов частот ЭЭГ в те же или другие диапазоны (“алгоритмов взаимодействия” между основными “ритмами” ЭЭГ) (Сороко и др., 2012; Soroko et al., 2015) или анализа длительности и вероятности изменения “микросостояний мозга”, характеризующихся определенным паттерном топического распределения колебаний потенциалов (Khanna et al., 2015; Michel, Koenig, 2018).
Мы предлагаем параметр Pt в качестве меры оценки временнóго масштаба упорядоченности ЭЭГ-процессов. Если согласиться с мнением, что для описания нейродинамики требуется целый набор характеристических временн${\text{ы}}'$х шкал, от миллисекунд до суток и более (Koenig et al., 2005; Harris, Gordon, 2015; Khambhatiab et al., 2018), то Pt будет принадлежать масштабу миллисекундного диапазона.
Полученные нами результаты демонстрируют тенденцию к увеличению значения параметра Pt у учащихся в зависимости от возраста. С учетом заданной шкалы изменений Pt (0, 1) такой рост означает, что чем старше ребенок, тем менее детерминированными во времени становятся ЭЭГ-процессы. Или, иными словами, с возрастом у школьников уменьшается временнóй масштаб связности, инерционность ЭЭГ-процессов.
Нами (Rozhkov et al., 2018; Рожков и др., 2019) было высказано предположение, что имеется содержательная связь между оценками инерционности – физиологической лабильности в смысле классической физиологии – и мерой временнóй связности ЭЭГ-процессов. Интервал времени до достижения СФ1 своей горизонтальной асимптоты, угол подъема начального сегмента СФ1, величина смещения СФ1 на заданном (минимальном) интервале (используемая в расчете Pt) могут характеризовать, как долго система удерживает след памяти о своем предыдущем состоянии, а следовательно, позволяют оценивать, насколько система инерционна.
А.А. Ухтомский определял лабильность как “скорость, с которой данный физиологический субстрат успевает переходить от состояния покоя к состоянию возбуждения и обратно … к состоянию физиологического покоя с готовностью к новой реакции”. При этом “интервал возбуждения (продолжительность состояния возбуждения) характеризует степень инерционности субстрата” (Ухтомский, 1951). Чем значение параметра Pt больше, тем выше функциональная подвижность (физиологическая лабильность) нервных процессов, и наоборот, чем оно меньше, тем меньше и уровень физиологической лабильности. Подтверждение обоснованности предположения о правомерности рассмотрения Pt как косвенной оценки параметра физиологической лабильности получено при анализе динамики Pt при исследовании воздействия на ЦНС острой гипоксии (Рожков и др., 2019). Так, для лиц с повышенной чувствительностью к дефициту кислорода, при пребывании в условиях острой гипоксии, на фоне снижения функционального состояния мозга, было характерно резкое уменьшение величины Pt, свидетельствующее о снижении уровня функциональной подвижности нервных процессов. Подобное снижение уровня физиологической лабильности ЦНС, повышение инерционности нервных процессов, могло служить основой замедления (впоследствии и нарушения) процессов восприятия, когнитивных функций и целенаправленной деятельности.
Соответственно, мы полагаем, что уменьшение временн${\text{ы}}'$х масштабов внутренней связности ЭЭГ-процессов (уменьшение величины интегрального временнóго параметра Pt) отражает повышение функциональной подвижности нейродинамических процессов у учащихся от младших к старшим классам, обеспечивая ускорение процессов восприятия и переработки информации, повышение скорости и точности умственной работы.
Изменения интегрального параметра Рs, являющегося мерой пространственной связности ЭЭГ-процессов, имели противоположную направленность, c возрастом величина параметра уменьшалась. Согласно своему определению, параметр Рs может характеризовать обобщенный уровень пространственной (дистантной) синхронизации колебаний потенциалов многоканальной ЭЭГ, уровень пространственной взаимосвязи ЭЭГ между всеми отведениями, поскольку величина Asf (значение асимптоты СФ1) связана с определителем выборочной корреляционной матрицы ЭЭГ (Trifonov, 2016). Предельные значения параметра Рs (0, 1) соответствуют абсолютной пространственной синхронизации (сугубо детерминированному процессу) и полному отсутствию пространственной синхронизации (“чисто” случайным процессам во всех отведениях). Сам термин “пространственная синхронизация” ввел М.Н. Ливанов (Ливанов, 1972). В зарубежной литературе в настоящее время используют понятие “функциональная связность” (functional connectivity) (Babiloni et al., 2020). Возрастание показателя пространственной синхронизации (функциональной связности) между ЭЭГ-процессами в различных отведениях ЭЭГ может отражать повышение согласованности в работе отдельных корковых зон (Ливанов, 1972; Цицерошин, Шеповальников, 2009; O’Neill et al., 2018).
Результаты нашей работы демонстрируют тенденцию к снижению величины параметра Рs с возрастом, что характеризует повышение уровня пространственной синхронизации ЭЭГ, функциональной связности ЭЭГ-процессов у учащихся от младших классов к старшим. Полученные данные о повышении уровня пространственной синхронизации ЭЭГ-процессов у детей и подростков согласуются с результатами кросс-корреляционного анализа ЭЭГ (Цицерошин, Шеповальников, 2009) и оценками интегральных показателей (VOL), характеризующих обобщенный уровень линейной взаимосвязи между колебаниями потенциалов в отведениях левого, правого полушария мозга, и в целом в совокупности всех отведений ЭЭГ (Сороко и др., 2012). Показано, что эта тенденция может сохраняться и в период юношества вплоть до достижения зрелого возраста (Кручинина и др., 2020). Данные анализа уровней когерентности отдельных гармонических составляющих в ЭЭГ, используемых при изучении топических особенностей формирования межполушарных и межрегиональных взаимосвязей ЭЭГ в левом и правом полушариях мозга, также показывают генеральную тенденцию к повышению уровня функциональной связности между ЭЭГ-процессами с возрастом у детей и подростков, по крайней мере в области частоты спектрального максимума основного ритма ЭЭГ (Мачинская и др., 2007; Gmehlin et al., 2011).
Топически-избирательное возрастание пространственной синхронизации (функциональной связности) между ЭЭГ-процессами в различных отведениях ЭЭГ может отражать повышение согласованности в работе отдельных корковых зон, обеспечивающих формирование функциональной системы для осуществления когнитивной или моторной деятельности (Ливанов, 1972; Цицерошин, Шеповальников, 2009; Stevens, 2016). Возрастание уровня пространственной синхронизации ЭЭГ-процессов в целом может быть связано с формированием состояния “оперативного покоя” (Ухтомский, 1951), мобилизационной готовности к ответной реакции, действию или деятельности.
Стабильную и однозначно воспроизводимую на обоих этапах исследования возрастную динамику показывал показатель “обобщенной дисперсии” GV, изменения которого говорили об уменьшении с возрастом у школьников общей “мощности” колебаний потенциалов. Обобщенная дисперсия является многомерным аналогом дисперсии, которая характеризует диапазон колебаний случайной величины. Она характеризует мощность переменной составляющей случайного процесса и является обобщающей характеристикой абсолютных размеров вариации амплитуды ЭЭГ-сигналов в совокупности отведений. Как известно по данным визуального и численного анализа, в раннем онтогенезе в целом амплитуда ЭЭГ растет, достигая в среднем наибольших значений у детей преддошкольного возраста, после чего снижается (Алферова, Фарбер, 1990). Такое снижение, от младшего школьного возраста к старшему, отражается в суммарных показателях спектра мощности колебаний потенциалов (Gasser et al., 1988), более выраженном для низкочастотного (дельта и тета) ЭЭГ-диапазона (Алферова, Фарбер, 1990; Демин и др. 2013). Это снижение мощности ЭЭГ-процессов может быть связано с уменьшением объема серого вещества мозга, выявляемого в МРТ-исследованиях у детей начиная с 10-летнего возраста (Giedd, 2004). Уменьшение массы нервных элементов и сокращение синаптической базы процессов локальной синхронизации в ЭЭГ может обусловливать снижение мощности электрогенеза коры, а приходящееся на этот же возрастной период постепенное уменьшение величины гемодинамической реакции (BOLD-сигнала на фМРТ) (Murphy et al., 1996) может отражать тенденцию к оптимизации уровня метаболических затрат и снижению энергетического “запроса” для обеспечения деятельности ЦНС (Lüchinger et al., 2012).
Одним из наиболее ярких показателей созревания мозга в детском возрасте является увеличение частоты основного ритма ЭЭГ. Полагают, что для определения соответствия уровня зрелости ЭЭГ возрасту лучше всего подходят оценки основного ритма ЭЭГ в затылочных зонах (Gasser et al., 1988; Takagaki et al., 2015), однако при этом приходится принимать во внимание также гетерохронность формирования ЭЭГ в других отделах коры мозга (Алферова, Фарбер, 1990; Горбачевская, Кожушко, 1990; Vijayakumar et al., 2018). В нашем исследовании использована обобщенная оценка частоты основного ритма, которую делали на основе расчета спектра СФ1. Структурная функция в нашем случае характеризует многоканальную ЭЭГ, и мы не можем привести аналитической формулы, связывающей эту обобщенную оценку с оценками частоты в каждом конкретном отведении. Квартильные (25–75%) границы от 8.8 до 9.9 Гц вполне соответствовали нормативным оценкам частоты основного ритма у детей и подростков школьного возраста, хотя у 8 школьников доминирующая частота ЭЭГ на том или ином этапе обследования была менее 7 Гц. По результатам проведенного исследования на одной (3-й) сессии обнаружилась слабая статистическая связь между частотой основного ритма и возрастом, для остальных трех сессий такой связи выявлено не было. Имеются сведения о тенденции к увеличению частоты основного ритма ЭЭГ у детей и подростков в школьном возрасте (Gasser et al., 1988), мы также отмечали данную тенденцию при обследовании детей на севере Магаданской области (Сороко и др., 2012). Между тем, по данным (Алферова, Фарбер, 1990; Горбачевская, Кожушко, 1990), частота основного ритма в целом достигает индивидуальных дефинитивных значений у детей в возрасте 11–13 лет, после чего либо стабилизируется на достигнутом уровне, либо может даже снижаться в середине пубертатного периода, вновь возрастая к его окончанию (Комкова, 2018).
При изучении физиологических функций организма и поведения человека невозможно обойти вниманием вопрос полового диморфизма (Etchell et al., 2019). Отличия в морфологии головного мозга у лиц разного пола (Murphy et al., 1996) находят отражение и в организации биоэлектрической активности мозга (Clarke et al., 2004, Комкова, 2018, Kurgansky et al., 2020). На основании результатов метаанализа работ различных научных коллективов за последние 40 лет, в которых сопоставлялись данные нейровизуализационных методов исследования межполушарной асимметрии и оценки половых различий когнитивных процессов, был сделан вывод, что половые различия несомненно обнаруживаются, они устойчивы, но сравнительно невелики по численным значениям оцениваемых параметров (Hirnstein et al., 2019).
Нам не удалось выявить статистически значимого влияния фактора Пол на величины интегральных параметров Pt, Ps и GV. Впрочем, окончательно не решен и вопрос о возрасте, в котором проявляются различия между мальчиками и девочками как при осуществлении той или иной когнитивной деятельности, так и в показателях нейродинамических процессов. Имеются основания полагать, что критическим для формирования таких различий является период полового созревания (Безруких и др., 2009; Комкова, 2018; Кручинина и др., 2020). В подростковом возрасте, по данным МРТ-исследований, время созревания различных мозговых структур у девушек опережает в среднем на 1 год (Gedd, 2004), формирование организации ЭЭГ-процессов – на 1–2 года (Алферова, Фарбер, 1990; Горбачевская, Кожушко, 1990). В исследовании, охватывающем возрастной диапазон от 8 до 30 лет, показано, что в состоянии спокойного бодрствования связанные с полом различия в изменениях уровня пространственной синхронизации ЭЭГ сравнительно невелики, но начинают значимо проявляться при деятельности, причем гетерохронно, например, при восприятии текста на слух с 12 лет, а при чтении текста – с 15 лет (Кручинина и др., 2020).
На количественные оценки динамики функционального созревания ЦНС, выраженности половых различий в ЭЭГ в конкретной группе обследуемых существенное влияние оказывает межиндивидуальная вариабельность изучаемых параметров. Так, численные значения статистической связи параметров Pt, Ps и GV с возрастом (см. табл. 3) отличаются для 1-го и 2-го этапов исследования. По существу на этих двух этапах мы имели дело с двумя различными, хотя и частично пересекающимися выборками детей и подростков. Применение процедуры нормирования, ориентация на интегральные, оцениваемые по всей совокупности ЭЭГ-процессов, параметры в известной степени позволяют уменьшить выборочную дисперсию оценок. Однако причина вариабельности заложена уже изначально в генетическом коде, определяющем индивидуальные особенности ЭЭГ и задающем программу ее формирования (Безруких и др., 2009).
Реализация этой программы, формирование физиологических функций и развитие организма в целом происходит при тесном взаимодействии организма и среды. Процесс развития на всех этапах онтогенеза носит адаптивный приспособительный характер, который определяется двумя важнейшими факторами: морфофункциональной зрелостью физиологических систем и адекватностью функциональных возможностей организма условиям внешней среды.
Сочетанное воздействие близких к экстремальным природных, социально-экономических и бытовых условий жизни на Севере вызывает не только функциональные отклонения, связанные с напряжением регуляторных процессов, но может также и обусловливать замедление темпов “созревания” электрогенеза мозга. Исследования, проведенные в Архангельской области, показали, что у 53% детей и подростков школьного возраста выявляются признаки незрелости (в сравнении с возрастными нормативами) электрической активности мозга в виде неустойчивости основного ритма, повышенного содержания медленноволновых (тета- и дельта-) составляющих ЭЭГ, эпизодов пароксизмальных проявлений физиологической активности. При этом у 29% учащихся наблюдается отставание в темпах формирования частотного спектра ЭЭГ на 1.5–2 года по сравнению со сверстниками из средней полосы России (Сороко и др., 2005). Отмечен своеобразный “широтный” эффект в степени выраженности признаков незрелости ЦНС: по оценкам амплитуды и индекса активности в основных диапазонах частот, по темпам созревания ЭЭГ подростки Заполярья отстают от сверстников из Приполярного района, что может быть обусловлено более суровыми условиями жизни в Заполярье (Демин и др., 2013).
Непосредственный эффект влияния факторов среды (точнее, их комплексных сезонных перестроек) на функциональное состояние ЦНС у школьников-северян в нашем исследовании проявился в динамике значений параметров Pt, Ps и GV при их сопоставлении в осенней и весенней сессиях. Значения параметра Pt были выше, а параметров GV и Ps были ниже в весенний период в сравнении с осенним. Для параметров Pt и GV эти изменения были выражены в сравнительно равной степени для левого и правого полушария мозга. В динамике параметра Ps выявлен эффект билатеральной асимметрии. Уменьшение в весеннем периоде величины Ps, характеризующее повышение пространственной связности процессов, уровня дистантной синхронизации ЭЭГ, выявлено в левом и не выявлено в правом полушарии мозга. В работе В.В. Аршавского и соавт. (Аршавский и др., 1989) было показано значимое влияние процесса адаптации к сложным природным условиям Севера на характер и выраженность функциональной асимметрии мозга, что нашло отражение в оценках пространственной синхронизации биопотенциалов в покое и при нагрузках, адресованных преимущественно правой или левой гемисфере.
Изменения параметра GV – обобщенной дисперсии, характеризующей абсолютные размеры вариации амплитуды сигналов в многоканальной ЭЭГ, свидетельствовали об уменьшении в весенний период в сравнении с предыдущим осенним периодом величины интегральной амплитуды ЭЭГ. Более высокие значения параметра GV объяснимы значимо большей представленностью в осенний период в спектрах ЭЭГ частот медленноволнового – дельта- и тета-диапазона, что отмечено также в исследованиях Грибанова и соавт. (Грибанов и др., 2016б). Авторы этой работы обращают внимание на взаимосвязь динамики спектральных характеристик ЭЭГ с периодами естественной освещенности и полагают, что изменение продолжительности светового дня является одним из наиболее значимых факторов, влияющих на формирование биоэлектрической активности мозга у школьников-северян.
Действительно, два сезона исследований различаются противоположным характером фотопериодизма, в конце октября значительно больше продолжительность темного времени суток, в начале апреля – светлого, и имеется взаимосвязь между показателями гормонального статуса человека с длительностью светового дня (Кубасов и др., 2006). При этом изменяющийся гормональный фон создает предпосылки для развития процесса акклиматизации к изменениям температурного режима (Евдокимов и др., 2007). В весенний период по сравнению с осенним у школьников-северян в значимо большем числе случаев выявляются отклонения в ЭЭГ, свидетельствующие о дисбалансе подкорково-корковых регуляторных влияний, предположительно вследствие чрезмерного напряжения функциональных систем организма в процессе адаптации к условиям северной зимы (Rozhkov et al., 2018).
Возникает сложная проблема взаимосвязи не только факторов температуры и естественной освещенности, но также и погодных условий, состояния геомагнитного поля, оценки их раздельного и сочетанного влияния на развитие ЦНС у детей и подростков, подходы к решению которой еще только разрабатываются.
ЗАКЛЮЧЕНИЕ
Для оценки динамики возрастных изменений биоэлектрической активности мозга при изучении формирования ЦНС у детей и подростков мы ориентировались на применение интегральных показателей, являющихся обобщенными характеристиками ЭЭГ-процессов в совокупности отведений как левого, так и правого полушарий, а также конвекситальной поверхности мозга в целом. Эти параметры рассчитывались на основе структурной функции многоканальной ЭЭГ так, чтобы единый пространственно-временнóй процесс разделить на 2 составляющие, являющиеся отдельной мерой масштаба временнóй (Pt) и мерой пространственной (Ps) связности ЭЭГ-процессов. В качестве дополнительных параметров использовалась частота основного ритма, рассчитываемая по максимуму спектра мощности структурной функции, и показатель GV (general variability) как многомерный аналог дисперсии ЭЭГ-процесса, оцениваемый по совокупности ЭЭГ-процессов.
Полученные нами результаты демонстрируют тенденцию к увеличению значения параметра Pt у учащихся в зависимости от возраста. С учетом заданной шкалы изменений Pt (0, 1) такой рост означает, что чем старше ребенок, тем менее детерминированными во времени становятся ЭЭГ-процессы, тем меньше временнóй масштаб связности и, соответственно, инерционность ЭЭГ-процессов. По нашему мнению, уменьшение величины интегрального параметра Pt, уменьшение временн${\text{ы}}'$х масштабов внутренней связности ЭЭГ-процессов отражает повышение функциональной подвижности нейродинамических процессов у учащихся от младших к старшим классам, обеспечивая ускорение процессов восприятия и переработки информации, повышение скорости и точности умственной работы. Численные значения параметра Pt для левого и правого полушария мозга значимо не различались во всех возрастных группах школьников, что объясняется строгой сонастроенностью временных масштабов связности ЭЭГ-процессов обоих полушарий мозга для совместной работы.
Изменения интегрального параметра Рs, являющегося мерой пространственной связности ЭЭГ-процессов, имеют противоположную направленность, c возрастом величина параметра уменьшается. Предельные значения параметра Рs (0, 1) соответствуют абсолютной пространственной синхронизации (сугубо детерминированному процессу) и полному отсутствию пространственной синхронизации (“чисто” случайным процессам во всех отведениях). Возрастание уровня пространственной синхронизации ЭЭГ-процессов в целом отражает повышение согласованности в работе отдельных корковых зон, усиление функционального взаимодействия между ними и может быть связано с формированием состояния “оперативного покоя” (по А.А. Ухтомскому) как мобилизационной готовности к ответной реакции, действию или деятельности.
Изменения параметра GV – обобщенной дисперсии, характеризующей абсолютные размеры вариации амплитуды сигналов в многоканальной ЭЭГ, свидетельствовали об уменьшении с возрастом у школьников общей совокупной “мощности” ЭЭГ-процессов в различных отведениях. Сопоставление с нейроморфологическими данными позволяет предположить, что такое снижение мощности ЭЭГ-процессов, электрогенеза коры, может быть связано с уменьшением объема серого вещества мозга, выявляемого при МРТ-исследованиях у детей начиная с 10-летнего возраста.
Выявлены вариации интегральных параметров ЭЭГ, отражающие влияние сезонных изменений природно-климатических факторов и связанные с процессами акклиматизации у детей и подростков-северян. Значения интегрального временного параметра Pt увеличивались, тогда как значения GV – обобщенной характеристики мощности ЭЭГ-процесса – и интегрального пространственного параметра Ps уменьшались в весенний период в сравнении с осенним. В качестве одного из факторов, вызывающих эти перестройки в ЭЭГ, можно рассматривать контрастные изменения фотопериодизма в осеннее и весеннее время. При этом возникает сложная проблема взаимосвязи целого ряда факторов – освещенности, холодового фактора, с которым связаны процессы акклиматизации к суровым условиям северной зимы, и других, требующая разработки подходов к оценке раздельного и сочетанного влияния этих факторов на развитие ЦНС у детей и подростков-северян.
Работа поддержана Госзаданием № Гос. рег. ИСГЗ AAAA-A18-118012290142-9.
Список литературы
Алферова В.В., Фарбер Д.А. Отражение возрастных особенностей функциональной организации мозга в электроэнцефалограмме покоя. В кн.: Структурно-функциональная организация развивающегося мозга. 1990. Л. Наука. С. 45–65.
Аршавский В.В., Гельфгат Е.Л., Ротенберг В.С., Соловенчук Л.Л. Межполушарная асимметрия как фактор адаптации человека в условиях Севера. Физиология человека. 1989. 15 (5): 142–147.
Барвинок А.И., Рожков В.П. Особенности межцентральной координации корковых электрических процессов при умственной деятельности. Физиология человека. 1992. 18 (3): 5–16.
Бойко Е.Р. Физиолого-биохимические основы жизнедеятельности человека на Севере. Екатеринбург. УрО РАН. 2005. 190 с.
Безруких М.М., Мачинская Р.И., Фарбер Д.А. Структурно-функциональная организация развивающегося мозга и формирование познавательной деятельности в онтогенезе ребенка. Физиология человека. 2009. 35 (6): 10–24.
Гаврон А.А., Yacila Isabela Deza Araujo, Шарова Е.В., Смирнов А.С., Князев Г.Г., Челяпина М.В., Фадеева Л.М., Абдулаев А.А., Куликов М.А., Жаворонкова Л.А., Болдырева Г.Н., Верхлютов В.М., Пронин И.Н. Групповой и индивидуальный фМРТ-анализ основных сетей покоя здоровых испытуемых. Журнал высшей нервной деятельности им. И.П. Павлова. 2019. 69 (2): 150–163.
Горбачевская Н.Л., Кожушко Л.Ф. Динамика формирования ЭЭГ у мальчиков и девочек школьного возраста (по данным 9-летнего наблюдения). Журн. невропатол. 1990. 90 (8): 75–79.
Грибанов А.В., Гудков А.Б., Попова О.Н., Крайнова И.Н. Кровообращение и дыхание у школьников в циркумполярных условиях. Архангельск: САФУ. 2016а. 270 с.
Грибанов А.В., Джос Ю.С., Багрецова Т.В., Бирюков И.С. Фотопериодизм и изменения биоэлектрической активности головного мозга у школьников арктической зоны. Физиология человека. 2016б. 42 (2): 16–26.
Демин Б.Д., Поскотинова Л.В., Кривоногова Е.В. Варианты возрастного формирования структуры ЭЭГ подростков Приполярных и Заполярных районов Европейского Севера. Вестник Северного (Арктического) федерального университета. 2013. 1: 41–45.
Дзугаева С.Б. Проводящие пути головного мозга человека: в онтогенезе. М: Медицина, 1975. 253 с.
Евдокимов В.Г., Рогачевская О.В., Варламова Н.Г. Модулирующее влияние факторов Севера на кардиореспираторную систему человека в онтогенезе. Екатеринбург. УРО РАН. 2007. 257 с.
Комкова Ю.Н. Электрическая активность головного мозга у детей и подростков на разных стадиях полового созревания в состоянии покоя. Новые исследования. 2018. 1 (54): 4–25.
Кручинина О.В., Станкова Е.П., Гальперина Е.И. Возрастные особенности пространственно-временнóй организации ЭЭГ у испытуемых 8–30 лет мужского и женского пола при восприятии устных и письменных текстов. Физиология человека. 2020. 46 (3): 15–28.
Кубасов Р.В., Демин Д.Б., Ткачев А.В. Адаптивные реакции эндокринной системы у детей, проживающих в условиях контрастной фотопериодики. Физиология человека. 2006. 32 (4): 89–96.
Кулаичев А.П. Об информативности когерентного анализа в исследованиях ЭЭГ. Журнал высшей нервной деятельности им. И.П. Павлова. 2009. 59 (6): 766–775.
Лебедева Н.Н., Каримова Е.Д. Устойчивость паттернов ЭЭГ человека в различных задачах: проблема аутентификации личности. Журнал высшей нервной деятельности им. И.П. Павлова. 2020. 70 (1): 40–49.
Ливанов М.Н. Пространственная организация процессов головного мозга. М.: Наука, 1972. 181 с.
Мачинская Р.И., Курганский А.В., Ломакин Д.И. Возрастные изменения функциональной организации корковых звеньев регуляторных систем мозга у подростков. Анализ нейронных сетей покоя в пространстве источников. Физиология человека. 2019. 5 (45): 5–19.
Мачинская Р.И., Соколова Л.С., Крупская Е.В. Формирование функциональной организации коры больших полушарий в покое у детей младшего школьного возраста с различной степенью зрелости регуляторных систем мозга. Сообщение II. Анализ когерентности а-ритма ЭЭГ. Физиология человека. 2007. 33 (2): 5–15.
Рожков В.П., Трифонов М.И., Бурых Э.А., Сороко С.И. Оценка индивидуальной устойчивости человека к острой гипоксии по интегральным характеристикам структурной функции многоканальной ЭЭГ. Росс. физиол. журн. им. И.М. Сеченова. 2019. 105 (7): 832–852.
Сороко С.И., Бекшаев С.С., Рожков В.П. ЭЭГ корреляты генофенотипических особенностей возрастного развития мозга у детей аборигенного и пришлого населения северо-востока России. Росс. физиол. журн. им. И.М. Сеченова. 2012. 98 (1): 3–26.
Сороко С.И., Бурых Э.А., Сидоренко Г.В. Особенности возрастного развития мозга у детей в условиях Севера. Росс. физиол. журн. им. И.М. Сеченова. 2005. 91 (7): 729–739.
Ухтомский А.А. Собрание сочинений. Т. II. Парабиоз, физиологическая лабильность, усвоение ритма. Л. Изд-во Ленинградского государственного университета. 1951. 180 с.
Цицерошин М.Н., Шеповальников А.Н. Становление интегративной функции мозга. СПб.: Наука. 2009. 249 с.
Anderson A.J., Perone S. Developmental change in the resting state electroencephalogram: Insights into cognition and the brain. Brain Cogn. 2018. 126: 40–52.
Babiloni C., Barry R.J., Basar E., Blinowska K.J., Andrzej Cichocki A., Drinkenburg W. H.I.M., Klimesch W., Knight R.T., Lopes da Silva F., Nunez P., Oostenveld R., Jeong J., Pascual-Marqui R., Valdes-Sosa P., Hallett M. International Federation of Clinical Neurophysiology (IFCN) – EEG research workgroup: Recommendations on frequency and topographic analysis of resting state EEG rhythms. Part 1: Applications in clinical research studies. Clin. Neurophysiol. 2020. 131 (1): 285–307.
Barry R.J., Clarke A.R., McCarthy R., Selikowitz M., Johnstone S.J., Rushby J.A. Age and gender effects in EEG coherence: I. Developmental trends in normal children. Clin. Neurophysiol. 2004. 115 (10): 2252–2258.
Cekic S., Grandjean D., Renaud O. Time, frequency, and time-varying Granger-causality measures in neuroscience. Stat. Med. 2018. 37 (11): 1910–1931.
Etchell A., Adhikari A., Weinberg L.S., Choo A.L., Garnett E.O., Chow H.M., Chang S.E. A systematic literature review of sex differences in childhood language and brain development. Neuropsychologia. 2018. 114: 19–31.
Gasser T., Verleger R., Bacher P., Sroka L. Development of the EEG of school-age children and adolescents: I, analysis of band power. Electroenceph. Clin. Neurophysiol. 1988. 69: 91–99.
Giedd J.N. Structural magnetic resonance imaging of the adolescent brain/ Ann. N. Y. Acad. Sci. 2004. 1021: 77–85.
Gmehlin D., Thomas C., Weisbrod M., Walther S., Resch F., Oelkers-Ax R. Development of brain synchronisation within school-age–individual analysis of resting (α) coherence in a longitudinal data set. Clin. Neurophysiol. 2011. V. 122. № 10. P. 1973–1983.
Harris A.Z., Gordon J.A. Long-range neural synchrony in behavior. Annu. Rev. Neurosci. 2015. 38: 171–194.
Hirnstein M., Hugdahl K., Hausmann M. Cognitive sex differences and hemispheric asymmetry: A critical review of 40 years of research. Laterality. 2019. 24 (2): 204–252.
Jiruška P., Prokš J., Drbal O., Sovka P., Marusic P., Mares P. Comparison of Different Methods of Time Shift Measurement in EEG. Physiol. Res. 2005. 54 (4): 459–465.
Khanna A., Pascual-Leone A., Michel C.M., Farzan F. Microstates in Resting-State EEG: Current Status and Future Directions. Neurosci. Biobehav. Rev. 2015. 49: 105–113.
Khambhatiab A.N., Sizemorea A.E., Betzela R.F., Bassett D.S. Modeling and interpreting mesoscale network dynamics. NeuroImage. 2018. 180 (Part B): 337–349.
Koenig T., Studer D., Hubl D., Melie L., Strik W.K. Brain connectivity at different time-scales measured with EEG. Phil. Trans. R. Soc. B. Biol. Sci. 2005. 360 (1457): 1015–1023.
Kurgansky A.V., Lomakin D.I., Machinskaya R.I. Resting-state networks in adolescents with poor behavior regulation. an analysis of effective cortical connectivity in EEG source space. Журнал высшей нервной деятельности. 2020. 70 (6): 723–737.
Lüchinger R., Michels L., Martin E., Brandeis D. Brain state regulation during normal development: Intrinsic activity fluctuations in simultaneous EEG–fMRI. NeuroImage. 2012. 60: 1426–1439.
Michel Ch.M., Koenig T. EEG microstates as a tool for studying the temporal dynamics of whole-brain neuronal networks: A review. Neuroimage. 2018. 180 (Pt B): 577–593.
Murphy D.G., DeCarli C., McIntosh A.R., Daly E., Mentis M.J., Pietrini P., Szczepanik J., Schapiro M.B., Grady C.L., Horwitz B., Rapoport S.I. Sex differences in human brain morphometry and metabolism: an in vivo quantitative magnetic resonance imaging and positron emission tomography study on the effect of aging. Arch. Gen. Psychiatry. 1996. 53 (7): 585–594.
O’Neill G.C., Tewarie P., Vidaurre D., Liuzzi L., Woolrich M.W., Brookes M.J. Dynamics of large-scale electrophysiological networks: A technical review. Neuroimage. 2018. 180 (Pt B): 559–576.
Rozhkov V.P., Trifonov M.I., Soroko S.I. Study of Brain Bioelectrical Activity Maturation in Northern Schoolchildren with Integral Parameters of Multichannel EEG. Human Physiology 2018. 44 (6): 617–626.
Seghier M.L., Price C.J. Interpreting and Utilising Intersubject Variability in Brain Function. Trends Cogn. Sci. 2018. 22 (6): 517–530.
Seth A.K., Barrett A.B., Barnett L. Granger causality analysis in neuroscience and neuroimaging. J. Neurosci. 2015. 35 (8): 3293–3297.
Snyder A.C., Morais M.J., Willis C.M., Smith M.A. Global network influences on local functional connectivity. Nat. Neurosci. 2015. 18 (5): 736–743.
Soroko S.I., Bekshaev S.S., Rozhkov V.P., Nagornova Zh.V., Shemyakina N.V. General Features of the Formation of EEG Wave Structure in Children and Adolescents Living in Northern Europe. Human Physiology. 2015. 41 (4): 394–403.
Soroko S.I., Nagornova Zh.V., Rozhkov V.P., Shemyakina N.V. Age-Specific Characteristics of EEG Coherence in Children and Adolescents Living in the European North of Russia. Human Physiology. 2015. 41 (5): 517–531.
Stevens M.C. The contributions of resting state and task based functional connectivity studies to our understanding of adolescent brain network maturation. Neurosci. Biobehav. Rev. 2016. 70: 13–32.
Stokes P.A., Purdon P.L. A study of problems encountered in Granger causality analysis from a neuroscience perspective. PNAS. 2017. 114 (34): E7063–E7072.
Takagaki K., Russell J., Lippert M.T., Motamedi Gh.K. Development of the posterior basic rhythm in children with autism. Clinical Neurophysiology. 2015. 126 (2): 297–303.
Thatcher R.W., North D.M., Biver C.J. Development of cortical connections as measured by EEG coherence and phase delays. Hum. Brain Mapp. 2008. 29 (12): 1400–1415.
Trifonov M. The structure function as new integral measure of spatial and temporal properties of multi-channel EEG. Brain informatics. 2016. 3 (4): 211–220.
Vijayakumar N., Op de Macks Z., Shirtcliff E.A., Pfeifer J.H. Puberty and the human brain: Insights into adolescent development. Neurosci. Biobehav. Rev. 2018. 92: 417–436.
Whitford T.J., Rennie C.J., Grieve S.M., Clark C.R., Gordon E., Williams L.M. Brain maturation in adolescence: concurrent changes in neuroanatomy and neurophysiology. Hum. Brain Mapp. 2007. 28 (3): 228–237.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова