

Журнал высшей нервной деятельности им. И.П. Павлова, 2021, T. 71, № 1, стр. 126-135
Автоматический анализ данных поведенческого теста “водный лабиринт Морриса”
Д. П. Чернюк 1, *, А. Г. Зорин 2, К. З. Деревцова 1, Е. В. Ефимова 3, В. А. Приходько 4, Ю. И. Сысоев 4, 5, О. Л. Власова 1, М. В. Болсуновская 2, И. Б. Безпрозванный 1, 6, **
1 Лаборатория молекулярной нейродегенерации
Санкт-Петербургского политехнического университета Петра Великого
Санкт-Петербург, Россия
2 Лаборатория Промышленные системы потоковой обработки данных ЦНТИ Санкт-Петербургского политехнического университета Петра Великого
Санкт-Петербург, Россия
3 Лаборатория нейробиологии и молекулярной фармакологии, Институт трансляционной биомедицины,
Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
4 Санкт-Петербургский государственный химико-фармацевтический университет
Санкт-Петербург, Россия
5 Лаборатория нейропротезов, Институт трансляционной биомедицины,
Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
6 Отделение физиологии Юго-Западного медицинского центра Техасского университета
Даллас, США
* E-mail: dashachernuk@gmail.com
** E-mail: mnlabspb@gmail.com
Поступила в редакцию 22.06.2020
После доработки 03.09.2020
Принята к публикации 05.10.2020
Аннотация
Поведенческий тест “Водный лабиринт Морриса” является универсальным способом проверки когнитивных функций у подопытных грызунов, особенно он эффективен при выявлении отклонений функций памяти и обучения, что делает его незаменимым при исследовании нейродегенеративных заболеваний и тестировании терапевтических препаратов, направленных на улучшение когнитивных функций. Однако, несмотря на большой диапазон возможных применений, использование данного теста осложняется трудоемким анализом полученных данных. Поэтому на данный момент большую популярность приобретают программы автоматизированного трекинга и анализа движения грызунов. Но все известные качественные программы являются собственностью зарубежных коммерческих компаний и отличаются высокой ценой. Нашей задачей стали разработка и создание доступного качественного отечественного программного продукта, который позволит российским ученым осуществлять исследования с использованием многочисленных конфигураций “Водного лабиринта Морриса”. В данной статье мы подробно описываем процедуру создания нового программного продукта под названием Minopontikos. Мы также провели сравнительный анализ Minopontikos с двумя популярными коммерческими пакетами: VideoMot и EthoVision. Мы показали, что Minopontikos был способен быстро и качественно детектировать траекторию движения животного в воде и определять параметры для оценки когнитивных функций памяти и обучения. По своим характеристикам Minopontikos не уступает коммерческим программам.
ВВЕДЕНИЕ
“Водный лабиринт Морриса” (Morris Water Maze, MWM) является одним из самых популярных поведенческих тестов для изучения процессов формирования и консолидации памяти. Его изобрел британский нейробиолог Ричард Грэм Майкл Моррис (Morris, 1981; Morris, 1984). Идея данного поведенческого теста достаточно проста: животных, обычно крыс или мышей, помещают в большой круговой бассейн с водой, и им необходимо выбраться из воды на скрытую платформу. Новизна данной методики заключалась в том, что пройти этот тест возможно только с использованием пространственной памяти, так как никаких локальных сигналов нет – грызунам необходимо определить по памяти местоположение объекта, который они никогда не видели, не чуяли и не могли услышать. Крысы и мыши являются хорошими пловцами и легко находят скрытую под водой платформу, даже если они перенесли различные поражения мозга (Morris, 1984). Типичные испытания в водном лабиринте ограничены 60–90 с, в то время как испытания в сухом лабиринте часто длятся намного дольше. Это позволяет повысить пропускную способность и эффективность “Водного лабиринта Морриса” (Morgan, 2009). Благодаря своим особенностям MWM стал “золотым стандартом” для изучения пространственной памяти и процессов обучения (Morris, 2008), входит в перечень необходимых тестов для фенотипирования мутантных и трансгенных мышей (Crawley, 2008), а также часто используется в качестве общего анализа когнитивной функции при различных заболеваниях и воздействиях (Brandeis et al., 1989; Nunn et al., 1994; Hsiao et al., 1996; Gallagher, Rapp, 1997; D’Hooge, De Deyn, 2001; Bromley-Brits et al., 2011; Edwards et al., 2014; Wu et al., 2015; Shariatpanahi et al., 2016; Ge et al., 2018). Многочисленные протоколы этого теста настолько чувствительны к изменениям нормальной функции различных областей мозга, не только гиппокампа, что их можно использовать в качестве “индикатора” нормального функционирования когнитивных функций (Bryan et al., 2009). Главным параметром оценки процессов обучения и памяти во всех этих протоколах является латентный период нахождения платформы испытуемым животным. Однако для более объективной оценки поведения и различных когнитивных процессов латентного времени поиска платформы недостаточно (Maei et al., 2009; Morgan, 2009; Tomas Pereira, Burwell, 2015), сама траектория движения подопытного животного может стать очень информативным ресурсом данного поведенческого теста.
Так как в век цифровых технологий визуальная регистрация является достаточно необъективной и неточной методикой изучения поведения, широкое распространение получила автоматическая трассировка перемещения животного (Noldus et al., 2001; Spink et al., 2001; Khotskin, 2016). Значительным плюсом автоматизации видеонаблюдения и анализа являются надежность и последовательность алгоритма, чего сложно достичь при визуальной и ручной обработке данных. На данный момент на российском рынке не существует надежно работающего программного продукта для анализа поведения лабораторных грызунов в водном тесте Морриса, а предлагаемое ПО зарубежных разработчиков отличается высокой ценой.
В этой статье мы представляем инструмент с открытым исходным кодом для автоматизации тестирования “Водного лабиринта Морриса” – Minopontikos. Он написан на языках программирования C++ и Rust и является автономным. Разрабатываемая программа проста в установке и не требует дополнительного программного обеспечения. Хотя на данный момент ПО находится в активной стадии разработки, мы уже имеем отработанный протокол проведения “Водного лабиринта Морриса” и первые результаты, полученные при помощи разрабатываемой нами программы. Целью данной работы является разработка доступного качественного отечественного программного продукта, который позволит российским ученым осуществлять исследования с использованием многочисленных модификаций “Водного лабиринта Морриса”, апробация его в лабораторных условиях и сравнение результатов с двумя из самых популярных коммерческих продуктов – VideoMot компании TSE Systems и EthoVision разработки Noldus.
МЕТОДИКА
В настоящем исследовании использовали 10 мышей линии 5FAD (B6SJLF1/J background): 7 дикого типа (WT) и 3 трансгенные в возрасте 13 мес, инвентарный номер 006554 из Jackson Laboratory. Это исследование проводилось в соответствии с европейскими нормативами по работе с лабораторными животными – директива 2010/63/EU Европейского парламента (directive 2010/63/EU of the European parliament and of the counscil of 22 September 2010) и с соблюдением “Правил проведения работ с использованием экспериментальных животных” (приказ № 775 от 12.08.1977 г. МЗ СССР). Мыши содержались в виварии Лаборатории молекулярной нейродегенерации Санкт-Петербургского политехнического университета Петра Великого с 12-часовым световым циклом при свободном доступе к еде и воде.
Установка для проведения теста представляет собой бассейн для воды, изготовленный из полипропилена, установленный на стальную подставку с регулируемыми опорами (НПК “Открытая Наука” (Open Science), TS1004-M2). Диаметр бассейна 1.5 м, высота стенок 60 см. В комплектацию бассейна входит регулируемая по высоте платформа из белого акрила с металлическим утяжелителем в основании, диаметром 10 см. Навигация для поиска платформы осуществляется при помощи четырех черно-белых ориентиров, расположенных за пределами бассейна и ориентированных в соответствии со сторонами света. Установка дополнительно оборудована цифровой видеосистемой VS 1304-1 с переносным телескопическим штативом. Видеосистема состоит из высокочувствительной цифровой видеокамеры GigE Vision (DMK23GV024) и объектива Fujinon (YV5x2.7R4B-2). Управление видеосъемкой в ходе эксперимента происходит со стационарного компьютера при помощи бесплатного интерфейса Gigabit Ethernet.
Используемый протокол проведения теста “Водный лабиринт Морриса”
Поведенческий тест “Водный лабиринт Морриса” состоит из обучающих сессий, в которых мышь ищет спрятанную (погруженную на 0.5 см в непрозрачную воду) платформу в круглом бассейне. Обучающая сессия состоит из 4 испытаний в день и проводится подряд в течение 4 дней, в одно и то же время в одинаковых условиях в присутствии платформы, последний, 5-й день является тестовым и проводится в отсутствие платформы в 1 испытание. Время каждого испытания составляет 1.5 мин. Сессия проводится в бассейне, наполовину заполненном водой 22°С. Время поиска платформы отсчитывается с момента начала нахождения мыши в бассейне. Опускание мыши в бассейн осуществляется с 4 разных стартовых позиций, головой к стене. Время начала эксперимента – 15:00 ± 1 ч.
Процедура проведения теста
1. Животное опускается в воду в начальном положении. Когда оно достигает платформы, испытание завершается. Животные, не находящие платформу в течение 90 с, направляются к ней и оставляются там на 15 с.
2. Животное помещается в новое место старта, и испытание повторяется (шаг 1), пока животное не выполнит 4 испытания в день. Между испытаниями животному дается перерыв в 15 мин.
3. В последующие дни тренировочные сессии повторяются (пункты 1–2).
4. В последний – тестовый – день проводится пробное испытание без платформы. Животное плавает в течение 1.5 мин в поисках платформы, после чего убирается из бассейна.
Разрабатываемое программное обеспечение
В рамках данной работы было разработано ПО (Minopontikos), работающее в несколько этапов: предварительная обработка входных данных, анализ данных и постобработка. Архитектура ПО показана на рис. 1. Далее подробнее описан каждый этап.
Рис. 1.
Структура программного обеспечения Minopontikos.
Fig. 1. The structure of the Minopontikos software.
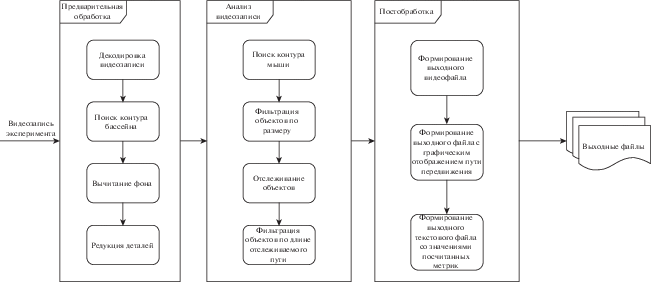
Предварительная обработка данных. Первый этап работы разрабатываемого программного обеспечения (ПО) – предварительная обработка. Заключается она в смене кодировки видеозаписей с DV VIDEO на H.265. Данная функциональность реализована в виде библиотеки, написанной на языке программирования Rust. Для корректной работы данной библиотеки необходимо наличие утилиты ffmpeg.
Для поиска мыши на видеозаписи также необходимы несколько этапов предобработки: определение бассейна для сужения области анализа, вычитание фона для уменьшения ложных обработок движения и размытие деталей, не представляющих интереса для анализа.
Детектирование бассейна. Поиск бассейна на кадре производится с помощью преобразования Хафа (Zivkovic, vd Heijden, 2006). Данный алгоритм использует процедуру голосования для поиска фигур, относящихся к определенному классу. Таким образом, использование данного алгоритма позволяет определить и ограничить анализируемую область размерами бассейна.
Вычитание фона. Один из главных этапов предварительной обработки во многих ПО на основе компьютерного зрения – вычитание фона. Такой этап позволяет удалить статический фон и оставить представляющие интерес динамические объекты. В тесте Морриса за динамический объект принимается мышь, плавающая в бассейне; статическим объектом, соответственно, является бассейн. Следовательно, при вычитании фона остается только передвигающаяся мышь.
В данной работе для вычленения динамического объекта (мыши) из статического фона (бассейн) использовался алгоритм MOG2. Данный алгоритм использует смеси гауссианов. Для постоянного обновления параметров в нем используются рекурсивные уравнения, которые также помогают в одновременном выборе соответствующего количества компонент для каждого пикселя (Zivkovic, 2004; Zivkovic, vd Heijden, 2006).
Редукция деталей. Как упоминалось ранее, для уменьшения влияния посторонних артефактов на анализ видеозаписи можно использовать фильтр размывания Гаусса. Такой фильтр, как видно из названия, относится к размывающим изображения фильтрам и используется в нем функция Гаусса (урав. 1) для подсчета трансформаций, применяемых к каждому пикселю на изображении
(1)
$G(x,y) = \frac{1}{{2\pi {{\sigma }^{2}}}}{{e}^{{ - \frac{{{{x}^{2}} + {{y}^{2}}}}{{2{{\sigma }^{2}}}}}}},$Использование данного фильтра в разрабатываемом ПО позволяет отсечь такие артефакты, как круги на воде и тени от анализируемой мыши.
Обнаружение и отслеживание. После выполнения всех шагов предобработки поиск мыши на кадре сводится к поиску контура. Для целей поиска контура используется алгоритм, предложенный Сатоси Сузуки и Кейчи Эйбом (Abe, 1985). Данный метод основан на прогонных данных, включает в себя трассировку прогонных данных по краям и использует данные прогона в парах, состоящих из левого и правого краев объекта, которые получаются с помощью горизонтального сканирования линий слева направо на изображении.
Несмотря на тот факт, что видимых объектов на видеозаписи не так много, во время обработки появляется множество ложноопределенных объектов. Для их фильтрации отсеиваются найденные объекты неподходящего размера (слишком маленькие и слишком большие) и те, которые выходят за рамки ранее найденного бассейна.
Помимо задачи поиска мыши на кадре необходимо также решить задачу отслеживания передвижения найденной мыши для дальнейшего измерения различных метрик. Для решения данной задачи был использован фильтр Калмана, который оценивает вектор состояния, содержащий параметры цели, такие как положение и скорость, на основе динамической/измерительной модели. Для отслеживания движущихся объектов с использованием фильтра Калмана необходимо разработать динамическую модель движения цели. Наиболее распространенной динамической моделью является модель постоянной скорости (CV), которая предполагает, что скорость является постоянной в течение интервала дискретизации. Эта модель использовалась во многих приложениях благодаря своей универсальности, эффективности и простоте (Saho, 2018). Данный алгоритм позволяет не только отслеживать обнаружение мышей и рисовать их траекторию, но и выбирать мышей из всех обнаруженных объектов. Предполагается, что самый длинный отслеживаемый путь принадлежит наблюдаемым мышам. Когда мыши не могли быть обнаружены на некоторых кадрах в случае окклюзии, используемый алгоритм отслеживания предполагает их размещение и продолжает отслеживание на протяжении небольшого количества кадров.
Для сравнения работы в данном исследовании также были использованы программные продукты VideoMot (TSE Systems) и EthoVision (Noldus et al., 2001).
Статистика. Полученные данные по всем независимым параметрам у всех животных обрабатывали с помощью программы GraphPad Prism. Использование критерия Андерсона–Дарлинга (согласуется с результатами теста Колмогорова–Смирнова) показало, что полученные данные не обладают свойствами нормального распределения, поэтому далее дисперсионный анализ проводили с использованием тестов Манна–Уитни (между двумя выборками) и Краскела–Уоллиса (сравнивая результаты всех трех программ). Также для предварительной оценки равенства дисперсий переменных был использован тест Брауна–Форсайта (для трех выборок) или тест Левена (для двух выборок).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Для сравнительного анализа работы нашего ПО (Minopontikos) было выбрано 30 видеофайлов: четвертое испытание каждой из подопытных мышей на первый день обучения (10 шт.), на последний день обучения (10 шт.) и пробное испытание, проводимое на пятый день поведенческого теста (10 шт.).
Общими параметрами для сравнения были выбраны латентное время нахождения платформы, процентное время нахождения в NW-квадранте, где находилась платформа, угловая ошибка поиска и количество пересечений зоны интереса, где ранее была размещена платформа (для пробного испытания). Результаты обсчета в трех программах представлены в табл. 1.
Таблица 1.
Средние величины значений параметров (± стандартные ошибки), обсчитанные тремя разными программами. Жирным шрифтом выделены статистически значимые различия между значениями параметров по критерию Брауна–Форсайта при p < 0.05 Table 1. Means of parameter values (± standard errors) calculated by three different programs. The bold font indicates statistically significant differences between parameter values according to the Brown–Foresight test, p < 0.05
| Параметры | Результаты обсчета параметров, среднее ± стандартная ошибка (для угловой ошибки приводится модуль) | ||
|---|---|---|---|
| Программы | |||
| VideoMot | EthoVision | Minopontikos | |
| Латентное время нахождения платформы в первый день обучения | 64.29 ± 11.16 | 63.22 ± 11.70 | 64.95 ± 11.20 |
| Латентное время нахождения платформы в последний день обучения | 57.10 ± 8.43 | 63.05 ± 9.56 | 57.69 ± 8.12 |
| Процентное время нахождения в NW-квадранте в первый день обучения | 35.46 ± 9.50 | 33.67 ± 8.88 | 31.55 ± 9.85 |
| Процентное время нахождения в NW-квадранте в последний день обучения | 25.46 ± 2.87 | 28.73 ± 3.15 | 21.95 ± 2.97 |
| Процентное время нахождения в NW-квадранте в тестовый день | 21.48 ± 3.34 | 20.54 ± 3.15 | 13.67 ± 2.80 |
| Угловая ошибка поиска в первый день обучения | 34.20 ± 17.84 | 7.13 ± 28.18 | 39.62 ± 6.68 |
| Угловая ошибка поиска в последний день обучения | 6.70 ± 27.93 | 18.65 ± 26.15 | 43.45 ± 10.84 |
| Угловая ошибка поиска в тестовый день | 1.25 ± 15.10 | 34.04 ± 28.14 | 35.47 ± 11.95 |
| Количество пересечений зоны интереса (где была платформа) в тестовый день | 0.9 ± 0.31 | – | 1.10 ± 0.41 |
Дисперсионный анализ результатов обсчета в трех программах выявил незначительные различия для двух ключевых параметров – латентного времени и процентного времени нахождения в NW-квадранте. Для латентного времени в первый день обучения p = 0.9941, F = 0.0059, в последний день обучения p = = 0.869, F = 0.141 (тест Брауна–Форсайта). Тест Краскела–Уоллиса: для первого дня обучения p = 0.915, для последнего p = 0.646. Для процентного времени нахождения в NW-квадранте бассейна в первый, последний день обучения и в тестовый день результаты следующие: p = 0.958 (F = 0.043), p = 0.295 (F = 1.278), p = 0.171 (F = 1.888) по тесту Брауна–Форсайта и p = 0.716, p = 0.200, p = 0.147 по тесту Краскела–Уоллиса. Результат статистической обработки свидетельствует, что наибольшие различия в обсчете программами данного параметра наблюдаются для последнего дня обучения и в тестовый день, однако эти различия статистически незначимы, что позволяет нам сделать вывод о незначительном различии при подсчете процентного времени во всех трех программах, как и для латентного времени поиска платформы.
Распределение значений латентного времени нахождения подопытными мышами платформы показано на рис. 2.
Рис. 2.
Распределение индивидуальных значений латентного времени поиска скрытой платформы (latency) в первый и в последний день обучений (День 1 и День 4 соответственно), высчитанных при помощи трех различных программ (VideoMot – выделено синим, EthoVision – зеленым, Minopontikos – оранжевым).
Fig. 2. Distribution of individual latency values for the hidden platform search (latency) on the first and last days of training (Day 1 and Day 4, respectively), calculated using three different programs (VideoMot – in blue, EthoVision – in green, Minopontikos – in orange).
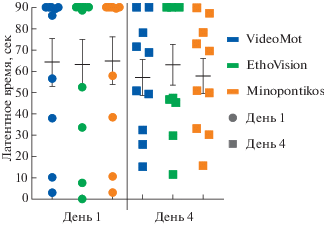
При более детальном анализе обработки видеофайлов программами выяснилось, что для латентного времени поиска платформы различие между двумя коммерческими программами чуть больше, чем между каждой из них и нашим программным продуктом. Так, по критерию Манна–Уитни при сравнении VideoMot и EthoVision p = 0.399, а для Minopontikos p = 0.814 при сравнении с VideoMot и p = 0.500 при сравнении с EthoVision.
Обратная ситуация обнаружилась для параметра процентного времени в квадранте. При сравнении суммарных выборок по всем трем дням тест Краскела–Уоллиса различий не выявил (p = 0.051), однако по критерию Манна–Уитни при сравнении VideoMot и EthoVision p = 0.997, а для Minopontikos p = = 0.032 при сравнении с VideoMot, p = 0.038 при сравнении с EthoVision.
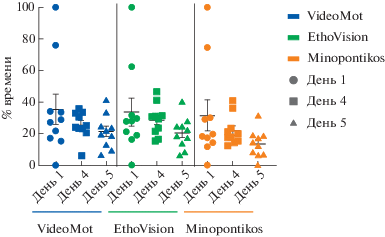
Распределение процентного времени в квадранте NW (где находилась платформа во время обучающих сессий) представлено на рис. 3.
Рис. 3.
Распределение индивидуальных значений процентного времени нахождения в одном из квадрантов бассейна (NW-квадрант, где при обучении находилась скрытая платформа) в первый и в последний день обучений (День 1 и День 4 соответственно), и в тестовый день (День 5), высчитанных при помощи трех различных программ (VideoMot – выделено синим, EthoVision – зеленым, Minopontikos – оранжевым).
Fig. 3. Distribution of individual values of the time spent percentage in one of the pool quadrants (NW quadrant, where the hidden platform was located during training) on the first and last days of training (Day 1 and Day 4, respectively) and on the test day (Day 5), calculated using three different programs ( VideoMot – in blue, EthoVision – in green, Minopontikos – in orange).
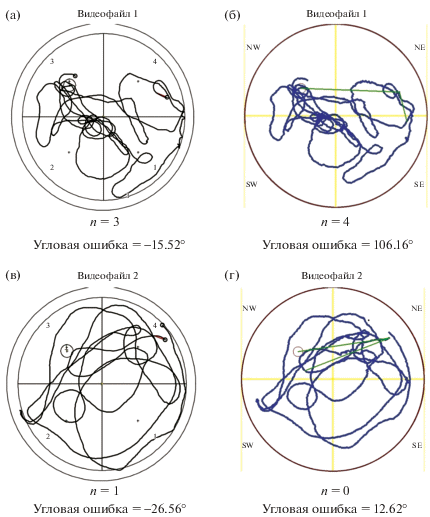
Для тестового испытания кроме процентного времени нахождения в NW-квадранте было подсчитано количество пересечений зоны интереса – места, где при обучении находилась платформа. Данный параметр был нами обнаружен в программе VideoMot, но не в EthoVision. При сравнении результатов обсчета VideoMot и Minopontikos тест Левена показал p = 0.042 (F = 4.261), была выявлена разница только при обсчете двух видео, для которых количество пересечений в VideoMot равно 3 и 1, а в Minopontikos – 4 и 0 соответственно. Иллюстрации траектории движения мышей в данных видео представлены на рис. 4.
Рис. 4.
Траектории движения двух из подопытных мышей (рис. 1 (а, б) – видеофайл 1; рис. 1 (в, г) – видеофайл 2) в тестовый день, детектированные двумя программами (VideoMot – рис. 1 (а, в); Minopontikos – рис. 1 (б, г)). Траектории наложены на контур детектируемой зоны и зоны интереса, где ранее при обучении находилась платформа, также присутствует обозначение квадрантов бассейна (VideoMot – 1, 2, 3, 4; Minopontikos – NW, SW, SE, NE). n – количество пересечений зоны интереса, рассчитанное программами.
Fig. 4. Movement trajectories from two of the experimental mice (fig. 1 (а, б) – video 1; fig. 1 (в, г) – video 2) on the test day, detected by two programs (VideoMot – fig. 1 (а, в); Minopontikos – fig. 1 (б, г)). The trajectories are superimposed on the detection zone contour and the zone of interest, where the platform was previously during training, there is also the designation of the pool quadrants (VideoMot – 1, 2, 3, 4; Minopontikos – NW, SW, SE, NE). n is the number of zone of interest crossings, calculated by the programs.
Статистическая обработка показала, что посчитанная угловая ошибка у всех программ значительно отличается, тест Брауна–Форсайта показал p = 0.0002 (F = 9.247). Это свидетельствует, что случайные различия в значениях данного параметра маловероятны.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные данные свидетельствуют о схожести работы трех программ: двух коммерческих продуктов VideoMot и EthoVision, и разработанной нами программы Minopontikos. Наиболее стабильным параметром для вычисления во всех трех программах является латентное время поиска платформы, полученные нами данные не дают оснований делать вывод о различиях в распределениях этой характеристики. Более того, результаты двух коммерческих продуктов между собой отличаются сильнее, чем результат обсчета нашей программы с каждой из них по отдельности. Также сильных различий не обнаружилось и при расчете процентного времени нахождения в квадранте интереса. Однако более детальный статистический анализ выявил небольшие расхождения при сравнении результатов нашей программы с каждой из коммерческих по отдельности. При сравнении расчета параметра угловой ошибки было выявлено значительное различие результатов всех трех программ, что не дает нам сделать вывод о схожести распределений трех выборок. При более глубоком анализе принципа работы программ выяснилось, что вычисления угловой ошибки во всех программах происходят с использованием различных подходов. Так, в программе VideoMot угол считается между двумя прямыми, соединяющими три точки: платформу, начальное положение животного и положение, которое занимает животное через 10 см пути (количество см можно менять); наша программа за третью точку берет положение мыши через 5% от всего пройденного пути (процент также можно менять). Так как скорость передвижения и длина траектории за одно и то же время у подопытных животных сильно различаются и зависят от индивидуальных особенностей, критерий в процентах от всего пройденного пути нам кажется более объективным, чем конкретное расстояние в единицах измерения длины.
Сравнительный анализ последнего параметра – количества пересечений зоны интереса – в данном исследовании стал возможен только между нашим программным продуктом и VideoMot. Дисперсионный анализ показал различие двух выборок, но более детальное рассмотрение расхождений работы двух программ выявило различие только в двух из десяти видеофайлов. Детектируемые траектории двух программ в этих видео являются практически идентичными (рис. 4), а разница в количестве пересечений обусловлена точностью выбора зоны интереса. В Minopontikos, как и в VideoMot, есть возможность использовать раннее сохраненную зону интереса из других файлов (видео, снятых в обучающих сессиях), которое до этого было выбрано пользователем вручную. А значит, положение выделенной зоны зависит только от человека, использующего программу.
При использовании коммерческих программ и нашего программного продукта выявилось одно интересное отличие, которое, на наш взгляд, является плюсом Minopontikos. Это автоматический отсчет времени начала движения испытуемого грызуна и автоматическое обрезание трека заданным пользователем временным интервалом. При обработке видеофайлов данный подход был более удобен, чем ручная обрезка в коммерческих программах. Кроме того, VideoMot и EthoVision являются универсальными трекинговыми программами для обработки большого набора поведенческих тестов, поэтому их интерфейс очень большой и сильно перегружен, и требует достаточного времени, чтобы разобраться в их функционале.
ЗАКЛЮЧЕНИЕ
В данной статье для автоматизации тестирования MWM были рассмотрены различные методы, которые объединены нами в один инструмент, в отличие от существующих коммерческих и некоммерческих инструментов – открытый и автономный, и создана уникальная программа для трассировки движения мышей и обработки полученных данных. С ее помощью мы получили корректные данные о траектории движения мыши во время процедуры проведения поведенческого теста, а также сравнили их с результатами работы двух самых популярных иностранных коммерческих продуктов: VideoMot и EthoVision. Сейчас наша программа Minopontikos находится в активной стадии разработки, многие возможности еще не реализованы, но полученные в данном исследовании результаты позволяют нам сделать вывод, что имеющиеся уже интерфейс и функционал ничем не уступают популярным коммерческим продуктам.
Публикация подготовлена при поддержке Санкт-Петербургского политехнического университета Петра Великого в рамках программы повышения конкурентоспособности 5-100 (разработка и создание программы Minopontikos) и за счет гранта Российского научного фонда (проект № 20-65-46004) (сравнение Minopontikos с коммерческими продуктами), работа Е. В. Ефимовой выполнялась за счет финансирования СПбГУ ID: 51143531.
Список литературы
Abe S.S.K. Topological structural analysis of digitized binary images by border following. Computer Vision, Graphics, and Image Processing. 1985; 30 (1): 32–46.
Brandeis R., Brandys Y., Yehuda S. The use of the Morris Water Maze in the study of memory and learning. Int J Neurosci. 1989 Sep; 48 (1–2): 29–69.
Bromley-Brits K., Deng Y., Song W. Morris water maze test for learning and memory deficits in Alzheimer’s disease model mice. J Vis Exp. 2011 Jul 20 (53).
Bryan K.J., Lee H., Perry G., Smith M.A., Casadesus G. Transgenic Mouse Models of Alzheimer’s Disease: Behavioral Testing and Considerations. In: Methods of Behavior Analysis in Neuroscience (editor: Buccafusco JJ). 2 edition. Frontiers in Neuroscience. Boca Raton (FL): 2009.
Crawley J.N. Behavioral phenotyping strategies for mutant mice. Neuron. 2008 Mar 27; 57(6): 809–818.
D’Hooge R., De Deyn P.P. Applications of the Morris water maze in the study of learning and memory. Brain Res Brain Res Rev. 2001 Aug; 36 (1): 60–90.
Edwards S.R., Hamlin A.S., Marks N., Coulson E.J., Smith M.T. Comparative studies using the Morris water maze to assess spatial memory deficits in two transgenic mouse models of Alzheimer’s disease. Clin Exp Pharmacol Physiol. 2014 Oct; 41 (10): 798–806.
Gallagher M., Rapp P.R. The use of animal models to study the effects of aging on cognition. Annu Rev Psychol. 1997; 48: 339–370.
Ge M., Zhang Y., Hao Q., Zhao Y., Dong B. Effects of mesenchymal stem cells transplantation on cognitive deficits in animal models of Alzheimer’s disease: A systematic review and meta-analysis. Brain Behav. 2018 Jul; 8(7): e00982.
Hsiao K., Chapman P., Nilsen S., Eckman C., Harigaya Y., Younkin S., Yang F., Cole G. Correlative memory deficits, Abeta elevation, and amyloid plaques in transgenic mice. Science. 1996 Oct 4; 274 (5284): 99–102.
Khotskin N.V., Kulikov V.A., Zavyalov E.L., Fursenko D.V., Kulikov A.V. Conducting and automating Morris water maze test under SPF conditions. Russ. J. Genet. Appl. Res. 2016; 6: 394–399.
Maei H.R., Zaslavsky K., Teixeira C.M., Frankland P.W. What is the Most Sensitive Measure of Water Maze Probe Test Performance? Front Integr Neurosci. 2009. 3: 4.
Morgan D. Water Maze Tasks in Mice: Special Reference to Alzheimer’s Transgenic Mice. In: Methods of Behavior Analysis in Neuroscience (editor: Buccafusco JJ). 2 edition. Frontiers in Neuroscience. Boca Raton (FL): 2009.
Morris R.G.M. Spatial localization does not require the presence of local cues. Learning and Motivation. 1981. 12 (2): 239–260.
Morris R.G.M. Developments of a water-maze procedure for studying spatial learning in the rat Journal of Neuroscience Methods. 1984; 11 (1): 47–60.
Morris R.G.M. Morris water maze. Scholarpedia. 2008. p. 6315.
Noldus L.P., Spink A.J., Tegelenbosch R.A. EthoVision: a versatile video tracking system for automation of behavioral experiments. Behav Res Methods Instrum Comput. 2001 Aug; 33 (3): 398–414.
Nunn J.A., LePeillet E., Netto C.A., Hodges H., Gray J.A., Meldrum B.S. Global ischaemia: hippocampal pathology and spatial deficits in the water maze. Behav Brain Res. 1994 May 30; 62 (1): 41–54.
Rakos D. Efficient Gaussian blur with linear sampling. https://tinyurl.com/398mgyj: RasterGrid Blogosphere; 2010.
Saho K. Kalman Filter for Moving Object Tracking: Performance Analysis and Filter Design. Kalman Filters – Theory for Advanced Applications. 2018.
Shariatpanahi M., Khodagholi F., Ashabi G., Bonakdar Yazdi B., Hassani S., Azami K., Abdollahi M., Noorbakhsh F., Taghizadeh G., Sharifzadeh M. The involvement of protein kinase G inhibitor in regulation of apoptosis and autophagy markers in spatial memory deficit induced by Abeta. Fundam Clin Pharmacol. 2016 Aug; 30(4): 364–375.
Spink A.J., Tegelenbosch R.A., Buma M.O., Noldus L.P. The EthoVision video tracking system–a tool for behavioral phenotyping of transgenic mice. Physiol Behav. 2001 Aug; 73(5): 731–744.
Tomas Pereira I., Burwell R.D. Using the spatial learning index to evaluate performance on the water maze. Behav Neurosci. 2015 Aug; 129 (4): 533–539.
Wu M.N., Zhou L.W., Wang Z.J., Han W.N., Zhang J., Liu X.J., Tong J.Q., Qi J.S. Colivelin ameliorates amyloid beta peptide-induced impairments in spatial memory, synaptic plasticity, and calcium homeostasis in rats. Hippocampus. 2015 Mar; 25 (3): 363–372.
Zivkovic Z. Improved adaptive Gaussian mixture model for background subtraction. Proceedings of the 17th International Conference on Pattern Recognition, ICPR 2004. Cambridge 2004. P. 28–31.
Zivkovic Z., vd Heijden F. Efficient adaptive density estimation per image pixel for the task of background subtraction. Pattern recognition letters. 2006; 27 (7): 773–780.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова