

Журнал высшей нервной деятельности им. И.П. Павлова, 2021, T. 71, № 2, стр. 279-285
Поведение крыс двух линий, контрастных по признаку “аудиогенная эпилепсия”
Н. М. Сурина 1, Г. М. Николаев 1, И. И. Полетаева 1, *, И. Б. Федотова 1
1 Кафедра высшей нервной деятельности, Московский государственный университет имени М.В. Ломоносова
Москва, Россия
* E-mail: ingapoletaeva@mail.ru
Поступила в редакцию 16.08.2020
После доработки 10.10.2020
Принята к публикации 22.12.2020
Аннотация
Сравнивали поведение в тесте “открытое поле” у крыс двух линий, контрастных по проявлению судорог в ответ на звук (аудиогенная эпилепсия – АЭ). Эти линии (“4” и “0”) были выведены на основе единой гибридной популяции. Тест проводили перед серией экспозиций действию звука (21 день) и после нее. В ходе последовательных предъявлений звука у крыс линии “4” формировались миоклонические судороги, тогда как у крыс “0” АЭ и, соответственно, миоклонических судорог не было. В исходном тесте у крыс линии “4” уровень локомоции и исследовательской активности были выше, чем у крыс линии “0”. Во втором тесте у крыс линии “4” значимо изменились многие показатели поведения, отражая общее угнетение исследовательской активности и локомоции. У крыс линии “0”, подвергавшихся действию звука (но не обнаруживших признаков АЭ), поведение во втором тесте было более активным, чем в первом, отражая привыкание животных к обстановке опыта, однако сдвиги ряда показателей второго теста свидетельствовали о сложном характере изменений в уровне тревожности этих животных (как следствия серийного действия сильного звука). Сходный генетический фон двух линий крыс, контрастных по АЭ, позволяет заключить, что изменения поведения крыс линии “4” явились следствием индукции серии эпилептиформных феноменов АЭ. Данная пара линий может быть четкой генетической моделью судорожных состояний, позволяющей надежно оценивать эффекты и внешних воздействий, и фармакологических агентов.
ВВЕДЕНИЕ
Использование лабораторных моделей для исследования заболеваний человека и, в частности, суждение о наличии или отсутствии “коморбидности” в проявлении патологических признаков в поведении животных разных генотипов ставит вопрос о влиянии на эти явления генетического фона сравниваемых линий (Garcia-Cairasco et al., 2017). Сопоставление проявлений аудиогенной эпилепсии (АЭ) с показателями поведения в тесте Порсолта и приподнятого крестообразного лабиринта у крыс 4 генотипов, попарно различавшихся по наличию или отсутствию аудиогенной эпилепсии (Sarkisova et al., 2017), продемонстрировало четкое влияние генетического фона животных исследуемых генотипов. Было показано, что коморбидность АЭ с признаками депрессии и тревожности в поведении обнаружилась только в одной “паре” линий крыс (линия Крушинского-Молодкина, КМ, с интенсивной АЭ) – и крыс Вистар (нет АЭ). Однако в другой “паре” линий, в которой были линия “0” с отсутствием АЭ и линия “4” с максимальной интенсивностью АЭ (селектированные из одной гибридной популяции КМ × Вистар), такая коморбидность обнаружена не была. Эти данные не позволяют говорить об отсутствии подобной коморбидности, но показывают, что эта проблема достаточно сложна, поскольку в поведении животных выявляется существенная роль генетического фона линии.
В связи с этим для оценки влияния АЭ (точнее, влияния “аудиогенного миоклонуса”) на поведение крыс в тесте “открытое поле” были использованы крысы второй упоминавшейся выше пары линий. Обе эти линии (“4” и “0”) имеют общее происхождение (и более близкий друг к другу генетический фон). Их раздельное разведение насчитывает 36 поколений, тогда как крысы линии КМ и Вистар разводятся независимо в течение многих десятков поколений, начиная с конца 1940 г. Это обстоятельство не позволяет игнорировать возможное влияние на ЦНС значительного числа независимых мутаций, которые могли возникнуть и у КМ, и у Вистар. Сходный (или близкий) генетический фон сравниваемых линий позволяет делать более уверенные заключения о влиянии на поведение (или об отсутствии такого влияния) неврологических отклонений, связанных именно с АЭ. В данной работе приводятся результаты тестирования крыс в “открытом поле”, которое животные этих линий проходили дважды – до серийного ежедневного (21 день) предъявления сильного звука и после этого. В процессе такого эксперимента у крыс с предрасположенностью к АЭ развивается явление “аудиогенного киндлинга” (по аналогии с явлением киндлинга, или “раскачки”, при ритмической электростимуляции гиппокампа или миндалины). Эти быстрые “тикообразные” миоклонические судороги затрагивают лицевую мускулатуру, мышцы шеи, передних конечностей, а иногда и туловища (Федотова, Семиохина, 2002).
Формирование миоклонических судорог связано с вовлечением в процесс эпилептогенеза структур переднего мозга, в частности, гиппокампа, неокортекса и миндалины (Федотова, Семиохина, 2002; Garcia-Cairasco et al., 2017), что не может не сказаться на функции этих структур и, следовательно, на поведении животных. В связи с этим сравнение поведения крыс линий “0” и “4” в тесте “открытое поле” до и после “выработки” миоклонических судорог может показать, как формирование патологического процесса в переднем мозге (линия “4”) может повлиять на уровень локомоции, поведение “замирания” и исследовательскую активность животных по сравнению с влиянием серийной экспозиции животных действию звука без развития судорог (линия “0”). Такое проводимое впервые сопоставление позволило выявить важные межлинейные различия в поведении, но в то же время продемонстрировало необходимость использования не одиночного теста “открытое поле”, а набора (батареи) тестов для более детального и информативного сравнения такого рода.
МЕТОДИКА
Экспериментальные животные. В работе были использованы 4-мес. крысы-самцы линии “4” (F36, n = 14) и линии “0” (F40, n = 13). Селекция линии “0” (на отсутствие АЭ при действии звука) была начала в середине 1990 г., селекция линии “4” была инициирована на основе крыс F4 линии “0”, проявлявших невысокую интенсивность АЭ. Началом этого селекционного эксперимента было получение гибридов F2 между крысами линии КМ и крысами Вистар, специально строго отобранными на отсутствие АЭ в ходе троекратной экспозиции их действию звука с интервалами 4–6 дней (Федотова и др., 2012). Затем были проведены два последовательных возвратных скрещивания с крысами линии КМ, после чего была начата собственно селекция на отсутствие АЭ (см. ниже).
Эксперименты были проведены в соответствии с биоэтическими требованиями DIRECTIVE 2010/63/EU.
Аудиогенная эпилепсия (АЭ). Тестирование АЭ, а в дальнейшем многократная экспозиция всех животных действию звука в течение 21 дня, проводились в звукоприглушающей камере (из твердого пластика). Использовали звук силой 120 дБ (аудиторный звонок). Через 6–9 сек у крыс линии “4” начиналось развитие типичного припадка АЭ: стадия быстрого (клонического) бега, затем клонические и тонические судороги, захватывающие все мышцы туловища и конечностей. У всех крыс линии “4” интенсивность приступа АЭ была максимальной. Ежедневная экспозиция крыс этой линии действию звука привела к появлению судорог другого типа – миоклонических (Poletaeva et al., 2017), которые проявлялись сразу после окончания тонического судорожного припадка. Их первое появление у некоторых животных было отмечено на 8-й день данной серии, но на 9-й день миоклонические судороги были зарегистрированы у всех животных линии “4”. У крыс линии “0” аналогичные воздействия никаких проявлений АЭ не вызвали ни у одного животного (ни генерализованного припадка, ни миоклонических судорог).
Тест “открытое поле”. Изготовленная из жесткого пластика установка “открытого поля” имела диаметр арены 1.5 м с высотой стенок 50 см. Пол арены был расчерчен на квадраты со стороной 10 см. Тестирование проводили в отдельном помещении при освещенности 50–60 люкс. Животное помещали в центр арены на 3 мин. Показатели поведения вводили в компьютер online в полуавтоматическом режиме (программа М.Г. Плескачевой). Регистрировали число пересеченных сторон квадратов на периферии арены и в ее центре, число эпизодов и длительность как эпизодов замирания, так и чистки шерсти (груминга), число вертикальных стоек и болюсов дефекации.
Оценка статистической значимости различий была проведена с использованием непараметрического критерия Манна–Уитни. Различия считались значимыми при р < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Показатели 1-го теста “открытое поле” были получены за 1 день до начала ежедневной экспозиции крыс обеих линий действию звука. На следующий день после окончания этой серии тест “открытое поле” был проведен повторно. Ежедневное действие сильного звука в течение длительного периода как несомненно негативного для состояния организма фактора могло вызвать изменения в физическом состоянии животных обеих линий. Никаких изменений шерстного покрова, состояния кожи или других внешних признаков неблагоприятных изменений у крыс, использованных в эксперименте, обнаружено не было. Потери веса тела у крыс обеих линий не было, было отмечено небольшое (статистически незначимое) его увеличение. У крыс линии “4” – с 383.1 ± 9.5 г до 392 ± ± 10.4 г, у крыс линии “0” – с 390 ± 90.1 г до 415.7 ± 9.6 г.
Результаты двух предъявлений теста сравнивали как внутри каждой линии (до и после воздействия), так и между линиями.
Сравнение показателей поведения крыс линий “4” и “0” в тесте “открытого поля” до начала ежедневных экспозиций действию звука
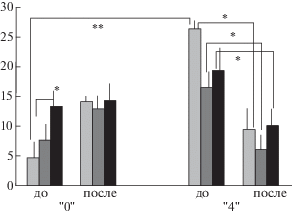
Число пересечений сторон квадратов на периферии арены за 1-ю минуту теста было статистически значимо больше у крыс линии “4”, по сравнению с линией “0” (p < 0.01, см. рис. 1). Таким образом, общий уровень двигательной активности в исходном, “фоновом” состоянии был выше у крыс линии “4”. Такие же различия были обнаружены и по суммарным показателям локомоции за 3 мин теста. Общий паттерн различий в локомоторной активности (как межлинейных, так и для каждой линии между двумя тестами) представлен на рис. 1.
Рис. 1.
Число пересечений сторон квадратов на периферии арены “открытого поля” у крыс линий “0” и “4” по 3 мин теста до и после звуковых экспозиций. Светло-серые столбики – 1-я минута, темно-серые – 2-я минута, черные – 3-я минута теста. *, ** – статистически значимое различие между показателями (р < 0.05 и 0.01, критерий Манна–Уитни).
Fig. 1. The numbers of square sides crossed in the “open field” arena periphery by rats of lines “0” and “4” in 3 min before and after sound exposures. Light grey columns – 1st min, dark grey ones – 2nd min, black – 3rd min. *,** significant difference in scores (р < 0.05 and 0.01, Mann–Witney crit.).
Число пересеченных сторон “центральных” квадратов (область из 16 квадратов в середине арены) также было несколько (незначимо) выше у крыс линии “4”: 8.31 ± 1.13, “0” – 6.36 ± 1.08. Эти различия можно трактовать как отсутствие значительных межлинейных различий в избегании центра “поля”, т.е. в тревожности этих животных, развивающейся после помещения животного на освещенную открытую площадку.
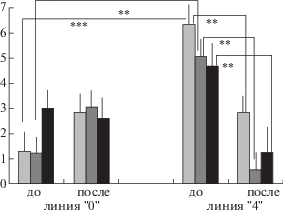
Межлинейные различия в “фоновом” поведении крыс этих линий обнаружились и в числе вертикальных стоек, наиболее четко выраженные в 1-ю и 2-ю минуту теста. За 3 мин теста число стоек у крыс линий “0” и “4” также статистически значимо различалось и было выше у крыс линии “4”: 6.23 ± 0.86, “0” – 1.35 ± 0.83 (p < 0.01). Следует отметить в целом невысокий уровень исследовательской активности у крыс обеих линий (рис. 2). Следует отметить, что в первом тесте “динамика” числа стоек за 3 последовательные минуты пребывания в “открытом поле” у крыс линии “0” и линии “4” была противоположной – у крыс линии “0” число стоек стало больше в 3-ю минуту, тогда как у другой линии обнаружилось некоторое снижение. По всей видимости, для крыс линии “0” новая обстановка вызвала подавление исследовательских движений (и проявление замирания, см. ниже), тогда как у линии “4” этого не произошло. Возможно, что это отражает наличие межлинейных различий в “фоновом” поведении крыс этих линий.
Рис. 2.
Число вертикальных стоек в тесте “Открытое поле” у крыс двух линий до и после серии звуковых экспозиций, вызвавшей миоклонические судороги у крыс линии “4”. Обозначения как на рис. 1, **, ***– статистически значимые различия (р < 0.01 и 0.001).
Fig. 2. The numbers of rearings in the “open field” text in rats of two lines before and after the sound exposure which provoked the myoclonic seizures in line “4” rats. Designations as in fig. 1. **, *** – significant differences (р < 0.01 and 0.001).
Число эпизодов груминга как показатель “конфликта” между тревожностью и исследовательским поведением (Delprato et al., 2017) было статистически незначимо ниже у крыс “0”. У “4” – 1.38 ± 0.38, у “0” – 0.78 ± ± 0.37. Время, в течение которого животные чистили шерсть, у крыс двух исследуемых линий практически не различалось (у “4” – 1.31 ± ± 0.98 с, у “0” – 1.79 ± 0.95 с).
Число эпизодов замирания (как показателя страха-тревоги при помещении животного на открытую арену) за 1-ю минуту теста было статистически значимо выше у крыс линии “0” (у “4” – 0.38 ± 0.39, у “0” – 2.0 ± 0, р < < 0.01). Суммарное время замирания было также значимо выше у крыс линии “0” (у “4” – 0.50 ± 2.47 с, у “0” – 8.48 ± 2.38 с, р < 0.01). Это свидетельствует о более высокой тревожности крыс линии “0” в обстановке теста “открытое поле” при их первом помещении на арену. Об этом же свидетельствуют и несколько более высокие показатели дефекации (как проявления эмоциональной реактивности, у линии “4” – 0.38 ± 0,25, у “0” – 0.57 ± 0.24, различия недостоверны). Как “знак” межлинейных различий, так и абсолютные величины показателей поведения за 2-ю и 3-ю минуту теста были сходны с данными по 1-й минуте.
Изменения поведения крыс двух линий в тесте “открытое поле” после серии экспозиций действию сильного звука
Крысы линии “4”. У этих животных поведение в “открытом поле” после выработки аудиогенных миоклонических судорог претерпело значительные изменения. Произошло резкое (и статистически значимое, р < < 0.05) снижение уровня локомоторной активности, выразившееся в снижении числа пересечений сторон квадратов на периферии арены как для каждой из минут теста (см. рис. 1), так и суммарно за все время теста (суммарные данные: 1-й тест – 34.91 ± 4.02, 2-й тест – 14.67 ± 3.85, p < 0.01). Такое же различие в числе пересечений сторон квадратов в центре арены не было значимым (тенденция, р = = 0.065). Таким образом, после ежедневных припадков АЭ у крыс линии “4” произошло резкое снижение локомоторной активности: число вертикальных стоек (как маркер исследовательской активности животного) во втором тесте резко и статистически значимо (p < < 0.01) стало ниже (рис. 2).
Эпизодов груминга у крыс этой линии стало статистически значимо больше во втором тесте (1-я минута, р = 0.013): 1-й тест – 1.18 ± ± 0.43, 2-й тест – 3.00 ± 0.42 (р < 0.05). Время, занятое грумингом, во втором тесте также статистически значимо увеличилось (p < 0.01) (1-й тест – 0.80 ± 2.54 с, 2-й тест – 10.87 ± 2.44 с). Число эпизодов замирания между тестами не различалось, но время замирания (проявление тревожной реакции на обстановку теста) в 1-ю минуту теста статистически значимо увеличилось: 1-й тест – 0.39 ± 3.51 с, 2-й тест – 14.65 ± 3.36 с (р < 0.05).
Крысы линии “0”. У крыс этой линии различия в показателях теста “открытое поле” до и после звуковых воздействий были практически во всех случаях статистически незначимыми, за одним исключением. Уровень дефекации (число болюсов суммарно за 3 мин теста) у крыс линии “0” достоверно снизился (1-й тест – 1.6 ± 0.4, 2-й тест – 0.4 ± 0.4, р < < 0.05). Это может быть свидетельством снижения уровня тревожности у крыс “0” ко 2-му тесту, хотя делать этот вывод на основании только данного показателя может быть не очень надежно, поскольку другие индексы тревожности у этих животных не свидетельствуют о снижении тревожности. Отметим, что суммарно за 3 мин теста локомоторная активность во 2-м тесте стала незначимо выше (1-й тест – 36.69 ± 9.26, 2-й тест – 56.42 ± ± 9.64). Отсутствие статистически значимых различий между показателями двух тестов у крыс линии “0” отражает, по всей видимости, большой индивидуальный разброс этих показателей. Это, в свою очередь, может быть связано с разным влиянием дискомфортного воздействия (серии звуковых экспозиций) на этих крыс. Известно, что в тесте “открытого поля” реакцией животного может быть как повышенная локомоция (реакция “панического бега”), так и “затаивание” (Edut, Eilam, 2004; Miguel et al., 2010; Sun et al., 2020]). Оказалось, что число эпизодов замирания за 3 мин теста у крыс линии “0” при повторном тестировании стало больше (5.8 ± 1.0 и 7.3 ± ± 1.0), как стало больше и число эпизодов груминга (2.9 ± 0.7 и 4.3 ± 0.8), т.е. действие сильного звука отразилось на этих показателях. Из этого следует, что суждение об изменении такой “внутренней переменной”, как уровень тревожности следует делать на основе показателей не одного теста, а их батареи, что в данной работе сделано не было.
Сравнение показателей поведения крыс двух линий в тесте “открытого поля” у крыс после множественных ежедневных экспозиций действию звука
Число квадратов, пересеченных в 1-ю минуту теста, было несколько (статистически незначимо) выше у крыс линии “0”. Число стоек в 1-ю минуту теста было почти одинаковым, а эпизодов груминга было статистически значимо больше у крыс линии “4” (“4” – 3.0 ± ± 0.46, “0” – 1.2 ± 0.43, р < 0.01). Время, занятое грумингом в 1-ю минуту теста, также было у них больше: линия “4” – 10.87 ± 2.7 с, линия “0” – 3.2 ± 2.5 с, р < 0.05). Длительность эпизодов замирания была больше у крыс линии “4”, т.е. их испуг проявлялся в более длительном и частом замирании. У крыс линии “4” это было 10.58 ± 3.52 с, у крыс линии “0” – 5.01 ± 3.38 с. Данные за 3 мин теста имели, в целом, такой же “рисунок” межлинейных различий, как и в 1-ю минуту теста.
В то же время стоек (за 3 мин суммарно) у крыс линии “0” стало статистически значимо больше, чем у линии “4” (8.7 ± 1.2 и 2.5 ± 1.4, p < 0.01), число болюсов дефекации было больше у крыс линии “4” (2.0 ± 0.4 и 0.7 ± 0.4, p < 0.05).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
И межлинейные сравнения поведения в тесте “открытого поля”, и сравнение этих показателей в каждой линии “до” и “после” многократных экспозиций действию звука показали, что в “фоне” крысы линии “0” были значительно менее активны и обнаруживали несколько более выраженную тревожность, чем крысы линии “4”. В ходе множественных предъявлений звука у крыс линии “0” не было проявлений АЭ, тогда как у крыс “4” были и тонические, и миоклонические судорожные припадки. Поведение крыс линии “0” во 2-м тесте свидетельствует о проявлении у них “привыкания” к обстановке теста – возросла их двигательная активность и снизились знаки тревожности (стало больше исследовательских стоек, произошло снижение уровня дефекации и др.). Поведение крыс линии “4” во 2-м тесте изменилось противоположным образом. У них достоверно снизились и уровень локомоции, и число вертикальных стоек, а показатели тревожности – усилились. Таким образом, в противоположность крысам “0” (у которых поведение стало более активным), у крыс “4” после серии судорожных припадков имело место угнетение активности, которое следует отнести за счет регулярного проявления судорожных состояний.
Рисунки 1 и 2 четко показывают, что изначально существовавшие (1-й тест) межлинейные различия приобретают во 2-м тесте другой “знак”. Подавление локомоции и исследовательской активности у крыс линии “4” после длительной серии индуцированных звуком судорожных припадков и отсутствие такого феномена у крыс линии “0” могут быть указанием на реальное существование связи между судорожными феноменами АЭ и “депрессивно-подобным” феноменом в виде изменившейся картины теста у линии “4”. У крыс американской линии GEPR-3 (с невысокой интенсивностью судорог АЭ) была показана ассоциация АЭ и особенностей поведения (при сопоставлении с показателями исходной линии Sprague-Dawley, в частности, в тесте “открытого поля”). Однако, как уже обсуждалось выше, такое сопоставление не учитывает возможных различий в генетическом фоне сравниваемых групп. Данных по такой ассоциации между крысами Sprague-Dawley и крысами линии GEPR-9 (у которой судороги АЭ более интенсивны) авторы не приводят (Aguilar et al., 2018). Сопоставление поведения крыс бразильской линии WAR (Wistar Audiogenic Rats) с исходной популяцией Wistar позволило выявить особенности поведения крыс с АЭ, однако прямых свидетельств коморбидности АЭ с проявлениями депрессии и тревожности найдено не было (Castro et al., 2017). Влияние генетического фона линии (точнее, особенностей генотипа Wistar и Sprague-Dawley, т.е. линий, служивших исходными популяциями для селекции на АЭ) выявилось в новом морфологическом исследовании мозга двух линий с АЭ: GEPR-3 и WAR (Lee et al., 2018). При практически полном сходстве фенотипа АЭ, у этих линий различались размеры ряда структур мозга, связанных с развитием судорог, характерных для АЭ. Понятие “генетический фон (genetic background) предполагает особенности набора генов одной линии в отличие от такового другой, которые могут быть причиной различий в эффектах идентичного воздействия. Показано, например, что генотип линии может определять особенности реакции мозга на изменения микробиоты после активации иммунной системы беременной самки мыши (Morais et al., 2018). Изменение генотипа (нокауты и нок-ины) может неодинаково изменять поведение животных с разным генетическим фоном (Löscher et al., 2017; Meng et al., 2018).
Отметим также, что ни в одном исследовании не рассматривался вопрос о различиях во влиянии на поведение крыс “простой” экспозиции серии звуковых воздействий (у животных без АЭ) с поведением животных после судорожных припадков, развивавшихся вследствие такого воздействия. Результаты такого анализа, представленные в настоящей статье, являются в этом отношении приоритетными.
ЗАКЛЮЧЕНИЕ
Суммируя представленные данные, следует отметить, что АЭ, как специфический тип рефлекторно вызванного судорожного состояния, представляет собой достаточно распространенный феномен, который успешно используется в моделировании судорог в интересах клинических исследований. У животных с предрасположенностью к развитию судорог обнаруживаются каналопатии (есть отклонения в строении и функции ионных каналов), причем во многих случаях эта патология оказывается специфичной для определенных отделов мозга, что характерно также и для ряда форм эпилепсии человека. Следуя современной концепции “эндофенотипов” (Gould, Gottesman, 2006), можно с уверенностью говорить о существовании эндофенотипа аудиогенной эпилепсии, дальнейшее подробное исследование которого с большой вероятностью даст много четких данных и по эпилептогенезу в целом, и по особенностям функции ЦНС у животных с АЭ (Полетаева и др., 2017). Важность такого показателя, как генетический фон, при оценке информативности моделей судорожных состояний достаточно велика.
Работа поддержана РФФИ, грант № 18-05-00173 и Госпрограммой № AAA-A16-1160216600515-1.
Список литературы
Полетаева И.И., Костына З.А., Сурина Н.М., Федотова И.Б., Зорина З.А. Генетическая линия крыс Крушинского–Молодкиной как уникальная экспериментальная модель судорожных состояний. Вавил. журн. генет. селек. 2017. 21 (4): 427–434. https://doi.org/10.18699/VJ17.261
Федотова И.Б., Семиохина А.Ф. Изменения аудиогенной эпилепсии и миоклонуса у крыс линии КМ. Журнал высшей нервной деятельности им. И.П. Павлова. 2002. 52 (2): 261–265.
Федотова И.Б., Костына З.А., Сурина Н.М., Полетаева И.И. Cелекция лабораторных крыс на признак “отсутствие предрасположенности к аудиогенной эпилепсии”. Генетика. 2012. 48 (6): 685–691.
Aguilar B.L., Malkova L., N’Gouemo P., Forcelli P.A. Genetically epilepsy-prone rats display anxiety-like behaviors and neuropsychiatric comorbidities of epilepsy. Front. Neurol. 2018. 9:476.https://doi.org/10.3389/fneur.2018.00476
Castro G.P., Medeiros D.C., Guarnieri L.O., Mourão F.A.G., Pinto H.P.P., Pereira G.S., Moraes M.F.D. Wistar audiogenic rats display abnormal behavioral traits associated with artificial selection for seizure susceptibility. Epilepsy Behav. 2017. 71 (Pt B): 243–249. https://doi.org/10.1016/j.yebeh.2015.08.039
Delprato A., Algéo M.P., Bonheur B., Bubier J.A., Lu L., Williams R.W., Chesler E.J., Crusio W.E. QTL and systems genetics analysis of mouse grooming and behavioral responses to novelty in an open field. Genes Brain Behav. 2017. 16 (8): 790–799. https://doi.org/10.1111/gbb.12392
Edut S., Eilam D. Protean behavior under barn-owl attack: voles alternate between freezing and fleeing and spiny mice flee in alternating patterns. Behav Brain Res. 2004. 155 (2): 207–216. https://doi.org/10.1016/j.bbr.2004.04.018
Garcia-Cairasco N., Umeoka E.H.L., Cortes de Oliveira J.A. The Wistar Audiogenic Rat (WAR) strain and its contributions to epileptology and related comorbidities: History and perspectives. Epilepsy Behav. 2017; 71 (Pt B): 250–273. https://doi.org/10.1016/j.yebeh.2017.04.001
Gould T.D., Gottesman I.I. Psychiatric endophenotypes and the development of valid animal models. Genes Brain Behav. 2006 5 (2): 113–9. https://doi.org/10.1111/j.1601-183X.2005.00186
Lee Y., Rodriguez O.C., Albanese C., Santos V.R., Cortes de Oliveira J.A., Donatti A.L.F., Fernandes A., Garcia-Cairasco N., N’Gouemo P., Forcelli P.A. Divergent brain changes in two audiogenic rat strains: A voxel-based morphometry and diffusion tensor imaging comparison of the genetically epilepsy prone rat (GEPR-3) and the Wistar Audiogenic Rat (WAR). Neurobiol Dis. 2018 Mar; 111: 80–90. https://doi.org/10.1016/j.nbd.2017.12.014
Löscher W., Ferland R.J., Ferraro T.N. The relevance of inter- and intrastrain differences in mice and rats and their implications for models of seizures and epilepsy. Epilepsy Behav. 2017. 73: 214–235. https://doi.org/10.1016/j.yebeh.2017.05.040
Meng X.W., Wang J., Ma Q.W. The genetic background and application of Down syndrome mouse models. Yi Chuan. 2018. 40 (3): 207–217. https://doi.org/10.16288/j.yczz.17-279
Morais L.H., Felice D., Golubeva A.V., Moloney G., Dinan T.G., Cryan J.F. Strain differences in the susceptibility to the gut-brain axis and neurobehavioural alterations induced by maternal immune activation in mice. Behav. Pharmacol. 2018. 29 (2 and 3-Spec Issue): 181–198. https://doi.org/10.1097/FBP.0000000000000374
Miguel T.L., Pobbe R.L., Spiacci Jn A., Zangrossi Jn H. Dorsal raphe nucleus regulation of a panic-like defensive behavior evoked by chemical stimulation of the rat dorsal periaqueductal gray matter. Behav. Brain Res. 2010. 213 (2): 195–200. https://doi.org/10.1016/j.bbr.2010.04.055
Poletaeva I.I., Surina N.M., Kostina Z.A., Perepelkina O.V., Fedotova I.B. The rats of Krushinsky-Molodkina strain. Study of audiogenic epilepsy during 65 years. Epilepsy Behav. 2017. 71 (Pt B): 130–141. https://doi.org/10.1016/j.yebeh.2015.04.072
Sarkisova K.Y., Fedotova I.B., Surina N.M., Nikolaev G.M., Perepelkina O.V., Kostina Z.A., Poletaeva I.I. Epilepsy Behav. 2017. 68: 95–102. https://doi.org/10.1016/j.yebeh.2016.12.025
Sun Y., Gooch H., Sah P. Fear conditioning and the basolateral amygdala. F1000 Res. 2020 Jan 28;9: F1000 Faculty Rev-53. https://doi.org/10.12688/f1000research.21201.1
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова