

Журнал высшей нервной деятельности им. И.П. Павлова, 2020, T. 70, № 5, стр. 635-649
Реактивность зеркальной системы мозга и уровень интеллекта у детей школьного возраста
С. А. Махин 1, *, А. И. Кайда 1, Е. В. Эйсмонт 1, В. Б. Павленко 1
1 Крымский федеральный университет им. В.И. Вернадского
Симферополь, Россия
* E-mail: smakhin@inbox.ru
Поступила в редакцию 29.04.2020
После доработки 14.05.2020
Принята к публикации 01.06.2020
Аннотация
Анализировали различия в показателях вербального и невербального интеллекта (тест Векслера – WISC) у 98 детей в зависимости от принадлежности к возрастной группе (младшая – 8–11 лет/старшая – 12–15 лет), сенсомоторной специфичности при выполнении инструментальных движений компьютерной мышью (снижение амплитуды мю-ритма ЭЭГ в низкочастотном/высокочастотном поддиапазонах) и наличия подавления мю-ритма при наблюдении и слуховом восприятии аналогичных движений, выполняемых другим человеком. В ситуации наблюдения за движениями отсутствие снижения амплитуды мю-ритма во фронтальных регионах коры (т.е. отсутствие “зеркальной” активации) у детей старшей группы с высокочастотным мю-ритмом было характерно для испытуемых с более низким уровнем развития невербального интеллекта. Отсутствие “зеркальной” активации в центральных регионах выявлено у детей обеих возрастных групп с высокочастотным мю-ритмом, имеющих более низкие показатели невербального интеллекта, а в париетальных – высокий уровень вербального интеллекта. В ситуации слухового восприятия движений менее выраженная “зеркальная” активация была обнаружена у детей с низкочастотным мю-ритмом, отличающихся более высоким уровнем невербального интеллекта. Таким образом, характер связи реактивности зеркальной системы мозга с уровнем интеллекта школьников зависит от их возраста, частотных особенностей мю-ритма, анализируемого региона коры и экспериментальной ситуации, в которой регистрируется динамика ритмов ЭЭГ.
ВВЕДЕНИЕ
В ЦНС человека принято выделять ряд сетей нейронного взаимодействия в связи с соответствующей им функциональной специализацией [Курганский, 2018; Tompson et al., 2020]. В последние два десятилетия одной из наиболее обсуждаемых в научном сообществе тем является вопрос о наличии у человека и функциональной роли так называемой зеркальной системы мозга (ЗСМ), сходной с обнаруженной первоначально у обезьян системой зеркальных нейронов [di Pellegrino et al., 1992]. К настоящему времени накоплен значительный объем экспериментальных данных, говорящих в пользу гипотезы о существовании человеческого аналога такой системы, ключевой особенностью которой является способность реагировать сходным образом на процессы реализации собственных действий и наблюдение за аналогичными действиями, выполняемыми другими [Molenberghs et al., 2012]. Наиболее популярны гипотезы о вовлеченности ЗСМ в процессы распознавания целей (и намерений), стоящих за поведением окружающих [Gallese, Sinigaglia, 2011; Mottonen et al., 2016], подражания [Wadsworth et al., 2017; Binder et al., 2017], эмпатии [Krautheim et al. 2019; Jospe et al., 2020], а также развития и функционирования человеческой способности к языковой коммуникации [Glenberg, Gallese, 2012; Bidet-Ildei et al., 2020]. Другими словами, функционально работа данной системы связывается с реализацией различных аспектов социального познания [Лебедева и др., 2017; Alcala-Lopez et al., 2019]. Принимая во внимание тот факт, что когнитивное развитие ребенка происходит в тесном взаимодействии со сверстниками и взрослыми, представляется закономерным предположить, что индивидуальные особенности функционирования ЗСМ могут быть связаны как с особенностями развития отдельных когнитивных способностей, так и с показателями его общего интеллектуального развития. Несмотря на правдоподобность данной гипотезы, в тематической научной литературе исследования такого рода практически не представлены. Косвенно на такую возможность могут указывать, например, свидетельства о вовлеченности “зеркальных” корковых механизмов в процессы восприятия временных интервалов взрослыми людьми [Bushov et al., 2019].
В контексте изучения мозговой организации общей интеллектуальной способности человека наибольшую эмпирическую поддержку получила так называемая теория парието-фронтальной интеграции [Jung, Haier, 2007]. Согласно ее ключевым положениям, более высокий уровень интеллекта обеспечивается процессами более скоростной и точной передачи информации между теменными и лобными участками коры. В период взросления связи между этими областями коры постепенно развиваются и усложняются [Power et al., 2010]. Показано, что уже у детей 6–8 лет более высокому уровню невербального интеллекта соответствует более высокая степень функциональной связанности теменных и лобных участков коры правого полушария в состоянии покоя [Langeslag et al., 2013]. При изучении взрослых испытуемых показано, что уровень вербального интеллекта может зависеть от особенностей взаимодействия между теменной корой и мозжечком [Chen, Desmond, 2005; Macher et al., 2014].
Важно указать, что классические области ЗСМ человека также располагаются в лобных и теменных участках коры [Риццолатти, Синигалья, 2012]. В “расширенную” версию данной системы дополнительно включают также первичные сенсомоторные области [Pineda, 2008]. Совместная активация упомянутых областей при наблюдении за действиями других людей рассматривается как связанная с функционированием ЗСМ и может быть зарегистрирована в форме реакции падения амплитуды (так называемой десинхронизации) разновидности альфа-активности ЭЭГ – сенсомоторного мю-ритма в центральных и прилегающих к ним областях коры. Ключевое свойство мю-ритма, заключающееся в снижении его амплитуды в ответ на произвольное осуществление, представление и наблюдение моторных действий, позволяет рассматривать его как один из возможных маркеров активации ЗСМ человека [Fox et al., 2016; Лебедева и др., 2018].
Наблюдение за чужими действиями вызывает, как правило, менее локализованную, по сравнению с условием выполнения собственных движений, реакцию снижения амплитуды мю-ритма, которая может распространяться как на премоторную, так и париетальную кору [Frenkel-Toledo et al., 2013; Thorpe et al., 2016]. Исходя из этого, появляется возможность оценить соответствующие паттерны реактивности ЭЭГ в связи со свойствами неокортекса в целом, лежащими в основе индивидуальных психологических особенностей человека, в частности, показателей общего интеллектуального развития. Такого рода исследование представляется особенно обоснованным, так как оценка стабильности индивидуальных паттернов реактивности ЭЭГ показала, что в активных задачах наиболее устойчивые показатели демонстрируют модуляции ЭЭГ в диапазоне альфа-ритма именно при наблюдении моторных действий [Лебедева, Каримова, 2020]. При этом следует учитывать модуляции не только его амплитудных, но и частотных параметров. Эта необходимость обусловлена двумя теоретическими предпосылками. Во-первых, считается, что амплитуда мю-ритма в более высоком частотном поддиапазоне отражает синхронную активность более локализованных участков сенсомоторной коры, в то время как на частоте ниже 10 Гц мю-ритм скорее отражает уровень общей моторной корковой активации [Pfurtscheller et al., 2000]. Во-вторых, доминирующая частота альфа-ритма положительно коррелирует с успешностью выполнения ряда когнитивных тестов и может быть ассоциирована с эффективностью нейронного взаимодействия между отдельными участками коры в ходе решения актуальных задач [Grandy et al., 2013].
Настоящая работа является продолжением серии исследований, посвященных анализу паттернов реактивности сенсомоторных ритмов ЭЭГ у детей в задачах, которые принято связывать с процессами активации ЗСМ [Кайда и др., 2019]. Целью работы является изучение возможных взаимосвязей между уровнем развития вербального и невербального компонентов общего интеллекта у детей школьного возраста и особенностями реактивности, частоты и топографии мю-ритма при условии зрительно-слухового и исключительно слухового восприятия инструментальных движений. Мы предполагаем, что у детей, имеющих относительно низкие или высокие показатели интеллекта, будут присутствовать характерные отличия в реактивности фронтального, центрального и париетального корковых компонентов ЗСМ в зависимости от частотной специфичности модуляций мю-ритма ребенка при самостоятельном выполнении соответствующих движений. Принимая во внимание, что к началу подросткового возраста происходит существенное усиление роли фронтальной коры в процессах когнитивного контроля [Steinbeis, Crone, 2016], можно ожидать, что показатели реактивности премоторного компонента ЗСМ у старшей возрастной группы будут более специфично связаны с их интеллектуальным развитием по сравнению с младшей группой.
МЕТОДИКА
Процедура эксперимента. Испытуемый и экспериментатор сидели за отдельными столами (экспериментатор справа, на расстоянии 1.5 м), на каждом из которых размещали монитор и компьютерную мышь (КМ). На экран монитора перед испытуемым выводили видеоизображение участка стола экспериментатора с расположенной на ней КМ (в перспективе от первого лица). Движения КМ испытуемый и экспериментатор осуществляли в среднем темпе 1–2 вращения в секунду с помощью правой руки по часовой стрелке таким образом, чтобы курсор КМ не выходил за границы окна с видеоизображением. Предварительно каждый участник в течение одной минуты отрабатывал данное движение, чтобы оно могло выполняться в режиме достаточной плавности. Последовательность выполнения заданий состояла из серии этапов длительностью 30 с каждый. Перед каждым из них давали соответствующую инструкцию. Полная серия заданий включала в себя этапы пассивной зрительной фиксации на видеоизображении неподвижной КМ, выполнения самостоятельных движений, наблюдения за движениями экспериментатора, имитации движений экспериментатора в режиме реального времени, бодрствующего состояния с закрытыми глазами и слухового восприятия характерных звуков, сопровождающих движения КМ экспериментатора, при условии закрытых глаз испытуемого. Остальные подробности методики описаны ранее [Кайда и др., 2019].
В данной работе анализировали показатели ЭЭГ участников в ситуациях самостоятельной двигательной активности (“Двиг”), наблюдения (“Набл”) и слухового восприятия (“Слух”) движений с КМ относительно предшествующих им исходных условий. Для “Двиг” и “Набл” это была ситуация зрительной фиксации (ЗФ) на видеоизображении неподвижной КМ, для “Слух” – бодрствующего расслабленного состояния с закрытыми глазами (ГЗ). Для сохранения устойчивого внимания испытуемых к предъявляемым стимулам при наблюдении и слуховом восприятии чужих движений им давали задание сосчитать количество циклов вращения КМ в течение каждой экспериментальной серии.
Регистрация ЭЭГ. Запись, обработка и анализ ЭЭГ осуществлены по общепринятой методике с помощью автоматизированного комплекса, состоящего из электроэнцефалографа “Нейрон-Cпектр-3” (фирма “Нейрософт”, Россия) и персонального компьютера. ЭЭГ-потенциалы отводили монополярно от 19 электродов, стандартно расположенных по системе 10–20. Областью интереса в рамках настоящего исследования являлась реактивность ЭЭГ во фронтальных (F3, F4), центральных (C3, C4) и париетальных (P3, P4) отведениях. В качестве референтного электрода использовали объединенные контакты, закрепленные на мочках ушей. Частоты среза фильтров высоких и низких частот составляли соответственно 1.5 и 35 Гц, частота оцифровки ЭЭГ-сигналов – 250 Гц, эпохи анализа – 4 с (с перекрытием 50%). Сигналы обрабатывали с помощью быстрого преобразования Фурье. Для обработки данных использовали компьютерную программу WinEEG (фирма “Мицар”, Россия). Артефакты удаляли с помощью встроенного в приложение метода независимых компонент с дополнительным зрительным контролем качества записи.
Индивидуальная частота мю-ритма. Индивидуальные частотные диапазоны мю-ритма определяли как интервалы шириной 2 Гц (с шагом в 0.5 Гц) в границах частот ЭЭГ от 6 до 13 Гц, имеющие максимальную реакцию десинхронизации амплитуды в отведении C3 при выполнении самостоятельных движений правой рукой с помощью КМ (см. подробное описание [Кайда и др., 2019]).
Индексы реактивности амплитуды мю-ритма. Индексы реактивности (ИР) мю-ритма рассчитывали по формуле [k = ln(B/A)], где k – индекс реактивности мю-ритма, B – амплитуда мю-ритма (стимул), A – амплитуда мю-ритма (фон) [Кайда и др., 2019]. Положительные значения индекса соответствовали реакции синхронизации амплитуды мю-ритма, а отрицательные – реакции десинхронизации.
Уровень интеллекта. Уровень интеллектуального развития испытуемых измеряли с помощью детского варианта теста Векслера (WISC) [Филимоненко, Тимофеев, 2016] непосредственно до процедуры проведения эксперимента. Методика включает в себя 12 субтестов, по результатам выполнения которых рассчитывают уровни развития вербального и невербального интеллекта, а также интегральный балл общего интеллекта. Суммы первичных оценок переводили в стандартные баллы в соответствии с возрастными нормами для каждого участника исследования. В настоящей работе анализировали показатели вербального и невербального интеллекта. Вербальные субтесты состоят из заданий, стимульный материал которых представлен в языковой форме. Их содержанием является установление логико-функциональных связей в стимулах, опосредованных речью. Невербальные тесты интеллекта состоят из заданий, в которых стимульный материал представлен либо в наглядной форме (в виде графических изображений, рисунков, чертежей), либо в предметной форме (кубики, части объектов и пр.). В этих тестах знание языка требуется только для понимания инструкций, которые намеренно делаются простыми и по возможности короткими. В связи с этим можно считать, что вербальные тесты интеллекта отражают в большей степени показатели словесного (понятийного) логического мышления, а с помощью невербальных тестов оценивается преимущественно наглядно-образное и наглядно-действенное мышление [Дружинин, 2007].
Испытуемые и критерии их отбора. В исследовании приняли участие 116 детей в возрасте 8–15 лет. Родителям детей были предоставлены все необходимые сведения о процедуре исследования, и они дали письменное согласие на бесплатное участие ребенка в экспериментах. Настоящее исследование соответствовало этическим принципам Хельсинкской декларации 1964 г. и было одобрено этическим комитетом Крымского федерального университета им. В.И. Вернадского.
Первичным критерием отбора испытуемых для дальнейшего анализа в рамках настоящего исследования служили наличие нормального (или скорректированного до нормального) уровня зрения и слуха, предпочтение правой руки в работе с КМ, а также достаточный уровень когнитивного развития (уровень общего интеллекта не менее 80 баллов). Дополнительным требованием было наличие как минимум 20 с безартефактной записи ЭЭГ в рамках анализируемых этапов эксперимента, а также возможность выделения индивидуального частотного диапазона мю-ритма в заданных границах частот в соответствии с его ключевым свойством – реакцией десинхронизации в ответ на запуск и осуществление произвольных движений. По причине неполного соответствия критериям отбора из исходной выборки были исключены 18 детей. Итоговая выборка составила 98 человек. Она была разделена на две группы: младшую (8–11 лет; n = 56; 31 мальчик, 25 девочек) и старшую (12–15 лет; n = 42; 12 мальчиков, 30 девочек).
Выделение факторов для дисперсионного анализа различий в показателях амплитуды мю-ритма. Для оценки значимости изменений амплитуды индивидуального мю-ритма в каждой из трех экспериментальных ситуаций (самостоятельные движения, наблюдение и слуховое восприятие движений другого человека) применяли многофакторный дисперсионный анализ для связанных переменных (repeated measures ANOVA). Значения амплитуды мю-ритма предварительно подвергали log-трансформации с использованием натурального логарифма. Отдельно рассчитывали эффекты факторов ситуации “Двиг” (Двиг/ЗФ), “Набл” (Набл/ЗФ) и “Слух” (Слух/ГЗ) и их взаимодействие с факторами возраста (“Возраст”: 8–11/12–15 лет), индивидуальной частоты мю-ритма (“Частота”: низкочастотный/высокочастотный мю-ритм), областей коры (“Область”: фронтальная/центральная/париетальная) и полушарий (“Полушарие”: левое/правое).
Выделение факторов для дисперсионного анализа различий в уровне интеллекта. Различия в уровне вербального и невербального интеллекта оценивали в зависимости от влияния и взаимодействия факторов возраста, индивидуальной частоты мю-ритма, а также наличия реакции десинхронизации амплитуды мю-ритма отдельно в ситуациях наблюдения и слухового восприятия. Показатели реактивности мю-ритма в анализируемых парах отведений и полушариях объединяли при условии отсутствия значимых различий в их модуляциях для совокупной выборки детей в соответствующих ситуациях.
Статистическую обработку результатов исследования осуществляли в программном пакете StatSoft Statistica 10.0.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Индивидуальная частота мю-ритма. Усредненные значения индивидуальных границ частотных диапазонов мю-ритма (шириной 2 Гц), соответствующих реакции максимального снижения амплитуды мю-ритма в отведении С3 при выполнении самостоятельных движений правой рукой с КМ, в младшей и старшей группах значимо не различались. В группе детей 8–11 лет они составили соответственно 9.1 ± 1.3 и 11.1 ± 1.3 Гц, а в группе детей 12–15 лет — 9.4 ± 1.2 и 11.4 ± 1.2 Гц (здесь и далее в тексте приводятся средние значения ± стандартное отклонение). Каждая из двух возрастных групп была разделена на две подгруппы. Дети, у которых амплитуда мю-ритма максимально падала в индивидуальном окне частот со срединным значением индивидуального частотного диапазона в пределах от 7 до 10 Гц, были отнесены к подгруппам с низкочастотным мю-ритмом. Те, у кого срединная частота мю-ритма была определена в пределах от 10.5 до 12.0 Гц, включены в подгруппы с высокочастотным мю-ритмом. Количество детей с высокочастотным мю-ритмом в младшей группе составило 27 человек (48.2%), в старшей – 22 человека (52.4%).
Показатели интеллекта. Средние показатели вербального и невербального интеллекта в группах детей, разделенных по возрасту и частоте максимальной супрессии мю-ритма при выполнении самостоятельных движений с КМ, представлены в табл. 1.
Таблица 1.
Оценки вербального и невербального интеллекта детей в зависимости от возрастной группы и частоты максимальной десинхронизации мю-ритма ЭЭГ Table 1. Scores of verbal and nonverbal intelligence in children according to their age groups and mu rhythm peak desynchronization frequency
| Младшая группа, 8–11 лет | Старшая группа, 12–15 лет | |||
|---|---|---|---|---|
| низкочастотный мю-ритм (n = 29) | высокочастотный мю-ритм (n = 27) | низкочастотный мю-ритм (n = 20) | высокочастотный мю-ритм (n = 22) | |
| Вербальный интеллект | 112.2 ± 11.5 | 107.9 ± 13.4 | 109.0 ± 10.9 | 110.4 ± 14.2 |
| Невербальный интеллект | 111.6 ± 15.1 | 109.8 ± 11.2 | 106.9 ± 10.9 | 108.4 ± 13.2 |
Дисперсионный анализ модуляций амплитуды мю-ритма в ситуации “Двиг”. Условие самостоятельного выполнения инструментальных движений приводило к статистически значимому падению амплитуды мю-ритма как без учета других факторов (F1, 94 = 55.90, $\eta _{p}^{2}$ = 0.37, p < 0.001), так и при взаимодействии факторов в следующих сочетаниях: Двиг × Область (F2, 188 = 45.06, $\eta _{p}^{2}$ = 0.32, p < 0.001), Двиг × × Полушарие (F1, 94 = 40.35, $\eta _{p}^{2}$ = 0.32, p < 0.001), Двиг × Область × Полушарие (F2, 188 = 13.32, $\eta _{p}^{2}$ = 0.12, p < 0.001) и Двиг × Полушарие × Частота (F1, 94 = 7.92, $\eta _{p}^{2}$ = 0.08, p = 0.006). Обращают на себя внимание различия в полушарной латерализации эффектов в зависимости от частоты мю-ритма, особенно в париетальных отведениях. Так, у детей с высокочастотной реакцией десинхронизации мю-ритма (рис. 1 (б), (г)) его амплитуда в левом полушарии (отведение Р3) подавляется существенно в большей степени, чем в правом (Р4) в отличие от таковых реакций у детей с низкочастотным мю-ритмом (t = –2.8, p = 0.006).
Рис. 1.
Индексы реактивности (с ошибкой среднего) мю-ритма ЭЭГ в отведениях F3, F4, C3, C4, P3, P4 в ситуациях выполнения самостоятельных инструментальных движений (Двиг), наблюдения за движениями другого человека (Набл) и слуховом восприятии аналогичных движений при условии закрытых глаз (Слух). Фрагмент (а) – младшая группа с низкочастотной десинхронизацией мю-ритма, (б) – с высокочастотной десинхронизацией мю-ритма; (в) — старшая группа с низкочастотной десинхронизацией мю-ритма, (г) – с высокочастотной десинхронизацией мю-ритма.
Fig. 1. Reactivity indices (with mean error) for the EEG mu rhythm in loci F3, F4, C3, C4, P3, P4 under conditions of performing self-paced instrumental movements (Двиг), observing movements performed by another person (Набл) and auditory perception of similar movements, with eyes closed (Слух). Fragments (a) – younger group with low-frequency mu rhythm desynchronization, (б) – with high-frequency mu rhythm desynchronization; (в) – older group with low-frequency mu rhythm desynchronization, (г) – with high-frequency mu rhythm desynchronization.
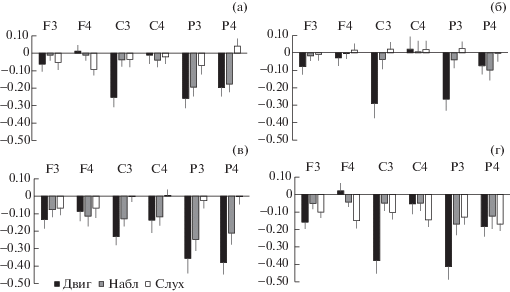
Дисперсионный анализ модуляций амплитуды мю-ритма в ситуации “Набл”. В условиях наблюдения за движениями экспериментатора значимое влияние на амплитуду мю-ритма оказывали лишь фактор ситуации “Набл” (F1, 94 = 24.08, $\eta _{p}^{2}$ = 0.20, p < 0.001) и его взаимодействие с фактором “Область” (F2, 188 = 17.42, $\eta _{p}^{2}$ = 0.16, p < 0.001). Таким образом, ситуация наблюдения за движениями другого человека сопровождалась десинхронизацией мю-ритма, величина которой различалась во фронтальных, центральных и париетальных отведениях независимо от конкретного полушария, индивидуальной частоты мю-ритма и возраста участников.
Дисперсионный анализ модуляций амплитуды мю-ритма в ситуации “Слух”. В условиях слухового восприятия движений значимое влияние на амплитуду мю-ритма оказывал фактор “Слух” (F1, 94 = 8.58, $\eta _{p}^{2}$ = 0.08, p = 0.004), его взаимодействие с фактором “Возраст” (F1, 94 = 4.50, $\eta _{p}^{2}$ = 0.05, p = 0.046), а также взаимодействие с факторами “Возраст” и “Частота” (F1, 94 = 5.94, $\eta _{p}^{2}$ = 0.06, p = 0.017). Отсутствие значимого взаимодействия между факторами “Слух”, “Область” и “Полушарие” позволяет рассматривать реакцию десинхронизации амплитуды мю-ритма при восприятии звуков, сопровождающих знакомые действия как относительно широко распространенный и единообразный процесс активации сенсомоторных и прилегающих к ним париетальных и премоторных областей коры. При этом его величина демонстрирует значимые различия в зависимости от возраста и индивидуальной частоты мю-ритма испытуемых. У испытуемых с низкочастотным мю-ритмом наблюдается некоторая тенденция к падению его амплитуды (рис. 1 (а), (в)), однако не достигающая уровня статистической значимости. В выборке, характеризующейся высокочастотным мю-ритмом, его амплитуда в младшей группе не показывает явной тенденции к изменению (рис. 1 (б)), в то время как в старшей десинхронизация мю-ритма выражена максимально (t = 4.01, p < 0.001) (рис. 1 (г)).
Выделение факторов реактивности мю-ритма для дисперсионного анализа различий в уровне интеллекта. В связи с тем, что в ситуации наблюдения за чужими движениями значимое влияние на десинхронизацию мю-ритма оказывал лишь топографический фактор “Область”, но не “Полушарие”, в последующем анализе показатели ИР мю-ритма усредняли для пар отведений в соответствующих областях коры: фронтальных (F3–F4), центральных (C3–C4) и париетальных (P3–P4). Усредненные показатели ИР для каждой области составили –0.04 ± 0.15, –0.05 ± 0.20 и –0.15 ± 0.26 соответственно. Десинхронизацию мю-ритма во фронтальной, центральной и париетальной областях в ситуации наблюдения за движениями выявили соответственно у 62, 63 и 75 испытуемых. Факт наличия (отрицательные значения ИР) или отсутствия (положительные значения ИР) реакции десинхронизации мю-ритма в данных областях далее рассматривали как дополнительный фактор (“Набл-F” для фронтальных областей, “Набл-С” – для центральных, “Набл-P” – для париетальных), который может быть связан с показателями интеллекта испытуемых.
Условие исключительно слухового восприятия чужих движений вызывало относительно широкую десинхронизацию в изучаемых областях коры, на которую не оказывали значимое влияние факторы “Область” и “Полушарие”. В связи с этим вычисляли общий ИР мю-ритма, который усредняли для индексов реактивности во всех анализируемых отведениях и который был равен 0.04 ± 0.16. Количество испытуемых, показавших десинхронизацию мю-ритма в данной экспериментальной ситуации, составило 58 человек. Аналогично ситуации “Наблюдения”, наличие или отсутствие десинхронизации мю-ритма в ситуации “Слух” также рассматривали в качестве фактора (“Слух-FCP”), который может быть связан с показателями интеллекта.
Различия в показателях вербального и невербального интеллекта (отдельно для каждого) оценивали с помощью дисперсионного анализа при взаимодействии таких факторов, как “Набл-F”, “Набл-C”, “Набл-P”, “Слух-FCP” (отдельно для каждого), “Возраст” и “Частота”. Различия в уровне интеллекта между отдельными подгруппами испытуемых, у которых проявлялась или отсутствовала реакция десинхронизации амплитуды мю-ритма, анализировали с помощью метода априорных контрастов (F-статистика).
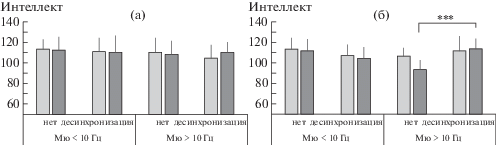
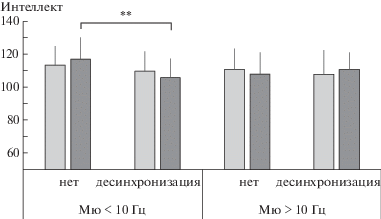
Анализ различий в уровне интеллекта в ситуации “Набл”. Фактор реактивности мю-ритма во фронтальных отведениях ЭЭГ не оказывал значимого влияния на уровень вербального интеллекта, в том числе и при взаимодействии с другими выделенными факторами. При этом показатели невербального интеллекта зависели от взаимодействия факторов Набл-F × Возраст (F1, 90 = 8.56, $\eta _{p}^{2}$ = = 0.09, p = 0.004) и Набл-F × Возраст × Частота (F1, 90 = 4.49, $\eta _{p}^{2}$ = 0.05, p = 0.037). Показатели интеллекта соответствующих подгрупп испытуемых представлены на рис. 2. Анализ контрастов между подгруппами с разным характером реактивности высокочастотного мю-ритма показал наиболее выраженное различие в старшей группе школьников (рис. 2 (б)). Те из них, у кого при наблюдении за чужими движениями амплитуда мю-ритма во фронтальных отведениях не уменьшалась, характеризовались существенно (t = 3.44, p < < 0.001) более низким уровнем развития невербального интеллекта (в среднем 93.7 ± ± 9.0 баллов), чем те, у кого мю-ритм десинхронизировался (113.9 ± 9.9).
Рис. 2.
Средние оценки вербального (светлые столбцы) и невербального (темные столбцы) интеллекта у детей 8–11 (а) и 12–15 (б) лет в зависимости от индивидуальной частоты мю-ритма и наличия реакции его десинхронизации во фронтальных отведениях ЭЭГ в ситуации наблюдения за инструментальными движениями. *** – различия значимы при p < 0.001.
Fig. 2. Mean scores for verbal (lighter columns) and nonverbal (darker columns) intelligence in children aged 8–11 (a) and 12–15 (б) years, grouped by individual mu rhythm frequency and presence of its desynchronization reaction in EEG frontal loci, when observing instrumental movements. *** – significant differences with p < 0.001.
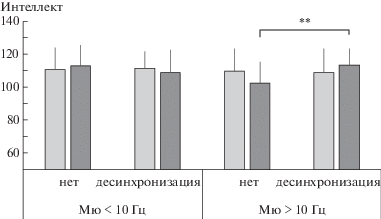
Анализ зависимости показателей интеллекта от характера реактивности мю-ритма в центральных отведениях ЭЭГ в сочетании с другими выделенными факторами продемонстрировал в целом сходные результаты. Было обнаружено значимое взаимодействие факторов Набл-C × Частота (F1, 90 = 9.74, $\eta _{p}^{2}$ = = 0.1, p = 0.002). Однако эффект взаимодействия факторов Набл-C × Возраст × Частота на показатели невербального интеллекта не достигал уровня статистической значимости (F1, 90 = 3.33, $\eta _{p}^{2}$ = 0.04, p = 0.071). Так как фактор “Возраст” не оказывал значимого влияния на показатели интеллекта, анализ контрастов по факту наличия десинхронизации мю-ритма проводили для детей обеих возрастных групп в совокупности. Усредненные оценки уровня развития каждого типа интеллекта испытуемых в соответствующих подгруппах представлены на рис. 3. Значимые различия в оценках невербального интеллекта были выявлены также лишь у детей с преимущественно высокочастотной десинхронизацией мю-ритма при совершении самостоятельных движений с КМ. У детей с отсутствием реакции десинхронизации высокочастотного мю-ритма в центральных отведениях показатели невербального интеллекта были значимо (t = 2.98, p = 0.004) ниже (102.5 ± ± 12.7 балла), чем у тех, у кого десинхронизация имела место (113.0 ± 9.9 балла).
Рис. 3.
Средние оценки вербального (светлые столбцы) и невербального (темные столбцы) интеллекта у детей 8–15 лет в зависимости от индивидуальной частоты мю-ритма и наличия реакции его десинхронизации в центральных отведениях ЭЭГ в ситуации наблюдения за инструментальными движениями. ** – различия значимы при p < 0.01.
Fig. 3. Mean scores for verbal (lighter columns) and nonverbal (darker columns) intelligence in children aged 8–15 years, grouped by individual mu rhythm frequency and presence of its desynchronization reaction in EEG central loci, when observing instrumental movements. ** – significant differences with p < 0.01.
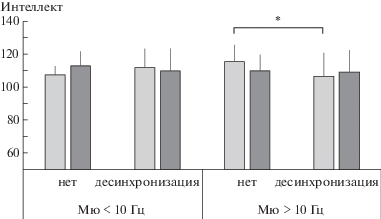
Фактор реактивности мю-ритма в париетальных отведениях также достоверно взаимодействовал с фактором индивидуальной частоты мю-ритма, но в этом случае эффект данного взаимодействия оказался значимым для уровня вербального интеллекта (Набл-P × × Частота: F1, 90 = 4.59, $\eta _{p}^{2}$ = 0.03, p = 0.049). Соответствующие выделенным подгруппам оценки интеллекта показаны на рис. 4. Так как фактор возраста не оказывал значимого влияния на показатели интеллекта, при расчете контрастов выборка по возрасту не делилась. В отличие от закономерностей, описанных для лобных и центральных отведений ЭЭГ, в теменных отведениях значимые различия были обнаружены для оценок детей по шкале вербального интеллекта. Эти различия так же выявлены лишь для детей с высокочастотным мю-ритмом. Дети, у которых наблюдение за движениями с КМ не сопровождалось падением амплитуды мю-ритма в париетальных отведениях, имели значимо (t = –2.61, p = 0.011) более высокие оценки по шкале вербального интеллекта, в среднем равные 115.3 ± 10.3 балла, против 105.9 ± 14.2 у тех, у кого мю-ритм десинхронизировался.
Рис. 4.
Средние оценки вербального (светлые столбцы) и невербального (темные столбцы) интеллекта у детей 8–15 лет в зависимости от индивидуальной частоты мю-ритма и наличия реакции его десинхронизации в париетальных отведениях ЭЭГ в ситуации наблюдения за инструментальными движениями. * – различия значимы при p < 0.05.
Fig. 4. Mean scores for verbal (lighter columns) and nonverbal (darker columns) intelligence in children aged 8–15 years, grouped by individual mu rhythm frequency and presence of its desynchronization reaction in EEG parietal loci, when observing instrumental movements. * – significant differences with p < 0.05.
Анализ различий в уровне интеллекта в ситуации “Слух”. Фактор реактивности амплитуды мю-ритма всей совокупности фронтальных, центральных и париетальных отведений в ситуации слухового восприятия инструментальных движений оказывал значимое влияние на уровень невербального интеллекта при взаимодействии с фактором индивидуальной частоты мю-ритма (Слух-FCP×Частота: F1, 90 = 5.52, $\eta _{p}^{2}$ = 0.06, p = 0.021). Соответствующие подгруппам испытуемых показатели интеллекта представлены на рис. 5. Расчет индивидуальных контрастов между подгруппами детей с разным характером реактивности мю-ритма в ситуации слухового восприятия движений показал значимые различия уровня интеллекта в выборке детей, для которых условие самостоятельных манипуляций с КМ сопровождалось преимущественным падением амплитуды низкочастотного мю-ритма. Значимо (t = –2.67, p = 0.009) более высокие оценки невербального интеллекта (116.7 ± 13.3 балла) были в подгруппе участников, у которых амплитуда мю-ритма, усредненная для совокупности всех рассматриваемых отведений ЭЭГ, не показывала реакции десинхронизации. В подгруппе детей, у которых низкочастотный мю-ритм десинхронизировался, уровень невербального интеллекта составил в среднем 105.3 ± 12.1 балла.
Рис. 5.
Средние оценки вербального (светлые столбцы) и невербального (темные столбцы) интеллекта у детей 8–15 лет в зависимости от индивидуальной частоты мю-ритма и наличия реакции его десинхронизации для всей совокупности фронтальных, центральных и париетальных отведений ЭЭГ в ситуации слухового восприятия инструментальных движений другого человека. ** – различия значимы при p < 0.01.
Fig. 5. Mean scores for verbal (lighter columns) and nonverbal (darker columns) intelligence in children aged 8–15 years, grouped by individual mu rhythm frequency and presence of its desynchronization reaction in EEG frontal, central and parietal loci combined, when hearing sounds characteristic for instrumental movements. ** – significant differences with p < 0.01.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Выполненное исследование исходило из теоретических предпосылок, согласно которым характер и динамика реакций ЗСМ при наблюдении действий окружающих, хотя и имеют определенную врожденную предопределенность, в значительной степени все же являются результатом процессов индивидуального ассоциативного научения [Cook et al., 2014]. Другими словами, чтобы возникла “зеркальная” реакция, необходимо, чтобы наблюдаемое действие уже находилось в индивидуальном репертуаре действий наблюдателя. При этом величина активации релевантных наблюдаемому действию участков сенсомоторной коры (т.н. “сенсомоторный резонанс”) будет тем более выраженной, чем в большей степени освоено это действие самим наблюдателем. Поэтому при изучении особенностей функционирования зеркальной системы мозга всегда следует учитывать индивидуальную специфичность наблюдаемого действия, которое используется в качестве экспериментального стимула, способного вызвать “зеркальную” реакцию.
В рамках настоящего исследования анализировали реактивность осцилляций мю-ритма при относительно продолжительном (в течение 30 с) выполнении инструментальных движений, а также зрительно-слуховом (условие наблюдения, при котором основным каналом информации был зрительный, но испытуемые также могли слышать характерные звуки движения КМ) и исключительно слуховом восприятии аналогичных движений, выполняемых другим человеком. Благодаря этому достигалось относительно устойчивое функциональное состояние ЦНС, необходимое для выполнения соответствующей экспериментальной задачи. Вследствие эффектов нейрональной адаптации к повторяющимся стимулам усредненные показатели мю-ритма в каждой ситуации могли быть менее выражены, чем в случае анализа коротких интервалов ЭЭГ непосредственно до и после предъявления стимулов [Александров, Тугин, 2010]. Преимуществом данного подхода является уменьшение эффектов колебания внимания, сопровождающих ориентировочную реакцию после начала действия нового стимула. Тем самым при оценке модуляций мю-ритма можно было снизить вклад реактивности классического альфа-ритма (прежде всего теменно-затылочной локализации), ассоциированной с различными аспектами обработки сигналов.
При использовании процедуры выделения индивидуального мю-ритма было необходимо учитывать тот факт, что его пиковая частота в детском возрасте может быть несколько ниже, чем у взрослых. Как известно, частота пика данного ритма быстро растет от 3 до 8 Гц у детей в течение первого года жизни, после чего ее рост постепенно замедляется и достигает близкой к 10 Гц величины к периоду ранней взрослости [Berchicci et al., 2011]. Более того, некоторые авторы полагают, что мю-ритму взрослого человека соответствуют осцилляции ЭЭГ конкретно в области частот 10–13 Гц [Thorpe et al., 2016]. Также распространена точка зрения, что низкочастотная полоса мю-ритма чувствительна к общей моторной активации, в то время как высокочастотная отражает более узко локализованные модуляции активности сенсомоторной коры [Pfurtscheller et al., 2000]. Исходя из этого мы выделяли относительно узкий частотный диапазон мю-ритма, наиболее чувствительный к тому типу произвольного моторного контроля, который использовался в нашем эксперименте, – длительной манипуляции компьютерной мышью таким образом, чтобы курсор на экране монитора циклически двигался по кругу в рамках заданной области. Индивидуально определив показатели мю-ритма для каждого участника, можно в дальнейшем с бо́льшим основанием утверждать, что регистрируемые при наблюдении и слуховом восприятии модуляции ЭЭГ могут быть интерпретированы как имеющие “зеркальную” природу.
Нами были выделены подгруппы испытуемых, имеющих преимущественно низко- (≤10 Гц) или высокочастотный (>10 Гц) диапазоны реактивности мю-ритма при выполнении самостоятельных манипуляций с КМ. Их соотношение в рамках каждой возрастной группы было примерно равным. В дальнейшем мы рассматривали детей с низкочастотным диапазоном мю-ритма как тех, для кого манипуляции с КМ являются сенсомоторно менее специфическими (работа с КМ у них приводит к общей моторной активации), а для детей с высокочастотным диапазоном – более специфическими (работа с КМ сопровождается более узко локализованными реакциями активации сенсомоторной коры) [Pfurtscheller et al., 2000]. Мы полагаем, что более выраженная локальность процесса могла быть обусловлена, например, бо́льшим опытом использования компьютерной мыши в повседневной жизни или же большей степенью сосредоточенности на выполнении экспериментальной задачи и соответственно более тонким произвольным контролем собственных движений.
Условие выполнения самостоятельных движений с КМ вызывало наиболее существенную супрессию мю-ритма в левом полушарии: менее выраженную во фронтальной области и более – в центральной и париетальной. В правом полушарии выраженная десинхронизация имела место только в париетальном отведении. При этом не было значимых различий в характере реактивности мю-ритма между младшей и старшей возрастными группами. Реакция десинхронизации мю-ритма в париетальных отведениях при осуществлении самостоятельных движений, сопоставимая по силе с найденной для левого центрального отведения, может объясняться их относительной продолжительностью. В то время как “быстрые” движения реализуются в соответствии с предварительно сформированным моторным планом и не требуют периферических сигналов обратной связи, “медленные” (соответствующие условиям нашего эксперимента) осуществляются с учетом поступающей от соматосенсорных рецепторов информации [Stancak, Pfurtscheller, 1996]. При этом в подгруппе с высокочастотным мю-ритмом его амплитуда значимо больше снижалась в париетальных отведениях левого полушария (по сравнению с правым), в то время как у детей с низкочастотным мю-ритмом она уменьшалась в париетальных отведениях обоих полушарий примерно в равной степени.
При анализе “зеркальных” реакций в ходе зрительно-слухового наблюдения и исключительно слухового восприятия движений другого человека мы рассматривали возможность выделения отдельных компонентов “сенсомоторного резонанса”: низкоуровневого, связанного с активацией первичной сенсомоторной коры (отведения С3–С4) и высокоуровневого, связанного с активацией фронтальной (F3–F4) и париетальной (P3–P4) коры. Активация премоторной коры, с нашей точки зрения, могла быть результатом актуализации релевантных наблюдаемому движению схем моторного контроля, которую можно связать с имплицитным пониманием целей действия, а париетальной – соответствующих зрительно-пространственных паттернов восприятия. Дополнительно оценивали возможность наличия межполушарных различий в регистрируемых реакциях.
Анализ паттернов реактивности индивидуального мю-ритма детей при наблюдении инструментальных движений другого человека позволил обнаружить его “зеркальную” десинхронизацию без существенной полушарной латерализации, но значимо дифференцированную по величине для фронтальных (минимальная), центральных и париетальных (максимальная) отведений. Такие факторы, как возраст и индивидуальный частотный диапазон мю-ритма, не оказывали значимого влияния на данные показатели. Однако при оценке их взаимодействия с факторами наличия/отсутствия десинхронизации непосредственно в каждой из трех областей коры были выявлены специфические различия в показателях вербального и невербального интеллекта участников исследования. Что интересно, оценка индивидуальных контрастов выявила значимые различия в уровне интеллектуального развития исключительно в подгруппе испытуемых с высокочастотным мю-ритмом. Модуляции амплитуды мю-ритма у детей с низкочастотным мю-ритмом не показали связи с их интеллектуальными показателями.
При анализе показателей, зарегистрированных в ситуации наблюдения за движениями, наиболее контрастное различие в уровне интеллекта было обнаружено в старшей подгруппе с высокочастотным мю-ритмом между детьми, характеризовавшимися разным направлением модуляций его амплитуды. Те из них, у кого амплитуда мю-ритма в указанной ситуации не снижалась, характеризовались существенно более низкими показателями невербального интеллекта. Важно указать, что в младшей группе данная закономерность не прослеживалась. Известно [Steinbeis, Crone, 2016], что в подростковом возрасте (а дети старшей группы находились в пубертатном или препубертатном периоде) роль фронтальной коры в процессах саморегуляции и когнитивного контроля существенно возрастает. Исходя из этого, можно высказать предположение, что отсутствие реакции “зеркальной” активации релевантных схем моторного контроля, связанных с наблюдаемыми действиями другого человека, может оказаться для детей одной из возможных причин сложностей перехода на более высокий уровень когнитивной организации, соответствующий подростковому возрасту.
Отсутствие низкоуровневой “зеркальной” десинхронизации в областях первичной сенсомоторной коры (отведения С3–С4) аналогичным образом указывало на значимо более низкий уровень развития невербального интеллекта. Однако в данном случае эта закономерность характеризовала испытуемых с высокочастотным мю-ритмом уже независимо от их принадлежности к конкретной возрастной группе. Можно высказать предположение, что отсутствие низкоуровневой “зеркальной” активации при наблюдении знакомых инструментальных движений также может оказаться одним из факторов замедления процессов естественного развития невербального интеллекта в период взросления, как минимум, начиная с раннего школьного возраста.
Закономерность другого рода была обнаружена для испытуемых, не продемонстрировавших реакцию десинхронизации высокочастотного мю-ритма в париетальных областях коры, которые связывают преимущественно с процессами визуомоторной интеграции. Уровень их вербального интеллекта оказался несколько выше, чем у тех, у кого амплитуда мю-ритма в париетальных отведениях (P3–P4) падала при наблюдении за движениями. Данные различия, хоть и достигали уровня статистической значимости, были выражены менее контрастно по сравнению с описанными выше. Мы полагаем, что, возможно, дети с более высокоразвитой второй сигнальной системой могут более эффективно распределять когнитивные ресурсы при выполнении поставленных задач, произвольно тормозя индуцированные наблюдением процессы имплицитной активации соответствующих зон зрительно-пространственного восприятия. По-видимому, данная логика применима в первую очередь к конкретным условиям нашего эксперимента, так как задача наблюдения циклически повторяющихся инструментальных движений на протяжении полуминуты не предъявляет повышенных требований к процессам восприятия.
Ситуация слухового восприятия инструментальных движений при условии закрытых глаз обнаружила “зеркальные” эффекты, которые значимо не дифференцировались ни по полушариям, ни по отдельным областям коры, активность в которых анализировалась. В связи с этим фактор наличия или отсутствия реакции десинхронизации мю-ритма рассматривался нами далее для всех шести отведений ЭЭГ в совокупности. Поэтому в определенной степени предсказуемо, что данный фактор обнаружил связь с показателями интеллекта лишь у тех испытуемых, для которых выполнение соответствующих инструментальных движений сопровождалось десинхронизацией амплитуды преимущественно низкочастотного мю-ритма, отражающего неспецифическую сенсомоторную активацию. Дети, не продемонстрировавшие такого рода “зеркальной” активации, обладали значимо более высокими показателями невербального интеллекта по сравнению с теми, у кого наблюдали значимое снижение амплитуды мю-ритма.
При интерпретации данного наблюдения следует учитывать особенности экспериментальной ситуации, заключающиеся в том, что испытуемых заранее не предупреждали о том, что им предстоит услышать знакомые звуки, которые у них уже устойчиво ассоциировались с ранее выполнявшимися движениями. Их задачей было сохранять расслабленное состояние при условии закрытых глаз и не предпринимать никаких действий без инструкции. Поэтому нельзя исключить, что реакция широкой десинхронизации низкочастотного мю-ритма может отражать как неспецифическую “зеркальную” реакцию, так и аттенциональные эффекты, сопровождающие мыслительные рефлексивные процессы в сознании детей. Отсутствие такого рода реакций у испытуемых, имеющих в среднем более высокий уровень невербального интеллекта, может условно говорить в пользу концепции нервной эффективности. Другими словами, более высокий уровень когнитивного развития детей школьного возраста снижает необходимость в выраженной “зеркальной” реакции сенсомоторных областей коры в ситуации относительно простого социального восприятия, связанного со слуховым узнаванием действий других людей. Однако данная закономерность ограничена условием того, что воспринимаемые инструментальные движения имеют для испытуемого относительно низкую сенсомоторную специфичность. Мы полагаем, что данный тип специфичности может быть обусловлен либо менее развитым соответствующим моторным навыком, либо меньшей степенью концентрации внимания ребенка на выполнении задания.
ЗАКЛЮЧЕНИЕ
Результаты настоящего исследования со всей определенностью указывают на то, что при изучении нейронных “зеркальных” механизмов в ситуации восприятия чужих действий, особенно при условии использования модуляций мю-ритма ЭЭГ в качестве маркера активации зеркальной системы мозга, необходимо предварительно оценивать специфичность реакций ЦНС индивида при самостоятельном осуществлении подобных действий. Дополнительно необходимо принимать во внимание, какой перцептивный канал является в акте восприятия основным. Данная рекомендация представляется особенно актуальной в случае экспериментальных исследований с участием детей. От этого могут в значительной степени различаться как сами “зеркальные” реакции, так и особенности их взаимосвязи с индивидуальными особенностями когнитивного развития.
В случае условно низкой для субъекта сенсомоторной специфичности конкретных инструментальных действий (их самостоятельное выполнение сопровождается десинхронизацией преимущественно низкочастотного мю-ритма) “зеркальные” эффекты в широких сенсомоторных проекциях, индуцированные исключительно слуховым восприятием сопутствующих этим действиям звуков, оказываются менее выраженными у детей школьного возраста, имеющих более высокий уровень развития невербального интеллекта.
В случае относительно высокой сенсомоторной специфичности процесса осуществления инструментальных движений (их самостоятельное выполнение сопровождается десинхронизацией преимущественно высокочастотного мю-ритма), при зрительно-слуховом восприятия движений другого человека “зеркальные” эффекты оказываются дифференцированными для премоторных, париетальных и сенсомоторных областей коры. И в каждом случае они специфично взаимодействуют с показателями интеллектуального развития. При этом не выявлено значимой полушарной латерализации в характере выявленных закономерностей.
Отсутствие “зеркальной” активации премоторных участков коры оказалось ассоциировано с относительно низким уровнем развития невербального интеллекта. Причем данная закономерность была значимо выражена лишь в группе детей старшего школьного возраста. Относительно низкие показатели невербального интеллекта характеризовали всех детей школьного возраста, у которых не регистрировалась “зеркальная” активация в области сенсомоторной коры. В свою очередь отсутствие “зеркальной” активации париетальных участков коры было более характерно для детей, имеющих несколько более высокий уровень развития вербального интеллекта.
Таким образом, с определенной степенью осторожности можно предполагать, что недостаток функциональной активности зеркальной системы мозга у детей школьного возраста может предрасполагать как к более низким, так и более высоким показателям интеллекта. Отсутствие имплицитных “зеркальных” реакций при наблюдении ими хорошо знакомых инструментальных действий может предсказывать более низкие показатели невербального интеллекта. В то же время отсутствие таких реакций у детей при слуховом узнавании менее свойственных им самим действий может указывать, наоборот, на более высокий уровень развития невербального интеллекта. При этом для вербального интеллекта обнаружена тенденция к более высоким показателям, если “зеркальные” реакции на наблюдение знакомых действий снижены. Другими словами, высокая активность ЗСМ не является однозначно благоприятной для развития общего интеллекта ребенка. Многие аспекты учебной или другой деятельности в бо́льшей степени требуют от детей и подростков самостоятельности и оригинальности мышления, чем хорошо развитой способности к подражанию.
Исследования выполнены с использованием оборудования ЦКП научным оборудованием “Экспериментальная физиология и биофизика” КФУ им. В.И. Вернадского при финансовой поддержке РФФИ в рамках научного проекта № 18-015-00074 А.
Список литературы
Александров А.А., Тугин С.М. Изменения µ-ритма при различных формах двигательной активности и наблюдении движений. Российский физиологический журнал им. И.М. Сеченова. 2010. 96 (11): 1088–1096.
Дружинин В.Н. Психология общих способностей. 3-е изд. СПб.: Питер: Техническая книга, 2007. 368 с.
Кайда А.И., Махин С.А., Эйсмонт Е.В., Павленко В.Б. Возрастная динамика и топография реактивности индивидуального мю-ритма ЭЭГ у детей 4–14 лет. Вестник Томского государственного университета. Биология. 2019. 45: 106–127.
Курганский А.В. Функциональная организация мозга человека в состоянии покоя. Журн. высш. нерв. деят. им. И.П. Павлова. 2018. 68 (5): 567–580.
Лебедева Н.Н., Зуфман А.И., Мальцев В.Ю. Система зеркальных нейронов мозга: ключ к обучению, формированию личности и пониманию чужого сознания. Успехи физиологических наук. 2017. 48 (4): 16–28.
Лебедева Н.Н., Каримова Е.Д. Устойчивость паттернов ЭЭГ человека в различных задачах: проблема аутентификации личности. Журн. высш. нерв. деят. им. И.П. Павлова. 2020. 70 (1): 40–49.
Лебедева Н.Н., Каримова Е.Д., Карпычев В.В., Мальцев В.Ю. Зеркальная система мозга при наблюдении, выполнении и представлении моторных задач – нейрофизиологическое отражение восприятия чужого сознания. Журн. высш. нерв. деят. им. И.П. Павлова. 2018. 68 (2): 204–215.
Риццолатти Дж., Синигалья К. Зеркала в мозге: О механизмах совместного действия и сопереживания. М: Языки славянских культур, 2012. 208 с.
Филимоненко Ю.И. Тест Д. Векслера. Диагностика структуры интеллекта (детский вариант): методическое руководство. СПб.: ИМАТОН, 2016.
Alcala-Lopez D., Vogeley K., Binkofski F., Bzdok D. Building blocks of social cognition: Mirror, mentalize, share? Cortex. 2019. 118: 4–18.
Berchicci M., Zhang T., Romero L., Peters A., Annett R., Teuscher U., Bertollo M., Okada Y., Stephen J., Comani S. Development of mu rhythm in infants and preschool children. Developmental Neuroscience. 2011. 33: 130–143.
Bidet-Ildei C., Beauprez S.-A., Badets A. A review of literature on the link between action observation and action language: advancing a shared semantic theory. New ideas in psychology. 2020. 58: 100777.
Binder E., Dovern A., Hesse M.D., Ebke M., Karbe H., Saliger J., Fink G.R., Weiss P.H. Lesion evidence for a human mirror neuron system. Cortex. 2017. 90: 125–137.
Bushov Yu.V., Svetlik M.V., Esipenko E.A., Kartashov S.I., Orlov V.A., Ushakov V.L. The activity of human mirror neurons during observation and time perception. Sovremennye Tehnologii v Medicine. 2019. 11 (1): 69–74.
Chen S.A., Desmond J.E. Cerebrocerebellar networks during articulatory rehearsal and verbal working memory tasks. Neuroimage. 2005. 24: 332–338.
Cook R., Bird G., Catmur C., Press C., Heyes C. Mirror neurons: from origin to function. Behav Brain Sci. 2014. 37 (2): 177–192.
di Pellegrino G., Fadiga L., Fogassi L., Gallese V., Rizzolatti G. Understanding motor events: a neurophysiological study. Exp. Brain Res. 1992. 91: 176–180.
Fox N.A., Bakermans-Kranenburg M.J., Yoo K.H., Bowman L.C., Cannon E.N., Vanderwert R.E., Ferrari P.F. Assessing human mirror activity with EEG mu rhythm: A meta-analysis. Psychol Bull. 2016.142 (3): 291–313.
Frenkel-Toledo S., Bentin S., Perry A., Liebermann D.G., Soroker N. Dynamics of the EEG power in the frequency and spatial domains during observation and execution of manual movements. Brain Res. 2013. 1509: 43–57.
Gallese V., Sinigaglia C. What is so special about embodied simulation? Trends in Cognitive Sciences. 2011. 15: 512–519.
Glenberg A.M., Gallese V. Action-based language: a theory of language acquisition, comprehension, and production. Cortex. 2012. 48: 905–922.
Grandy T.H., Werkle-Bergner M., Chicherio C., Lovden M., Schmeidek F., Lindenberger U. Individual alpha peak frequency is related to latent factors of general cognitive abilities. Neuroimage. 2013. 79: 10–18.
Jospe K., Floel A., Lavidor M. The interactive effect of empathy and motor cortex stimulation on hand gesture comprehension. Neuropsychologia. 2020. 141: 107412.
Jung R.E., Haier R.J. The parieto-frontal integration theory (P-FIT) of intelligence: converging neuroimaging evidence. Behav Brain Sci. 2007. 30 (2): 135–154.
Krautheim J.T., Dannlowski U., Steines M., Neziroglu G., Acosta H., Sommer J., Straube B., Kircher T. Intergroup empathy: Enhanced neural resonance for ingroup facial emotion in a shared neural production-perception network. Neuroimage. 2019. 194: 182–190.
Langeslag S.J., Schmidt M., Ghassabian A., Jaddoe V.W., Hofman A., van der Lugt A., Verhulst F.C., Tiemeier H., White T.J. Functional connectivity between parietal and frontal brain regions and intelligence in young children: the Generation R study. Hum. Brain Mapp. 2013. 34: 3299–3307.
Macher K., Bohringer A., Villringer A., Pleger B. Cerebellar-parietal connections underpin phonological storage. J. Neurosci. 2014. 34: 5029–5037.
Molenberghs P., Cunnington R., Mattingley J.B. Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies. Neuroscience and Biobehavioral Reviews. 2012. 36: 341–349.
Mottonen R., Farmer H., Watkins K.E. Neural basis of understanding communicative actions: Changes associated with knowing the actor’s intention and the meanings of the actions. Neuropsychologia. 2016. 81: 230–237.
Pfurtscheller G., Neuper C., Krausz G. Functional dissociation of lower and upper mu rhythms in relation to voluntary limb movement. Clinical neurophysiology. 2000. 111 (10): 1873–1879.
Pineda J. Sensorimotor cortex as a crucial component of an ‘extended’ mirror neuron system: Does it solve the development, correspondence, and control problem in mirroring? Behavioral and Brain Functions. 2008. 4 (1): 47.
Power J.D., Fair D.A., Schlaggar B.L., Petersen S.E. The development of human functional brain networks. Neuron. 2010. 67: 735–748.
Stancák A. Jr., Pfurtscheller G. Mu rhythm changes in brisk and slow self-paced finger movements. Neuroreport. 1996. 7: 1161–1164.
Steinbeis N., Crone E.A. The link between cognitive control and decision-making across child and adolescent development. Current Opinion in Behavioral Sciences. 2016. 10: 28–32.
Thorpe S.G., Cannon E.N., Fox N.A. Spectral and source structural development of mu and alpha rhythms from infancy through adulthood. Clinical Neurophysiology. 2016. 127 (1): 254–269.
Tompson S.H., Kahn A.E., Falk E.B., Vettel J.M., Bassett D.S. Functional brain network architecture supporting the learning of social networks in humans. Neuroimage. 2020. 210: 116498.
Wadsworth H.M., Maximo J.O., Lemelma A.R., Clayton K., Sivaraman S., Deshpande H.D., Ver Hoef L., Kana R.K. The action imitation network and motor imitation in children and adolescents with autism. Neuroscience. 2017. 343: 147–156.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова