

Доклады Российской академии наук. Науки о жизни, 2023, T. 513, № 1, стр. 528-532
Mustela strandi (mustelidae, carnivora) из раннего плейстоцена Крыма
Д. О. Гимранов 1, *, Ц. Цзянцзуо 2, 3, А. В. Лавров 4, академик РАН А. В. Лопатин 4
1 Институт экологии растений и животных
Уральского отделения Российской академии наук
Екатеринбург, Россия
2 Ключевая лаборатория орогенных поясов
и эволюции земной коры, Школа наук о Земле и космосе, Пекинский университет
Пекин, Китай
3 Ключевая лаборатория эволюции позвоночных
и происхождения человека Китайской академии наук, Институт палеонтологии позвоночных
и палеоантропологии Китайской академии наук
Пекин, Китай
4 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: djulfa250@rambler.ru
Поступила в редакцию 20.06.2023
После доработки 10.07.2023
Принята к публикации 12.07.2023
- EDN: GTZZKA
- DOI: 10.31857/S2686738923600449
Аннотация
Описана нижнечелюстная кость Mustela strandi Kormos, 1934 из нижнеплейстоценовых отложений (поздний виллафранк, 1.8–1.5 млн л.н.) пещеры Таврида в Крыму. Это первая находка в России M. strandi – вымершего вида куньих, известного по редким остаткам в нижнем и среднем плейстоцене Центральной Европы.
Мелкие куньи рода Mustela появились в палеонтологической летописи в раннем плиоцене, причем их находки на протяжении всего плиоцена очень скудны. Самые древние виды Mustela plioerminea Stach, 1959 и M. pliocaenica Stach, 1959 описаны из местонахождений Венже 1 (Польша, 3.6–3.2 млн л.н.) и Вельферсхайм (Германия, 3.6–3.5 млн л.н.) [1, 2]. В плиоцене в пределах рода произошла дивергенция двух крупных линий – горностаев и ласок, что подтверждается молекулярно-генетическими данными [3].
В первой половине плейстоцена представители рода Mustela стали более многочисленными и распространились в Азию. Пика своего видового разнообразия род достиг, по-видимому, в позднем плейстоцене и голоцене, в современной фауне он представлен 17–18 видами [4].
Из нижнего плейстоцена Европы описаны Mustela palerminea (Petenyi, 1864), M. praenivalis Kormos, 1934, M. strandi Kormos, 1934, M. putorius stromeri Kormos, 1934, M. cf. eversmanii (Lesson, 1827) и M. nivalis Linnaeus, 1766 [5–8]. В Азии отмечены M. cf. sibirica Pallas, 1773 [9] и Mustela sp. [10]. Самыми распространенными и многочисленными видами рода в раннем плейстоцене были M. palerminea и M. praenivalis, остатки которых найдены во многих европейских местонахождениях [11–13]. Недавно M. palerminea была обнаружена нами в Крыму [14].
Намного хуже известен весьма редкий европейский вид M. strandi. Впервые этот вид был обнаружен в Венгрии в местонахождении Брассо (средний плейстоцен), откуда он был описан как Putorius (Arctogale) sp. [15]. Впоследствии данная форма была выделена в самостоятельный вид M. strandi Kormos, 1934 [5]. Позднее M. strandi была определена еще из пяти центральноевропейских местонахождений. В раннем плейстоцене M. strandi обитала на территории Германии (Шернфельд, 1.9–1.6 млн л.н.) и Польши (пещера Жабья, 1.7–1.5 млн л.н.) [13]. На границе раннего и среднего плейстоцена остатки M. strandi обнаружены в этих же странах (пещера Сакдиллинг, Германия, 0.9–0.7 млн л.н.; Кози Гжбет, Польша, 0.8–0.7 млн л.н.) [13]. Серединой среднего плейстоцена датирована находка M. strandi в пещере Полудневой (Południowa Cave, Kitzelhöhle) на юго-западе Польши [16].
В настоящей статье нами рассматривается находка M. strandi в нижнем плейстоцене пещеры Таврида в Крыму (Белогорский район, п. Зуя). По составу фауны позвоночных основной костеносный слой пещеры Таврида датируется ранним плейстоценом (поздний виллафранк, псекупский фаунистический комплекс, около 1.8–1.5 млн л.н.) [17]. Среди млекопитающих отсюда определены многочисленные крупные хищные семейств Ursidae, Canidae, Felidae и Hyaenidae и редкие мелкие Mustelidae [14].
Нами был дополнительно изучен голотип M. strandi, хранящийся в Геологическом музее Будапешта (Венгрия). Для сравнения использовались коллекции современных M. erminea Linnaeus, 1758, M. nivalis Linnaeus, 1766, M. eversmanii (Lesson, 1827), M. putorius Linnaeus, 1758, M. sibirica Pallas, 1773, M. altaica Pallas, 1811, M. lutreola Linnaeus, 1761 и Neogale vison (Schreber, 1777), хранящиеся в Институте экологии растений и животных УрО РАН (ИЭРЖ, Екатеринбург), Зоологическом музее Московского государственного университета им. М.В. Ломоносова и Зоологическом институте РАН (Санкт-Петербург).
Описываемый материал из пещеры Таврида представлен левой нижнечелюстной костью (экз. ИЭРЖ, № 727/2263, сборы 2021 г.). Измерения проводились штангенциркулем с точностью до 0.01 мм. Визуализация данных была выполнена в программе R 4.1 [18] с помощью пакета ggplot2 [19].
Левая ветвь нижней челюсти (экз. ИЭРЖ, № 727/2263) содержит нестертые p4–m2; резцовая часть челюсти, клык и симфиз утрачены, челюсть обломана на уровне р3. Вершина венечного отростка отсутствует, сочленовный и угловой отростки большей частью утрачены (рис. 1). Челюсть довольно массивная, массетерная впадина (fossa masseterica) овальная, умеренно глубокая, ее передний край заходит за уровень заднего края m1. Задний край венечного отростка (proc. coronoideus) субвертикальный. Основание углового отростка (proc. angularis) массивное. Сочленовный отросток (proc. articularis) расположен чуть ниже уровня альвеолярного края нижней челюсти. Край области крепления медиальной порции височной мышцы (musculus temporalis pars medialis) на поверхности венечного отростка хорошо выражен в рельефе кости. Его передняя часть находится вентральнее заднего края m2, а вентральная часть располагается на уровне середины сочленовного отростка. Рельефность и низкое положение края области крепления на челюсти указывают на специализированное мощное развитие височной мышцы.
Рис. 1.
Mustela strandi Kormos, 1934, экз. ИЭРЖ, № 727/2263, левая нижнечелюстная кость с p4–m2: а – с буккальной стороны, б – с окклюзиальной стороны, в – с лингвальной стороны (с прорисовкой зеркально); Крым, пещера Таврида; нижний плейстоцен. Обозначения: Hycd – гипоконид, Pacd – параконид, Prcd – протоконид.
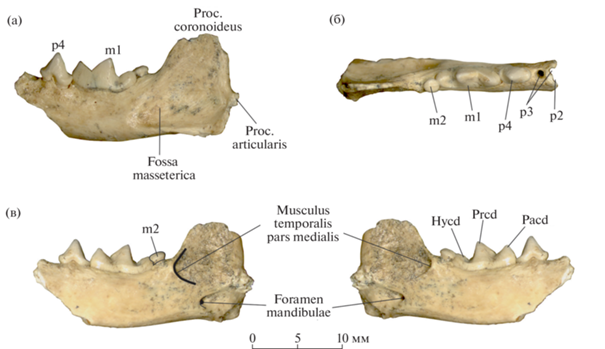
От р2 сохранилась только задняя часть альвеолы, поэтому неясно, был ли он однокорневым или двухкорневым. Судя по альвеолам, р3 – двухкорневой, крупнее р2, но меньше, чем р4. Задняя альвеола р3 немного больше передней.
Последний премоляр р4 – довольно крупный, массивный зуб. Коронка слегка асимметричная, слабо расширятся в задней части. Главный бугорок конический, с развитым дистальным гребнем. Выраженный передний цингулид отсутствует, задний цингулид без бугорков. Задний конец коронки р4 слегка перекрывает буккально параконид m1. Вершина главного бугорка р4 выше параконида и протоконида m1 (рис. 1а). Поверхность эмали р4 и m1 гладкая, без выраженной струйчатости.
Коронка хищнического зуба m1 крупная, продольно вытянутая, без выступов в середине и сужения в области передней части талонида. Лингвальный цингулид почти полный, прерывается только на небольшом участке в средней части, буккальный цингулид не развит. Параконид m1 ниже протоконида (рис. 1а), хищническая вырезка глубокая и широкая, метаконидный гребень отсутствует. Талонид режущего типа – без талонидного бассейна. Талонид удлиненный, почти равен по длине паракониду. Гипоконид увеличен, имеет вид режущего гребня и расположен на продольной оси зуба. Задний цингулидный бугорок на талониде отсутствует. Дополнительный корень между передним и задними корнями не развит.
Последний моляр m2 редуцированный, но довольно крупный, смещен лингвальнее продольной оси m1 (рис. 1б). Окклюзиальная поверхность m2 расположена выше уровня талонида m1 (рис. 1в). Коронка m2 имеет форму слабо удлиненного овала. На уплощенной окклюзиальной поверхности имеется невысокий узкий гребень, вытянутый продольно.
Размеры нижнечелюстной кости и зубов экз. ИЭРЖ, № 727/2263 приведены в табл. 1.
Таблица 1.
Размеры (в мм) нижнечелюстной кости Mustela strandi Kormos, 1934 из плейстоцена Европы (n – число экземпляров)
| Промеры | пещера Таврида, Крым, Россия (экз. ИЭРЖ, № 727/2263) | Кози Гжбет*, Польша | Брассо, Венгрия | ||
|---|---|---|---|---|---|
| пределы | среднее | n | |||
| Длина основания венечного отростка | 9.03 | – | – | – | – |
| Высота за p4 | 5.56 | – | – | – | – |
| Высота за m1 | 6.51 | 5.5–6.3 | 5.7 | 3 | – |
| Толщина под p4 | 3.0 | – | – | – | – |
| Толщина под m1 | 2.96 | 2.8–3.4 | 3.0 | 3 | – |
| Длина p3–m2 | 14.47 | – | – | – | – |
| Длина p4–m1 | 10.27 | – | – | – | – |
| Длина p4–m2 | 11.36 | – | – | – | – |
| Длина m1–m2 | 7.98 | – | – | – | – |
| Длина p4 | 3.62 | 3.3–3.7 | 3.5 | 4 | 3.4 |
| Ширина p4 | 1.97 | 1.4–1.9 | 1.6 | 5 | 1.8 |
| Длина m1 | 6.8 | 6.2–6.7 | 6.5 | 4 | 6.65 |
| Длина тригонида m1 | 5.18 | – | – | – | – |
| Ширина тригонида m1 | 2.48 | 2.2–2.7 | 2.4 | 4 | 2.35 |
| Ширина талонида m1 | 2.07 | 1.5–1.8 | 1.7 | 4 | 1.6 |
| Длина m2 | 1.55 | 1.6–2.0 | 1.8 | 4 | 1.5 |
| Ширина m2 | 1.45 | 1.5–1.9 | 1.7 | 4 | 1.4 |
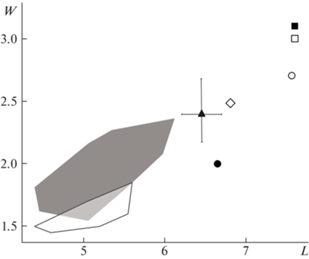
По длине и ширине хищнический зуб m1 экз. ИЭРЖ, № 727/2263 близок к таковым M. strandi из среднеплейстоценового местонахождения Брассо в Венгрии и раннеплейстоценового (конец раннего плейстоцена) местонахождения Кози Гжбет в Польше (рис. 2). Следует отметить, что экз. ИЭРЖ, № 727/2263 по размерам (высота тела нижней челюсти за m1, длина и ширина р4, длина m1, ширина талонида m1) превосходит образцы из местонахождений Кози Гжбет и Брассо (табл. 1). На графике (рис. 2) видно, что M. palerminea имеет существенно меньшие размеры m1, тогда как M. cf. sibirica из Китая, M. putorius stromeri и M. cf. eversmanii из Европы по размерам m1 крупнее, чем M. strandi.
Рис. 2.
Размеры m1 представителей Mustela из плейстоцена Евразии. Обозначения: L – длина m1, W – наибольшая ширина m1; черный квадрат – M. putorius stromeri Kormos, 1934 [5], белый квадрат – M. cf. eversmanii (Lesson, 1827) [7], белый круг – M. cf. sibirica Pallas, 1773 [9]; M. strandi Kormos, 1934: черный треугольник (с пределами изменчивости) [6], черный круг [5], белый ромб – экз. ИЭРЖ, № 727/2263; M. palerminea (Petenyi, 1864): серая область [13], белая область [5].
От современных среднеразмерных представителей рода Mustela, населяющих Северную Евразию (M. erminea и M. altaica), и от ископаемой M. palerminea экз. ИЭРЖ, № 727/2263 отличается более крупными размерами. Также описываемый экземпляр от упомянутых видов отличают следующие морфологические признаки: (1) рельефный край области прикрепления височной мышцы спереди не заходит за задний край m2 и вентрально достигает уровня сочленовного отростка; (2) сочленовный отросток расположен ниже уровня альвеолярного края нижней челюсти; (3) массетерная впадина протягивается вперед за уровень заднего края m1; (4) р4 крупный, массивный; (5) передний цингулид на р4 почти полностью отсутствует; (6) главный бугорок р4 имеет развитый дистальный гребень; (7) вершина главного бугорка р4 выше вершин параконида и протоконида m1; (8) эмаль р4 и m1 без струйчатости; (9) m2 смещен лингвально относительно оси m1 (в окклюзиальной проекции); (10) m2 довольно крупный относительно размеров m1. Указанные номера признаков (с дополнениями) ниже использованы для сравнения с другими видами.
От M. sibirica экз. ИЭРЖ, № 727/2263 отличается признаками 1, 2 и 8, а также (11) отсутствием лингвального бассейна на талониде m1, (12) отсутствием дополнительного корня в средней части m1, (13) отсутствием расширения (вздутия) m1 в средней части коронки с лингвальной стороны.
От M. putorius и M. eversmanii отличается признаками 1, 2, 6, 8, 12, а также (14) почти полным лингвальным цингулидом, прерывающимся в средней части m1, (15) отсутствием гипоконулида на m1, (16) лингвальным смещением m2 относительно m1.
От M. lutreola отличается признаками 1, 2, 8, 11, 13, 14, а также (17) талонидом m1 режущего типа, сформированным из гипоконида.
От Neogale vison отличается признаками 8, 13, 14, 16, 17, а также (18) отсутствием бугорков на заднем цингулиде р4 и (19) отсутствием буккального цингулида на m1.
Таким образом, отмеченные на экз. ИЭРЖ, № 727/2263 признаки 1 и 2 (рельеф и положение края области крепления медиальной порции височной мышцы и расположение сочленовного отростка) не встречаются у других видов рода Mustela. Т. Кормош [5] ранее указывал на захождение рельефного края области крепления височной мышцы за уровень заднего края m2 как видоспецифичный признак M. strandi.
Т. Вишневская [6] сближала M. strandi с современным колонком M. sibirica. Мы тоже отмечаем, что эти виды разделены наименьшим числом признаков по сравнению с другими современными представителями рода Mustela.
Морфологические признаки нижнечелюстной кости и зубов экз. ИЭРЖ, № 727/2263 сходны с описанными ранее для M. strandi [5, 6, 13, 15]. Размерные характеристики экз. ИЭРЖ, № 727/2263 сближают его с находками из Брассо и Кози Гжбет. Однако при этом форма из пещеры Таврида выделяется особо крупными размерами нижней челюсти, р4 и m1 при наличии сравнительно небольшого m2. Можно предположить, что в первой половине плейстоцена у M. strandi стала проявляться тенденция к уменьшению размеров; это позволяет объяснить относительно мелкие размеры более поздних представителей вида в Европе.
Список литературы
Stach J. On some Mustelinae from the Pliocene bone breccias of Węże // Acta Palaeontologica Polonica. 1959. V. 4. № 2. P. 101–118.
Morlo M., Kundrát M. The first carnivoran fauna from the Ruscinium (Early Pliocene, MN 15) of Germany // Paläontologische Zeitschrift. 2001. V. 75. P. 163–187.
Sato J.J., Wolsan M., Prevosti F.J. et al. Evolutionary and biogeographic history of weasel-like carnivorans (Musteloidea) // Molecular Phylogenetic and Evolution. 2012. V. 63. № 3. P. 745–757.
Wilson D.E., Reeder D.M., eds. Mammal species of the World: a taxonomic and geographic reference. Baltimore: Johns Hopkins University Press, 2005.
Kormos T. Neue und wenig bekannte Musteliden aus dem ungarischen Oberpliozän // Folia Zoologica et Hydrobiologica. 1934. V. 5. P. 129–158.
Wiszniowska T. Middle Pleistocene Carnivora (Mammalia) from Kozi Grzbiet in the Swietokrzyskie Mts, Poland // Acta Zoologica Cracoviensia. 1989. V. 32. № 14. P. 589–630.
Thenius E. Die Carnivoren-Reste aus dem Altpleistozän von Voigtstedt bei Sangerhausen in Thüringen // Paläontologische Abhandlungen. Abt. A. 1965. V. 2. № 2–3. P. 537–564.
Thabard C., Fourvel J.-B. Les restes de Mustela nivalis Linnaeus, 1766 (Carnivora, Mustelidae) du gisement Pléistocène inférieur de Montoussé 5 (Hautes-Pyrénées, France) // Comptes Rendus Palevol. 2022. Art. 21 (28). P. 595–618.
Pei W. On the Carnivora from Locality 1 of Choukutien // Palaeontologia Sinica. Ser. C. 1934. V. 8. P. 1–217.
Мотузко А.Н. Фауна млекопитающих верхнего плиоцена, нижнего и начала среднего плейстоцена внеледниковой области Западной Сибири и ее палеогеографическое значение // Автореферат на соиск. уч. степени канд. географ. наук. Москва: МГУ, 1971. С. 1–16.
Rabeder G. Die Carnivoren (Mammalia) aus dem Altpleistozän von Deutsch-Altenburg 2. Mit Beiträgen zur Systematik einiger Musteliden und Caniden // Beiträge zur Paläontologie von Österreich. 1976. V. 1. P. 5–119.
Palombo M.R., Sardella R., Novelli M. Carnivora dispersal in Western Mediterranean during the last 2.6 Ma // Quaternary International. 2008. V. 179. № 1. P. 176–189.
Marciszak A., Ambros D., Hilpert B. Mustelids from Sackdilling Cave (Bavaria, Germany) and their biostratigraphic significance // Geobios. 2021. V. 68. P. 83–107.
Гимранов Д.О., Цзянцзуо Ц., Лавров А.В., Лопатин А.В. Mustela palerminea (Mustelidae, Carnivora) из раннего плейстоцена Крыма // Доклады Российской академии наук. Науки о жизни. 2023. Т. 511. С. 375–380.
Éhik J. Die präglaciale Fauna von Brasso (Vorläufiger Bericht) // Földtany Közlony. 1913. V. 43. P. 23–26.
Marciszak A., Matyaszczyk L. Południowa Cave – first mid-Middle Pleistocene site from Sudety Mts (Silesia, SW Poland) and their biostratigraphic significance // Proceedings of INQUA SEQS 2021 Conference. Wrocław, Poland: University of Wrocław and Polish Geological Society, 2021. P. 64.
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
R Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, 2020. https://www.R-project.org.
Wickham H. ggplot2: elegant graphics for data analysis. New York: Springer-Verlag, 2016.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни