

Журнал высшей нервной деятельности им. И.П. Павлова, 2019, T. 69, № 4, стр. 493-504
У ВЫСОКОИМПУЛЬСИВНЫХ КРЫС РЕАКЦИЯ УСЛОВНОРЕФЛЕКТОРНОГО СТРАХА ВЫРАЖЕНА СЛАБЕЕ, ЧЕМ У НИЗКОИМПУЛЬСИВНЫХ ЖИВОТНЫХ
И. В. Павлова 1, *, М. И. Зайченко 1, Г. Х. Мержанова 1, Г. А. Григорьян 1
1 Федеральное государственное бюджетное учреждение науки
Институт высшей нервной деятельности и нейрофизиологии Российской академии наук
Москва, Россия
* E-mail: pavlovfml@mail.ru
Поступила в редакцию 28.02.2018
После доработки 02.03.2018
Принята к публикации 14.05.2018
Аннотация
С помощью методики delay discounting крыс классифицировали на высокоимпульсивных (предпочитавших малоценное немедленное пищевое подкрепление), низкоимпульсивных (выбирающих задержанное на 5 с высокоценное подкрепление) и амбивалентных, у которых предпочтение выбора подкрепления отсутствовало. У высокоимпульсивных крыс условнорефлекторная реакция страха, оцениваемая по реакции замирания, была выражена менее интенсивно, чем у низкоимпульсивных и амбивалентных животных. Различия между группами проявились на стадии обучения и при тестировании рефлекса спустя 24 ч после обучения. Различия наблюдались как при действии сигнального раздражителя (звука), так и при исследовании контекста. Угашение условнорефлекторной реакции страха у высокоимпульсивных крыс осуществлялось быстрее, чем у низкоимпульсивных и амбивалентных животных. Пороги болевых реакций (вздрагиваний и прыжков) не отличались у трех групп крыс. Полученные данные рассматриваются с позиции представлений о функциональной и дисфункциональной форм импульсивности, и соответствующих им проявлений условнорефлекторного страха и аффективных негативных расстройств.
Импульсивность – это склонность субъекта к быстрым, необдуманным и зачастую неоправданно рискованным действиям, способным привести к тяжким негативным последствиям, хотя благоприятный исход также не является исключением. Оценка импульсивности часто встречается в характеристиках нормального индивидуального поведения человека. Между тем чрезмерная или “патологическая” импульсивность является важным симптомом ряда серьезных психопатологий, таких как синдром дефицита внимания и гиперактивного расстройства [Miller et al., 2010], наркомании [Jupp, Dalley, 2014], шизофрении [Yu et al., 2017], биполярного расстройства [Bauer et al., 2017], болезни Паркинсона [Nombela et al., 2014 ] и др. Не случайно в последние годы резко усилился интерес к этой проблеме и разработке для ее исследования адекватных методических приемов. На сегодняшний день почти все существующие оценки импульсивного поведения построены на методиках с положительным подкреплением. Среди них наиболее часто применяется методика delay discounting, которая позволяет экспериментатору оценить психологическое восприятие субъектом (животным) величины подкрепления, зависящее от времени его достижения. Оценка осуществляется на основе выбора субъектом слабого (по размеру/качеству) подкрепления или сильного (большего или более ценного), но задержанного подкрепления. Предпочтение слабого немедленного подкрепления характеризует импульсивное поведение, а выбор сильного, но задержанного подкрепления – способность к самоконтролю [Green, Sayderman, 1980; Мержанова, Берг, 1991; Cardinal et al., 2002]. Из других приемов оценки импульсивного поведения применяют методы 5-местной условнорефлекторной дифференцировки и стоп-сигнальной реакции [Eagle et al., 2008; Robbins, 2002]. В первом случае изучают состояние механизмов внимания при импульсивности, а во втором – способность сдерживать или прекращать начатое действие.
Исследование реакции при использовании болевого подкрепления у крыс с высокой и низкой степенью импульсивности практически не проводилось, если не считать нашей недавней работы [Зайченко и др., 2018] и работы южно-корейских авторов П. Ким и соавт. [Kim et al., 2012]. В работе [Зайченко и др., 2018] было показано, что высокоимпульсивные животные лучше, чем низкоимпульсивные обучаются реакции двустороннего избегания в челночной камере. П. Ким и соавт. [Kim et al., 2012] исследовали влияние электрокожного болевого раздражения на частоту подходов к поилке и попыток питья из нее. При этом животные, чтобы подойти к поилке, должны были преодолеть участок пола, на который подавался электрический ток. Но главное, это то, что авторы использовали для этой цели не обычных крыс, а специально выведенную линию спонтанно гипертензивных крыс, обладающих помимо высокого артериального давления, еще и хорошо выраженными импульсивными свойствами [Calzavara et al., 2009; Togashi et al , 2004; Ledoux et al., 1983]. Спонтанно гипертензивные крысы делали больше попыток подойти к поилке через электрифицированный пол камеры, чем контрольные нормотензивные крысы (Wistar-Kyoto), что косвенно свидетельствует об их меньшем страхе перед электрокожным болевым раздражением. Пороги болевых реакций при этом у контрольных и спонтанно гипертензивных крыс были одинаковыми. Еще раньше Дж. Леду и соавт. [Ledoux et al, 1983], а после них Х. Тогаши и соавт. [Togashi et al., 2004], используя процедуру выработки классического оборонительного условного рефлекса, показали, что у спонтанно гипертензивных крыс страх проявляется слабее, чем у контрольных животных. Таким образом, кроме данных, полученных на линии спонтанно гипертензивных крыс, отличающихся еще и повышенной импульсивностью, в литературе нет прямых результатов, указывающих на зависимость проявлений условнорефлекторного страха от степени импульсивности человека и животных.
В настоящей работе мы поставили задачу исследовать вопрос, как связана импульсивность, определяемая по методике delay discounting, c проявлением условнорефлекторного страха, оцениваемого по реакции замирания.
МЕТОДИКА
Объект исследования. Опыты проводили в условиях хронического эксперимента на 22 крысах-самцах линии Вистар массой 330–400 г, полученных из филиала “Столбовая” ФГБУН НЦБМТ ФМБА России. Животные содержались в виварии при обычном 12-часовом световом режиме со свободным доступом к воде и стандартному корму в клетках по 5–6 крыс. В экспериментах соблюдали принципы гуманности, изложенные в директивах Европейского Сообщества (2010/63/ЕU), и положения ИВНД и НФ РАН о работе с экспериментальными животными.
Определение уровня импульсивности с применением методики “права выбора ценности пищевого подкрепления (delay discounting)”. При обучении крыс для поддержания пищевой мотивации в виварии количество подаваемой пищи ограничивали так, чтобы масса крысы составляла приблизительно 80% от массы животного при свободном доступе к пище. Экспериментальная установка представляла собой прямоугольную камеру размером 30 × 60 × 30 см (камера № 1). В середине передней стенки камеры была встроена кормушка, доступ в которую загораживала подвижная шторка из прозрачного пластика. С двух сторон от кормушки размещались выдвижные педали. В кормушке и над каждой из педалей находились сигнальные лампочки. Предварительно крыс обучали в экспериментальной установке нажимать на две педали для получения пищи – стандартной пищевой гранулы весом 45 мг производства фирмы “Bio3Serv” (США). Обучение проводилось до тех пор, пока крыса с равной вероятностью не нажимала на обе педали. Затем животным предоставлялась возможность выбирать либо малоценное немедленное подкрепление, либо ценное, но задержанное пищевое подкрепление. Схема экспериментальной пробы была такова. После запуска экспериментальной программы в камере зажигалась лампочка общего освещения камеры и одновременно лампочка в кормушке. В течение 10 с крыса могла отодвинуть носом шторку, закрывающую доступ в кормушку, что приводило к выдвижению обеих педалей, которые оставались выдвинутыми в течение 10 с. При нажатии на правую педаль крыса получала 1 пелетку сразу, на левую – 4 пелетки с задержкой 5 с. Во время ежедневных экспериментов без перерыва на выходные дни крыса осуществляла 25 проб до достижения критерия стабильности – предпочтение одной и той же педали в течение 10 дней. Это занимало от 1 до 2 мес. В каждом опыте анализировали число нажатий на педали для получения малоценного немедленного подкрепления (k1), число нажатий для получения более ценного, но задержанного подкрепления (k2). Коэффициент импульсивности рассчитывали по формуле Кимп = k2/(k1 + k2).
После разделения крыс на группы с высокой, низкой и средней (амбивалентные животные) степенью импульсивности переходили к выработке у них реакции условнорефлекторного страха.
Определение порогов болевой чувствительности. Крыс всех групп тестировали на предмет болевой чувствительности при электрокожном раздражении. Опыты проводили в камере № 2 (Startle and Fear Combined System производства PanLab Harvard apparatus, Spain, 2000). Внутренняя камера размером 24 × 24 × 24 см, куда помещали животное, располагалась на четырех датчиках, фиксирующих движения крысы. Пол камеры представлял собой решетку, на которую подавался ток нарастающей силы (0, 0.025, 0.05 мА и далее с шагом 0.05 до 0.35 мА) с межстимульным интервалом 90–120 с, длительностью 0.5 с. Ток определенной силы предъявлялся только один раз. Реакцию крысы на ток определяли визуально и по записи механограммы. Для оценки характера реакции крысы и для более четкого выделения реакции замирания механограмму анализировали за 5 с до предъявления электрокожной стимуляции, в течение 0.5 с во время ее действия и еще в течение 50 с после окончания стимуляции. Определяли пороги вздрагивания, прыжков и вокализации в слышимом для человека диапазоне. После каждой крысы камеру протирали 20% раствором этилового спирта для устранения запаха. После определения порогов болевой чувствительности в течение двух дней угашали реакцию страха на обстановку, для чего крыс помещали в камеру на 10 мин. Во всех последующих экспериментах в той же камере существенно изменяли контекст: покрывали опилками пол, заклеивали бумагой стены и т.д.
Выработка, тестирование и угашение реакции условнорефлекторного страха. Через 27–28 дней после определения порогов проводили выработку классического оборонительного условного рефлекса в камере № 2 (Startle and Fear Combined System производства PanLab Harvard apparatus, Spain, 2000). При об-учении после 120-секундного периода ознакомления животных с камерой предъявляли 3 сочетания звука (30 с, 80 дБ, 2000 Гц) и электрокожного раздражения лап через решетку пола (2 с, 0.8 мА, задержка 28 с от начала действия звука) с 40–60 с межстимульными интервалами. Через 24 ч после обучения тестировали реакцию условнорефлекторного страха (Тест 1). При этом после 120 с периода исследования камеры предъявляли звук (120 с, 80 дБ, 2000 Гц), после которого следовал 120 с период без звуковых стимулов (последействие). После каждой крысы камеру протирали 1–2% раствором уксуса для устранения запаха.
В дальнейшем (через 2–3 дня) в трех опытах угашали реакцию условнорефлекторного страха, при этом после 120 с периода исследования камеры следовало 10 изолированных звуковых стимулов (30 с, 80 дБ, 2000 Гц) без электрокожных раздражений с 20 с межсигнальными интервалами. Через 24 ч после каждого опыта с угашением тестировали сохранность рефлекса (соответственно Тесты 2, 3 и 4).
Исследовали поведение крыс в интервалы времени до, во время и после действия звука. По механограмме с помощью амплитудного и временного порогов выделяли периоды неподвижности (замирания) крыс длительностью не менее 2 с, когда наблюдались только периодические дыхательные движения животного. Подсчитывали процент времени замирания от общего времени каждого поведенческого этапа, а также число эпизодов замирания и среднее время таких эпизодов. Обработка проводилась с помощью стандартной программы, прилагающейся к установке фирмы Panlab. Кроме того, после каждого опыта для определения степени эмоционального напряжения подсчитывали число уринаций и дефекаций.
Статистический анализ. Для статистической обработки результатов использовали стандартную программу STATISTICA 8.0. При сопоставлении групп крыс применяли дисперсионный анализ: многофакторный анализ (Factorial ANOVA), однофакторный анализ (One-way ANOVA) и для повторных измерений (Repeated measures ANOVA). Анализировали влияние факторов “ГРУППА КРЫС”, “НОМЕР ОПЫТА”, “ИНТЕРВАЛ ВРЕМЕНИ”. При post-hoc анализе использовали критерий Fisher LSD. Различия считали статистически значимыми при p < 0.05, отмечали наличие тенденции при 0.05 ≤ р ≤ 0.1. С применением Basic Statistics строили гистограммы распределения крыс в зависимости от их коэффициента импульсивности. Данные на рисунках представлены в виде средних величин ± ошибки средних.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
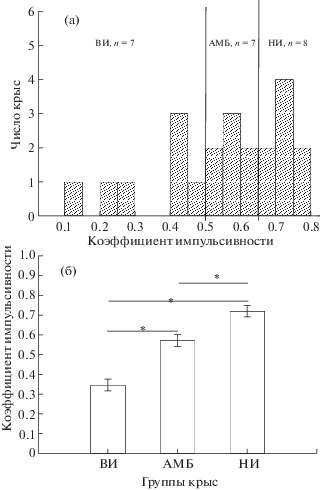
Выделение групп крыс с высокой, средней и низкой степенью импульсивности. На рис. 1 (а) показано распределение крыс в зависимости от коэффициента импульсивности, вычисленного на основе методики delay discounting, т.е. оценки предпочтения выбора небольшого немедленного или большого задержанного подкреплений. Среднее значение распределения составляло 0.553, стандартная ошибка среднего 0.037, стандартное отклонение 0.175. Учитывая форму гистограммы распределения, выборку крыс делили примерно на три равные части. Крыс с низким коэффициентом импульсивности Кимп. < 0.5 относили к группе высокоимпульсивных животных (ВИ, n = 7). Эти крысы предпочитали небольшое, но немедленное подкрепление. Крысы с коэффициентом импульсивности от 0.5 до 0.65 составляли промежуточную или амбивалентную группу (АМБ, n = 7), а животные с показателем Кимп > 0.65 составляли группу низкоимпульсивных или способных к самоконтролю крыс (НИ, n = 8). Животные этой группы были в состоянии ждать большое, но задержанное подкрепление. Амбивалентные животные проявляли реакции обоих типов, т.е. они могли чередовать выбор, как небольшого немедленного, так и большого задержанного подкреплений. Анализ с помощью One-way ANOVA выявило влияние фактора “ГРУППА КРЫС” [F(2, 19) = 40.976, p < 0.000] (1 (б)), post-hoc анализ обнаружил, что крысы НИ группы отличались, как от ВИ, так и АМБ группы, статистически значимые различия были обнаружены также между ВИ и АМБ группой.
Рис. 1.
Выделение и сопоставление групп высокоимпульсивных (ВИ), амбивалентных (АМБ) и низкоимпульсивных крыс (НИ). (а) – Распределение крыс в зависимости от коэффициента импульсивности. По оси абсцисс – коэффициент импульсивности в относительных единицах, по оси ординат – число крыс. (б) – средний коэффициент импульсивности в разных группах. По оси ординат – коэффициент импульсивности в относительных единицах, по оси абсцисс – группы крыс. n – число крыс в группе. * – статистически значимые различия между группами (One-way ANOVA, p < 0.05).
Fig. 1. A selection and comparison of highly impulsive (ВИ), low impulsive (НИ) and ambivalent groups of rats. (а) – distribution of rats depending on the coefficient of impulsivity. On horizontal axis – the coefficient of impulsivity in relative units, on vertical line – a number of rats. (б) – an average coefficient of impulsivity in the different groups. On ordinate axis – the coefficient of impulsivity in relative units, on abscissa axis – the groups of rats. n – a number of rats in a group. * – the significant differences between groups (One-way ANOVA, p < 0.05).
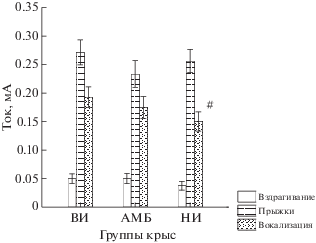
Сопоставление порогов болевых и эмоциональных реакций не выявило статистически значимых различий между группами с разной степенью импульсивности. Пороги реакции вздрагивания и прыжков (рис. 2) были примерно одинаковыми у всех групп, а пороги вокализации в слышимом человеком диапазоне были чуть ниже (тенденция) для группы НИ крыс по сравнению с крысами ВИ группы. В среднем для всех групп пороги вздрагивания составляли 0.047 ± 0.024 мА, пороги прыжков – 0.255 ± 0.057 мА, а пороги вокализации – 0.180 ± 0.061 мА.
Рис. 2.
Пороги вздрагивания, прыжков и вокализации у крыс разных групп. По оси ординат – величина тока электрического раздражения решетки пола, мА. По оси абсцисс – группы крыс. # – тенденция к снижению порога в группе НИ по сравнению с ВИ крысами (p = 0.099).
Fig. 2. The threshold of flinch, jumpings and vocalizations in rats of the different groups. On ordinate axis – the current intensity of electrical stimulation of a floor grid, mА. On abscissa axis – the groups of rats. # – p = 0.099.
Выработка, тестирование и угашение условнорефлекторной реакции страха у крыс с разной степенью импульсивности. На рис. 3 (а, б) видно, что три последовательных, сочетанных с болевым электрокожным раздражением предъявления звука вызывают отчетливую выработку реакции условнорефлекторного страха. Это проявляется в прогрессивном увеличении времени замирания у всех групп животных, начиная со второго и третьего применения условного раздражителя, а также в межсигнальные интервалы. Высокий процент замирания остается и в последействии. Дисперсионный факторный анализ (Factorial ANOVA) показал влияние факторов “ГРУППА КРЫС” [F(2,133) = 13.25, p < 0.001] и “ИНТЕРВАЛ ВРЕМЕНИ” [F(6,133) = 7.27, p = = 0.000] (рис. 3 (б)) на процент времени замирания крыс. Post-hoc анализ выявил, что различия между группами обусловлены существенной разницей в процентах времени замирания между крысами групп ВИ и АМБ (р < 0.000), а также ВИ и НИ (р < 0.000), между животными группы АМБ и НИ статистически значимых различий не наблюдалось (р = 0.261). Различия между группами проявились при втором и третьем действии сигнального раздражителя и во время второго межсигнального периода. На рис. 3 (б) видно, что проявления реакции замирания в ходе обучения быстрее увеличиваются у крыс НИ и АМБ групп по сравнению с ВИ группой. При тестировании рефлекса через 24 часа (Тест 1) (рис. 3 (в)) влияние факторов ГРУППА КРЫС [F(2,57) = 7.85, p = 0.001] и ИНТЕРВАЛ ВРЕМЕНИ [F(2,57) = 5.60, p = 0.006] оставалось высокозначимым. Post-hoc анализ показал, что время замирания крыс группы НИ было больше при действии звука и в последействии, чем крыс ВИ группы (рис. 3 (в)). Однофакторный дисперсионный анализ (One-way ANOVA) не обнаружил значимых различий в числе болюсов и уринаций у всех групп крыс в Тесте 1 (рис. 3 (г)). Вместе с тем средняя длительность эпизодов замирания была значимо больше у крыс НИ группы по сравнению с ВИ животными (рис. 3 (г)).
Рис. 3.
Поведение крыс разных групп при выработке и тестировании классического оборонительного рефлекса. (а) – схема эксперимента. (б) – время замирания крыс (%) при выработке условного рефлекса; по оси абсцисс – периоды обучения: “До” – до первого сочетания звука и тока, Зв1, Зв2, Зв3 – при действии первого, второго и третьего звука, МС1, МС2 – межсигнальные периоды, “После” – после трех сочетаний. (в) – время замирания (%) до звука, во время действия звука и после звука в Тесте 1. (г) – средняя длительность эпизодов замирания (с), число болюсов и уринаций в Тесте 1. (д) – сопоставление времени замирания (%) при исследовании контекста и при действии звука до обучения и в Тесте 1. * – статистически значимые различия в группах АМБ или НИ по сравнению с группой ВИ (p < 0.05), # – тенденция (0.05 < < p < 0.1). ↑ – значимое увеличение времени замирания в Тесте 1 по сравнению с периодом до обучения (Repeated measures ANOVA). Остальные обозначения как на рис. 1.
Fig. 3. The behavior of different groups of rats at elaboration and testing of classical defensive reflex. (а) – the plan of the experiment. (б) – the freezing reaction time (%) at learning; on abscissa axis – the stages of learning: “До” – before the first pairing of tone and shock, Зв1, Зв2, Зв3 – at presentations of the first, second and third tones, МС1, МС2 – intersignal periods, “После” – after three pairings. (в) – the freezing time (%) before a tone, during its presentation and after termination of tone in Test 1. (г) – the mean duration of freezing episodes (c), a number of boluses and urinations in Test 1. (д) – comparison of freezing duration (%) at context investigation and a tone presentation before learning and in the Test 1. * – the significant differences in the groups of АМБ or НИ relative to the group ВИ (p < 0.05), # – a tendency (0.05 < p < 0.1). ↑ – a significant increase the freezing time in the Test 1 compared with a period before learning (Repeated measures ANOVA). The rest designations are the same as on Fig. 1.
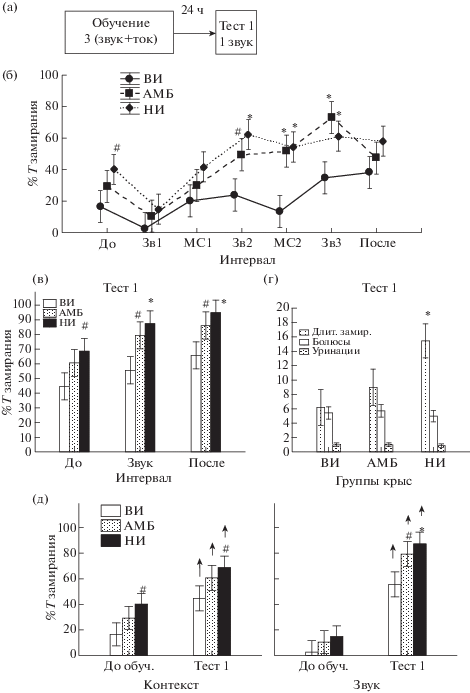
У всех групп крыс сравнивали время замирания в условиях только контекста камеры и контекста вместе с действием звука, до обучения и во время тестирования выработанного навыка спустя 24 ч после обучения (рис. 3 (д), Repeated Measures ANOVA). Было обнаружено значимое влияние фактора ГРУППА КРЫС [F(2,38) = 5.3, p = 0.009] и фактора НОМЕР ОПЫТА [F(1,38) = 102.8, p = 0.000]. Post-hoc анализ показал увеличение времени замирания у всех обученных групп животных по сравнению с необученными как в контексте, так и при действии звука на фоне контекста. Взаимодействия факторов НОМЕР ОПЫТА/ГРУППА КРЫС не было обнаружено [F(2,38) = 0.5, p = 0.615]. Однако межгрупповое различие лучше проявилось при действии контекста вместе со звуком, чем в условиях только одного контекста. В последнем случае различия проявлялись на уровне тенденции.
При угашении классического оборонительного условного рефлекса (рис. 4 (а,б)) было обнаружено влияние факторов ГРУППА КРЫС [F(2,63) = 1.58, p = 0.000] и НОМЕР ОПЫТА [F(3,189) = 34.24, p = 0.000] (Repeated Measures ANOVA). У всех групп крыс происходило снижение процента времени замирания в Тестах 3 и 4 по сравнению с Тестом 1. Межгрупповые различия, а именно больший процент времени замирания у НИ и АМБ крыс по сравнению с ВИ животными, выявленные в Тесте 1, сохранялись до последнего дня процедуры угашения. Детальный анализ по интервалам времени в Тесте 4 (рис. 4 (в)) показал, что НИ по сравнению с ВИ крысами больше времени замирали до и после звука, т.е. в ответ на контекст, а АМБ крысы больше замирали в ответ на звук по сравнению с ВИ животными.
Рис. 4.
Поведение крыс разных групп при угашении условного оборонительного рефлекса. (а) – схема эксперимента. (б) – время замирания (%) крыс в Тестах 1–4 (суммарные данные по всем периодам). (в) – время замирания (%) крыс до, во время и после действия звука в Тесте 4. (г) – время замирания (%) при исследовании контекста и при действии звука в Тестах 1 и 4. ↓ – значимое снижение в Тестах 2–4 по сравнению с Тестом 1 (Repeated measures ANOVA). * – статистически значимые различия в группах АМБ или НИ по сравнению с группой ВИ (p < 0.05), # – тенденция (0.05 < p < 0.1). Остальные обозначения как на рис. 1.
Fig. 4. The behavior of rats of the different groups at extinction of the defensive conditioned reflex. (а) - the plan of experiment. (б) – the time of freezing (%) in Tests 1–4 (the integral data on all periods). (в) – the freezing time (%) of rats before, at the tone presentation and after termination of sound in Tests 4. (г) – the freezing time (%) at the context and tone presentation in Tests 1 and 4. ↓ – a significant decrease in Tests 2–4 relative to Test 1 (Repeated measures ANOVA). * – the significant differences in groups АМБ or НИ compared with the group ВИ (p < 0.05), # – a tendency (0.05 < p < 0.1). The rest designations are the same as on Fig. 1.
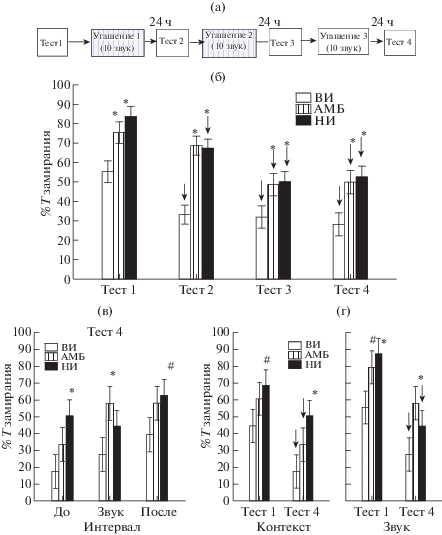
При анализе эффективности угашения сравнивали поведение в тестах 1 и 4 в ответ на контекст и на звук (рис. 4 (г), Repeated Measures ANOVA). Было обнаружено (post-hoc анализ), что лучше всего угашается рефлекс у крыс ВИ группы. У этой группы время замирания снижалось как на изолированный контекст, так и на звук. У крыс АМБ группы время замирания статистически значимо уменьшалось в ответ на контекст, а у крыс НИ группы в ответ на звук. Таким образом, угашение классического оборонительного условного рефлекса было более быстрым у ВИ группы, чем у НИ и АМБ. У НИ крыс было затруднено угашение страха на контекст, а у АМБ животных – на звук.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В настоящей работе мы показали, что условная реакция страха, оцениваемая по поведению замирания в ответ на условный раздражитель и контекст, проявляется менее интенсивно у высокоимпульсивных крыс по сравнению с низкоимпульсивными и амбивалентными животными. Эта разница наблюдается уже при втором сочетанном применении звука, сохраняется в межсигнальные интервалы, и в последействии после окончания действия трех сочетанных применений звука с электрокожным раздражением. Спустя 24 ч при повторном тестировании эти отношения между группами устойчиво сохраняются, причем разница остается высоко- значимой при действии сигнального раздражителя (звука) и проявляет тенденцию к достоверности в условиях влияния изолированного контекста. Полученные данные свидетельствуют о том, что способность крыс сдерживать активные моторные реакции может проявляться как в оборонительной ситуации в виде замирания, так и пищевой ситуации в виде долгого ожидания подкрепления.
Угашение условной реакции страха у высокоимпульсивных крыс также протекает быстрее, чем у низкоимпульсивных и амбивалентных животных, практически достигая фоновых значений на контекст и слегка превышая их в ответ на звук. Угашение условного рефлекса по своим механизмам можно рассматривать как выработку нового условного рефлекса с другим подкрепляющим (положительным) раздражителем, а именно отменой отрицательного. Ранее нами было показано [Зайченко и др., 2016], что высокоимпульсивные крысы по сравнению с низкоимпульсивными и амбивалентными животными быстрее обучаются и совершают меньше ошибок при решении пространственных и сигнальных задач в 8-канальном радиальном лабиринте. Поэтому неудивительно, что угашение у высокоимпульсивных крыс проходит быстрее, чем у крыс двух других групп.
Таким образом, у высокоимпульсивных животных условнорефлекторная реакция страха, оцениваемая по реакции замирания, проявляется слабее и угашается быстрее, чем у амбивалентных и низкоимпульсивных крыс. До нашей работы исследование зависимости условнорефлекторного страха от степени импульсивности животных проводили только на специально выведенной линии животных – спонтанно гипертензивных крысах [Togashi et al , 2004; Calzavara et al., 2009; Ledoux et al., 1983]. Эти крысы характеризуются высокоимпульсивными свойствами в модели delay discounting [Mill et al., 2005], но не проявляют импульсивных свойств в модели условнорефлекторной дифференцировки по месту [van den Bergh et al., 2006], что ограничивает их использование в качестве модели импульсивного поведения [Kim et al., 2012]. В наших опытах мы не исследовали импульсивность крыс в моделях 5-местной условнорефлекторной дифференцировки и стоп-сигнальной реакции. Но в литературе описаны случаи подобно поведению спонтанно гипертензивных крыс, у которых импульсивность в одной модели проявилась, и в других не проявилась [Winstanley, 2011]. Интересно, что пороги болевых реакций на электрокожное раздражение у крыс спонтанно гипертензивной линии [Kim et al., 2012; Calzavara et al., 2009; Togashi et al., 2004] по сравнению с контрольными животными существенно не отличались друг от друга, также как и пороги болевых реакций у высокоимпульсивных и низкоимпульсивных крыс в наших опытах. Это свидетельствует о том, что чувствительность к болевому электрокожному раздражению у разных групп животных была одной и той же, а значимые различия в проявлениях у них условнорефлекторного страха не были связаны с их разной болевой чувствительностью.
О наличии связи между уровнем страха и уровнем импульсивности косвенно свидетельствуют данные, полученные для “подросткового периода” (adolescence) человека [Cohn et al., 2013; 2016] и соответствующего этому периоду возраста животных [Pattwell et al., 2013]. Так, известно, что подростковый период человека характеризуется повышенной импульсивностью [Humphreys et al., 2016], связанной с активным исследовательским поведением и быстрым познанием окружающего мира [Spear, 2000]. С другой стороны, имеются данные, что в этом возрасте условнорефлекторный страх выражен слабее и угашается не так быстро, как в детстве и в зрелом возрасте [Humphreys et al., 2016; Baker et al., 2016].
Какие могут быть причины взаимосвязанного проявления импульсивного поведения и условнорефлекторной реакции страха у человека и животных? Одной из причин может быть участие в их реализации [Cohn et al., 2013; 2016] одних и тех же структур мозга (миндалина, гипоталамус, гиппокамп, медиальная префронтальная кора и др.). Причем важно не только, какие структуры вовлекаются в обе эти формы поведения, но и какие между ними устанавливаются функциональные связи. Так Г.Х. Мержанова и соавт. [Merzhanova, 2012] в исследованиях на кошках показали, что при самоконтролируемом поведении в условиях выбора, связанного с качеством подкрепления и разными возможностями его получения, активно вовлекаются фронто-гиппокампальные межнейронные взаимодействия, а импульсивность характеризуется активированием амигдало-фронтальной системы. У кроликов, предпочитающих пассивно-оборонительные реакции (замирание) на аверсивные воздействия по сравнению с животными с активно-оборонительными реакциями были обнаружены особенности во взаимодействии нейронов миндалины [Рысакова, Павлова, 2011], гиппокампа и неокортекса [Павлова, 2009]. У животных с длительным замиранием уровень активации миндалины был высоким. Функциональное магнитно-резонансное картирование мозга у подростков с импульсивно-агрессивным поведением показало снижение активности в миндалине и ослабление связей между миндалиной и орбито-фронтальной корой [Marsh et al., 2011]. В других подобных работах была обнаружена высокая активность в миндалине и передней цингулярной коре (структурах, связанных со страхом) во время выработки [Cohn et al., 2013] и угашения классического оборонительного условного рефлекса [Cohn et al., 2016]. В исследованиях на человеке было показано, что индивидуальное поведение может зависеть от особенностей функционирования обратных временных связей [Арзуманов, 1985].
В литературе существует точка зрения, что импульсивность отражает собой защитную реакцию организма на экстремальные негативные влияния [Cyders, Smith, 2008]. Выплескивая своими безудержными действиями негативные эмоции, импульсивные субъекты (а возможно и животные) пытаются, как бы освободиться от них. В этом смысле, рассматриваемая гипотеза не противоречит описанной выше зависимости, поскольку чем более импульсивными являются субъекты, тем больше попыток они делают, чтобы “сбросить” негативные эмоции, и при этом испытывают все меньше страха [Tomko et al., 2015; Cyders, Smith, 2008]. Близкими к этой точке зрения являются представления И. Селби и соавт. [Selby et al., 2009], согласно которым накопления негативных влияний в организме предстают в виде “эмоционального каскада”, захватывающего все помыслы и размышления субъекта. Импульсивные действия приостанавливают этот каскад и отвлекают субъект от тягостных дум и размышлений. Описанные выше гипотезы в большей своей части имеют отношение к так называемой “дисфункциональной” импульсивности [Dickman, 1990], которая ассоциируется с негативным “патологическим” воздействием на организм. С. Дикман определил ее как “тенденцию действовать непредусмотрительно в отличие от действия большинства других людей в аналогичной ситуации, приводящую в итоге к проблемам и трудностям”. В отличие от дисфункциональной импульсивности функциональную он определил, как “тенденцию действовать с относительно меньшей предусмотрительностью по сравнению с другими людьми, но в данных условиях такое поведение является оптимальным” [Dickman, 1990].
ВЫВОДЫ
1. У высокоимпульсивных крыс условно-рефлекторная реакция страха, оцениваемая по реакции замирания, была выражена менее интенсивно, чем у низкоимпульсивных и амбивалентных животных. Различия между группами проявились на стадии обучения и при тестировании рефлекса спустя 24 ч после обучения. Они сохранялись как при действии сигнального раздражителя (звука), так и при исследовании контекста.
2. Угашение условнорефлекторной реакции страха у высокоимпульсивных крыс осуществлялось быстрее, чем у низкоимпульсивных и амбивалентных животных.
3. Пороги болевых реакций (вздрагивания и прыжки) на электрокожное раздражение были одинаковыми у всех групп крыс с разным уровнем импульсивности.
4. Полученные данные сопоставляются с данными на человеке с привлечением гипотез о функциональной и дисфункциональной форм импульсивности, и соответствующих им проявлений условнорефлекторного страха и аффективных негативных расстройств.
Список литературы
Арзуманов Ю.Л. Функциональное значение обратных временных связей в структуре условного рефлекса у человека. Диссертация на соискание ученой степени доктора медицинских наук. М. 1985. 281 с. Институт высшей нервной деятельности АН СССР.
Зайченко М.И., Григорьян Г.А., Мержанова Г.Х. Реакции избавления и избегания на собственное и чужое болевое электрокожное раздражение у крыс с высокой и низкой степенью импульсивности Журн. высш. нервн. деят. 2018. 68 (4): 477–487.
Зайченко М.И., Григорьян Г.А., Мержанова Г.Х. У высокоимпульсивных крыс сигнальная память в 8-канальном радиальном лабиринте проявляется лучше, чем у низкоимпульсивных животных. Журн. высш. нервн. деят. 2016.66 (5): 600–610.
Мержанова Г.Х., Берг A.M. Выбор качества подкрепления, зависящего от времени задержки инструментальной реакции у кошек. Журн. высш. нервн. деят. 1991. 41 (5): 948–954.
Павлова И.В. Взаимодействие нейронов гиппокампа и неокортекса в эмоционально-негативных ситуациях у активных и пассивных кроликов. Журн. высш. нервн. деят. 2009. 59 (1): 75–86.
Рысакова М.П., Павлова И.В. Взаимодействие нейронов базального и центрального ядер миндалины у кроликов с активной и пассивной стратегией поведения в эмоционально-негативных ситуациях. Журн. высш. нервн. деят. 2011. 61 (2): 190–203.
Baker K.D., Bisby M.A., Richardson R. Impaired fear extinction in adolescent rodents: Behavioural and neural analyses. Neurosci. Biobehav. Rev. 2016. 70: 59–73.
Bauer I.E., Meyer T.D., Sanches M., Spiker D., Zunta-Soares G., Soares J.C. Are self-rated and behavioural measures of impulsivity in bipolar disorder mainly related to comorbid substance use problems? Cogn Neuropsychiatry. 2017. 22 (4): 298–314.
Cardinal R.N., Daw N., Robbins T.W., Everitt B.J. Local analysis of behaviour in the adjusting-delay task for assessing choice of delayed reinforcement. Neural Networks. 2002. (15): 617–634.
Calzavara M.B., Medrano W.A., Levin R., et al. Neuroleptic drugs revert the contextual fear conditioning deficit presented by spontaneously hypertensive rats: A potential animal model of emotional context processing in schizophrenia? Schizophrenia Bulletin. 2009. 35 (4): 748–759.
Cohn M.D., Popma A., van den B.W., et al. Fear conditioning, persistence of disruptive behavior and psychopathic traits: an fMRI study. Translational Psychiatry, 2013. 3, e319.
Cohn M.D., van Lith K., Kindt M., et al. Fear extinction, persistent disruptive behavior and psychopathic traits: fMRI in late adolescence Soc. Cogn. Affect. Neurosci. 2016. 11 (7): 1027–1035.
Cyders M.A., Smith, G.T. Emotion-based dispositions to rash action: positive and negative urgency. Psychol. Bull.2008. 134. 807–828.
Dickman S.J. Functional and dysfunctional impulsivity: personality and cognitive correlates. J. Pers. Soc. Psychol. 1990. 58 (1): 95–102.
Eagle D.M., Baunez C., Hutcheson D.M., Lehmann O., Shah A.P., Robbins T.W. Stop signal reaction time task performance: role of prefrontal cortex and subthalamic nucleus. Cereb. Cortex. 2008. 18 (1): 178–188.
Green L., Sayderman M. Choice between rewards differing in amount and delay toward a choice model of self-control//J. Exptl. Anal. Behav. 1980. 34: 135–147.
Humphreys K.L., Telzer E.H., Flannery J., Goff B., Gabard-Durnam L., Gee D.G., Lee S.S., Tottenham N. Risky decision making from childhood through adulthood: Contributions of learning and sensitivity to negative feedback. Emotion. 2016. 16: 101–109.
Jupp B., Dalley J.W. Convergent pharmacological mechanisms in impulsivity and addiction: insights from rodent models. Br. J. Pharmacol. 2014. 171 (20): 4729–4766.
Kim P., Choi I., Pena I.C. et al. Impulsive Behavior in an Animal Model of Attention Deficit Hyperactivity Disorder (ADHD) of the Spontaneously Hypertensive Rats. Biomol. Ther (Seoul). 2012. 20 (1): 125–131.
Ledoux J.E., Sakaguchi A., Reis D.J. Strain differences in fear between spontaneously hypertensive and normotensive rats. Brain Res. 1983. 277: 137–143.
Marsh A.A., Finger E.C., Fowler K.A., Jurkowitz I.T.N., Schechter J.C., Yu H.H., Pine D.S., Blair R.J.R. Reduced amygdala-orbitofrontal connectivity during moral judgments in youths with disruptive behavior disorders and psychopathic traits. Psychiatry Res. 2011. 194 (3): 279–286.
Merzhanova G.Kh. Impulsivity and Self-control in Animals and Humans during Choice of Behavioral Strategy. In: Psychology of Impulsivity. Ed: M.A. Cyders. Nova. 2012. Chapter 6: 183–208.
Mill J., Sagvolden T., Asherson P. Sequence analysis of Drd2, Drd4, and Dat1 in SHR and WKY rat strains. Behav. Brain. Funct. 2005. 1: 24.
Miller D.J, Derefinko K.J., Lynam D.R., Milich R., Fillmore M.T. Impulsivity and Attention Deficit-Hyperactivity Disorder: Subtype Classification Using the UPPS Impulsive Behavior Scale J Psychopathol Behav. Assess. 2010. 32 (3): 323–332.
Nombela C., Rittman T., Robbins T.W., Rowe J.B. Multiple modes of impulsivity in Parkinson’s disease. PLoS One. 2014. 9 (1): e85747.
Pattwell S.S., Lee F.S., Casey B.J. Fear learning and memory across adolescent development. Hormones and Behavior Special Issue: Puberty and Adolescence Horm. Behav. 2013. 64 (2): 380–389.
Robbins T.W. The 5 choice serial reaction time task: behavioural pharmacology and functional neurochemistry. Psychopharmacology. 2002. 163 (3–4): 362–380.
Selby E.A., Anestis M.D., Bender T.W., Joiner T.E. Jr. An exploration of the emotional cascade model in borderline personality disorder. Journal of Abnormal Psychology. 2009. 118: 375–387.
Spear L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000. 24: 417–463.
Togashi H., Ueno K.-Ichi, Yamaguchi T., Matsumoto M., Higuchi K., Saito H., Yoshioka M. Anxiety-related behavior in juvenile stroke-prone spontaneously hypertensive rats – an animal model of attention-deficit/hyperactivity disorder. Biogenic Amines. 2004. 18 (3-6): 247–261.
Tomko R.L., Lane S.P., Pronove L.M., Treloar H.R., Brown W.C., Solhan M.B., Wood P.K., Trull T.J. Undifferentiated negative affect and impulsivity in borderline personality and depressive disorders: A momentary perspective. J. Abnorm. Psychol. 2015. 124 (3): 740–753.
Van den Bergh F. S., Bloemarts E., Chan J. S., Groenink L., Olivier B., Oosting R.S. Spontaneously hypertensive rats do not predict symptoms of attention-deficit hyperactivity disorder. Pharmacol. Biochem. Behav. 2006. 83: 380–390.
Winstanley C.A. The utility of rat models of impulsivity in developing pharmacotherapies for impulse control disorders Br. J. Pharmacol. 2011. 164 (4): 1301–1321.
Yu L.Q., Lee S., Katchmar N., Satterthwaite T.D., Kable J.W., Wolf D.H. Steeper discounting of delayed rewards in schizophrenia but not first-degree relatives. Psychiatry Res. 2017. 252: 303–309.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова