

Журнал высшей нервной деятельности им. И.П. Павлова, 2020, T. 70, № 5, стр. 590-600
Сенсорный гейтинг и сенсорная фасилитация: перспективная парадигма для изучения нарушений непроизвольного внимания
А. А. Варламов 1, 2, *, И. В. Скороходов 2, Г. В. Портнова 3
1 Центр нейрокоммуникативных исследований, Государственный институт русского языка им. А.С. Пушкина
Москва, Россия
2 АНО “Центр реабилитации инвалидов детства “Наш солнечный мир”
Москва, Россия
3 Федеральное государственное бюджетное учреждение науки
Институт высшей нервной деятельности и нейрофизиологии Российской академии наук
Москва, Россия
* E-mail: antonvarlamov@gmail.com
Поступила в редакцию 30.12.2019
После доработки 26.02.2020
Принята к публикации 26.02.2020
Аннотация
Разработка экспериментальных схем для изучения ЭЭГ-маркеров нарушений внимания является одной из актуальных задач в современной нейропсихиатрии. В данной работе был проведен анализ вызванных потенциалов (ВП) и вызванных изменений ритмов ЭЭГ при выполнении задания c редким целевым тоном (2000 Гц, 10%), частым нецелевым тоном (1500 Гц, 80%) и отсутствием физического стимула (10%). Межстимульный интервал (SOA) был фиксированным (1000 мс). Показано, что данное экспериментальное задание позволяет получить высокодостоверные эффекты для компонентов ответа мозга, связанных с фильтрацией сенсорной информации (значительное увеличение амплитуды компонентов ВП P50, N100 и P200 на нецелевые стимулы, следующие после пропущенных, а также сопутствующее кратковременное увеличение амплитуды сигнала в диапазонах от дельта до альфа-2), и выраженный комплекс P50-N100-N200-P300, связанный с произвольным вниманием (реакция на стимул-мишень). Представляется, что подобные экспериментальные схемы можно успешно использовать как для изучения взаимосвязи между различными аспектами внимания, так и в исследованиях биомаркеров нарушений внимания и процессов сенсорной интеграции.
В последние годы мы можем наблюдать значительное увеличение количества исследований, посвященных сенсорным компонентам вызванных потенциалов (ВП), сенсорному гейтингу и в целом фильтрации сенсорной информации. На момент написания статьи поиск в системе Pubmed по запросу ERP sensory gating выдает 857 статей, из них более 400 опубликованы с 2010 г. Это связано с тем, что эффективность фильтрации сенсорной информации является одним из наиболее значимых биомаркеров для многих психических расстройств и нарушений развития, так или иначе связанных с нарушениями внимания, таких как расстройства аутистического спектра [Orekhova, Stroganova, 2014; Matsuzaki et al., 2014], шизофрения [Bramon et al., 2004; De Wilde et al., 2007; Patterson et al., 2008], биполярное расстройство [Lijffijt et al., 2012; Schulze et al., 2007], шизотипическое расстройство личности [Wan et al., 2007; Wang et al., 2004], болезнь Альцгеймера [Thomas et al., 2010] и антисоциальное расстройство личности [Lijffijt et al., 2012], панические расстройства [Ghisolfi et al., 2006] и посттравматическое стрессовое расстройство [Karl et al., 2006].
Наиболее изученный ЭЭГ-коррелят процессов сенсорного гейтинга (sensory gating, встречается также термин “сенсорное дозирование” [Александров и др., 2015]) – снижение амплитуды компонентов P50, N100 и P200 ответа ВП на стимул, предъявленный вскоре после предшествующего стимула (обзор см. [Дмитриева, 2016; Chien et al., 2019]). Считается, что компонент P50 является преимущественно сенсорным и отражает восходящие процессы, связанные с восприятием стимула [Freedman et al., 1987; Jerger et al., 1992; White, Yee, 1997, 2006]; тем не менее такое снижение его амплитуды отражает нисходящие влияния более высоких в иерархическом плане когнитивных процессов и является проявлением тормозных механизмов фильтрации избыточной сенсорной информации, предохраняющих нервную систему от перегрузки [Freedman et al., 1991; Jerger et al., 1992; Wan et al., 2008]. Снижение амплитуды компонентов N100 и P200 может отражать несколько иные компоненты активности мозга [Boutros et al., 2004a, 2004b; Wan et al., 2008], и предположительно связывается, соответственно, с триггером переключения внимания [Наатанен, 1998; Näätänen, Picton, 1987] и с подавлением выделения дополнительных ресурсов внимания на нецелевой стимул [Näätänen, 1992]. В работе М. Лийффийта [Lijffijt et al., 2009] показано, что снижение амплитуды этих компонентов является относительно независимым и связано с различными аспектами внимания и рабочей памяти (см. также [Boutros et al., 2004a, 2004b; Kisley et al., 2004]).
Большая часть работ, посвященных фильтрации сенсорной информации, выполнены на одной экспериментальной парадигме – парадигме двойных щелчков (double click paradigm), в соответствии с классическим вариантом которой идентичные стимулы (короткие щелчки длительностью 40–50 мс) предъявляются парами, при этом расстояние между щелчками в паре составляет 500 мс, а между парами – от 8 до 10 с [Lijffijt et al., 2009]. При этом для второго щелчка в паре наблюдается значительное снижение амплитуды компонентов P50, N100 и P200 по сравнению с первым щелчком. Эта парадигма хорошо стандартизована и активно используется для исследования ЭЭГ-маркеров психических расстройств, связанных с нарушениями фильтрации сенсорной информации, однако ее значительным недостатком является достаточно высокая длительность (как правило, более 15 минут). Обратная ситуация, а именно увеличение амплитуды ответа мозга при увеличении бдительности (vigilance), обусловленном спецификой задания, остается практически неизученной. Мы предположили, что при увеличении бдительности будет наблюдаться сенсорная фасилитация – процесс, обратный сенсорному гейтингу и проявляющийся в увеличении амплитуды компонентов P50, N100 и P200, обусловленный неспецифическим увеличением нисходящей регуляции сенсорных компонентов восприятия.
Для проверки этой гипотезы авторами статьи было использовано экспериментальное задание, сходное с классической парадигмой Т. Пиктона [Пиктон и др., 1974]. Это задание представляет собой вариант парадигмы oddball с фиксированным межстимульным интервалом (1000 мс), частым нецелевым стимулом (тон 1500 Гц, вероятность условия 80%), редким стимулом-мишенью (тон 2000 Гц, вероятность условия 10%) и редкими нецелевыми пропусками стимулов (вероятность условия 10%). В соответствии с экспериментальной гипотезой ожидалось, что для нецелевых стимулов, предъявленных после пропуска ожидаемого стимула, будет наблюдаться увеличение компонентов ВП, связанных с фильтрацией сенсорной информации, причем это увеличение может быть по-разному выражено для компонентов P50, N100 и P200.
МЕТОДИКА
1. Испытуемые. В исследовании приняли участие 26 здоровых испытуемых, из них 7 мужчин и 19 женщин, в возрасте от 19 до 31 года (средний возраст 23.1 года, SD = 2.1 года), имевшие нормальное или скорректированное до нормального зрение, нормальный слух и являющиеся правшами. Испытуемые были предварительно ознакомлены с процедурой и дали согласие на участие в исследовании.
2. Стимулы и экспериментальная процедура. Использовавшееся задание представляет собой вариант парадигмы с редким целевым стимулом (oddball) и с фиксированным межстимульным интервалом (1000 мс). В качестве нецелевого стимула использовался тон частотой 1500 Гц и длительностью 150 мс, в качестве стимула-мишени использовался тон частотой 2000 Гц с той же длительностью. Стимулы предъявлялись через наушники, их громкость составляла 70 дБ. Экспериментальные условия предъявлялись в псевдослучайном порядке со следующей вероятностью: нецелевой стимул 80%, стимул-мишень 10%, пропуск стимула (отсутствие физического стимула) 10%. Единственное ограничение строго случайного порядка заключалось в том, что не допускалось предъявление двух стимулов-мишеней или двух пропусков стимула подряд. Испытуемые располагались с открытыми глазами в удобном кресле в затемненной комнате, во время всего выполнения задания испытуемые смотрели на фиксационный символ “+” в центре экрана 21’ монитора. Всего испытуемым предъявлялось 8 основных блоков по 60 стимулов (всего 480 стимулов) и тренировочный блок из 20 стимулов для ознакомления с заданием, который не включался в анализ данных ЭЭГ. Между блоками делались перерывы длительностью около 10 сек. В соответствии с экспериментальной гипотезой анализировались два экспериментальных условия: нецелевой стимул, предъявленный после нецелевого стимула и нецелевой стимул, предъявленный после неожиданного пропуска стимула.
3. Регистрация ЭЭГ и предварительный анализ данных. ЭЭГ регистрировали с использованием 32-канального усилителя Neuroscan Synamps и 32-канальной шапочки с встроенными Ag/AgCl электродами (названия электродов соответствуют международной схеме 10–10). Для референтного отведения использовали объединенный ушной электрод, заземляющий электрод располагался в центре лба, сопротивление электродов было не более 10 кОм. В качестве первого этапа обработки данных осуществлялась полосовая фильтрация (0.3–45 Гц). Окулографический артефакт удалялся с использованием метода независимых компонент, качество удаления артефакта проверялось экспертом и было устойчиво высоким для всех испытуемых. Для каждого из двух анализируемых экспериментальных условий (нецелевой стимул, предъявленный после нецелевого стимула, нецелевой стимул, предъявленный после неожиданного пропуска стимула) выделялись эпохи ЭЭГ, эпохи брались избыточной длины (от –1300 до 2000 мс от начала предъявления стимула), чтобы исключить влияние краевых артефактов, связанных с фильтрацией и сглаживанием. После этого проводилось дополнительное исключение эпох с миографическими, двигательными и иными артефактами (число исключенных эпох по каждому экспериментальному условию составило не более 20% для каждого испытуемого). При анализе ВП коррекция базового уровня проводилась с использованием предстимульного интервала от –50 мс до 0 мс. Для оценки вызванных изменений ритмов ЭЭГ (ВИР ЭЭГ) использовался модифицированный алгоритм вычисления вызванной синхронизации/десинхронизации [Klimesch et al., 1998b] по следующей схеме: 1) осуществлялась полосовая фильтрация в анализируемых частотных диапазонах (дельта, 2–4 Гц; тета1, 4–6 Гц; тета2, 6–8 Гц; альфа1, 8–10 Гц, альфа2 (10–12 Гц); 2) вычислялся модуль амплитуды ритма (использование модуля вместо традиционного возведения в квадрат позволяет уменьшить влияние высокоамплитудных выбросов и улучшить соотношение сигнал-шум при усреднении; см. тж. [Jongen et al., 2006]); 3) проводилась коррекция базового уровня (по предстимульному интервалу от –200 мс до 0), усреднение и нормализация (вычисление изменений амплитуды ритма в дБ по отношению к предстимульному интервалу от –200 мс до 0), 4) осуществлялось сглаживание полученной кривой с использованием скользящего среднего (ширина окна была равна удвоенной средней частоте для частотного диапазона, напр., для тета1 она составляла 100 мс, а для альфа2 – 45 мс). Для предварительного анализа данных ЭЭГ использовалось программное обеспечение Brainvision Analyser 1.0 и EEGLAB.
4. Статистический анализ данных. Поскольку в нашем задании для усреднения бралось разное количество предъявлений для разных экспериментальных условий, было решено отказаться от поиска пиков компонентов ВП на индивидуальном уровне, так как такой подход способствовал бы завышенным оценкам амплитуды компонентов для условия с меньшим количеством предъявлений. Использовался более строгий критерий: для статистического анализа для каждого испытуемого экспортировались значения временных отчетов в точках, соответствующих пикам компонентов ВП для усредненных кривых для всей выборки (50 мс для Р50, 112 мс для N100 и 200 мс для P200). Тот же подход применялся и при анализе ВИР ЭЭГ. Для оценки достоверности по каждому компоненту и каждому частотному диапазону использовался многофакторный дисперсионный анализ с повторными измерениями ANOVA по схеме Отведение × ЭУ (2) для электродов центральной линии, дальнейшая топография различий уточнялась с использованием post hoс сравнений. Для коррекции достоверности полученных результатов использовалась поправки Гринхауза–Гейссера и Бонферрони. При проведении post hoс сравнений использовался критерий Тьюки. При проведении статистических анализов использовалось программное обеспечение Statistica 6.0.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
1. Анализ вызванных потенциалов
Качественный анализ кривых вызванных потенциалов, усредненных по группе испытуемых, показал, что при предъявлении стимула-мишени наблюдался классический комплекс P50-N100-N200-P300 (P3a, см. [Polich, 2007]), в ответ на все нецелевые стимулы наблюдался комплекс компонентов P50-N100-P200-N350 с последующим возвратом к базовому уровню приблизительно через 600 мс с момента предъявления стимула, а в ответ на пропуск стимула выраженные компоненты ВП не формировались (рис. 1). Дополнительное сравнение кривых ВП для нецелевых стимулов, предъявленных после нецелевых стимулов и для нецелевых стимулов, предъявленных после пропусков стимула, показало, что амплитуда компонентов P50, N100 и P200 была значительно выше для нецелевых стимулов, следующих после пропущенных (рис. 2).
Рис. 1.
Усредненные кривые ВП для нецелевых стимулов, стимулов-мишеней и пропусков, предъявленных после нецелевых стимулов (отведение Fz), и карты компонентов ВП для стимулов-мишеней.
Fig. 1. ERP grand averages for nontargets, targets and stimulus omissions (Fz electrode) with maps for ERP components for target condition.
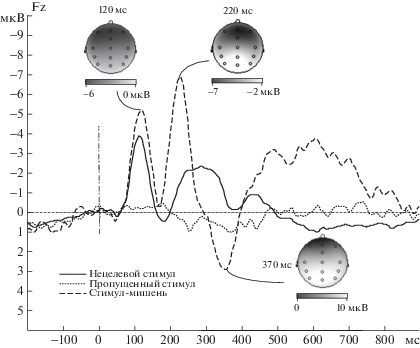
Рис. 2.
Усредненные кривые ВП для нецелевых стимулов, предъявленных после нецелевых стимулов и после пропусков (отведение Сz), и карты компонентов ВП для нецелевых стимулов, предъявленных после пропусков.
Fig. 2. ERP grand averages for nontarget-nontarget condition and for omission-nontarget condition (Cz electrode), maps for omission-nontarget condition.
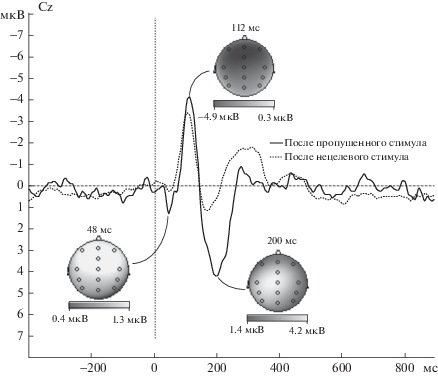
Анализ ANOVA для амплитуды компонента P50 для электродов средней линии (Отведение 6 (Fz, FCz, Cz, CPz, Pz, Oz) × ЭУ 2) показал наличие эффекта экспериментального условия (F(1, 25) = 19.54, p < 0.0002), свидетельствующего о том, что амплитуда компонента Р50 для нецелевых стимулов, следующих после пропущенных, была достоверно выше, чем для нецелевых стимулов, следующих после нецелевых (рис. 2). Отсутствие достоверности взаимодействия Отведение × ЭУ (p > 0.1) показало, что наблюдавшийся эффект был выражен для всех электродов средней линии. Анализ средних показал, что, как и ожидалось, различия были несколько более выражены для фронтальных и центральных электродов.
Анализ ANOVA для амплитуды компонента N100 для электродов средней линии показал схожие результаты: наличие чуть менее выраженного эффекта экспериментального условия (F(1, 25) = 11.30, p < 0.003) и отсутствие достоверности взаимодействия Отведение × ЭУ показали увеличение амплитуды компонента N100 для нецелевых стимулов, следующих после пропущенных (см. рис. 2).
Анализ ANOVA для амплитуды компонента Р200 для электродов средней линии показал, что увеличение амплитуды Р200 после пропуска стимулов по-разному проявлялось для различных отведений: были обнаружены как высокодостоверный эффект экспериментального условия (F(1, 25) = 47.97, p < < 0.00001), так и взаимодействие Отведение × × ЭУ (F(5, 125) = 5.25, p < 0.0003). Анализ полученного взаимодействия показывает, что различия были высокодостоверны для всех отведений, но выраженность их была выше для фронтоцентральных и центральных отведений (рис. 3).
Рис. 3.
Средние значения и стандартные ошибки амплитуды компонента P200 для различных ЭУ и отведений срединной линии.
Fig. 3. Means and SEs for P200 amplitude for midline electrodes.
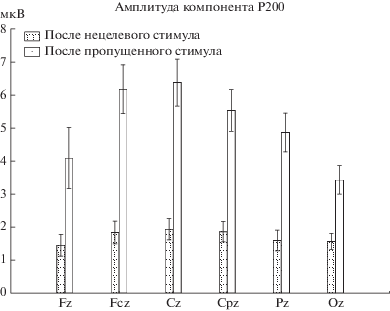
Полученные нами данные подтвердили экспериментальную гипотезу: амплитуда всех анализируемых компонентов ВП была существенно выше для нецелевых стимулов, следующих после пропущенных; наибольшие различия (более чем в три раза по амплитуде) наблюдались для компонентов P50 и P200.
2. Анализ вызванных изменений ритмов ЭЭГ
Во всех проанализированных частотных диапазонах наблюдались высокодостоверные эффекты экспериментального условия, подтверждающие нашу экспериментальную гипотезу (все p < 0.00005): вызванное увеличение амплитуды ритмов ЭЭГ для нецелевых стимулов, следующих после пропущенных, было достоверно более выражено, чем для нецелевых стимулов, следующих после нецелевых.
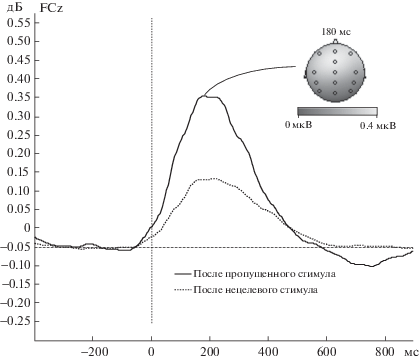
Кроме того, в тета1-, тета2-, альфа1- и альфа2-диапазонах наблюдалось достоверное взаимодействие Отведение × ЭУ (все p < 0.005), свидетельствующее о том, что этот эффект был больше выражен для префронтальных отведений (динамику и топографию такого увеличения амплитуды в тета1-диапазоне см. на рис. 4).
Рис. 4.
Усредненные кривые вызванных изменений амплитуды тета1-ритма (4–6 Гц) для различных ЭУ (отведение FСz).
Fig. 4. Event-related oscillations for lower theta band (4–6 Hz) for nontarget-nontarget condition and for omission-nontarget condition (FСz electrode).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Сенсорная фасилитация: увеличение амплитуды компонентов комплекса P50-N100-P200. Полученные результаты полностью подтверждают исходную экспериментальную гипотезу и указывают на то, что пропуск ожидаемого стимула приводит к значительной амплификации ответа мозга при восприятии последующего стимула. На наш взгляд, этот эффект может объясняться сочетанием двух причин.
1. В рамках классической интерпретации увеличение амплитуды компонентов ВП и ритмической активности ЭЭГ может обусловливаться увеличением межстимульного расстояния [Boutros et al., 2004; Kisley et al., 2004]. Действительно, расстояние до предшествующего стимула для стимулов, следующих после пропусков, увеличено в два раза и составляет две секунды вместо одной. Однако, как нам кажется, такое увеличение межстимульного расстояния само по себе не способно объяснить выраженность наблюдаемого увеличения компонентов ответа, связанных с фильтрацией сенсорной информации. В нашем случае соотношение межстимульного расстояния составляет 1:2, в то время как в классической парадигме двойных щелчков, обеспечивающей сопоставимую разницу в амплитуде компонентов ВП, соотношение межстимульного расстояния составляет 1:16.
2. Еще одной причиной увеличения амплитуды комплекса P50-N100-P200 может являться временное увеличение уровня внимания (бдительности, vigilance), связанное с предшествующим нарушением ожиданий. Это хорошо укладывается в рамки теории предсказывающего кодирования (predictive coding), сформулированной К. Фристоном и описывающей процессы формирования ожиданий и реакции на нарушение ожиданий [Friston, 2005; Arnal, Giraud, 2012]. Можно предположить, что при наличии фиксированного небольшого межстимульного интервала у человека формируются устойчивые ожидания относительно времени предъявления следующего стимула. Это наблюдается как в нашем задании для последовательности стимулов, так и в парадигме двойных щелчков для второго стимула, который каждый раз подается на фиксированном расстоянии в 500 мс от первого щелчка. Это предположение достаточно легко проверить, проведя дополнительные исследования с варьирующимся уровнем однозначности ожиданий и изменяющимся межстимульным интервалом. Такое предположение косвенно подкрепляется исследованиями, установившими, что выраженность эффекта сенсорного гейтинга положительно коррелирует с амплитудой негативности рассогласования [Kisley et al., 2004], которая является, пожалуй, наиболее изученным компонентом ВП, связанным с нарушением ожиданий [Наатанен, 1998]. Следует особо отметить, что наибольший эффект усиления был получен для компонента Р200. Этот компонент, как правило, связывается с тормозным нисходящим воздействием, предотвращающим выделение излишних ресурсов внимания для обработки нерелевантного стимула [Наатанен, 1998]. Увеличение амплитуды этого компонента логично вытекает из контекста задания: если предположить, что увеличение амплитуды более ранних компонентов P50 и N100 отражает привлечение дополнительного внимания к обработке стимула, в дальнейшем это должно компенсироваться более активным “вытормаживанием” нерелевантного стимула с целью экономии ресурса внимания.
Сенсорная фасилитация: вызванные изменения ритмов ЭЭГ. Анализ вызванных изменений амплитуды низко- и среднечастотных ритмов ЭЭГ также свидетельствует о сенсорной фасилитации и привлечении больших ресурсов для обработки информации в ситуации увеличения внимания. Этот эффект кратковременного увеличения амплитуды (“вызванной синхронизации”) хорошо описан в литературе и свидетельствует об увеличении когнитивной нагрузки или уровня физиологической активации при обработке эмоционально или контекстно значимых стимулов [Афтанас и др., 2002; Афтанас и др., 2003]. Кратковременное увеличение амплитуды свидетельствует об увеличении активации не только для низкочастотных ритмов, но и для альфа-диапазона; этот эффект получил название парадоксальной альфа-синхронизации [Cooper et al., 2006]. В продолжение обсуждения эффекта вызванного увеличения амплитуды ритмической активности в широком частотном диапазоне следует отметить прежде всего высокую статистическую силу наблюдаемых эффектов и высокую достоверность полученных различий. Это позволяет предположить, что данные показатели могут также быть использованы в качестве ЭЭГ-маркеров нарушений внимания для широкого круга психических расстройств. Действительно, ряд работ уже показал информативность ЭЭГ-маркеров, связанных с вызванными изменениями амплитуды ритмов, при исследованиях тревожности, алекситимии, шизофрении и СДВГ (обзор см. [Афтанас и др., 2003]). Кроме того, результаты ряда работ показывают, что фазосвязанная активность (evoked activity, т.е. вызванные потенциалы) и фазонезависимая активность (induced activity, вызванные изменения ритмов ЭЭГ) могут представлять собой относительно независимые компоненты ответа мозга [Cooper et al., 2006]. Как известно, при использовании совокупности относительно независимых предикторов, устойчиво связанных с определенным состоянием или устойчивой характеристикой объекта, точность классификации с использованием алгоритмов машинного об-учения резко возрастает по сравнению с моделями, основанными на меньшем количестве предикторов [Варламов, Стрелец, 2013; Iosifescu, 2011]. Это позволяет надеяться на то, что одновременное использование ВП-маркеров и ВИР-маркеров нарушений внимания может оказаться перспективным не только для изучения механизмов этих нарушений, но и при использовании в клинической практике для уточнения диагноза и при выборе схемы коррекции (см., например, рандомизованное исследование, показавшее эффективность ЭЭГ-маркеров депрессии при выборе фармакологического воздействия [DeBattista et al., 2011].
Общие характеристики экспериментального задания и возможности его использования. В данной работе использовалась активная версия задания oddball task, позволяющая оценить особенности произвольного внимания (комплекс P50-N100-N200-P300, наблюдающийся в ответ на стимул-мишень, рис. 1) и особенности непроизвольного внимания, связанные с фильтрацией сенсорной информации (комплекс компонентов P50-N100-P200, см. рис. 2). По сравнению с классической парадигмой двойных щелчков, которая в настоящее время является “золотым стандартом” для изучения сенсорного гейтинга, разработанная нами экспериментальная схема отличается следующими преимуществами:
а) существенно меньшая длительность (8–9 мин по сравнению с 14–20 мин[Lijffijt et al., 2009]);
б) возможность оценить в рамках одного краткого задания не только комплекс компонентов P50-N100-P200, связанный с сенсорным гейтингом, но и особенности произвольного внимания и ответа на стимул-мишень (компоненты N200 и P3a) для активной версии задания, либо особенности непроизвольного внимания на отклоняющийся стимул (предположительно, компоненты N200 и P3b; см. [Polich, 2007]) для пассивной версии задания;
в) межстимульный интервал оптимизирован для анализа вызванных изменений ритмов ЭЭГ и позволяет оценивать вызванные изменения ритмов в ответ на каждое экспериментальное условие без контаминации ответом на предшествующий стимул, в то время как в парадигме двойных щелчков ранние компоненты ответа на второй щелчок накладываются на поздние компоненты ответа на первый щелчок, что может затруднять интерпретацию полученных результатов.
Описанные особенности делают подобные экспериментальные схемы перспективными вариантами для оценки особенностей произвольного и непроизвольного внимания при широком круге нарушений развития и психических расстройств. Представляется, что активная версия задания, использованная в данной работе, будет наиболее информативна для исследования психических расстройств (антисоциальное расстройство личности), нарушений развития (синдром дефицита внимания и гиперактивности) и иных состояний, для которых характерно нарушение волевого контроля и произвольности внимания. Пассивная версия задания, не требующая ответа на редкий отклоняющийся стимул, может быть использована для оценки особенностей непроизвольного внимания у детей и взрослых с расстройствами аутистического спектра, шизофренией и иными психическими расстройствами, для которых характерно нарушение сенсорного гейтинга и других когнитивно-перцептивных процессов, а также у пациентов со сниженным уровнем сознания. Кроме того, отдельно следует отметить, что описанный нами феномен сенсорной фасилитации может исследоваться в рамках кросс-модальных экспериментальных схем (слуховые и зрительные стимулы, слуховые и тактильные стимулы и т.д.) для исследования нарушения сенсорной интеграции при расстройствах аутистического спектра и СДВГ [Withrow, 2007; Doumas et al., 2016; Panagiotidi et al., 2018].
Использование подобных экспериментальных схем может также быть продуктивным в исследовании общих механизмов формирования ожиданий в рамках теории предсказывающего кодирования. В настоящее время уже достаточно полно описан ответ мозга на стимулы разных модальностей, соответствующие ожиданиям (например, комплекс компонентов P50-N100-P200), и на стимулы, нарушающие ожидания или связанные с новизной (такие компоненты и комплексы компонентов, как негативность рассогласования (mismatch negativity, MMN) [Наатанен, 1998], N1 [Arnal, Giraud, 2012] и комплекс N2-P3a [Polich, 2007; Folstein, Van Petten, 2008]), однако эти исследования проводились в рамках различных экспериментальных схем и в какой-то степени в рамках различных концептуальных подходов. Нам представляется, что дальнейшие исследования в этом направлении могли бы способствовать более полному пониманию характера взаимоотношений восходящих и нисходящих влияний при обработке сенсорной информации.
ЗАКЛЮЧЕНИЕ
Полученные результаты позволяют однозначно утверждать, что пропуск ожидаемого стимула приводит к выраженной амплификации ответа мозга на последующий стимул, проявляющейся как в увеличении амплитуды комплекса P50-N100-P200, так и в вызванном увеличении амплитуды ритмов ЭЭГ в широком частотном диапазоне, от дельта-ритма до высокочастотного альфа-ритма. Это позволяет использовать данное задание для изучения процессов, связанных с фильтрацией сенсорной информации; можно также предположить, что при различных вариантах нарушений внимания эффекты амплификации сенсорных компонентов и подавления сенсорных компонентов могут быть выражены в разной степени. Кроме того, использованное в данной работе экспериментальное задание предоставляет возможность одновременно оценивать не только сохранность механизмов фильтрации сенсорной информации, но и механизмов избирательного внимания (реакция на стимул-мишень); при этом данное экспериментальное задание отличается простотой, небольшой продолжительностью и возможностью использования в пассивном варианте. В связи с этим предложенное задание может успешно использоваться для комплексных исследований нарушений внимания и устойчивых индивидуальных характеристик, отражающих различные аспекты избирательного и непроизвольного внимания.
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 18-00-01670 (18-00-01616).
Список литературы
Александров А.А., Дмитриева Е.С., Станкевич Л.Н. Корреляция между дозированием сенсорной информации, негативностью рассогласования и временем реакции. Журн. высш. нервн. деят. им. И.П. Павлова. 2015. 65 (5): 626–634.
Афтанас Л.И., Варламов А.А., Павлов С.В., Рева Н.В., Махнев В.П. Отражение знака эмоции в эффектах вызванной синхронизации и десинхронизации ЭЭГ. 2002. Рос. физиол. журн. 88 (6): 790–802.
Варламов А.А., Стрелец В.Б. Анализ когерентности ЭЭГ при депрессивных расстройствах: современное состояние и перспективы клинического применения. Журн. высш. нервн. деят. им. И.П. Павлова. 2013. 63 (6): С. 613–625.
Дмитриева Е.С. Исследование механизмов сенсорного дозирования (сенсорный гейтинг) с помощью слуховых вызванных потенциалов мозга человека. Дис. Санкт-Петербург: [СПБГУ]. 2016.
Наатанен Р. Внимание и функции мозга: Учеб. Пособие. Пер. с англ. Под ред. Е.Н. Соколова. М.: Изд-во МГУ, 1998. 559 с.
Пиктон Т.В., Хиллард С.Н., Галамбос Р. Корковые вызванные ответы на пропущенные стимулы. Основные проблемы электрофизиологии головного мозга. М.: Наука, 1974: 302–312.
Aftanas L.I., Varlamov A.A., Pavlov S.V., Makhnev V.P., Reva N.V. Time-dependent cortical asymmetries induced by emotional arousal: EEG analysis of event-related synchronization and desynchronization in individually defined frequency bands. 2002. Int J Psychophysiol. 44 (1): 67–82.
Arnal L.H., Giraud A.L. Cortical oscillations and sensory predictions. Trends Cogn Sci. 2012. 16 (7): 390-8.
Boutros N.N., Korzyukov O., Jansen B., Feingold A., Bell M. Sensory gating deficits during the mid-latency phase of information processing in medicated schizophrenic patients. Psychiatry Research. 2004a. 126: 203–215.
Boutros N.N., Korzyukov O., Oliwa G., Feingold A., Campbell D., McClain-Furmanski D., Struve F., Jansen B.H. Morphological and latency abnormalities of the mid-latency auditory evoked responses in schizophrenia: A preliminary report. Schizophrenia Research. 2004b. 70 (2-3): 303–313.
Bramon E., Rabe-Hesketh S., Sham P., Murray R.M., Frangou S. Meta-analysis of the P300 and P50 waveforms in schizophrenia. Schizophrenia Research. 2004. 70 (2–3): 315–329.
Chien Y.L., Hsieh M.H., Gau S.S.F. P50-N100-P200 sensory gating deficits in adolescents and young adults with autism spectrum disorders. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2019. 95: 109683.
Cooper N.R., Burgess A.P., Croft R.J., Gruzelier J.H. Investigating evoked and induced electroencephalogram activity in task-related alpha power increases during an internally directed attention task. Neuroreport. 2006. 17 (2): 205–208.
De Wilde O.M., Bour L.J., Dingemans P.M., Koelman J.H.T.M., Linszen D.H. A meta-analysis of P50 studies in patients with schizophrenia and relatives: Differences in methodology between research groups. Schizophrenia Research. 2007. 97 (1–3): 137–151.
DeBattista C., Kinrys G., Hoffman D., Goldstein C., Zajecka J., Kocsis J., Teicher M., Potkin S., Preda A., Multani G., Brandt L., Schiller M., Iosifescu D., Fava M. The use of referenced EEG (rEEG) in assisting medication selection for the treatment of depression. Journal of psychiatric research. 2011. 45 (1): 64–75.
Doumas M., McKenna R., Murphy B. Postural control deficits in autism spectrum disorder: the role of sensory integration. Journal of autism and developmental disorders. 2016. 46 (3): 853–861.
Folstein J.R., Van Petten C. Influence of cognitive control and mismatch on the N2 component of the ERP: a review. Psychophysiology. 2008. 45 (1): 152–170.
Freedman R., Adler L.E., Gerhardt G.A., Waldo M.C., Baker N., Rose G.M., Drebing C., Nagamoto H., Bickford-Wimer P., Franks R. Neurobiological studies of sensory gating in schizophrenia. Schizophrenia Bulletin. 1987. 13 (4): 669–678.
Freedman R., Waldo M., Bickford-Wimer P., Nagamoto H. Elementary neuronal dysfunction in schizophrenia. Schizophrenia Research. 1991. 4 (2): 233–243.
Friston K. A theory of cortical responses. Philos. Trans. R. Soc. Lond. B: Biol. Sci. 2005. 360 (1456): 815–836.
Ghisolfi E.S., Heldt E., Zanardo A.P., Strimitzer I.M. Jr., Prokopiuk A.S., Becker J., Cordioli A.V., Manfro G.G., Lara D.G. P50 sensory gating in panic disorder. Psychiatry Research. 2006. 40 (6): 535–540.
Iosifescu D.V. Electroencephalography-derived biomarkers of antidepressant response. Harvard review of psychiatry. 2011. 19 (3): 144–154.
Jerger K., Biggins C., Fein G. P50 suppression is not affected by attentional manipulations. Biological Psychiatry 1992. 31 (4): 365–377.
Jongen E.M., Smulders F.T., van Breukelen G.J. Varieties of attention in neutral trials: linking RT to ERPs and EEG frequencies. Psychophysiology. 2006. 43 (1): 113–25.
Karl A., Malta L.S., Maercker A. Meta-analytic review of event-related potential studies in post-traumatic stress disorder. Biological Psychology. 2006. 71 (2): 123–147.
Kisley M.A., Noecker T.L., Guinther P.M. Comparison of sensory gating to mismatch negativity and self-reported perceptual phenomena in healthy adults. Psychophysiology. 2004. 41 (4): 604–612.
Klimesch W., Russegger H., Doppelmayr M., Pachinger T. A method for the calculation of induced band power: Implications for the significance of brain oscillations. 1998b. Electroencephalography and Clinical Neurophysiology. 108 (2): 123–130.
Lijffijt M., Lane S.D., Meier S.L., Boutros N.N., Burroughs S., Steinberg J.L., Moeller F.G., Swann A.C. P50, N100 and P200 sensory gating: relationships with behavioral inhibition, attention and working memory. Psychophysiology. 2009. 46 (5): 1059–1068.
Lijffijt M., Cox B. Acas M.D., Lane S.D., Moeller F.G., Swann A.C. Differential relationships of impulsivity or antisocial symptoms on P50, N100, or P200 auditory sensory gating in controls and antisocial personality disorder. Journal of psychiatric research. 2012. 46 (6): 743–750.
Matsuzaki J., Kagitani-Shimono K., Sugata H., Hirata M., Hanaie R., Nagatani F., Taniike M. Progressively increased M50 responses to repeated sounds in autism spectrum disorder with auditory hypersensitivity: a magnetoencephalographic study. PLoS One. 2014. 9 (7): e102599.
Näätänen R., Picton T.W. The N1 wave of the human electric and magnetic response to sound: A review and an analysis of the component structure. Psychophysiology. 1987. 24 (4): 375–425.
Näätänen R. Attention and Brain Function. Hillsdale, New Jersey: Lawrence Erlbaum Associate. 1992. 494 p.
Orekhova E.V., Stroganova T.A. Arousal and attention re-orienting in autism spectrum disorders: evidence from auditory event-related potentials. Frontiers in Human Neuroscience. 2014. 8: 34.
Panagiotidi M., Overton P.G., Stafford T. The relationship between ADHD traits and sensory sensitivity in the general population. Comprehensive psychiatry. 2018. 80: 179–185.
Patterson J.V., Hetrick W.P., Boutros N.N., Jin Y., Sandman C., Stern H., Potkin S., Bunney Jr, W.E. P50 sensory gating ratios in schizophrenics and controls: A review and data analysis. Psychiatry Research. 2008. 158 (2): 226–247.
Polich J. Updating P300: an integrative theory of P3a and P3b. Clinical neurophysiology. 2007. 118 (10): 2128–2148.
Schulze K.K., Hall M.H., McDonald C., Marshall N., Walshe M., Murray R.M., Bramon E. P50 auditory evoked potential suppression in bipolar disorder patients with psychotic features and their unaffected relatives. Biological Psychiatry. 2007. 62 (2): 121–128.
Thomas C., vom Berg I., Rupp A., Seidl U., Schröder J., Roesch-Ely D., Kreisel S.H., Mundt C., Weisbrod M. P50 gating deficit in Alzheimer dementia correlates to frontal neuropsychological function. Neurobiology of aging. 2010. 31 (3): 416–424.
Wan L., Friedman B.H., Boutros N.N., Crawford H.J. P50 sensory gating and attentional performance. International Journal of Psychophysiology. 2008. 67 (2): 91–100.
Wang J., Miyazato H., Hokama H., Hiramatsu K., Kondo T. Correlation between P50 suppression and psychometric schizotypy among non-clinical Japanese subjects. International Journal of Psychophysiology. 2004. 52 (2): 147–157.
White P.M., Yee C.M. Effects of attentional and stressor manipulations on the P50 gating response. Psychophysiology. 1997. 34 (6): 703–771.
White P.M., Yee C.M. P50 sensitivity to physical and psychological state influences. Psychophysiology. 2006. 43 (3): 320–328.
Withrow R.L. Sensory Integration Dysfunction: Implications for Counselors Working with Children. Journal of School Counseling. 2007. 5 (18): n18.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова