

Журнал высшей нервной деятельности им. И.П. Павлова, 2021, T. 71, № 4, стр. 500-514
Поздние компоненты связанных с событиями потенциалов у детей, подростков и взрослых при чтении слов
Е. И. Гальперина 1, 2, *, О. В. Кручинина 1, 2, Е. П. Станкова 2, А. Н. Корнев 1
1 Санкт-Петербургский государственный педиатрический медицинский университет Минздрава РФ
Санкт-Петербург, Россия
2 Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН
Санкт-Петербург, Россия
* E-mail: galperina-e@yandex.ru
Поступила в редакцию 12.11.2020
После доработки 16.12.2020
Принята к публикации 22.12.2020
Аннотация
Известно, что различия при чтении повторяющихся/новых слов у взрослых отражаются в компонентах связанных с событием потенциалов (ССП) N400 и P600, однако вопрос о функциональном значении этих компонентов и возрастной траектории их развития все еще остается открытым. Целью нашего исследования было оценить амплитуду и латентность компонентов N400 и P600 в ответ на повторяющиеся и новые слова в трех возрастных группах: у детей предподросткового возраста (N = 25, 9–11 лет), подростков (N = 17, 12–14 лет) и взрослых (N = 22, 18–36 лет). Возрастные различия амплитуды были выявлены для компонента N400, усредненного по отведениям F3, Fz, F4, C3, Cz, C4, и компонента P600, усредненного по отведениям Р3, Pz, Р4, различий по латентности выявлено не было. Сравнение амплитуды усредненных ССП на повторяющиеся и новые слова выявило различия у всех возрастных групп: у детей во временном интервале 750–850 мс, у подростков во временном интервале 650–750 мс, а у взрослых – в интервале 600–800 мс. При этом во всех возрастных группах амплитуда Р600 на повторяющееся слово выше, чем на новое. Для N400 различия в ответ на повторяющееся и новое слова выявлены только в группе взрослых: латентность на новое слово меньше, чем на повторяющееся. По-видимому, процесс сопоставления новой и ранее предъявленной вербальной зрительной информации многостадийный. Эти стадии формируются на разных этапах онтогенеза и отражаются в разных компонентах ССП. Сначала проявляются различия между повторяющимся и новым словом по компоненту Р600: в 12–14 лет паттерн различий аналогичен таковому у взрослых. В то время как различия по компоненту N400 в детском и подростковом возрасте не выявляются и присутствуют только у взрослых.
ВВЕДЕНИЕ
Многочисленные исследования показали, что овладение элементарными навыками чтения (декодированием) происходит на протяжении первых 3–4 лет обучения (Maïonchi-Pino et al., 2009; Безруких, Крещенко, 2011; Корнев и др., 2019). Примерно начиная с 4–5-го года обучения у детей особенно активно формируются навыки смысловой обработки текста, что продолжается до конца подросткового возраста, а во многих случаях и в юношеском возрасте (Корнев, 2006). Одновременно в процессе созревания мозга происходит изменение объема и толщины серого вещества, площади поверхности коры больших полушарий мозга (Giedd et al., 1999, 2006, Wierenga et al., 2014; Ducharme et al., 2015; Mills et al., 2016; Vijayakumar et al., 2016), синаптический прунинг (Huttenlocher et al., 1982), а также усиление синаптической передачи (Singh, Su, 2013; Toffoletto, et al., 2014; Rossetti, et al., 2016). Происходящие изменения влияют на нейрофизиологическое обеспечение когнитивных процессов, отражаясь в изменениях амплитуды, латентности и длительности многих компонентов вызванных потенциалов (ВП) и связанных с событием потенциалов (ССП) (Holcomb, et al., 1992; Atchley et al., 2006; McNealy et al., 2009; Benau, 2011; Maurer et al., 2006; Сороко и др., 2018; Портнова и др., 2014). Процесс взросления, таким образом, представляет собой, с одной стороны, создание новых связей и новых функциональных систем, а, с другой стороны, некоторую реорганизацию уже существующих систем. Существуют свидетельства изменения стратегий выполнения одних и тех же когнитивных задач с возрастом (Castel et al., 2011; Seassau, Bucci, 2013). Важно понимать, что внешняя схожесть деятельности у детей и взрослых может иметь несколько отличающуюся системную организацию. Например, при чтении текста увеличивается вклад процессов антиципации и селективности у взрослых по сравнению с детьми (Federmeier, 2007). В нашей работе мы сравнивали нейрофизиологические механизмы чтения и дискриминации отдельных слов у детей младшего школьного возраста, подростков и взрослых по данным ССП. В эксперименте моделировалась ситуация, весьма близкая к той, что происходит в процессе чтения текста: чтение слов и сопоставление их с предыдущими прочитанными словами с опорой на рабочую память. Испытуемым в процесс чтения была поставлена задача сопоставления воспринимаемого слова с двумя только что прочитанными словами. Соответственно сложившимся представлениям об операциональном обеспечении чтения слов, этот процесс включает зрительное распознавание слов, актуализацию фонологической структуры слова и возможное (необязательное) соотнесение с ментальным лексиконом (Frost, 1998; Fiebach et al., 2002). Также в соответствии с дизайном эксперимента от испытуемого требовалось сравнение отдельно предъявленного слова с двумя ранее прочитанными словами и принятия решения о том, присутствовало ли данное слово среди первых двух.
Ранее было показано, что анализ формы слова начинается на 250-й мс от начала предъявления слова (Tarkiainen et al., 2002), а обращение к ментальному лексикону происходит позже. Компонент ССП N400 (250–500 мс) отражает манипуляции с лексическими переменными, такими как семантическая вероятность, лексическая частотность и лексическая близость, и наблюдается как у взрослых, так и у детей (Friederici, 2004, Crandall, 2010; Kutas, Federmeier, 2011). Компонент N400 может отражать общий процесс, вызванный потенциально значимым стимулом. Височные, префронтальные области и передняя поясная извилина интегрированы в распределенную кортикальную сеть и могут обеспечивать специализированные вклады различных факторов, в том числе таких как частота, повторение или семантические ассоциации, контекст и более широкий дискурс (Kutas, Federmeier, 2011). Эти процессы в ходе конструирования значения, которое наилучшим образом соответствует данному контексту, могут происходить как параллельно, так и запускать друг друга, т.е. быть взаимозависимыми.
Функциональное значение компонента Р600 весьма разнообразно, появление этого пика показано при включении в синтаксические конструкции различного рода ошибок, включая ошибки субкатегоризации, числа, времени, пола, падежной согласованности и др. (см. обзор Gouvea et al., 2010). Кроме того, компонент P600 связывают с таким процессом обработки, как реанализ (Friederici, 1995; Canseco-Gonzalez, 2000), а также повышенные потребности рабочей памяти при обработке сложных предложений, а амплитуда Р600 отражает “сложность синтаксической интеграции” (Kaan et al., 2000). Кроме того, Р600 связывают с глобальными процессами повторной обработки информации, когда ожидания, основанные на правилах, не соблюдаются (Schmidt-Kassow, Kotz, 2009).
В контексте сопоставления слов с ранее прочитанным, значение компонентов N400 и Р600 относят к таким процессам памяти, как узнавание и вспоминание (familiarity and recollection) (Mandler et al., 2010). Узнаванием считается субъективное переживание того, что стимул встречался раньше, при невозможности вспомнить какую-либо контекстную информацию относительно данного стимула. Вспоминание, напротив, позволяет восстановить качественную информацию о происходящем событии, например, где и когда оно произошло (Yonelinas, 2010). Узнавание ассоциируют с негативной волной, имеющей максимум во фронтальных отведениях, которая начинается примерно через 300 мс после предъявления стимула, а вспоминание – с позитивной волной, имеющей максимум в париетальных отведениях и начинающейся примерно через 500 мс после предъявления стимула (Woodruff et al., 2006). Таким образом, если процессы восприятия и сличения только что прочитанного и ранее предъявленного слова отличаются у детей, подростков и взрослых, то эти отличия вероятнее всего найдут свое отражение в изменениях параметров компонентов N400 с фронто-центральной локализацией и Р600 – с центро-париетальной. Примечательно, что достоверных различий в топографическом распределении, амплитуде и латентности P600 при сопоставлении детей 8–15 лет и взрослых не выявлено (Atchley et al., 2006), тогда как они описаны для компонента N400 (Holcomb et al., 1992; Friederici, 2006). Локализация N400 у детей не ограничивается затылочными и центро-париетальным отведениями, как у взрослых (Holcomb et al., 1992), а захватывает лобные области (Friederici, Hahne, 2001, Atchley et al., 2006). Эти данные позволили предположить, что различия компонентов N400 и P600, связанные с дискриминацией стимула, формируются в онтогенезе не одновременно. Цель работы – проверить эту гипотезу и оценить онтогенетические изменения различий компонентов N400 и P600 ССП при зрительном предъявлении новых и повторяющихся слов у детей, подростков и взрослых.
МЕТОДИКА
Испытуемые
В исследовании приняли участие дети 9–11 лет (средний возраст = 9.9 ± 0.7, N = 25, 10 мальчиков), подростки 12–14 лет (средний возраст = 12.9 ± 0.8, N = 17, 8 мальчиков) и взрослые (средний возраст 23.5 ± 5.7, N = 22, 7 мужчин). Дети и подростки обучались в государственных общеобразовательных школах, взрослые имели высшее образование или обучались в ВУЗах. Родным языком для всех испытуемых был русский. Родители (законные представители) несовершеннолетних участников исследования представили добровольное письменное информированное согласие, подписанное ими после разъяснения им потенциальных рисков и преимуществ, а также характера предстоящего исследования. Все испытуемые участвовали в исследованиях добровольно. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Комиссией по этике биомедицинских исследований ИЭФБ РАН.
Психологическое тестирование
Оценка невербального интеллекта производилась с использованием методики теста Кеттелла (Cattell, 1973).
Оценка показателей рабочей памяти производилась на основе субтеста на повторение цифровых рядов (в прямом и обратном порядке) методики Д. Векслера – тест WISC (Wechsler Intelligence Scale for Children) в адаптированной и стандартизированной версии (Филимоненко, Тимофеев, 1994). В качестве интегральной оценки приведен суммарный показатель результатов двух тестов.
Скорость чтения и качество понимания текста оценивали только у детей и подростков по методике СМИНЧ (Корнев, Ишимова, 2010). Методика СМИНЧ включает 2 текста, различающихся по уровню сложности, и позволяет оценить следующие параметры: скорость чтения, число ошибочно прочитанных слов, качество понимания текста, коэффициент техники чтения (КТЧ). Объем каждого текста около 200 слов. Понимание оценивалось по количеству верных ответов на 10 стандартных вопросов к фрагменту текста объемом в 95–110 слов. В соответствии с методикой для учащихся 3-го класса нормальная скорость чтения должна превышать 54 слова в мин, КТЧ больше 85; 4-го класса – 74 слова в мин, КТЧ больше 87, 5-го класса – 80 слов в мин, КТЧ больше 88, 6-го класса – 84 слова в мин, КТЧ больше 90.
Все вышеописанные тесты выполнялись с целью установить соответствие развития детей и подростков возрастной норме.
Стимульный материал
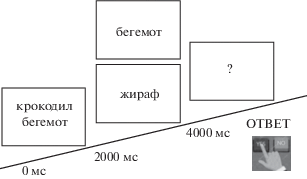
Тест чтения слов в парадигме дискриминационного выбора содержал три типа стимулов: первый стимул содержал 2 слова (существительных в именительном падеже, из 2–3 слогов, частотность в корпусе письменного русского языка (https://ruscorpora.ru/new/) не менее 10 ipm, например, “крокодил бегемот”), второй стимул – существительное в именительном падеже, которое могло повторять любое из первой пары (например, “крокодил”), либо быть новым (например, “верблюд”), третий стимул – знак вопроса, после появления которого испытуемый должен был дать ответ, нажав на соответствующую кнопку (рис. 1). Длительность каждого стимула составляла 2000 мс, интервал между 1-м и 2-м, 2-м и 3-м стимулами варьировал от 0 до 300 мс, таким образом, длительность пробы составляла от 6000 до 6600 мс. Данная парадигма исследования позволяет анализировать отдельно каждый этап чтения: 1) чтение первых двух слов; 2) чтение повторяющегося слова и 3) чтение нового слова.
Рис. 1.
Структура пробы и временная последовательность событий в тесте дискриминации слов.
Fig. 1. Structure and timing in a discrimination task.
Процедура исследования
В ходе предварительного обследования у испытуемых оценивали показатели чтения, рабочей памяти и интеллекта. В случае, если эти показатели соответствовали возрастной норме, испытуемый принимал участие в ЭЭГ-исследовании, в ходе которого он выполнял тест дискриминации стимулов. В ходе тренировочной серии, целью которой было усвоение инструкции, испытуемым предъявлялось по 5 проб каждого типа. Экспериментальная серия содержала по 70 проб каждого типа, количество проб лимитировано допустимой для детей продолжительностью исследования, которая не должна превышать 25 мин, включая тренировочную сессию и запись фоновой ЭЭГ. Стимулы предъявлялись на экране монитора (Dell P2213, размер видимой области по диагонали – 55.88 см), расположенного на расстоянии 1 м от глаз испытуемого. Шрифт – 36, Arial, черный шрифт на белом фоне, контраст изображения 50%. Угол обзора соответствовал оптимальным углам от –15° до 15° (Чулков, 2011). Парные стимулы располагались по центру экрана один под другим, одиночные располагались по центру экрана. Уровень освещенности помещения – 200–300 Лк.
Во время выполнения теста проводилась регистрация монополярной ЭЭГ от 19 электродов, расположенных по системе 10/20 (Fp1, Fp2, F3, Fz, F4, F7, F8, C3, Cz, C4, T3, T4, T5, T6, Р3, Pz, Р4, O1, O2), с использованием общего усредненного референтного монтажа, заземляющий электрод располагался на голове испытуемого в области вертекса. Для регистрации ЭЭГ использован электроэнцефалограф Нейровизор БММ NVX 52 фирмы Neurobotics (Зеленогорск, Россия). Сопротивление электродов не превышало 10 кОм. ЭЭГ регистрировалась в полосе 0.53–50 Гц, частота дискретизации составляла 500 Гц, сетевой фильтр 45–55 Гц. Для регистрации движений глаз и морганий использовали биполярное отведение с расположением электродов в районе височного угла глаза и верхнего края глаза.
Анализ поведенческих данных
По результатам выполнения испытуемым теста чтения слов рассчитывали следующие показатели: а) процент правильных ответов – количество верно принятых решений, включает преждевременные нажатия; б) процент преждевременных нажатий – количество правильных ответов при нажатии на кнопку до появления знака вопроса; в) процент правильных нажатий – количество верно принятых решений, в которых испытуемый своевременно (после знака вопроса) нажал на кнопку; д) время реакции – время, прошедшее между появлением знака вопроса и нажатием на кнопку (рассчитывалось на основе правильных нажатий).
Статистический анализ производили в пакете программ IBM SPSS Statistics, версия 26. Соответствие распределения нормальному определяли с помощью критерия Шапиро–Уилка. Поскольку не все исследованные параметры подчинялись закону нормального распределения, то достоверность различий перечисленных выше показателей в трех возрастных группах оценивали с помощью непараметрического H-критерия Краскела–Уоллиса. Апостериорные сравнения проводились с помощью U-критерия Манна–Уитни, применялась поправка Бонферрони на количество сравнений (три возраста между собой). В тексте приведены средние значения показателей и 95%-й доверительный интервал для среднего.
Анализ ЭЭГ данных
Обработку ЭЭГ и расчет ССП производили в программе “WinEEG” (версия 2.140.113). Для удаления глазодвигательных артефактов и миограммы был использован метод независимых компонент (Chaumon et al., 2015). Фрагменты записи, содержащие другие виды артефактов, удалялись из обработки на основе визуального анализа. Из усреднения также исключались пробы, ошибочно дискриминированные испытуемым, и пробы с преждевременным нажатием. Расчет вызванных потенциалов производили для второго стимула пробы (повторяющееся/новое слово) для каждого из 19 отведений (для построения топограмм распределения амплитуды пиков вызванного ответа). Для каждой пробы производилась коррекция изолинии. Расчет вызванных потенциалов производили на интервале от 300 мс до предъявления стимула до 1000 мс после предъявления стимула (всего 1300 мс). На этом интервале выделяли для последующего анализа два пика после предъявления стимула: N400 и P600. Интервалы для расчета компонентов N400 и P600 брали в соответствии с литературными данными (Hoeks et al., 2004; Kim, Osterhout, 2005; Nieuwland, van Berkum, 2005; Kutas, Federmeier, 2011; Delogu et al., 2019) и на основе визуальной оценки: соответственно, 350–550 и 550–900 мс после предъявления стимула.
Для статистического анализа ССП компонент N400 усредняли по 6 отведениям ЭЭГ (F3, Fz, F4, C3, Cz, C4), а P600 – по 3 отведениям (Р3, Pz, Р4). Выбор каналов для усреднения производили на основе локализации максимума ответа в соответствующем временном окне. Пиковые амплитуды вычисляли для каждого испытуемого отдельно: для N400 – это максимальное значение амплитуды сигнала во временном окне 350–550 мс, а для P600 – 550–900 мс. Кроме того, для каждого значения пиковой амплитуды определялась латентность. В тексте приведены средние значения показателей и 95%-й доверительный интервал для среднего. Соответствие распределения нормальному определяли с помощью критерия Шапиро–Уилка. Поскольку не все исследованные параметры подчинялись закону нормального распределения, то достоверность межгрупповых различий амплитуд и латентностей пиков, вычисленных для каждого испытуемого отдельно, оценивали с помощью непараметрического H-критерия Краскела–Уоллиса. Апостериорные сравнения проводились с помощью U-критерия Манна–Уитни, применялась поправка Бонферрони на количество сравнений. Амплитуды и латентности пиков на повторяющиеся и новые слова сравнивали попарно внутри каждой возрастной группы с использованием непараметрического Т-критерия Вилкоксона для связанных выборок с последующей корректировкой уровня значимости на количество возрастных групп (3) и количество сравниваемых показателей (2, амплитуда и латентность).
Помимо сравнения пиковых амплитуд внутри каждой группы сопоставляли средние амплитуды компонентов ССП на повторяющееся и новое слово на последовательных интервалах длительностью 50 мс с помощью Т-критерия Вилкоксона для связанных выборок. Для пика N400 был исследован интервал от 350 до 550 мс, на котором располагалось четыре “эпохи” по 50 мс (350–400, 400–450, 450–500, 500–550 мс). Для пика P600 был изучен интервал от 600 до 900 мс, на котором располагалось шесть “эпох” по 50 мс. Применяли поправку Бонферрони для корректировки уровня значимости гипотезы на количество сравнений. Статистический анализ производили в пакете программ IBM SPSS Statistics, версия 26.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Поведенческие данные
Анализ чтения текстов по методике СМИНЧ выявил, что хотя скорость чтения у детей 9–11 лет была ниже, чем у подростков 12–14 лет (87.98 ± 29.04 и 133.07 ± 24.32 слов в мин соответственно, H = 15.560, p < 0.001), однако характеристики понимания прочитанных текстов в этих группах не различались, коэффициент техники чтения, учитывающий как скорость чтения, так и понимание прочитанного, составлял 100.8 и 103.6 соответственно. Таким образом, по всем исследованным показателям дети и подростки соответствовали возрастной норме.
Средние показатели интеллекта, оцениваемого по методике Кеттелла, в группах детей, подростков и взрослых также соответствовали возрастной норме и составили соответственно 115.889 ± 11.14; 106.929 ± 13.58; 117.3 ± 13.3. Объем рабочей памяти у детей, подростков и взрослых оценивался на основе субтеста методики Векслера “повторение цифровых рядов” (в прямом и обратном порядке). Интегральный показатель выполнения субтеста составил соответственно 8.3 ± 1.3; 9.9 ± 2.1 и 12.1 ± 4.5. С использованием критерия Краскела–Уоллиса выявлены значимые различия между возрастными группами ($\chi _{{(2)}}^{2}$ = 30.341, скорректированный p = 0.001), при этом нет достоверных отличий между детьми и подростками, однако обе группы детей отличаются от взрослых (U = 40.50, p = 0.000 и U = = 83.50, p = 0.000 соответственно).
Испытуемые всех возрастных групп успешно справились с дифференциацией повторяющегося или нового слова (табл. 1). Процент ошибочных ответов не превышал 2.5% у взрослых, 4% у подростков и 9.5% у детей 9–11 лет. При анализе результатов выполнения теста с использованием критерия Краскела–Уоллиса выявлены значимые различия между возрастными группами в показателях реакции на предъявление повторяющегося слова по проценту правильных ответов ($\chi _{{(2)}}^{2}$ = 13.889, скорректированный p = 0.003) и корректных нажатий ($\chi _{{(2)}}^{2}$ = 16.338, p = 0.001), а также на предъявление нового слова по проценту правильных ответов ($\chi _{{(2)}}^{2}$ = 13.334, p = = 0.003), корректных ($\chi _{{(2)}}^{2}$ = 20.597, p = 0.001) и преждевременных ($\chi _{{(2)}}^{2}$ = 19.813, p = 0.001) нажатий. Процент правильных ответов на повторяющееся слово у детей 9–11 лет был достоверно ниже, чем у подростков (U = 115.00, скорректированный p = 0.03) и взрослых (U = = 110.50, p = 0.000), как и процент корректных нажатий (U = 105.00, p = 0.01 и U = 96.50, p = 0.000 соответственно), по проценту преждевременных нажатий достоверных различий выявлено не было. При выборе нового слова процент правильных ответов и корректных нажатий у детей 9–11 лет был достоверно ниже, чем у взрослых (U = 106.50, p = 0.001; U = = 70.00, p = 0.001 соответственно). Кроме того, при выборе нового слова дети и подростки достоверно отличались по проценту преждевременных нажатий от взрослых (U = 86.50, p = 0.001 и U = 94.00, p = 0.01 соответственно), что может свидетельствовать о незрелой системе управляющих функций у детей и подростков. Не выявлено достоверных возрастных отличий по времени реакции ни на предъявление повторяющегося, ни нового слова. Поскольку у испытуемого не было задачи нажимать как можно быстрее, строго говоря, этот показатель нельзя считать напрямую связанным со скоростью принятия решения.
Таблица 1.
Процент правильных ответов, корректных нажатий и преждевременных реакций у детей, подростков и взрослых при выборе повторяющегося или нового слова Table 1. Percentage of correct answers, correct clicks and advanced reactions in children, adolescents and adults during discrimination of old/new word
| % правильных ответов | % преждевременных ответов | % правильных нажатий | время реакции ответа на стимул (мс) | |
|---|---|---|---|---|
| Повторяющееся слово | ||||
| дети 9–10 лет | 90.745 ± 3.799 | 6.014 ± 2.295 | 84.731 ± 4.752 | 675.948 ± 63.229 |
| подростки 12–14 лет | 96.322 ± 2.236 | 2.001 ± 1.234 | 94.322 ± 2,511 | 674.235 ± 82.628 |
| взрослые | 97.515 ± 1.466 | 1.223 ± 0.761 | 96.292 ± 1.617 | 618.730 ± 62.248 |
| Новое слово | ||||
| дети 9–10 лет | 90.243 ± 3.836 | 3.915 ± 1.874 | 86.329 ± 4.331 | 742.430 ± 71.810 |
| подростки 12–14 лет | 95.248 ± 2.805 | 1.746 ± 0.983 | 93.502 ± 3.305 | 745.525 ± 96.083 |
| взрослые | 97.440 ± 1.662 | 0.376 ± 0.221 | 97.065 ± 1.661 | 664.239 ± 73.346 |
Между двумя типами проб (повторяющееся/новое слово) достоверные отличия выявлены только по времени реакции: у взрослых (критерий знаковых рангов Уилкоксона Z = = –3.425, скорректированный p = 0.004), подростков (Z = –3.148, p = 0.008) и детей (Z = = –3.350, p = 0.004) время реакции на новое слово меньше.
Анализ связанных с событием потенциалов
Анализ топограмм вызванных ответов показал, что локализация компонента N400 имеет сходный характер во всех возрастных группах, захватывая лобные отделы с максимумом в Fz для повторяющегося слова (рис. 2 (а), I), и слегка смещается в сторону Cz у подростков и взрослых в ситуации предъявления нового слова (рис. 2 (б), I). Поэтому для анализа компонента N400 производили усреднение по отведениям F3, F4, Fz, C3, C4, Cz. Максимум амплитуды P600 наблюдался во временном интервале 700–800 мс в Pz у детей и подростков при предъявлении повторяющегося слова (рис. 2 (а), III) и был смещен вправо при предъявлении нового слова (рис. 2 (б), III). У взрослых небольшая правосторонняя латерализация наблюдалась в обоих случаях на интервале 600–700 мс (рис. 2, II). Поэтому для анализа компонента P600 производили усреднение по отведениям Р3, Pz, Р4.
Рис. 2.
Топографическое распределение эффектов N400 (усреднено по отведениям F3, F4, Fz, C3, C4, Cz на интервале 400–500 мс (I)) и Р600 (усреднено по отведениям Р3, Pz, Р4 на интервалах 600–700 (II) и 700–800 (III) мс) при чтении повторяющегося (а) и нового (б) слов в группах детей, подростков и взрослых.
Fig. 2. Topographic distribution of N400 (averaged F3, F4, Fz, C3, C4, Cz at the interval 400–500 ms (I)) and P600 (averaged Р3, Pz, Р4 at intervals 600–700 (II) and 700–800 (III) ms) during reading repetitive (а) and new (б) words in children, adolescents and adults.

Возрастные различия
Межгрупповой анализ значений максимальной амплитуды пиков N400 и P600, определенных индивидуально у каждого испытуемого, показал, что с возрастом амплитуда обоих исследуемых компонентов ССП уменьшается. Это справедливо как для предъявления повторяющегося, так и нового слова (табл. 2). Значения χ2 (при p < 0.001 на повторяющееся слово N400 – $\chi _{{(2)}}^{2}$ = 26.084, P600 – $\chi _{{(2)}}^{2}$ = 30.354) и результаты апостериорных сравнений показали, что максимальная амплитуда компонентов N400 и P600 в ответ на предъявление повторяющегося слова не различалась достоверно между детьми и подростками, в то же время были выявлены достоверные (p < 0.001) отличия между детьми и взрослыми (N400 – U = 53.000; P600 – U = = 44.000), а также между подростками и взрослыми (U = 48.000, U = 34.000 соответственно). Что касается нового слова, то амплитуда пика N400 достоверно различалась между всеми группами испытуемых ($\chi _{{(2)}}^{2}$ = = 31.866, p < 0.001), тогда как амплитуда P600 ($\chi _{{(2)}}^{2}$ = 24.777, p < 0.001) различалась только между детьми и взрослыми (U = 59.500, p < < 0.001) и между взрослыми и подростками (U = 74.500, p = 0.004), однако наблюдалась тенденция к отличиям между подростками и детьми (скорректированный p = 0.08). Достоверных межвозрастных различий латентности пиков N400 и P600 для каждого типа стимулов выявлено не было.
Таблица 2.
Значения амплитуды (мкВ) и латентности (мс) пиков N400 и P600 у детей, подростков и взрослых Table 2. Values of amplitude (μV) and latency (ms) of N400 and P600 peaks in children, adolescents and adults
| N400 | Р600 | |||
|---|---|---|---|---|
| Амплитуда, мкВ | Латентность, мс | Амплитуда, мкВ | Латентность, мс | |
| Ответ на повторяющееся слово | ||||
| Дети | –2.439 ± 0.337 | 474.080 ± 25.077 | 4.060 ± 0.709 | 748.000 ± 43.887 |
| Подростки | –2.312 ± 0.472 | 470.118 ± 30.087 | 3.281 ± 0.579 | 731.765 ± 42.501 |
| Взрослые | –1.130 ± 0.266 | 486.909 ± 33.193 | 1.643 ± 0.260 | 688.000 ± 32,527 |
| Ответ на новое слово | ||||
| Дети | –2.931 ± 0.431 | 470.720 ± 23.767 | 3.691 ± 0.771 | 736.160 ± 43.026 |
| Подростки | –2.198 ± 0.343 | 462.588 ± 30.703 | 2.409 ± 0.629 | 686.353 ± 55.619 |
| Взрослые | –1.420 ± 0.204 | 441.091 ± 25.600 | 1.328 ± 0.266 | 674.546 ± 47.432 |
Отличия, связанные с заданием
Внутригрупповые сравнения максимальной амплитуды пика N400 не выявили достоверных различий ответа на повторяющееся и новое слово ни в одной из возрастных групп (рис. 3, I). Средние значения амплитуды ответов на интервалах 350–400, 400–450, 450–500 и 450–550 мс не различались у детей и подростков. Однако в группе взрослых были выявлены различия на интервалах 350–400 мс (Z = –2.549, скорректированный p = 0.044) и 400–450 мс (Z = –2.906, скорректированный p = 0.016). Различия в латентности пика N400 между двумя типами стимулов не проявлялись у детей и подростков, а были обнаружены только в группе взрослых: латентность пика N400 в ответ на новое слово достоверно меньше, чем на повторяющееся, Z = –2.640, скорректированный p = 0.048.
Рис. 3.
Различия вызванного ответа (I) – N400 (усредненного по отведениям F3, F4, Fz, C3, C4, Cz), (II) – Р600 (усредненного по отведениям Р3, Pz, Р4) на предъявление повторяющегося (черный цвет) и нового (серый цвет) слов в группах детей (а), подростков (б) и взрослых (в). Выделены интервалы достоверных различий. По оси абсцисс время от начала предъявления стимула (мс), по оси ординат – амплитуда ответа (мкВ).
Fig. 3. (I) – N400 (averaged over leads F3, F4, Fz, C3, C4, Cz), (II) – Р600 (averaged over leads Р3, Pz, Р4) differences to repeated (black) or new (grey) words in children (а), adolescents (б) and adults (в). Intervals of significant differences are marked. The abscissa is the time from the beginning of the stimulus presentation (ms), the ordinate is the response amplitude (μV).
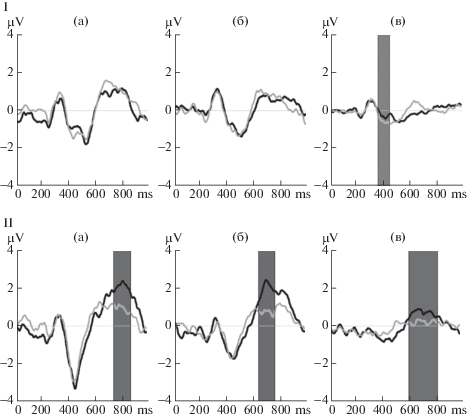
Внутригрупповые межстимульные различия амплитуды Р600 во всех возрастных группах оценивали двумя способами: по индивидуальным амплитудам пика, а также по средним значениям амплитуды в последовательных окнах по 50 мс. По амплитудам пика различия между стимулами были выявлены в группах подростков и взрослых, по средним значениям – во всех группах испытуемых. При этом пиковая амплитуда ответа, вызванного повторяющимся словом, была достоверно выше, чем амплитуда ответа, вызванного новым словом (табл. 2). Достоверность отличий оценивалась Т-критерием Вилкоксона для связанных выборок, для подростков Z = –2.911, скорректированный р = 0.024, для взрослых Z = –2.841, скорректированный р = 0.030. В группе детей достоверных отличий максимальной амплитуды пика P600 между двумя типами стимулов выявлено не было. Достоверных отличий латентности пика P600 в ответ на повторяющееся и новое слово не было выявлено ни в одной из возрастных групп.
При сопоставлении средней амплитуды P600 на повторяющееся и новое слово на последовательных временных интервалах от 600 до 900 мс с шагом 50 мс выявлены интервалы отличий для разных возрастных групп (табл. 3, рис. 3, II). Так, у детей с использованием непараметрического Т-критерия Вилкоксона для связанных выборок выявлены различия (при р < 0.01) на интервалах от 750 до 850 мс, у подростков – 650–750 мс, а у взрослых – 600–800 мс.
Таблица 3.
Различия вызванного ответа P600 (усредненного по отведениям Р3, Pz, Р4) на повторяющееся и новое слово в группах детей, подростков и взрослых Table 3. P600 differences (averaged over leads Р3, Pz, Р4) to repeated and new words in children, adolescents and adults
| 600–650 мс | 650–700 мс | 700–750 мс | 750–800 мс | 800–850 мс | |
|---|---|---|---|---|---|
| 9–11 лет | Z = 0.444 | Z = 1.224 | Z = 1.682 | Z = 2.597 | Z = 3.216 |
| p = 1.314 | p = 0.442 | p = 0.186 | p = 0.014 | p = 0.002 | |
| 12–14 лет | Z = 1.302 | Z = 3.243 | Z = 3.337 | Z = 2.249 | Z = 2.438 |
| p = 0.386 | p = 0.002 | p = 0.002 | p = 0.050 | p = 0.030 | |
| взрослые | Z = 3.263 | Z = 3.685 | Z = 3.230 | Z = 2.873 | Z = 2.224 |
| p = 0.002 | p = 0.000 | p = 0.002 | p = 0.008 | p = 0.052 |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Возрастные различия
Результаты проведенного исследования показали, что с возрастом происходит, с одной стороны, упрочение навыка чтения слов, которое выражается в снижении ошибочных и преждевременных нажатий, увеличении скорости чтения, а, с другой стороны, происходит созревание нейрофизиологических механизмов, обеспечивающих этот процесс, выражающееся в изменении характеристик вызванного ответа. Так, при предъявлении как повторяющихся, так и новых слов, происходит снижение амплитуды компонентов N400 и P600 с возрастом (табл. 2). Это соотносится с литературными данными об уменьшении величины как этих (Holcomb et al., 1992, Hahne et al., 2004; Atchley et al., 2006; McNealy et al., 2009; Benau, 2011), так и более ранних (Maurer et al., 2006; Сороко и др., 2018; Портнова и др., 2014) компонентов вызванного ответа с возрастом при обработке вербальной и невербальной информации. Снижение амплитуды может отражать снижение потенциала постсинаптических нейронов, активацию меньшего числа нейронов популяции, а также снижение временной синхронности генерации в популяции нейронов (Kutas, Federmeier, 2011). Это обусловлено, с одной стороны, “общими” процессами, связанными с развитием мозга как таковым. К ним можно отнести, например, изменения объема серого вещества мозга (Whitford et al., 2007) и изменение толщины костей черепа, которые приводят к снижению амплитуды (Ставцев и др., 2007) и мощности ЭЭГ-сигнала (Фарбер и соавт., 1990). С другой стороны, по мере становления навыка происходят существенные структурно-функциональные перестройки нейронной организации мозга, формирование нейронных ансамблей и связей внутри и между ними (Фарбер и др., 1990), приводящие к возрастной минимизации функций (Шеповальников и др., 1991).
В контексте данного исследования обращает на себя внимание тот факт, что возрастные различия между детьми и подростками менее выражены, чем различия между подростками и взрослыми. На наш взгляд, отчасти это может быть обусловлено траекторией развития серого вещества коры лобной и теменной долей мозга. Показано, что с 9 до 13 лет объем серого вещества изменяется незначительно, а после этого возраста начинает уменьшаться (Giedd et al., 1999), в основном за счет изменения объема нейропиля и количества синаптических контактов, что приводит к снижению синхронизации работы нейронов, участвующих в вызванном ответе (Whitford et al., 2007). С другой стороны, снижение амплитуды может быть связано с упрочением навыка чтения и ростом языковой компетенции. Так, уменьшение амплитуды N400 наблюдается у детей и взрослых, имеющих более высокие показатели языковых способностей (Neville et al., 1993; Weber-Fox et al., 2003; Hampton, Weber-Fox, 2013).
Различия, связанные с заданием
В нашем исследовании испытуемый должен был читать слово, а затем сопоставлять его с ранее прочитанными словами. Данные о временном ходе обработки слов (Marinković, 2004) показывают, что у взрослых понимание слова происходит примерно через 400 мс после начала его предъявления. Таким образом, мы исходим из того, что компонент N400 должен отражать лексико-семантическую обработку слов как у взрослых, так и у детей (Friederici, 2004; Crandall, 2010; Kutas, Federmeier, 2011). Кроме того, примерно в этом же временном интервале (300–500 мс) выделяют фронтальный негативный компонент, связанный с узнаванием (при невозможности детализировать воспоминание) (Woodruff et al., 2006). В используемой нами парадигме из-за того, что между запоминанием информации и извлечением ее из памяти происходит слишком малое время, отдельно узнавание выделить сложно. Более поздние компоненты, такие как P600 и LPC (later positive component), должны отражать процессы, происходящие уже после восприятия слова, в том числе воспоминание ранее прочитанных слов (Friedman, Johnson, 2000) и повторный анализ данного слова. В то же время компонент P600 связывают с синтаксическим и семантическим повторным анализом (reanalysis) (Friederici, 1997; Canseco-Gonzalez, 2000).
В литературе описан так называемый “old/new effect”, который заключается в том, что ответ на новое слово характеризуется у взрослых большей амплитудой негативного компонента во временном интервале 400–500 мс и меньшей амплитудой позитивного ответа в интервале 600–800 мс (Rugg, Curran, 2007). Подобный эффект в нашем исследовании был показан у взрослых, но не у детей и подростков. Таким образом, различия по компонентам N400 и P600, связанные с определением новизны слова, выявляются в разные периоды онтогенеза. Сначала проявляются различия вызванного ответа между повторяющимся и новым словом по компоненту Р600: сравнение на интервалах по средним значениям показывает различия между стимулами во всех обследованных возрастных группах (табл. 3). Амплитуда вызванного ответа выше на повторяющееся слово, чем на новое, у всех испытуемых, сравнения по пикам показали достоверные отличия у взрослых и у подростков. Это соотносится с литературными данными (Wilding, 2000; Tsivilis et al., 2015) и может быть объяснено тем, что при анализе повторяющегося слова вспоминание (recollection) вызывает дополнительный нейронный ответ.
Различия по компоненту N400 присутствуют только у взрослых, при этом на повторяющееся слово амплитуда была меньше, а латентность пика больше, чем на новое. Возможно, решение относительно нового слова принимается взрослыми раньше, по первым слогам или опорным буквам, до полного прочтения слова, тогда как повторяющееся слово требует повышенных ресурсов внимания и перепроверки. Важно, что в детском и подростковом возрасте межстимульных различий по N400 не выявляется. Это соотносится с данными, полученными при восприятии предложений на слух: различия по N400 выявляются в группе взрослых, но не детей 8–9 и 12–13 (Schneider et al., 2019), поэтому Шнейдер и соавт. полагают, что компонент N400 “слишком груб” для оценки семантических процессов у детей. В других исследованиях показана нечувствительность N400 у детей к степени неконгруэнтности слова, при том, что у взрослых эта зависимость выражена явно (Benau, 2011). Таким образом, хотя N400 регистрируется у детей в разных экспериментальных ситуациях, он проявляет признаки “незрелости” и “созревает” достаточно поздно в онтогенезе.
Изменение чувствительности компонента N400 к типу стимула может быть связано с реорганизацией сетей, ответственных за управляющие функции мозга (Moisala et al., 2018), усилением когнитивного контроля (Taylor et al., 2015; Best, Miller, 2010) и top-down-регуляции функций (Vettera et al., 2015), которые происходят в позднем подростковом и раннем взрослом возрасте. У подростков по сравнению с детьми существенно повышается эффективность внимания и рабочей памяти (Crone, Dahl, 2012; Crone, Konijn, 2018), возрастает когнитивная гибкость (Steinbeis, Crone, 2016). Все это позволяет обрабатывать информацию более избирательно (Castel et al., 2011) и, возможно, как бы “заранее”, еще до предъявления нового слова, выделять признаки, по которым оно будет сопоставляться с ранее прочитанными. Такая избирательность, в свою очередь, может отразиться на временном ходе обработки вербальной информации. Анализ слова в контексте задания с опорой на заранее выделенные признаки может происходить уже на этапе его восприятия, параллельно этому процессу, и приводить к различиям компонента N400 между повторяющимся и новым словом. Тогда как произвольное сопоставление воспринятого слова с ранее прочитанными происходит после его распознавания и отражается в компоненте P600.
Таким образом, мы предполагаем, что компоненты N400 и P600 могут отражать разные стадии и механизмы обработки информации, которые формируются в онтогенезе не одновременно. Однако детализация этих механизмов требует дальнейшего исследования.
ЗАКЛЮЧЕНИЕ
Нами проведены исследования возрастных изменений ССП, возникающих в ответ на предъявление повторяющегося и нового слова в процессе чтения у детей предподросткового, подросткового возраста и взрослых. Показано, что с возрастом происходит как упрочение навыка чтения слов, которое выражается в снижении ошибочных и преждевременных нажатий, увеличении скорости чтения, так и созревание нейрофизиологических механизмов, обеспечивающих этот процесс, выражающееся в изменении характеристик вызванного ответа. С возрастом происходит снижение амплитуды компонентов N400 и P600 при предъявлении как повторяющихся, так и новых слов.
Отличия компонентов, возникающие в ответ на новое и повторяющееся слово, появляются в онтогенезе не одновременно. Сначала выявляются различия в амплитуде пика P600, отражающие, на наш взгляд, последовательную обработку информации. Они присутствуют как у детей, так и у взрослых, при этом амплитуда на повторяющееся слово выше, чем на новое. Различия по компоненту N400, который может отражать процессы обработки слова, происходящие параллельно с распознаванием его значения, выявляются только у взрослых, показывая меньшие значения латентности и большие значения амплитуды при предъявлении нового слова по сравнению с повторяющимся.
Список литературы
Безруких М.М., Крещенко О.Ю. Особенности формирования навыков письма и чтения у школьников младших классов. Известия Российской академии образования. 2011. 1 (17): 122–136.
Корнев А.Н. Основы логопатологии детского возраста: клинические и психологические аспекты. СПб.: Речь, 2006. 380 с.
Корнев А.Н., Ишимова О.А. Методика диагностики дислексии у детей. Методическое пособие. СПб.: Изд-во Политехнического ун-та, 2010. 72 с.
Портнова Г.В., Мартынова О.В., Иваницкий Г.А. Возрастные различия слуховых вызванных потенциалов при восприятии последовательных и пространственных компонентов звуковой информации. Физиология человека. 2014. 40 (1): 26.
Сороко С.И., Белишева Н.К., Нагорнова Ж.В., Шемякина Н.В. Анализ возрастной динамики и половых особенностей спонтанной биоэлектрической активности и компонентов слуховых вызванных потенциалов у младших школьников, проживающих в Арктической зоне РФ. Физиология человека. 2018. 2 (44): 84–95.
Ставцев A.Ю., Ушаков В.Л., Верхлютов В.М. Моделирование влияния толщины слоев и проводимости тканей головы и мозга на потенциалы ЭЭГ с использованием метода конечных элементов. Журн. высш. нервн. деят. им. И.П. Павлова. 2007. 57 (6): 742–752.
Фарбер Д.А., Семенова Л.К., Алферова В.В. Структурно-функциональное созревание развивающегося мозга. Л.: Наука, 1990. 198 с.
Филимоненко Ю.И., Тимофеев В.И. Руководство к методике исследования интеллекта у детей Д. Векслера (WISC): Адапт. вариант. СПб.: Госстандарт России, ГП “ИМАТОН”, 1994. 94 с.
Чулков Н.А. Безопасность жизнедеятельности. Национальный исследовательский Томский политехнический университет. 2011.
Шеповальников А.Н., Цицерошин М.Н., Левинченко Н.В. “Возрастная минимизация” областей мозга, участвующих в системном обеспечении психических функций: аргументы за и против. Физиология человека. 1991. 17 (5): 28.
Atchley R.A., Rice M.L., Betz S.K., Kwasnya K.M., Serenoc J.A., Jongmanc A. A comparison of semantic and syntactic event related potentials generated by children and adult. Brain Lang. 2006. 99 (3): 236–246.
Benau E.M., Morris J., Couperus J.W. Semantic Processing in Children and Adults: Incongruity and the N400. J. Psycholinguist. Res. 2011. 40 (3): 225–239.
Best J.R., Miller P.H. A developmental perspective on executive function. Child Dev. 2010. 81 (6): 1641–1660.
Canseco-Gonzalez E. Using the recording of event-related brain potentials in the study of sentence processing. Foundations of neuropsychology: A series of textbooks, monographs, and treatises. San Diego: Academic Press. 2000. P. 229–266.
Castel A.D., Humphreys K.L., Lee S.S., Galván A., Balota D.A., McCabe D.P. The development of memory efficiency and value-directed remembering across the life span: a cross-sectional study of memory and selectivity. Dev. Psychol. 2011. 47 (6): 1553–1564.
Cattell R.B. Culture Fair Intelligence Tests: CFIT. Institute for Personality and Ability Testing.1973.
Chaumon M., Bishop D.V., Busch N.A. A practical guide to the selection of independent components of theelectroencephalogram for artifact correction. J. Neurosci Methods. 2015. 250: 47.
Crandall M. The ELAN Event-Related Potential in Children 5 to 12 Years of Age. All Theses and Dissertations. 2010. 2151.
Crone E., Dahl R. Understanding adolescence as a period of social-affective engagement and goal flexibility. Nat. Rev. Neurosci. 2012. 13: 636–650.
Crone E.A., Konijn E.A. Media use and brain development during adolescence. Nat. Commun. 2018. 9 (1): 588–603.
Delogu F., Brouwer H.,Crocker M.W. Event-related potentials index lexical retrieval (N400) and integration (P600) during language comprehension. Brain Cogn. 2019. 135: 103569.
Ducharme S., Albaugh M.D., Nguyen T.-V., Hudziak J.J., Mateos-Pérez J.M., Labbe A., Evans A.C., Karama S. Trajectories of cortical surface area and cortical volume maturation in normal brain development. Data Brief. 2015. 5: 929–938.
Federmeier K.D. Thinking ahead: The role and roots of prediction in language comprehension. Psychophysiology. 2007. 44 (4): 491–505.
Fiebach C.J., Friederici A.D., Müller K., Cramon D.Y.V. fMRI evidence for dual routes to the mental lexicon in visual word recognition. J. cogn. Neurosci. 2002. 14 (1): 11–23.
Friederici A.D. The time course of syntactic activation during language processing: a model based on neurological and neurophysiological data. Brain and Language. 1995. 50:259281.
Friederici A.D. Neurophysiological aspects of language processing. Clin. Neurosci. 1997. 4 (2): 64–72.
Friederici A.D. Event-related brain potential studies in language. Current Neurology and Neuroscience Report. 2004. 4 (6): 466–470.
Friederici A.D. The neural basis of language development and its impairment. Neuron. 2006. 52. 941–952.
Friederici A.D., Hahne A. Developmental patterns of brain activity reflecting semantic and syntactic processes. In J. Weissenborn & B. Houle (Eds.), Approches to bootstrapping: Phonological, lexical, syntactic, and neurophysiologcal aspects of early language acquisition. Amsterdam/Philadelphia: John Benjamin. 2001. P. 231–246.
Friedman D., Johnson R. Event-related potential (ERP) studies of memory encoding and retrieval: A selective review. Microscopy Res. Techniq. 2000. 51 (1): 6–28.
Frost R. Toward a strong phonological theory of visual word recognition: true issues and false trails. Psychological bulletin. 1998. 123 (1): 71.
Giedd J.N., Blumenthal J., Jeffries N.O., Castellanos F.X., Liu H., Zijdenbos A., Paus T., Evans A.C., Rapoport J.L. Brain development during childhood and adolescence: a longitudinal MRI study. Nat. Neurosci. 1999. 2(10): 861–863.
Gouvea A.C., Phillips C., Kazanina N., Poeppel D. The linguistic processes underlying the P600. Lang. Cogn. Proc. 2010. 25 (2): 149188.
Hahne A., Eckstein K., Friederici A.D. Brain signatures of syntactic and semantic processes during children’s language development. J. Cogn. Neurosci. 2004. 16: 1302–1318.
Hampton W.A., Weber-Fox C. Specific aspects of cognitive and language proficiency account for variability in neural indices of semantic and syntactic processing in children. Dev. Cogn. Neurosci. 2013. 5: 149–171.
Hoeks J.C.J., Stowe L.A., Doedens G. Seeing words in context: The interaction of lexical and sentence level information during reading. Cogn.Brain Res. 2004. 19 (1): 59–73.
Holcomb P.J., Coffey S.A., Neville H.J. Visual and auditory sentence processing: a developmental analysis using event related brain potentials. Dev. Neuropsychol. 1992. 8: 203–241.
Huttenlocher P., De Courten C., Garey L., Van der Loossynapto H. Genesis in human visual cortex–evidence for synapse elimination during normal development. Neurosci Lett. 1982. 33: 247–252.
Kaan E., Harris A., Gibson E., Holcomb P. J. The P600 as an index of syntactic integration difficulty. Lang. Cogn. Proc. 2000. 15: 159201.
Kim A., Osterhout L. The independence of combinatory semantic processing: evidence from event-related potentials. J. Mem. Lang. 2005. 52 (2): 205–225.
Kutas M., Federmeier K.D. Thirty years and counting: finding meaning in the N400 component of the event-related brain potential (ERP). Ann. Rev. Psychol., 2011. 62: 621–647.
Maïonchi-Pino N., Magnan A., Écalle J. Syllable frequency effects in visual word recognition: Developmental approach in French children. J. Appl. Dev. Psychol. 2010. 31: 70–82.
Mandler G., Yonelinas A.P., Aly M., Wang W.C., Koen J.D. Recollection and familiarity: examining controversial assumptions and new directions. Hippocampus. 2010. 20 (11): 1178–1194.
Marinković K. Spatiotemporal dynamics of word processing in the human cortex. The Neuroscientist: a review journal bringing neurobiology, neurology and psychiatry. 2004. 10 (2): 142–152.
Maurer U., Brem S., Kranz F., Bucher K., Benz R., Halder P., Steinhausen H.-C., Brandeis D. Coarse neural tuning for print peaks when children learn to read. NeuroImage. 2006. 33: 749–758.
McNealy K., Dapretto M., Bookheimer S. Language and the developing brain: insights from neuroimaging. Neuroimaging in Developmental Clinical Neuroscience, eds. Judith M. Rumsey and Monique Ernst. Published by Cambridge University Press. Cambridge University Press. 2009. 91–107.
Mills K.L., Goddings A.-L., Herting M.M., Meuwese R., Blakemore S.-J., Crone E.A., Dahl R.E., Güroğlu B., Raznahan A., Sowell E.R., Tamnes C.K. Structural brain development between childhood and adulthood: Convergence across four longitudinal samples. Neuroimage. 2016. 141: 273–281.
Moisala M., Salmela V., Carlson S., Salmela-Aro K., Lonka K., Hakkarainen K., Alho K. Neural activity patterns between different executive tasks are more similar in adulthood than in adolescence. Brain Behav. 2018. 8 (9): e01063.
Neville H.J., Coffey S.A., Holcomb P.J., Tallal P. The neurobiology of sensory and language processing in language-impaired children. J. Cogn. Neurosci. 1993. 5: 235–253.
Nieuwland M.S., van Berkum J.J.A. Testing the limits of the semantic illusion phenomenon: ERPs reveal temporary semantic change deafness in discourse comprehension. Cogn. Brain Res. 2005. 24 (3): 691–701.
Rossetti M.F., Cambiasso M.J., Holschbach M.A., Cabrera R.J. Oestrogens and Progestagens: Synthesis and Action in the Brain. Neuroendocrinol. 2016. 28: 7.
Schmidt-Kassow M., Kotz S.A. Event-related brain potentials suggest a late interaction of meter and syntax in the P600. J. Cogn. Neurosci. 2009. 21: 1693–1708.
Rugg M.D., Curran T. Event-related potentials and recognition memory. TRENDS in Cognitive Sciences. 2007. 11 (6): 251–257.
Schneider J.M., Abel A.D., Ogiela D.A., Middleton A.E., Maguire M.J. Developmental differences in beta and theta power during sentence processing. Dev. Cogn. Neurosci. 2016. 19: 19–30.
Seassau M., Bucci M.P. Reading and visual search: a developmental study in normal children. PLoS One. 2013. 8 (7): e70261.
Singh M., Su C. Progesterone and neuroprotection. Horm. Behav. 2013. 63 (2): 284–290.
Steinbeis N., Crone E.A. The link between cognitive control and decision-making across child and adolescent development. Curr. Opin. Behav. Sci. 2016. 10: 28.
Taylor S.J., Barker L.A., Heavey L., McHale S. The longitudinal development of social and executive functions in late adolescence and early adulthood. Front Behav. Neurosci. 2015. 9: 252.
Toffoletto S., Lanzenberger R., Gingnell M., Sundström-Poromaa I., Comasco E. Emotional and cognitive functional imaging of estrogen and progesterone effects in the female human brain: a systematic review. Psychoneuroendocrinology. 2014. 50: 28–52.
Tsivilis D., Allan K., Roberts J., Williams N., Downes J.J., El-Deredy W. Old-new ERP effects and remote memories: the late parietal effect is absent as recollection fails whereas the early mid-frontal effect persists as familiarity is retained. Front. Hum. Neurosci. 2015. 14 (9): 532.
Vettera C., Pilhatscha M., Weige S., Ripkea S., Smolkaa M.N. Mid-adolescent neurocognitive development of ignoring and attending emotional stimuli. Dev. Cogn. Neurosci. 2015. 14: 23–31.
Vijayakumar N., Allen N.B., Youssef G., Dennison M., Yücel M., Simmons J.G., Whittle S. Brain development during adolescence: A mixed-longitudinal investigation of cortical thickness, surface area, and volume. Hum. Brain Mapp. 2016. 37: 2027–2038.
Weber-Fox C., Davis L.J., Cuadrado E. Event-related brain potential markers of high-language proficiency in adults. Brain Lang. 2003. 85: 231–244.
Whitford T.J., Rennie C.J., Grieve S.M., Clark C.R., Gordon E., Williams L.M. Brain maturation in adolescence: concurrent changes in neuroanatomy and neurophysiology. Hum. Brain Mapp. 2007. 28 (3): 228–237.
Wierenga L.M., Langen M., Oranje B., Durston S. Unique developmental trajectories of cortical thickness and surface area. NeuroImage. 2014. 87: 120–126.
Wilding E. In what way does the parietal ERP old/new effect index recollection? Internat. J. Psychophysiol. 2000. 35 (1): 81–87.
Woodruff C.C., Hayama H.R., Rugg M.D. Electrophysiological dissociation of the neural correlates of recollection and familiarity. Brain Res. 2006. 1100 (1): 125–135.
Yonelinas A.P., Aly M., Wang W.C., Koen J.D. Recollection and familiarity: examining controversial assumptions and new directions. Hippocampus. 2010. 20 (11): 1178–1194.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова