

Журнал высшей нервной деятельности им. И.П. Павлова, 2020, T. 70, № 5, стр. 609-615
Потенциалы, связанные с событиями при выполнении теста Go/NoGo в подгруппах испытуемых с разным уровнем выраженности вербальной креативности
Ю. А. Бойцова 1, *, М. Г. Старченко 1
1 Федеральное государственное бюджетное учреждение науки Институт мозга человека им. Н.П. Бехтеревой Российской академии наук (ИМЧ РАН)
Санкт-Петербург, Россия
* E-mail: Boytsova.ihb@gmail.com
Поступила в редакцию 17.02.2020
После доработки 13.04.2020
Принята к публикации 01.06.2020
Аннотация
В литературе неоднократно обсуждалась связь между творческим мышлением и вниманием, но вопрос об особенностях внимания у индивидуумов с разным уровнем выраженности творческих способностей остается малоизученным. В настоящей работе проведено сопоставление потенциалов, связанных с событиями (ПСС) у подгрупп испытуемых с разным уровнем выраженности вербальной креативности, оцениваемой по тесту отдаленных ассоциаций Медника РАТ, при выполнении двустимульной модификации теста Go/NoGo, который является традиционным инструментом для исследования механизмов внимания. В результате работы показано, что подгруппы испытуемых с высоким и средним уровнем вербальной креативности отличаются от подгруппы испытуемых с низким уровнем таковой большей амплитудой позднего позитивного компонента Р300. Направленность выявленных различий между подгруппами одинакова в пробах Go и NoGo, но достоверные различия выявлены только в пробе NoGo. Также подгруппа испытуемых с высоким уровнем вербальной креативности отличается от подгруппы с низким уровнем меньшим временем реакции в пробе Go. Полученные данные позволяют предположить, что подгруппы испытуемых с разным уровнем выраженности уровня вербальной креативности различаются механизмами внимания, которые связаны с процессами подавления ответа и, возможно, с процессами вовлечения в действие.
ВВЕДЕНИЕ
Исследования нейрофизиологических механизмов творческой деятельности имеют богатую историю, в этой области накоплен огромный опыт анализа нейровизуализационных данных, анализа параметров спонтанной ЭЭГ, вызванной синхронизации/десинхронизации и вызванных потенциалов [Dietrich, Kanso, 2010]. Однако в исследованиях нейрофизиологии творчества один вопрос остается недостаточно исследованным – это вопрос взаимосвязи творческого мышления с особенностями процессов внимания.
Предположения о том, что в процессе творческой деятельности (креативности) немаловажную роль играют внимание и способ обработки информации, впервые были сформулированы Айзенком и Мартиндейлом [Eysenck, 1993; Martindale, 1995]. Предполагается, что внимание креативных испытуемых более расфокусировано по сравнению с низкокреативными. Широкий фокус внимания у креативных испытуемых позволяет учитывать большее количество информации, релевантной и нерелевантной. Это, в свою очередь, облегчает возникновение нестандартных идей, построение отдаленных ассоциаций, что рассматривается как один из механизмов творческого мышления [Mednick, Mednick, 1967]. Существует и противоположная точка зрения, что творческие люди просто легче регулируют фокус внимания в зависимости от требований задачи [Ansburg, Hill, 2003].
Анализ вызванных потенциалов широко используется для исследования различных этапов и механизмов внимания в тестах на внимание [Luck, 2014]. Разные компоненты вызванных потенциалов также рассматриваются при выполнении творческих и нетворческих заданий, при исследовании инсайтной стратегии решения задач [Dietrich, Kanso, 2010]. Но эти исследования не дают возможности непосредственно сопоставить механизмы внимания и креативности.
Целью настоящей работы было исследование особенностей внимания у испытуемых с разным уровнем вербальной креативности.
МЕТОДИКА
В исследовании приняли участие 38 человек, все праворукие (по тесту [Oldfield, 1971]. В соответствии с этическими стандартами Хельсинкской декларации 1964 г. все испытуемые подписали добровольное согласие на участие в исследовании. Исследование было одобрено решением Этической Комиссии Института мозга человека им. Н.П. Бехтеревой РАН.
Разделение данной группы испытуемых на подгруппы с разным уровнем выраженности творческих способностей проводили на основе результатов выполнения русскоязычного адаптированного варианта теста для диагностики вербальной креативности С. Медника [адаптация А.Н. Воронин, 1994, цит. по Истратова, 2006]. В результате в подгруппу испытуемых с высоким уровнем вербальной креативности (ВК) вошли 11 испытуемых (средний возраст 25 лет, 1 мужчина). Подгруппу с низким уровнем вербальной креативности (НК) составили 11 испытуемых (средний возраст 25 лет, 3 мужчины). В подгруппу со средним уровнем вербальной креативности (СК) вошли 16 испытуемых (средний возраст 24 года, 4 мужчины).
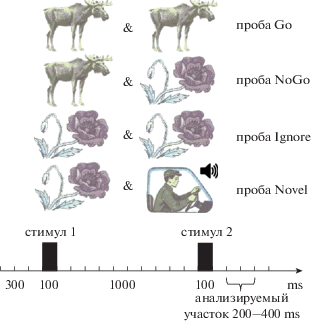
Используемый в работе двустимульный тест состоял из 400 проб в парадигме Go/NoGo (рис. 1). В качестве стимулов используются изображения животных, растений, человека. Пробами (тестовыми условиями) являются пары зрительных стимулов: “животное–животное” (проба Go), “животное–растение” (проба NoGo), “растение–растение” (проба Ignore (или p–p)), и “растение-человек” (проба Novel (или p–h)). Novel-проба сопровождалась звуковым сигналом, представляющим собой комбинацию тонов частотой 5000 Гц, 2500 Гц, 2000 Гц, 1500 Гц и 1000 Гц в случайном порядке. Каждый тон длился 20 мс, суммарная длительность звукового сигнала составляла 100 мс. Изображения животных, растений и людей предъявлялись на экране 19-дюймового монитора, в пробах “животное–животное” (Go) и “растение–растение” (Ignore) первое и второе изображения идентичны. Пробы подавались в случайном порядке с вероятностью 25%. Длительность проб – 3000 мс, престимульный интервал – 300 мс, длительность обоих стимулов – 100 мс, длительность интервала между двумя стимулами в паре – 1000 мс, интервал между парами стимулов 1800 мс. Общая продолжительность теста – 20 мин. Испытуемым давалась инструкция нажимать на кнопку указательным пальцем правой руки как можно быстрее в случае предъявления пары картинок “животное–животное” и не нажимать в случае предъявления других пар стимулов.
Рис. 1.
Схематическое изображение двустимульного теста в парадигме Go/NoGo.
Fig. 1. Schematic illustration of a two-stimulus test in the Go/NoGo paradigm.
Регистрацию электроэнцефалограммы (ЭЭГ) проводили для каждого испытуемого в состоянии покоя и во время выполнения тестового задания. Для отведения ЭЭГ использовались 19 электродов (международная система расположения электродов 10–20) с размещением объединенных референтных электродов на мочках ушей. Запись осуществляли монополярно посредством компьютерного электроэнцефалографа “Мицар-ЭЭГ” с частотой дискретизации 500 Гц. Параметры фильтра высокой частоты (ФВЧ) и фильтра низкой частоты (ФНЧ) составляли соответственно 0.03 и 40 Гц, также использовали режекторный фильтр – 45–55 Гц. Коррекцию артефактов моргания, кардиограммы и движения глаз в записях ЭЭГ проводили средствами коммерческого программного пакета WinEEG, версия 2.82 (НПФ “Мицар”, Санкт-Петербург, авторское право В.А. Пономарев, Ю.Д. Кропотов, РФ 2001610516, 08.05.2001), использовали метод независимых компонент (ICA), для разложения на компоненты использовался алгоритм “Infomax”.
По ЭЭГ-данным, зарегистрированным во время выполнения тестового задания, методом усреднения рассчитывали потенциалы, связанные с событиями (ПСС), для каждого типа проб и для каждого испытуемого в отдельности. Далее ВП усреднялись для каждого типа проб внутри каждой из подгрупп испытуемых. Полученные ПСС сравнивались между подгруппами испытуемых для каждого типа проб по отдельности. На основе полученных разностных изображений определялось методом визуальной инспекции, в каких типах проб и на каких временных интервалах существуют различия в амплитудах ПСС между подгруппами испытуемых. Таким образом, были выделены зоны интереса – пробы Go и NoGo – и временной интервал 200–400 мс от начала предъявления второго стимула в пробе. На данном временном интервале в пробах Go и NoGo отмечается позитивная волна с максимумом в лобно-центральных отведениях, соответствующая компоненту вызванных потенциалов Р300.
Для статистического анализа выявленных различий амплитуд ПСС использовался анализ (3 × 2 × 19) ANOVA. В качестве факторов повторных измерений рассматривали три подгруппы испытуемых (ВК/НК/СК), два условия (пробы Go/NoGo) и 19 ЭЭГ отведений. Анализировались средние значения амплитуды ПСС на временном интервале 200–400 мс. Для выявления зонального распределения различий использовался метод множественных сравнений (post-hoc comparisons) с использованием критерия Tukey HSD.
Для анализа поведенческих данных (время реакции в пробе Go и количество пропусков в пробе NoGo) в трех подгруппах испытуемых использовался статистический анализ one-way ANOVA.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Статистический анализ (one-way ANOVA) показал, что время реакции при нажатии на кнопку в пробе Go достоверно различается в подгруппах ВК и НК (F(1, 20) = 7.23, p = = 0.014). Подгруппа НК характеризуется большим временем реакции. Среднее время реакции в подгруппе ВК составило 306 мс, тогда как в подгруппе НК составило 369 мс. Достоверных различий между подгруппами по количеству пропусков нажатий в пробе NoGo не выявлено.
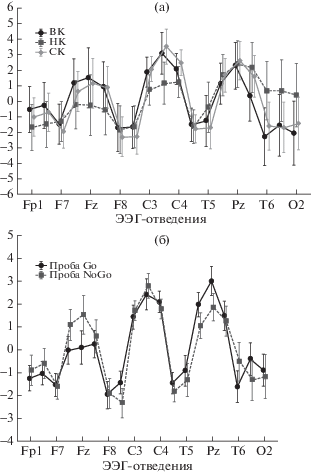
Дисперсионный анализ (3 × 2 × 19 ANOVA) показал достоверное взаимодействие факторов: Подгруппа × Отведения (F(36,630) = = 2.5102, p = 0.000005) (рис. 2 (а)) и достоверное взаимодействие факторов: Условия × Отведения (F(18,630) = 8.7556, p = 0.000000) (рис. 2 (б)). Таким образом, амплитуда компонента Р300 различается в трех подгруппах испытуемых, а также амплитуда компонента Р300 различается в пробах Go и NoGo.
Рис. 2.
Графическое отображение результатов дисперсионного анализа (3 × 2 × 19 ANOVA) амплитуд ПСС на временном интервале 200–400 мс от начала предъявления второго стимула в пробе. (а) – показано достоверное взаимодействие факторов Подгруппа (ВК/СК/НК) × Отведения (F(36,630) = 2.5102, p = 0.000005). (б) – показано достоверное взаимодействие факторов Условия (Go/NoGo) × Отведения (F(18,630) = 8.7556, p = 0.000000). Условные обозначения: по оси ординат – ЭЭГ-отведения, по оси абсцисс – средняя амплитуда ПСС (мкВ). Вертикальные полосы обозначают доверительный интервал (0.95).
Fig. 2. Graphic representation of the statistical analysis (3 × 2 × 19 ANOVA) in time interval 200–400 ms after the start of the second stimulus presentation. (а) shows a significant interaction of factors Subgroups (HC/MC/LC) × Leads (19 EEG electrodes) (F (36,630) = 2.5102, p = 0.000005). (б) shows a significant interaction of factors Conditions (Go/NoGo) × Leads (F (18,630) = 8.7556, p = 0.000000). Legend: ordinate axis – EEG leads, abscissa axis – the average amplitude of the ERP (μV). Vertical bars indicate a confidence interval (0.95).
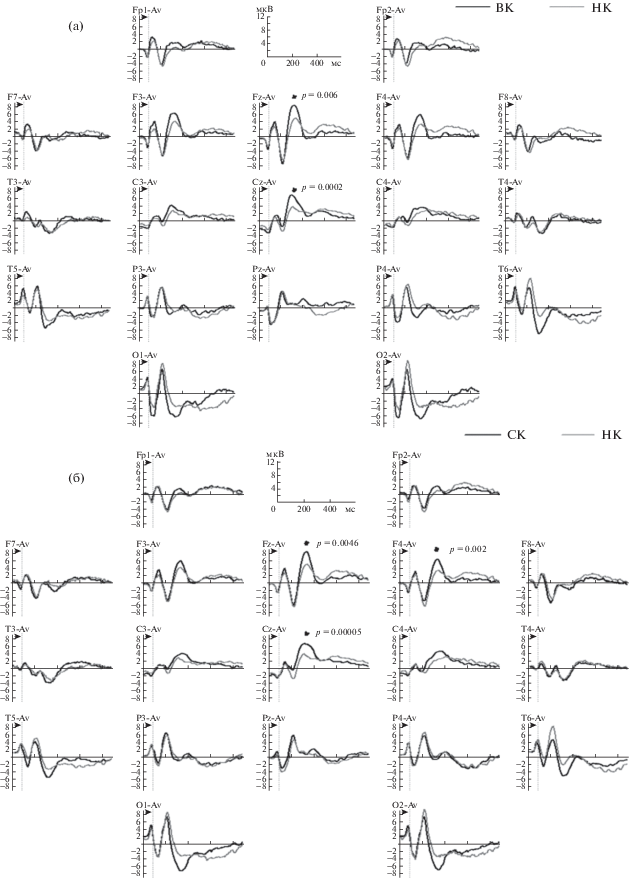
Для выявления зонального распределения различий использовался метод множественных сравнений (post-hoc comparisons) с использованием критерия Tukey HSD (р ≤ 0.05). Использование данного критерия выявило достоверные различия по амплитуде компонента Р300 между подгруппами ВК и НК в центральных отведениях Fz и Cz, в пробе NoGo (рис. 3 (а)). Также выявлены достоверные различия по амплитуде компонента Р300 между подгруппами СК и НК в отведениях Fz, F4 и Cz, а также в пробе NoGo (рис. 3 (б)). При рассмотрении рисунка групповых ПСС (рис. 3 (а), 3 (б)) видно, что подгруппы К и СК отличаются от подгруппы НК большей амплитудой позитивной волны на интервале 200–400 мс с максимумом на 332 мс. В пробе Go критерий Tukey HSD не выявил достоверных зональных различий между подгруппами испытуемых.
Рис. 3.
Групповые ПСС в пробе NoGo в подгруппах испытуемых ВК и НК (а) и в подгруппах испытуемых СК и НК (б). Условные обозначения: по оси ординат – амплитуда (мкВ), по оси абсцисс – время (мс), показан интервал 900 мс от начала предъявления второго стимула в пробе. Вертикальный пунктир – время окончания предъявления второго стимула в пробе. Звездочками отмечены конкретные отведения, в которых выявлены достоверные различия в амплитудах ПСС между двумя сравниваемыми подгруппами испытуемых (с использованием критерия Tukey HSD (р ≤ 0.05).
Fig. 3. Event related potentials (NoGo trial) in the subgroups of high-creative and low-creative subjects (a) and in the subgroups of middle-creative and low-creative (б). Legend: ordinate axis – amplitude (μV), abscissa axis – time (ms), an interval of 900 ms from the beginning of second stimulus in the trial is shown. The vertical dashed line indicates the end of the second stimulus presentation. Asterisks indicate leads, where significant differences in ERP between two compared subgroups were revealed (using the Tukey HSD criterion (p ≤ 0.05).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате анализа выявлены различия между подгруппами испытуемых с высоким (ВК) и низким уровнем (НК) вербальной креативности, а также между подгруппами со средним (СК) и низким уровнем (НК) вербальной креативности по амплитуде компонента вызванных потенциалов Р300. При этом подгруппа СК занимает промежуточное положение между подгруппами ВК и НК (рис. 2 (а)). Направленность выявленных различий между подгруппами одинакова в пробах Go и NoGo (рис. 2 (б)). Подгруппы ВК и СК отличаются от подгруппы НК большей амплитудой позитивной волны на временном интервале 200–400 мс, данная волна имеет максимум на 332 мс (рис. 3). Но уровня статистической достоверности различия достигают только в пробе NoGo. Локализация выявленных различий в переднецентральных отведениях и временной интервал компонента, по амплитуде которого выявлены различия, позволяет соотнести эту позитивную волну с компонентом вызванных потенциалов Р300.
Компонент Р300, выявленный в пробе NoGo, может соответствовать субкомпоненту “no-go P300”, который выявляется в различных вариантах oddball-парадигмы в ответ на игнорируемый стимул [Kok, 1986]. Данный субкомпонент имеет максимальную амплитуду в лобно-теменных областях и обычно связывается с механизмами подавления ответа [Falkenstein et al., 1999; Azizian et al., 2006]. В пробе NoGo после второго стимула испытуемый как раз должен воздержаться от нажатия на кнопку.
В исследованиях творческой деятельности компоненту Р300 уделяется достаточно большое внимание. Так, например, показано, что амплитуда компонента Р300 при выполнении задания с меняющимися правилами или меняющимися характеристиками стимулов может быть показателем степени когнитивной гибкости (flexibility), способности легко переключаться с задачи на задачу [Barcelo et al., 2002]. В исследованиях вербального творческого мышления, связанного с инсайтным типом решения творческих задач, который включает в себя разрушение старого гештальта (не приводящего к решению проблемы) и формирование нового гештальта, который как раз и обеспечивает успешное решение задачи, показано, что компонент P300 связан именно с формированием нового гештальта [Zhang et al., 2015]. Также при исследовании невербальной креативности показана связь компонента P300 со способностью к обучению музыкальной грамоте и музыкальной креативности в целом [Zioga et al., 2020] и связь компонента P3b со способностью к джазовой музыкальной импровизации и с показателями беглости и оригинальности по тесту, измеряющему дивергентное мышление [Przysinda et al., 2017]. В книге “Нейробиология исключительности” [Stough, 2005], в главе, посвященной нейробиологии креативности, делается предположение, что различия между индивидами с высоким и низким уровнем креативности могут быть выявлены при анализе именно компонента P300. Амплитуда компонента Р300 в отдельных работах также соотносится с определенными особенностями личности испытуемых, такими как уровень мотивации [Hansenne, 1999; Johnson, 1993] или уровень самоконтроля [Nash et al., 2013].
Анализ поведенческих данных показал, что подгруппа ВК отличается меньшим временем реакции от подгруппы НК. Сопоставление поведенческих данных и полученных данных ПСС при выполнении Go/NoGo теста позволяет предположить, что подгруппа испытуемых с высоким уровнем вербальной креативности отличается более эффективной работой механизмов внимания, обеспечивающих процесс подавления ответа и, возможно, процесс вовлечения в действие. Эти особенности внимания у данных подгрупп испытуемых могут быть обусловлены большей когнитивной гибкостью, большей мотивацией и большим уровнем самоконтроля при выполнении теста. Полученные в работе данные могут рассматриваться в пользу точки зрения, что творческие люди легче регулируют фокус внимания в зависимости от требований той или иной задачи.
ЗАКЛЮЧЕНИЕ
В результате работы показано, что подгруппы испытуемых с высоким и средним уровнем вербальной креативности отличаются от подгруппы испытуемых с низким уровнем вербальной креативности большей амплитудой позднего позитивного компонента Р300. Направленность выявленных различий между подгруппами одинакова в пробах Go и NoGo, но достоверные различия выявлены только в пробе NoGo. Полученные данные позволяют предположить, что данные подгруппы испытуемых различаются механизмами внимания, которые связаны с процессами подавления ответа и, возможно, с процессами вовлечения в действие.
Список литературы
Истратова О.Н., Эксакусто Т.В. Психодиагностика: коллекция лучших тестов. Ростов-на-Дону: Феникс, 2006. 375 с.
Ansburg P.I., Hill K. Creative and analytic thinkers differ in their use of attentional resources. Personality and Individual Differences. 2003. 34: 1141–1152.
Azizian A., Freitas A.L., Watson T.D., Squires N.K. Electrophysiological correlates of categorization: P300 amplitude as an index of target similarity. Biol. Psychol. 2006. 71: 278–288.
Barcelo F., Perianez J.A., Knight R.T. (2002). Think differently: a brain orienting response to task novelty. Neuroreport. 2002. 13 (15): 1887–1892.
Dietrich A., Kanso R. A Review of EEG, ERP, and Neuroimaging Studies of Creativity and Insight. Psychological Bulletin. 2010. 136 (5): 822–848.
Eysenck H.J. Creativity and personality: Suggestions for a theory. Psychological Inquiry. 1993. 4: 147–178.
Falkenstein M., Hoormann J., Hohnsbein J. ERP components in the Go/No– Go tasks and their relation to inhibition. Acta Psychol. (Amst.). 1999. 101: 267–291.
Hansenne M. P300 and personality: an investigation with the Cloninger’s model. BiolPsychol. 1999. 50 (2): 143–155.
Johnson R. Jr. On the neural generators of the P300 component of the event-related potential. Psychophysiology. 1993. 30 (1): 90–97.
Kok A. Effects of degradation of visual stimulation on components of event-related potentials (ERP) in Go/NOGO reaction task. Biol. Psychol. 1986. 23: 21–38.
Luck S.J. An Introduction to the Event-Related Potential Technique. Second Edition. A Bradford Book. 2014. 416 p.
Martindale C. Creativity and connectionism. The creative cognition approach. Ed. Smith S., Ward T., Finke R. Cambridge: MIT Press, 1995. 249–268.
Mednick S.A., Mednick M.T. Examiner’s manual: Remote Associates Test. Boston. MA: Houghton Mifflin, 1967.
Nash K., Schiller B., Gianotti L.R.R., Baumgartner T., Knoch D. Electrophysiological Indices of Response Inhibition in a Go/NoGo Task Predict Self Control in a Social Context. PLoS ONE. 2013. 8 (11): e79462.
Oldfield R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia. 1971. 9 (1): 97–113.
Przysinda E., Zeng T., Maves K., Arkin C., Loui P. Jazz musicians reveal role of expectancy in human creativity. Brain and Cognition. 2017. 119: 45–53.
Stough C. Neurobiology of exceptionality: the biology of normal and abnormal traits. New York: Kluwer Academic/Plenum Publishers, 2003. 219 p.
Zhang Z., Xing Q., Li H., Warren C. M., Tang Z., Che J. Chunk decomposition contributes to forming new mental representations: An ERP study. Neuroscience Letters. 2015. 598: 12–17.
Zioga I., Harrison P.M.C., Pearce M.T., Bhattacharya J., Di Bernardi Luft C. From learning to creativity: Identifying the behavioural and neuralcorrelates of learning to predict human judgements of musical creativity. NeuroImage. 2020. 206:116311.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова