

Журнал высшей нервной деятельности им. И.П. Павлова, 2019, T. 69, № 4, стр. 505-513
ИЗУЧЕНИЕ ОСОБЕННОСТЕЙ ПРОЦЕССА СИМВОЛИЗАЦИИ У СЕРЫХ ВОРОН
М. В. Самулеева 1, *, А. А. Смирнова 1
1 МГУ им. М.В. Ломоносова, биологический факультет
Москва, Россия
* E-mail: samuleeva@gmail.com
Поступила в редакцию 10.01.2019
После доработки 28.01.2019
Принята к публикации 11.02.2019
Аннотация
Способность к символизации, т.е. к установлению эквивалентных отношений между знаком и его референтом, является одним из базовых компонентов языка. Эквивалентные отношения обладают свойствами симметричности, рефлексивности и транзитивности. Для оценки факторов, влияющих на понимание симметричности отношений между знаком и обозначаемым, мы обучили серых ворон выбору по условному соответствию образцу: если образцом было изображение буквы “S”, подкрепляли выбор изображения пары фигур одной формы и одинакового размера; а если буква “V” – выбор пары фигур разного размера. После обучения с одной парой стимулов для выбора две серые вороны не справились с тестом на понимание симметричности отношений, в котором образцы и стимулы для выбора поменяли местами. Второй тест на понимание симметричности отношений провели после завершения обучения с еще пятью парами стимулов, и с ним одна из двух ворон справилась. Результаты тестов на перенос правила выбора по условному соответствию образцу на новые стимулы показали, что обе вороны связали буквы “S” и “V” с понятиями “сходство” и “различие”. С проведенным далее третьим тестом на понимание симметричности отношений справились обе вороны. Полученные результаты свидетельствуют о том, что серые вороны способны установить симметричность отношений между знаком и референтом без дополнительного обучения и что на понимание симметричности отношений влияет тип референта.
Способность к символизации, т.е. к установлению эквивалентных отношений между знаком и его референтом (обозначаемым), является одним из необходимых когнитивных компонентов языка человека [Carr, Felce, 2000]. В языке референт (чаще всего – понятие) и его устное и письменное обозначения эквивалентны [Carr, Felce, 2000; Zentall et al., 2014]. Эквивалентными называют [Sidman et al., 1982; Sidman, Tailbi, 1982] отношения, обладающие тремя основными свойствами. Свойство симметричности означает, что перестановка членов отношения R не ведет к изменению типа отношения: xRy ® yRx. Свойство рефлексивности подразумевает, что каждый член отношения находится в таком же отношении R к самому себе: xRy ® (xRx и yRy). Наконец, свойство транзитивности состоит в том, что из отношения R между х и у, и между у и z следует такое же отношение R между х и z: (xRy и yRz) ® xRz).
Исследование механизмов формирования эквивалентных отношений у животных вносит вклад в изучение биологических предпосылок возникновения языка человека. Для изучения процесса формирования эквивалентных отношений субъекта обучают выбору по условному соответствию образцу: например, подкрепляют выбор стимула В по образцу А (далее А–В) и стимула D по образцу C (далее С–D). Затем в тесте оценивают возможность спонтанного появления новых отношений: между А в роли образца и А в роли стимула для выбора (отношение рефлексивности), между B в роли образца и А в роли стимула для выбора (отношение симметричности В–А) и между А и C (отношение транзитивности, если образец А; и отношения транзитивности и симметричности, если образец С). Испытуемые-люди, в отличие от животных, с подобными тестами, как правило, справляются [например, Quezada Velázquez, 2018]. В одной из первых работ [Sidman et al., 1982] было показано, что понимание симметричности отношений демонстрируют дети дошкольного возраста, но не макаки-резусы и павианы. Последующие эксперименты подтвердили этот вывод [Kojima, 1984; Biro, Matsuzawa, 2001; Prichard et al., 2015].
С другой стороны, накапливаются данные о том, что некоторые животные могут справиться с тестом на понимание симметричности, если им продемонстрировать возможность таких отношений. Этого можно добиться, например, путем обучения выбору по условному соответствию образцу не только с прямым (А-В), но и с обратным (В-А) порядком стимулов. Так, шимпанзе Аи после такого обучения с несколькими парами образцов и стимулов для выбора справилась с тестом на понимание симметричности отношений с новыми стимулами знакомых категории [Kojima, 1984; Biro D., Matsuzawa, 2001]. Продемонстрировать возможность симметричных отношений можно и другим способом – об-учением выбору по сходству с образцом (А-А, В-В), что также приучает животное к тому, что образец и стимулы для выбора могут меняться местами [Frank, Wasserman, 2005; Tomonaga et al., 1991; Schusterman, Kastak, 1993; Swisher, Urcuioli, 2015; Campos et al., 2014; Urcuioli, Swisher, 2015]. Так, с тестом на понимание симметричности отношений справилась шимпанзе Хлоя [Tomonaga et al., 1991], параллельно обученная как выбору по условному соответствию образцу (А–В) с двумя парами стимулов, так и выбору по сходству с образцом (А–А, В–В). Мы также ранее обнаружили, что с тестом на понимание симметричности отношений справились серая ворона и венесуэльский амазон, у которых предварительно было сформировано обобщенное правило выбора по сходству с образцом [Смирнова и др., 2013].
Еще одним фактором, влияющим на результат этого теста, может быть тип референта. В человеческом языке референтом знаков часто являются понятия. Гипотезу о влиянии типа референта на результат теста [Medam et al., 2016] поддерживают данные, полученные на сером попугае Алексе [Pepperberg, Gordon, 2005; Пепперберг, 2017, Pepperberg, 2015; 2018]. Алекс понимал [Пепперберг, 2017; Pepperberg, 2018] значение обращенных к нему высказываний (рецептивное использование знаков) и отвечал на них (продуктивное использование). При исследовании его способности оперировать понятием “число” [Pepperberg, Gordon, 2005] попугая обучили называть числительное (В), соответствующее числу предъявляемых объектов (А; А–В), после чего он справился с тестом на понимание симметричности отношений: без дополнительного обучения выбирал множество, соответствующее устному числительному (В–А).
Для того, чтобы оценить способность к формированию эквивалентых отношений у обезьян, не относящиеся к человекообразным, был предложен подход, оценивающий скорость обучения. Оказалось, что капуцины, обученные выбору по условному соответствию образцу (А1–В1 и А2–В2), быстрее обучаются выбору в пробах с обратным и симметричным порядком стимулов (В1–А1, В2–А2), чем в пробах с обратным, но “антисимметричным” порядком [В1–А2, В2–А1; Picanço, Barros, 2015; Soares Filho et al., 2016].
Целью данной работы был анализ факторов, влияющих на понимание симметричности отношений между знаком и обозначаемым у серых ворон без предшествующего экспериментального опыта.
МЕТОДИКА
Исследование проводили на серых воронах (Corvus cornix L.) разного возраста (старше двух лет), не имевших предшествующего экспериментального опыта. При проведении экспериментов руководствовались правилами выполнения работ с использованием экспериментальных животных согласно требованиям Декларации EC 2010. Птиц содержали в вольере на открытом воздухе. В ходе экспериментов в их ежедневном рационе при необходимости уменьшали долю кормов, содержащих белки животного происхождения.
На время эксперимента каждую птицу помещали в отдельную клетку (размером 70 × 35 × × 35 см), оборудованную присадой и поилкой с водой. Экспериментатор находился сбоку от клетки с птицей за непрозрачным экраном (70 × 40 см), который во время осуществления выбора не позволял птице и человеку видеть друг друга (см. рисунок 1). Таким образом, возможность неосознанного влияния экспериментатора на выбор была сведена до минимума.
Рис. 1.
Обстановка эксперимента. А – экспериментатор, В – непрозрачный экран, С – ворона.
Fig. 1. Experimental design. A – experimenter, B – opaque screen, С – crow.
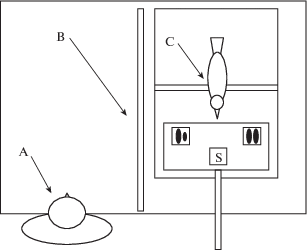
В клетку с птицей с помощью рукоятки длиной 30 см вдвигали “поднос” (пластину текстолита размером 20 × 30 см) с двумя кормушками (5.0 × 3.7 см) на его краях. Обе кормушки были накрыты ламинированными картонными карточками (70 × 70 мм) – стимулами для выбора. Между ними на середину подноса помещали карточку-образец. Подкрепление (две личинки мучного хрущака) помещали в кормушку, накрытую карточкой, которая условно (т.е. по произвольному решению экспериментатора) соответствовала образцу.
Подготовку подноса к предъявлению (в ходе которого двух личинок мучного хрущака помещали в одну из кормушек; накрывали обе кормушки стимулами для выбора и помещали между ними образец) проводили вне поля зрения вороны. Чтобы дать птице возможность рассмотреть предъявляемые ей стимулы, поднос на 3–5 с помещали снаружи от клетки так, чтобы ворона уже могла его видеть, но еще не могла дотянуться до карточек, накрывающих кормушки. После того, как птица осуществляла выбор и съедала подкрепление (или убеждалась в его отсутствии в выбранной кормушке), поднос извлекали из клетки, тем самым не давая птице возможности открыть вторую кормушку. Если в течение 2 мин птица не осуществляла выбор, поднос извлекали из клетки. Число проб в день зависело от уровня заинтересованности птиц (в среднем – 40).
При обучении выбору по условному соответствию образцу использовали два стимула-образца: на одном из них на белом фоне была изображена зеленая буква “S”, а на другом – буква “V”. На каждом из 12 возможных стимулов для выбора на белом фоне были изображены две черные фигуры одной формы одинакового или разного размера: круги, эллипсы, квадраты, прямоугольники, равносторонние и равнобедренные треугольники (суммарные площади двух фигур были выровнены). При обучении в одной пробе стимулами для выбора всегда служили изображения фигур одной формы: либо изображения кругов, либо эллипсов, либо квадратов и т.д. (далее мы их называем – пара стимулов). Если образцом было изображение буквы “S”, птицу подкрепляли за выбор фигур одинакового размера, а если образцом было изображение буквы “V”, то за выбор фигур разного размера. Обучение проводили до достижения критерия обученности – не менее 80% правильных выборов в 96 пробах подряд (p < < 0.0001).
В тестах, которые проводили после завершения обучения, собственно “тестовые пробы” (в которых использовали новые стимулы или меняли местами образцы и стимулы для выбора – см. в описании этапов эксперимента) чередовали с “фоновыми” (в которых использовали знакомые стимулы): каждая тестовая проба следовала после трех фоновых. В тестовых пробах личинок мучного хрущака помещали в обе кормушки. В фоновых пробах, как и при обучении, подкрепляли только “правильный” выбор.
Комбинации стимулов на каждом этапе эксперимента предъявляли в заранее организованной последовательности, в которой квазислучайно меняли как положение подкрепляемого стимула для выбора (в восьми последовательных пробах он четыре раза был слева и четыре раза справа, и при этом на одной стороне не больше двух раз подряд), так и тип образца (например, в четырех пробах из восьми это было изображение буквы “S”, а в других четырех – буквы “V”, при этом один и тот же образец использовали не более двух раз подряд).
Этапы эксперимента
1. Обучение двух серых ворон выбору по условному соответствию образцу: образцы – “S” и “V”; стимулы для выбора – шесть пар изображениями двух фигур одной формы одинакового или разного размера (1-й набор: круги, эллипсы, квадраты, прямоугольники, равносторонние или равнобедренные треугольники).
2. Обучение других двух серых ворон выбору по условному соответствию образцу с меньшим числом пар стимулов для выбора: образцы – “S” и “V”; стимулы для выбора – две пары из 1-го набора (круги и эллипсы). Здесь и на следующем этапе число стимулов для выбора уменьшили из-за отсутствия положительной динамики на предыдущем этапе.
3. Обучение этих же двух ворон выбору по условному соответствию образцу с единственной парой стимулов для выбора: образцы – “S” и “V”; стимулы для выбора – одна пара из 1-го набора (круги).
4. Первый тест на понимание симметричности отношений: в 48 тестовых пробах образцы – изображения кругов одинакового или разного размера (из 1-го набора), стимулы для выбора – “S” и “V”. В 144 фоновых пробах образцы – “S” и “V”; стимулы для выбора – одна пара из 1-го набора (круги).
5. Обучение выбору по условному соответствию образцу с каждой из оставшихся пяти пар стимулов из 1-го набора (образцы – “S” и “V”). Пару стимулов заменяли на следующую после достижения критерия обученности с предыдущей.
6. Продолжение обучения выбору по условному соответствию образцу при чередовании всех шести пар стимулов из 1-го набора (образцы – “S” и “V”).
7. Второй тест на понимание симметричности отношений: в 48 тестовых пробах образцы – 12 изображений фигур из 1-го набора; стимулы для выбора – “S” и “V”. В 144 фоновых пробах образцы – “S” и “V”; стимулы для выбора – все шесть пар стимулов из 1-го набора.
8. Тест на перенос правила выбора по условному соответствию образцу на новые стимулы, различающиеся по знакомому признаку (размеру фигур). И в фоновых, и в тестовых пробах образцы – “S” и “V”. В 24 тестовых пробах стимулы для выбора – шесть новых пар стимулов одной формы одинакового или разного размера (2-й набор: кресты, пятиугольники, восьмиугольники, полумесяцы, сердца и молнии). В 72 фоновых пробах стимулы для выбора – шесть знакомых пар стимулов (1-й набор).
9. Доучивание выбору по условному соответствию образцу со стимулами для выбора, использованными в тестовых пробах на предыдущем (восьмом) этапе: образцы – “S” и “V”; стимулы для выбора – шесть пар стимулов с изображениями двух фигур одной формы одинакового или разного размера (2-й набор).
10. Тест на перенос правила выбора по условному соответствию образцу на новые стимулы, различающиеся по новому признаку (форме фигур). И в фоновых, и в тестовых пробах образцы – “S” и “V”. В 24 тестовых пробах стимулы для выбора – 24 новые пары стимулов, составленные из 12 стимулов с изображениями двух фигур разного размера и одной формы из 1-го и 2-го наборов и 24 новых стимула с изображениями двух фигур разного размера и разной формы (например, треугольник и пятиугольник, крест и полумесяц). В 72 фоновых пробах стимулы для выбора – все 12 пар стимулов из 1-го и 2-го наборов.
11. Доучивание выбору по условному соответствию образцу со стимулами для выбора, использованными в тестовых пробах на предыдущем (десятом) этапе: образцы – “S” и “V”; стимулы для выбора – 24 пары (описаны выше).
12. Третий тест на понимание симметричности отношений: в 24 тестовых пробах образцы – 24 изображения фигур одинакового размера и одинаковой формы или разной формы (12 изображений фигур одинакового размера и одинаковой формы из 1-го и 2-го наборов и 12 новых изображений фигур одинакового размера и разной формы); стимулы для выбора – “S” и “V”. В 72 фоновых пробах образцы – “S” и “V”; стимулы для выбора – 12 пар (те же, что были использованы в тестовых пробах в качестве образцов).
Для статистического анализа данных использовали программу “STATISTICA for Windows” (версия 7): уровень достоверности правильных решений оценивали по биномиальному критерию (использовали “Binomial distribution probability calculator”); достоверность различий между долями правильных решений вычисляли с помощью метода определения ошибки разности между выборочными долями по критерию Стьюдента (“Difference between two proportions”; односторонний тест).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
1. При обучении двух серых ворон выбору по условному соответствию образцу со всеми шестью парами стимулов для выбора из 1-го набора положительной динамики обнаружено не было. С одной из этих двух птиц было выполнено 3600 проб, после чего она погибла. Со второй птицей обучение пришлось прервать после 1700 проб из-за усиления у нее невротических реакций.
2. При обучении двух других ворон (Веер и Шнобель) задачу упростили: использовали только две пары стимулов для выбора – круги и эллипсы. С каждой птицей было выполнено 2000 проб и положительной динамики обнаружено не было.
3. Далее этих же птиц обучали выбору по условному соответствию образцу с одной парой стимулов для выбора (кругами). При обучении наблюдалась положительная динамика; критерий обученности был достигнут обеими птицами за 2328 и 2248 проб соответственно.
4. В первом тесте на понимание симметричности отношений в тестовых пробах, в которых образцы и стимулы для выбора поменяли местами (B-A), доля правильных решений у обеих ворон не отличалась от случайного уровня: 45.8% (р = 0.68; n = 48) и 54.2% (p = 0.24; n = 48) соответственно. В фоновых пробах (A-B) птицы продолжали успешно решать ранее усвоенную задачу: 79.9% (p < 0.0001; n = 144) и 81.3% (p < 0.0001; n = 144).
5. При продолжении обучения выбору по условному соответствию образцу с каждой из оставшихся пяти пар стимулов из 1-го набора обе вороны достигали критерия обученности с каждой новой парой стимулов принципиально быстрее – за число проб, близкое к минимальным 96 (96–144; см. таблицу).
6. При продолжении обучения выбору по условному соответствию образцу при чередовании всех шести пар стимулов для выбора из 1-го набора вороны достигли критерия обученности за 96 и 184 пробы (таблица 1).
Таблица 1.
Число проб, потребовавшееся воронам для достижения критерия обученности, на разных этапах обучения выбору по условному соответствию образцу Table 1. Number of trials during each stage of matching-to-sample training
| Круги | Эллипсы | Квадраты | Прямоугольники | Треугольники равносторонние | Треугольники равнобедренные | Чередование всех 6 пар стимулов | |
|---|---|---|---|---|---|---|---|
| Веер | 2248 | 96 | 96 | 96 | 128 | 128 | 96 |
| Шнобель | 2328 | 96 | 144 | 144 | 144 | 144 | 184 |
7. Во втором тесте на понимание симметричности отношений в тестовых пробах (B-A), в которых образцами были 12 изображений фигур из 1-го набора, а стимулами для выбора – “S” и “V”, доля правильных решений у Веера превышала случайный уровень (64.6%, р = 0.02; n = 48), тогда как у Шнобеля – не отличалась от него (50.0%, р = 0.44; n = 48). В фоновых пробах обе птицы успешно решали ранее усвоенную задачу выбора по условному соответствию образцу: 84.7% (p < < 0.0001; n = 144) и 90.9% (p < 0.0001; n = 144). Доли правильных решений в тестовых и фоновых пробах у Веера достоверно отличались (p = 0.0015), т.е. он хуже справлялся с новой задачей в тестовых пробах (B–A), чем со знакомой задачей (A–B) в фоновых пробах.
8. В тесте на перенос правила выбора по условному соответствию образцу на новые стимулы, различающиеся по знакомому признаку (размеру фигур), доля правильных выборов в тестовых пробах у обеих ворон оказалась одинаковой и составила 79.2% (p = = 0.0008; n = 24). В фоновых пробах обе птицы продолжали успешно решать задачу выбора по условному соответствию образцу: 86.1% (p < 0.0001; n = 72) и 87.5% (p < 0.0001; n = 72). Доли правильных решений в тестовых и в фоновых пробах у обеих ворон не отличались (p = 0.2111 и p = 0.1604), т.е. птицы одинаково успешно решали данную задачу как со знакомыми, так и с новыми стимулами.
9. При доучивании выбору по условному соответствию образцу со стимулами для выбора, использованными в тестовых пробах на предыдущем (восьмом) этапе, каждая из ворон достигла критерия обученности за минимально возможное число проб (96). У обеих ворон доля правильных решений в первых 48 пробах (77.1% и 81.3%) не отличалась (p = 0.05 и p = 0.2949) от таковой в последних 48 пробах (89.6% и 85.4%).
10. В тесте на перенос правила выбора по условному соответствию образцу на новые стимулы, различающиеся по новому признаку (форме фигур), доля правильных выборов в тестовых пробах у двух ворон составила 66.7% (p = 0.03195; n = 24) и 75.0% (p = 0.0033; n = 24). В фоновых пробах вороны продолжали успешно решать задачу выбора по условному соответствию образцу со знакомыми стимулами: 88.9% (p < 0.0001; n = 72) и 79.2% (p < 0.0001; n = 72). Доли правильных решений в тестовых и в фоновых пробах достоверно отличались у первой вороны (p = 0.0067) и не отличались у второй (p = 0.3336). Таким образом, первая птица, в отличие от второй, хуже решала задачу выбора по условному соответствию образцу со стимулами, различающимися по новому признаку (форме фигур), чем со знакомыми.
11. При доучивании выбору по условному соответствию образцу со стимулами для выбора, использованными в тестовых пробах на предыдущем (десятом) этапе, каждая из ворон достигла критерия обученности за минимально возможное число проб (96). У обеих ворон доли правильных решений в первых 48 пробах (77.1% и 83.3%) не отличались (p = = 0.1488 и p = 0.3939) от таковой в последних 48 пробах (85.4% и 81.2%).
12. В третьем тесте на понимание симметричности отношений в тестовых пробах (B–A), в которых образцами были новые изображения фигур одинакового размера и одинаковой или разной формы, а стимулами для выбора – “S” и “V”, доли правильных решений у двух ворон составили 66.7% (p = = 0.03195, n = 24) и 79.2% (p = 0.00077, n = 24) соответственно. В фоновых пробах птицы продолжали успешно решать ранее усвоенную задачу выбора по условному соответствию образцу с этими же стимулами (A–B): 83.3% (p < 0.0001; n = 72) и 86.1% (p < 0.0001; n = 72). Доли правильных решений в тестовых и в фоновых пробах достоверно отличались у первой вороны (p = 0.0431), и не отличались у второй (p = 0.2111). Таким образом, первая птица, в отличие от второй, хуже справлялась с новой задачей в тестовых пробах (B–A), чем со знакомой задачей (A–B) в фоновых пробах.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Обучение ворон выбору по условному соответствию образцу при чередовании всех шести пар стимулов (как и при чередовании двух пар), оказалось неэффективным: ни у одной из птиц не наблюдалось положительной динамики даже после полутора тысяч проб. Эти данные согласуются с полученными нами ранее [Smirnova et al., 2000]: при обучении выбору по сходству с образцом с 12 вариантами стимулов для выбора ни одна из четырех ворон за 5000 проб так и не достигла критерия обученности. В данной работе положительная динамика стала заметной лишь при обучении птиц с одной парой стимулов для выбора (с кругами одинакового и разного размера). Скорость достижения критерия при этом оказалась примерно такой же, как при обучении ворон выбору по сходству с образцом (выбору черного стимула по черному образцу и белого стимула по белому образцу) [Smirnova et al., 2000]. Сходство скорости обучения может свидетельствовать об одинаковом механизме решения этих внешне различных задач на начальном этапе обучения – усвоении двух частных правил выбора “если …, то…” [Sidman, Tailby, 1982]. Столь длительное обучение выбору по образцу, по нашему мнению, связано со сложностью усвоения роли образца как знака, указывающего на подкрепляемый стимул для выбора.
С первым тестом на понимание симметричности отношений (В-А), проведенным после завершения обучения с кругами (А-В), ни одна из двух ворон не справилась. Таким образом, спонтанного понимания симметричности отношений между знаком и обозначаемым после обучения с одной парой стимулов у этих птиц, как и ожидалось, обнаружено не было. Сходный отрицательный результат – отсутствие спонтанного понимания отношений симметричности – был получен и в экспериментах с другими животными [например, Sidman et al., 1982; Kojima, 1984; Biro, Matsuzawa, 2001; Prichard et al., 2015] в отличие от экспериментов с людьми [Sidman et al., 1982; Quezada Velázquez, 2018].
Высокая скорость обучения ворон выбору по условному соответствию образцу с новыми парами стимулов из 1-го набора, вероятно, обусловлена тем, что на первых двух этапах обучения (2000 проб с кругами и эллипсами и примерно столько же с кругами) вороны долго усваивали и, наконец, усвоили особую роль образца как знака, указывающего на подкрепляемый стимул. В этом случае для успешного выбора по образцу с каждой новой парой стимулов им оставалось выучить два новых правила выбора типа “если…, то…” [Sidman M., Tailby, 1982]. Кроме того, быстрое достижения критерия обученности с новыми стимулами для выбора из 1-го набора может свидетельствовать о том, что параллельно с обучением выбору по условному соответствию шел процесс обобщения признаков изображенных фигур. Это могло привести к изменению типа референта (обозначаемого): вместо конкретных стимулов для выбора им могли становиться понятия “сходство” и “различие”.
Со вторым тестом на понимание симметричности отношений (В–А), который провели после завершения обучения со всеми стимулами из 1-го набора, одна из двух ворон справилась, хотя намного хуже, чем со знакомой задачей (А–В) в фоновых пробах. В этом тесте в качестве образцов использовали все 12 изображений из 1-го набора стимулов. Лишь два из них (круги одинакового и разного размера) вороны видели в роли образца ранее – в первом тесте на симметричность. Во всех тестах подкрепление было недифференцированным (его помещали в обе кормушки), что исключало возможность направленного обучения в ходе тестирования. Так, в первом тесте на понимание симметричности ворона получила подкрепление за выбор “правильного” стимула для выбора в 21 пробе и в 27 пробах – за выбор “неправильного”. Вероятность того, что такой опыт мог повлиять на результат второго теста, по нашему мнению, мала. Например, известно, что для того, чтобы обучить шимпанзе выбору по условному соответствию образцу с обратным порядком стимулов (B-A) после обучения выбору с прямым порядком стимулов (A–B), потребовалось более 2000 проб [Biro D., Matsuzawa, 2011]. Более вероятной причиной положительного результата второго теста на симметричность у одной из ворон, по нашему мнению, является изменение типа референта: к моменту проведения второго теста птица могла связать изображение букв “S” и “V” не с использованными при обучении конкретными стимулами, а с понятиями “сходство” и “различие”.
Для того, чтобы выяснить, с чем именно вороны связали буквы “S” и “V”, провели тесты на перенос правила выбора на новые стимулы. В первом из них в тестовых пробах использовали новые стимулы, различающиеся по знакомому признаку – размеру фигур. Во втором тесте новые стимулы различались по новому признаку – форме фигур. Он был принципиально сложнее еще и тем, что в тестовых пробах ранее подкрепляемые стимулы стали неподкрепляемыми, или наоборот. Например, если образец – “S”, то неподкрепляемым стимулом было новое изображение фигур разного размера и разной формы, а подкрепляемым – знакомое изображение фигур разного размера и одинаковой формы (в ходе обучения такой стимул был неподкрепляемым при образце “S”). Если образец – “V”, то неподкрепляемым стимулом было знакомое изображение фигур разного размера и одинаковой формы (в ходе обучения такой стимул был подкрепляемым при образце “V”), а подкрепляемым стимулом – новое изображение фигур разного размера и разной формы. Тот факт, что результаты обоих тестов оказались положительными, свидетельствует о том, что обе вороны действительно связали буквы “S” и “V” не только с конкретными знакомыми стимулами, но и с понятиями “сходство” и “различие”, применимыми не только к признаку размера, но и к форме фигур. Эти результаты подтверждают ранее полученные нами данные о способности ворон оперировать этими понятиями [Smirnova et al., 2000; Smirnova et al., 2015].
Проведенное после тестов на перенос правила выбора доучивание с новыми стимулами, различающимся по знакомому (размеру фигур) и новому (форме фигур) признакам, могло способствовать закреплению ранее сформированного обобщенного правила выбора по признакам сходства и отличия. Тот факт, что обе вороны одинаково успешно решали обе эти задачи как в первых, так и в последних 48 пробах, еще раз подтверждает, что у птиц было сформировано именно обобщенное правило выбора.
С третьим тестом на понимание симметричности отношений справились обе вороны. Его особенностью было то, что в тестовых пробах в качестве образца были применены новые стимулы – изображения фигур, одинаковых по размеру и одинаковых (например, два креста) или разных (например, круг и крест) по форме. Основной причиной отрицательного результата теста на симметричность у животных считают изменение места предъявления образца и стимула для выбора, поскольку место предъявления является важной характеристикой стимула [Frank, Wasserman, 2005; Swisher, Urcuioli, 2015; Urcuioli, Swisher, 2015]. Применение новых изображений могло способствовать положительному результату, поскольку они были меньше привязаны к конкретной роли (стимулов для выбора) и к месту предъявления, чем те, что были использованы в ходе обучения, а следовательно, десятки и сотни раз появлялись на месте и в роли стимулов для выбора. Кроме того, положительный результат третьего теста на симметричность отношений мог быть обусловлен типом референта, которым, как показали тесты на перенос правила выбора, к этому моменту стали понятия “сходство” и “различие”. Этот тип референта, вероятно, не столь привязан к месту предъявления, как конкретные изображения. Как было указано выше, один из немногочисленных примеров положительного результата в тесте на симметричность отношений ранее продемонстрировал серый попугай Алекс, у которого референтом знаков также были понятия [Pepperberg, Gordon, 2005; Pepperberg, 2018].
Еще одной возможной, хотя и маловероятной причиной положительного результата последнего теста на симметричность отношений у обеих ворон могло быть то, что он был для них уже третьим: несмотря на то, что во всех тестовых пробах подкрепляли любой выбор, им тем не менее демонстрировали, что образец и стимулы для выбора могут меняться местами. Чтобы исключить влияние этого фактора и подтвердить влияние типа референта на положительный результат теста на симметричность, необходимо обучить новых ворон обобщенному правилу выбора по условному соответствию образцу, убедиться, что референтами знаков стали понятия и затем провести единственный тест на понимание симметричности отношений.
ЗАКЛЮЧЕНИЕ
В целом полученные результаты свидетельствуют о том, что серые вороны способны справиться с тестом на понимание симметричности отношений между знаком и референтом без дополнительного обучения. Ранее нами было показано, что они способны к транзитивному заключению [Lazareva et al., 2004]. Тот факт, что они способны понимать симметричность и транзитивность отношений, может свидетельствовать о том, что в основе их способность к символизации, обнаруженной нами ранее [Смирнова, 2011], лежит формирование эквивалентных отношений между знаком и обозначаемым, а не только формирование условно рефлекторных ассоциаций.
Исследование выполнено при финансовой поддержке РФФИ (проект №16-04-01169), а также в рамках НИР Нейробиологические основы поведения животных АААА-А16-116021660055-1.
Список литературы
Пепперберг А. Алекс и я. М.: Языки славянских культур, 2017. 336 с.
Смирнова А.А. О способности птиц к символизации. Зоологический журнал. 2011. 90 (7): 803–810.
Смирнова А.А., Обозова Т.А., Самулеева М.В., Зорина З.А. Способность к символизации у птиц (врановые и попугаи): усвоение символов для обозначения признаков “сходство” и “различие”. Когнитивная наука в Москве: новые исследования. Тезисы конференции (19 июня 2013 г.). М.: БукиВеди, 2013. С. 1–5.
Biro D., Matsuzawa T. Use of numerical symbols by the chimpanzee (Pan troglodytes): Cardinals, ordinals, and the introduction of zero. Animal Cognition. 2001. 4: 193–199.
Campos H.C., Urcuioli P.J., Swisher M. Concurrent identity training is not necessary for associative symmetry in successive matching. Journal of the experimental analysis of behavior. 2014. 101 (1): 10–25.
Carr D., Felce D. Application of stimulus equivalence to language intervention for individuals with severe linguistic disabilities. Journal of Intellectual and Developmental Disability. 2000. 25 (3): 181–205.
Frank A.J., Wasserman E.A. Associative symmetry in the pigeon after successive matching-to-sample training. Journal of the Experimental Analysis of Behavior. 2005. 84 (2): 147–165.
Kojima T. Generalization Between Productive Use and Receptive Discrimination of Names in an Artificial Visual Language by a Chimpanzee. International Journal of Primatology. 1984. 5 (2): 161–182.
Lazareva O.F., Smirnova A.A., Bagozkaja M.S., Zorina Z.A., Rayevsky V.V., Wasserman E.A. Transitive responding in hooded crows requires linearly ordered stimuli. Journal of the experimental analysis of behavior. 2004. 82 (1): 1–19.
Medam T., Marzouki Y., Montant M., Fagot J. Categorization does not promote symmetry in Guinea baboons (Papio papio). Animal cognition. 2016. 19 (5): 987–998.
Pepperberg I.M. Numerical concepts: Grey parrot capacities. Mathematical Cognition and Learning. 2015. 1: 67–89.
Pepperberg I.M. Grey Parrots (Psittacus erithacus)–Cognitive and Communicative Abilities. Field and Laboratory Methods in Animal Cognition: A Comparative Guide. 2018. 329–347 pp.
Pepperberg I.M., Gordon J.D. Number comprehension by a grey parrot (Psittacus erithacus), including a zero-like concept //Journal of Comparative Psychology. 2005. V. 119. № 2. P. 197.
Picanço C.R.F., Barros R.S. Symmetry evaluation by comparing acquisition of conditional relations in successive (Go/No-Go) matching-to-sample training. The Psychological Record. 2015. 65 (1): 131–139.
Prichard A., Panoz Brown D., Bruce K., Galizio M. Emergent identity but not symmetry following successive olfactory discrimination training in rats. Journal of the experimental analysis of behavior. 2015. 10 4(2): 133–145.
Quezada Velázquez A.G., Padilla Vargas M.A., Flores Aguirre C.J. Equivalence class formation in 11-month-old pre-linguistic infants. Acta Colombiana de Psicología. 2018. 21 (1): 271–289.
Schusterman R.J., Kastak D.A. California sea lion (Zalophus californianus) is capable of forming equivalence relations. The Psychological Record. 1993. 43 (4): 823–839.
Sidman M., Rauzin R., Lazar R., Cunningham S., Tailby W., Carrigan P. A search for symmetry in the conditional discriminations of rhesus monkeys, baboons, and children. Journal of the Experimental Analysis of Behavior. 1982. 37 (1): 23–44.
Sidman M., Tailby W. Conditional discrimination vs. matching to sample: an expansion of the testing paradigm. Journal of the Experimental Analysis of Behavior. 1982. 37 (1): 5–22.
Soares Filho P.S., Silva Á.J., Velasco S.M., Barros R.S., Tomanari G.Y. Assessing symmetry by comparing the acquisition of symmetric and nonsymmetric conditional relations in a Capuchin Monkey. International Journal of Psychological Research. 2016. 9 (2): 30–39.
Swisher M., Urcuioli P. J. Symmetry in the pigeon with sample and comparison stimuli in different locations. II. Journal of the experimental analysis of behavior. 2015. 104 (2): 119–132.
Smirnova A.A., Lazareva O.F., Zorina Z.A. Use of number by crows: investigation by matching and oddity learning. J. Exp. Anal. Behav. 2000. 73: 163–176.
Smirnova A., Zorina Z., Obozova T., Wasserman E. Crows spontaneously exhibit analogical reasoning. Current Biology. 2015. 25 (2): 256–260.
Tomonaga M., Matsuzawa T., Fujita K., Yamamoto J.H. Emergence of symmetry in a visual conditional discrimination by chimpanzees (Pan troglodytes). Psychological Reports. 1991. 68 (1): 51–60.
Urcuioli P.J., Swisher M.J. Transitive and anti-transitive emergent relations in pigeons: Support for a theory of stimulus-class formation. Behavioural processes. 2015. 112: 49–60.
Zentall T.R., Wasserman E.A., Urcuioli P.J. Associative concept learning in animals. Journal of the Experimental Analysis of Behavior. 2014. 101 (1): 130–151.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова