

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 4, стр. 53-70
Pseudotorellia Florin из верхнеюрских–нижнемеловых отложений Буреинского бассейна, Дальний Восток России
Н. В. Носова 1, *, Е. И. Костина 2, Е. В. Бугдаева 3
1 Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
2 Геологический институт РАН
Москва, Россия
3 Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
Владивосток, Россия
* E-mail: natanosova@gmail.com
Поступила в редакцию 04.12.2020
После доработки 25.12.2020
Принята к публикации 31.12.2020
Аннотация
Проведена ревизия Pseudotorellia из верхнеюрских–нижнемеловых отложений Буреинского бассейна (Дальний Восток России). Выявлено сходство морфологических и эпидермальных признаков листьев Pseudotorellia angustifolia Doludenko и P. longifolia Doludenko, на основе чего предложено считать вид P. longifolia синонимом P. angustifolia. Листья этого рода из Буреинского бассейна, описанные ранее по морфологическим и эпидермальным признакам как P. ensiformis (Heer) Doludenko, выделены в новый вид P. doludenkoae sp. nov., поскольку типовые экземпляры P. ensiformis из Иркутского бассейна не имеют сохранившейся кутикулы и их эпидермальное строение неизвестно. Впервые изучено эпидермальное строение листьев, описанных ранее как Pseudotorellia pulchella и P. crassifolia только по морфологическим признакам. Обнаружено сходство их морфологических и эпидермальных признаков, на основе чего они объединены в один вид P. crassifolia, диагноз которого уточнен. Показано, что эпидермальные признаки Pseudotorellia позволяют уверенно различать виды этого рода даже по фрагментарным остаткам и дисперсным кутикулам. Это указывает на большой стратиграфический потенциал видов Pseudotorellia для внутрибассейновой и межрегиональной стратиграфии континентальных толщ, особенно при изучении кернового материала и углей, где обычно сохранность растительных остатков не позволяет судить об их морфологии. Ревизия всех находок Pseudotorellia angustifolia позволила предположить возможное место и время появления вида и его дальнейшее распространение в пространстве и во времени. По-видимому, этот вид появился в раннеюрскую эпоху на территории современной Восточной Сибири (Канский бассейн), в средней юре его ареал расширился как на северо-запад (Томская область и Ямало-Ненецкий автономный округ), где вид сохранился местами до поздней юры, так и на восток – до Иркутской области и Хабаровского края (Буреинский бассейн), где он сохранился до раннего мела.
ВВЕДЕНИЕ
Род Pseudotorellia Florin был выделен для линейных листьев из нижнемеловых отложений Шпицбергена, имеющих несколько параллельных жилок и своеобразное строение эпидермиса (Florin, 1936). Диагноз рода неоднократно уточнялся (Watson, 1969; Bose, Manum, 1990; Киричкова, Носова, 2009). Листья Pseudotorellia от линейных до ланцетовидных или обратнояйцевидных, верхушка тупая, округлая или заостренная, цельная или мелкозубчатая. В основание листа входят две жилки, дихотомирующие около основания и параллельные в средней части листа. Для большинства видов Pseudotorellia характерно расположение устьиц только на нижней поверхности листа (гипостоматность). Единственный вид с амфистоматными листьями – P. amphistomatica Nosova et Golovneva – был описан из альба Западной Сибири (Головнева, Носова, 2012). Устьица ориентированы продольно, реже косо, могут располагаться как беспорядочно на всей поверхности листа, так и образовывать прерывистые ряды и полосы. Замыкающие клетки устьиц погруженные, крыловидной формы, часто с длинными полярными выростами.
Изучая гинкговые и чекановскиевые из мезозойских отложений Буреинского бассейна, В.А. Красилов (1972) описал укороченные побеги с прикрепленными к ним листьями Pseudotorellia. В ассоциации с ними найдены женские репродуктивные структуры, выделенные Красиловым в род Umaltolepis Krassilov. На основе сходства эпидермальных признаков он отнес Pseudotorellia и Umaltolepis к новому семейству Pseudotorelliaceae в составе порядка Ginkgoales (Красилов, 1972). Ф.А. Станиславский (1973), а вслед за ним и Ч. Чжоу (Zhou, 1991) для растений Pseudotorellia–Umaltolepis использовали название семейства Umaltolepidaceae. Позднее совместные находки Pseudotorellia и Umaltolepis были описаны из нижней юры северного Ирана (Schweitzer, Kirchner, 1995), средней юры Центрального Китая (Dong et al., 2019), Узбекистана (Nosova, 2020) и Восточной Сибири, Россия (Nosova, 2021), а также из нижнего мела Северо-Востока Китая (Wang, 1984; Chen et al., 1988) и Центральной Монголии (Herrera et al., 2017).
Ф. Эррера с соавторами (Herrera et al., 2017), детально изучив репродуктивные структуры из апт-альбской формации Тевшиин Гови Монголии, а также проведя ревизию типового материала из юры Буреинского бассейна, пришли к выводу, что Umaltolepis существенно отличается от женских репродуктивных структур гинкговых и обнаруживает значительное сходство с фруктификациями Vladimaria Gordenko (2010) из средней юры европейской части России. Они относят Umaltolepis к предложенному Н.В. Горденко порядку Vladimariales (Gordenko, 2010). В статье, посвященной Umaltolepis из среднеюрских отложений Центрального Китая, Ч. Дун с соавторами рассматривают семейство Umaltolepidaceae в составе порядка Ginkgoales sensu lato (Dong et al., 2019). Однако не все виды Pseudotorellia можно с уверенностью относить к семейству Umaltolepidaceae. Например, в статье о женских репродуктивных структурах Umkomasia Thomas из нижнего мела Монголии Г. Ши с соавторами (Shi et al., 2019) приводят реконструкцию растения с листьями Pseudotorellia palustris Shi, Herrera, Herendeen, Leslie, Ichinnorov, Takahashi et Crane и женскими шишками Umkomasia mongolica Shi, Leslie, Herendeen, Herrera, Ichinnorov, Takahashi, Knopf et Crane и относят листья Pseudotorellia к семейству Umkomasiaceae порядка Umkomasiales.
Впервые остатки листьев Pseudotorellia из верхнеюрских–нижнемеловых отложений Буреинского бассейна (Хабаровский край, Дальний Восток России) были изучены М.П. Долуденко (Вахрамеев, Долуденко, 1961). Всего было выделено пять видов: Pseudotorellia angustifolia Doludenko, P. ensiformis (Heer) Doludenko, P. longifolia Doludenko, P. pulchella (Heer) Vassilevskaja и P. crassifolia (Prynada) Doludenko, из которых два последних – только на основе морфологических признаков.
О. Геер (Heer, 1876) описал изолированные ланцетовидные листья из юрского местонахождения Усть-Балей (Иркутский бассейн, Восточная Сибирь) как Podozamites ensiformis, фитолейма которых не сохранилась. Долуденко предложила комбинацию Pseudotorellia ensiformis на основе изученного ею морфологического и эпидермального строения листьев из Буреинского бассейна (Вахрамеев, Долуденко, 1961), включив в состав данного таксона и Podozamites ensiformis. Однако позже Долуденко и Е.С. Рассказова отметили, что отождествление буреинских листьев с иркутскими было неверным, поскольку эпидермальные признаки последних неизвестны (Долуденко, Рассказова, 1972, с. 21). Данные авторы предложили относить к виду Pseudotorellia ensiformis листья с неизвестными эпидермальными признаками, но имеющие такие же морфологические характеристики, как и выбранный ими лектотип из Усть-Балея. Красилов (1972) привел свое описание и расширенный диагноз вида P. angustifolia, считая P. ensiformis синонимом P. angustifolia. Авторы настоящей работы провели ревизию образцов Pseudotorellia из коллекций В.А. Вахрамеева и Красилова и пришли к выводу, что описанные Долуденко P. angustifolia и P. ensiformis существенно отличаются друг от друга как по морфологическим, так и по эпидермальным признакам. Поскольку эпидермальные признаки типовых экземпляров P. ensiformis из Усть-Балея неизвестны, выделен новый вид P. doludenkoae sp. nov. для листьев из Буреинского бассейна.
Детальное изучение и сравнение эпидермального строения буреинских листьев Pseudotorellia angustifolia и P. longifolia не обнаружило существенных различий между этими видами, поэтому они объединены в один вид, а видовое название P. longifolia принято за синоним P. angustifolia (см. описание и замечания ниже).
Комбинация Pseudotorellia crassifolia была предложена Долуденко только по морфологическим признакам (Вахрамеев, Долуденко, 1961). В настоящей работе изучено эпидермальное строение листьев этого вида с голотипа, на основе чего приведен уточненный видовой диагноз и расширенное описание.
Еще один вид из Буреинского бассейна, Pseudotorellia pulchella, был также выделен только по морфологическим признакам (Вахрамеев, Долуденко, 1961). Листья с такими признаками впервые были описаны Геером из нижнемеловых отложений Шпицбергена как Podozamites pulchellus Heer, 1876. Н.Д. Василевская (1959) на основе материала из нижнего мела Ленского бассейна (Восточная Сибирь) предложила комбинацию Pseudotorellia pulchella. Эпидермальное строение листьев как со Шпицбергена, так и из Ленского бассейна неизвестно. Изучив эпидермальное строение буреинских листьев P. pulchella, авторы настоящей работы пришли к выводу, что их морфологические и эпидермальные признаки сходны с таковыми у P. crassifolia. Поскольку, в отличие от P. pulchella, типовой материал P. crassifolia происходит из Буреинского бассейна, буреинские листья, описанные как P. pulchella, рассматриваются в составе вида P. crassifolia (см. описание ниже). К P. pulchella предлагается относить листья, эпидермальное строение которых неизвестно, а морфологические признаки сходны с таковыми у типовых экземпляров со Шпицбергена.
Авторы настоящей работы поддерживают точку зрения Эррера с соавторами (Herrera et al., 2017) и описывают листья Pseudotorellia из Буреинского бассейна в составе порядка Vladimariales, поскольку здесь они были найдены в ассоциации с фруктификациями Umaltolepis (Красилов, 1972).
МАТЕРИАЛ И МЕТОДЫ
Проведена ревизия образцов с остатками листьев Pseudotorellia из коллекции, собранной в 1958 г. В.А. Вахрамеевым и Е.Л. Лебедевым при участии М.Л. Едемского в Буреинском бассейне (Хабаровский край, Дальний Восток России). Коллекция хранится в Геологическом институте РАН (ГИН РАН) в Москве под № 3318.
Изученный нами материал происходит из обнажений правого берега р. Бурея в районе Катаевской протоки и ниже устья р. Умальта, а также из отвалов шахты № 3 около пос. Чегдомын (рис. 1).
Рис. 1.
(a) Схематическая геологическая карта юрских и меловых континентальных отложений Буреинского бассейна; (б) карта местонахождений остатков Pseudotorellia: I – обнажение ниже устья р. Умальта, II – разрезы вдоль берегов Катаевской протоки, III – место отвалов из шахты № 3; (в) обобщенный стратиграфический разрез талынджанской и дубликанской свит (по Маркевич, Бугдаева, 2014, с изменениями).

В отложениях, выступающих по берегам Катаевской протоки, были найдены остатки растений, описанные Долуденко как Pseudotorellia angustifolia (ГИН РАН, экз. №№ 3318/120-26, 3318/120-27) и P. ensiformis (ГИН РАН, экз. №№ 3318/118-1, 3318/118-2, 3318/120-25). Из обнажения вдоль правого берега р. Бурея, ниже устья р. Умальта, происходят остатки листьев, описанных Долуденко как P. ensiformis (ГИН РАН, экз. №№ 3318/52-3, 3318/52-4, 3318/54-9, 3318/57-6), P. pulchella (ГИН РАН, экз. № 3318/57-22) и P. crassifolia (ГИН РАН, экз. № 3318/58-3). Из отвалов шахты № 3 возле поселка Чегдомын были собраны образцы с ископаемыми листьями, описанными Долуденко как P. longifolia (ГИН РАН, экз. № 3318/9-38). К сожалению, нам не удалось обнаружить некоторые образцы из коллекции ГИН РАН № 3318 (экз. №№ 3318/52-3, 3318/54-9, 3318/118-1, 3318/118-2, 3318/120-25), но в нашем распоряжении имеются сделанные Долуденко препараты с кутикулами листьев с такими же номерами.
Также были изучены образцы с остатками листьев Pseudotorellia из коллекции, собранной В.А. Красиловым в том же местонахождении ископаемых растений на правом берегу р. Бурея, ниже устья р. Умальта. Коллекция хранится в Федеральном научном центре биоразнообразия наземной биоты Восточной Азии ДВО РАН (ФНЦ Биоразнообразия ДВО РАН) во Владивостоке под № 515 (экз. №№ 515/272 и 515/284).
Ископаемые кутикулы были обработаны по стандартной методике мацерации. Полученные препараты кутикул были изучены как в световом микроскопе Carl Zeiss Axio Scope.A1 с цифровой фотокамерой Lomo Microsystems MC-6.3, так и при помощи сканирующего электронного микроскопа JSM-6390 LA (СЭМ) в Ботаническом институте РАН (БИН РАН), в световом микроскопе Axiostar plus с цифровой фотокамерой Canon PS A640 в ГИН РАН (Москва), в световом микроскопе Carl Zeiss Axioscop 40, оборудованном цифровой фотокамерой Axiocam HRc, и при помощи СЭМ Carl Zeiss EVO 40 в ФНЦ Биоразнообразия ДВО РАН (Владивосток).
Препараты с кутикулами листьев хранятся в лаборатории палеофлористики ГИН РАН в Москве, в лаборатории палеоботаники БИН РАН в Санкт-Петербурге и в лаборатории палеоботаники ФНЦ Биоразнообразия ДВО РАН во Владивостоке.
СТРАТИГРАФИЯ ЮРСКИХ И НИЖНЕМЕЛОВЫХ ФЛОРОНОСНЫХ ОТЛОЖЕНИЙ БУРЕИНСКОГО БАССЕЙНА
Флороносные слои с остатками Pseudotorellia правого берега р. Бурея, ниже устья р. Умальта и вдоль Катаевской протоки (азановская толща), Вахрамеев относил к талынджанской свите, а близ пос. Чегдомын – сначала к солонийской (верхней) подсвите ургальской свиты (Вахрамеев, Долуденко, 1961), а позже к верхней части талынджанской свиты (Вахрамеев, Лебедев, 1967, с. 127).
Еще раньше Т.Н. Давыдова и Ц.Л. Гольдштейн (1949) предложили расчленить ургальскую свиту на две свиты: дубликанскую и солонийскую. Однако Вахрамеев и Долуденко (1961) использовали именно название “ургальская свита”, подразделяя ее на две подсвиты: дубликанскую и солонийскую. Позже эти подсвиты были переведены в ранг свит (Решения…, 1994).
Вахрамеев и Лебедев датировали ископаемые растения талынджанской и дубликанской свит поздней юрой, полагая, что они заметно отличаются от таковых солонийской свиты, залегающей в основании нижнего мела (Вахрамеев, Лебедев, 1967; Лебедев, Паракецов, 1975).
Красилов (1973), детально изучивший все известные разрезы Буреинского бассейна, заметил, что флористический комплекс из отложений, развитых вдоль Катаевской протоки (азановская толща), сходен с таковым дубликанской свиты. К этой же свите он отнес флороносные отложения на левом берегу р. Ургал, а также отложения близ пос. Чегдомын. Возраст флористического комплекса талынджанской свиты Красилов определил как келловей–оксфорд, дубликанской – как титон–берриас.
В последние годы было закончено крупно- и среднемасштабное геологическое картирование Буреинского бассейна, составлены карты нового поколения (Анойкин, 2003; Государственная…, 2009). Возраст свит и толщ указан в соответствии с решениями, принятыми на IV Межведомственном региональном стратиграфическом совещании по докембрию и фанерозою юга Дальнего Востока и Восточного Забайкалья (Решения…, 1994).
По литологическому составу и структурно-текстурным особенностям пород в талынджанской свите выделяются три пачки, из которых две нижние пачки объединены в нижнюю подсвиту, а верхняя соответствует верхней подсвите. Возраст нижней подсвиты установлен как келловей, верхней подсвиты – оксфорд (Решения…, 1994). Слои с остатками листьев Pseudotorellia относятся к верхней подсвите талынджанской свиты.
На талынджанской свите с местным размывом несогласно залегает дубликанская свита. В (Решения…, 1994) дубликанская свита датируется волжским веком. В.С. Маркевич (1995; Маркевич, Бугдаева, 2009) по данным спорово-пыльцевого анализа установила возраст дубликанской свиты как берриас. В объяснительной записке к Государственной геологической карте Российской Федерации масштаба 1 : 1 000 000 (лист М-52) возраст флористического комплекса дубликанской свиты трактуется как “верхи верхней юры (волжский или титонский ярус), хотя не исключаются и низы мела (берриас)” (Петрук и др., 2012, с. 34).
Авторы настоящей работы придерживаются более обоснованной точки зрения Красилова (1973) и принимают возраст талынджанской свиты как келловей–оксфорд, а дубликанской – как титон–берриас (рис. 1).
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ ВИДОВ PSEUDOTORELLIA
КЛАСС GINKGOOPSIDA
ПОРЯДОК VLADIMARIALES GORDENKO, 2010
СЕМЕЙСТВО UMALTOLEPIDACEAE STANISLAVSKY, 1973, EMEND. Z. ZHOU, 1991
Род Pseudotorellia Florin, 1936
Pseudotorellia angustifolia Doludenko, 1961, emend. Nosova
Табл. I , фиг. 1–4 ; табл. II
Pseudotorellia angustifolia: Вахрамеев, Долуденко, 1961, с. 114, табл. LVII , фиг. 1–9 .
Pseudotorellia angustifolia: Красилов, 1972, с. 58, pro parte, табл. XX , фиг. 1–11 ; табл. XXI , фиг. 1, 2, 4, 6–13 ; рис. 9, 10 а–10и, 10с, 10т.
Pseudotorellia angustifolia: Костина, 2004, с. 59, табл. XXXIX , фиг. 1–9 .
Pseudotorellia angustifolia: Киричкова и др., 2005, с. 107, табл. LI , фиг. 1–8 ; табл. LII , фиг. 1–5 .
Pseudotorellia angustifolia: Киричкова и др., 2020, с. 110, табл. CXV , фиг. 1–8 .
Pseudotorellia longifolia Doludenko: Вахрамеев, Долуденко, 1961, с. 115, табл. LVIII , фиг. 1–8 .
Pseudotorellia longifolia: Долуденко, Рассказова, 1972, с. 22, табл. XXX , фиг. 1–7 ; табл. XXXI , фиг. 1–4 .
Pseudotorellia angaranica Kiritchkova et Nosova: Киричкова и др., 2020, с. 108, табл. CXVII , фиг. 7, 8 ; табл. CXVIII , фиг. 2 а, 7–10; табл. CXXI , фиг. 1а, 2–11.
Голотип – ГИН РАН, № 3318/120-27b; Дальний Восток России, правый берег р. Бурея, Катаевская протока; дубликанская свита, титон–берриас; Вахрамеев, Долуденко, 1961, с. 114, табл. LVII , фиг. 6 ; обозначен здесь, табл. I , фиг. 2b, 3 .
Таблица I . Листья Pseudotorellia. Длина масштабной линейки 10 мм. 1–4 – Pseudotorellia angustifolia Doludenko, дубликанская свита, титон–берриас: 1–3 – правый берег р. Бурея, Катаевская протока: 1 – скопление листьев, экз. № 3318/120-26; 2 – скопление листьев: 2а – экз. № 3318/120-27a, 2b – экз. № 3318/120-27b; 3 – увеличенный лист с фиг. 2 , экз. № 3318/120-27b (голотип); 4 – пос. Чегдомын (шахта 3), скопление листьев, экз. № 3318/9-38; 5, 6, 8 – Pseudotorellia doludenkoae Nosova, Kostina et Bugdaeva, правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд: 5a–5d – четыре листа, экз. № 515/284; 6 – увеличенный лист с фиг. 5a , экз. № 515/284а (голотип); 8 – скопление листьев, экз. № 515/272; 7, 9, 10 – Pseudotorellia crassifolia Doludenko, правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд: 7a, 7b – листья, экз. № 3318/58-3 (7а – голотип); 9 – увеличенные верхушки листьев с фиг. 10 , экз. № 3318/57-22; 10 – скопление листьев, экз. № 3318/57-22.

Таблица II. Pseudotorellia angustifolia Doludenko, дубликанская свита, титон–берриас. Длина масштабной линейки для фиг. 1–6 , 8, 9 – 100 мкм; для фиг. 7, 10 – 50 мкм; для фиг. 11–13 – 20 мкм. 1, 4, 13 – правый берег р. Бурея, Катаевская протока, экз. № 3318/120-27b (голотип): 1 – устьица на нижней поверхности листа; 4 – клетки верхнего эпидермиса; 13 – устьице, видны полярные выросты замыкающих клеток и сохранившиеся внутренние стенки замыкающих клеток; 2, 3, 5–12 – пос. Чегдомын (шахта 3), экз. № 3318/9-38: 2 – смоляной тяж; 3 – верхняя (слева) и нижняя (справа) кутикулы; 5 – клетки верхнего эпидермиса; 6 – нечеткие устьичные полосы на нижней стороне листа; 7 – два устьица; 8 – устьица и клетки нижнего эпидермиса, СЭМ, вид изнутри; 9 – устьичные (три) и безустьичные (две) полосы; 10 – два устьица, одно из которых – с проксимальными папиллами; 11 – устьице с проксимальными папиллами; 12 – устьице, СЭМ, вид изнутри.
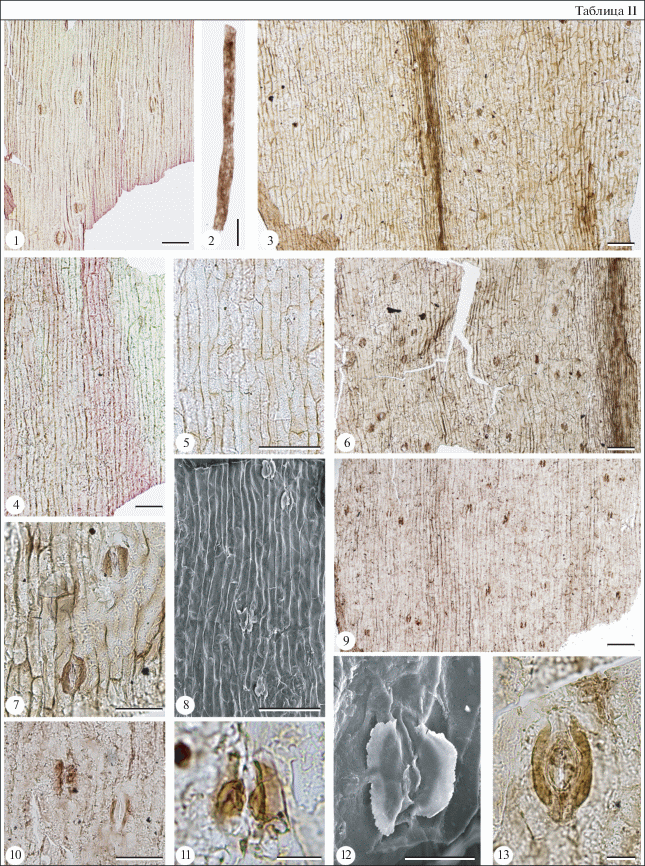
Таблица III. Pseudotorellia crassifolia Doludenko, правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд. Длина масштабной линейки для фиг. 1, 2, 4–6, 8 – 100 мкм; для фиг. 3 – 1000 мкм; для фиг. 7, 9–12 – 50 мкм. 1–6, 8–12 – экз. № 3318/57-22: 1 – кутикула верхней поверхности листа; 2 – клетки интеркостальной зоны верхнего эпидермиса; 3 – расположение смоляных тяжей; 4 – кутикула нижней поверхности листа; 5 – клетки безустьичной (костальной) зоны нижнего эпидермиса; 6 – фрагмент устьичной полосы; 8 – смоляной тяж; 9–10 – устьица; 11 – устьице с сохранившимися апертурными и эпидермальными стенками замыкающих клеток, вид изнутри, СЭМ; 12 – устьица, на одном из них видны сохранившиеся внутренние стенки замыкающих клеток, вид изнутри, СЭМ; 7 – устьица на нижней поверхности листа, экз. № 3318/58-3 (голотип).

Таблица IV. Pseudotorellia doludenkoae Nosova, Kostina et Bugdaeva, sp. nov. Длина масштабной линейки для фиг. 1–7 – 100 мкм; для фиг. 8 – 50 мкм. 1–3, 6 – правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд: 1, 6 – фрагменты нижнего эпидермиса, экз. № 515/284а (голотип); 2 – фрагмент верхнего эпидермиса, экз. № 515/284а (голотип); 3 – устьичные и безустьичные полосы нижнего эпидермиса, экз. № 515/272; 4, 5, 7, 8 – правый берег р. Бурея, Катаевская протока; дубликанская свита, титон–берриас: 4, 5 – эпидермальные клетки верхнего эпидермиса: 4 – экз. № 3318/54-90; 5 – экз. № 3318/52-4; 7 – клетки безустьичной полосы, экз. № 3318/120-28; 8 – устьице, экз. № 3318/52-4.

Таблица V. Pseudotorellia doludenkoae Nosova, Kostina et Bugdaeva, sp. nov. Масштабная линейка для фиг. 1, 3–7, 9, 11 – 100 мкм; для фиг. 2, 8, 10 – 50 мкм. 1, 2 – правый берег р. Бурея, Катаевская протока; дубликанская свита, титон–берриас: 1 – краевая устьичная полоса нижнего эпидермиса (слева) и клетки верхнего эпидермиса (справа), экз. № 3318/57-6; 2 – устьица, экз. № 3318/118; 3–11 – правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд: 3 – устьичные и безустьичные полосы, СЭМ, вид изнутри, экз. № 515/272; 4 – устьица, экз. № 515/272; 5 – устьице, экз. № 515/284а (голотип); 6 – смоляной тяж, экз. № 515/284а (голотип); 7 – клетки верхнего эпидермиса, СЭМ, вид изнутри, экз. № 515/272; 8 – устьице с сохранившимися внутренними стенками замыкающих клеток, СЭМ, вид изнутри, экз. № 515/272; 9 – устьица, СЭМ, вид изнутри, экз. № 515/284а (голотип); 10 – устьице, СЭМ, вид снаружи, экз. № 515/284а (голотип); 11 – устьичные полосы и смоляной тяж, СЭМ, вид изнутри, экз. № 515/284а (голотип).

Emended diagnosis. Leaves linear, narrow and long, apex rounded or acute. Veins 3–8 per widest part. Leaves hypostomatic. Ordinary epidermal cells of upper and lower epidermis elongated and rectangular, without cuticular ridges or papillae, anticlinal cell walls straight. Stomata sparse, arranged in bands, rarely irregularly scattered in the lower epidermis. Subsidiary cells without papillae, rarely with small proximal papillae.
Описание. Листья линейные (табл. I , фиг. 1–4 ). Ширина листьев 1.5–5 мм в средней части, длина 60–80 мм и более. Верхушка заостренная или округлая. В средней части листа 3–8 жилок (8–10 жилок на 5 мм ширины листовой пластинки). Между жилками расположены смоляные тяжи 65–116 мкм шириной, максимально сохранившаяся длина 1000 мкм (табл. II, фиг. 2 ).
Листья гипостоматные (табл. II, фиг. 3 ). Верхний эпидермис сложен удлиненными узкими четырехугольными клетками (8–38 × 60–130 мкм, ширина/длина = 1/3–1/13, чаще 1/5–1/7), часто образующими ряды (табл. II, фиг. 3–5 ). Углы эпидермальных клеток острые, реже закругленные; антиклинальные стенки прямые, периклинальные ровные.
Нижний эпидермис состоит из чередующихся устьичных (139–383 мкм) и безустьичных (157–306 мкм) полос (табл. II, фиг. 3, 6, 9 ). На некоторых фрагментах листьев устьичные полосы выражены нечетко, местами сливаясь друг с другом, в отдельных случаях устьица не образуют полос (табл. II, фиг. 6 ). Основные клетки как устьичных, так и безустьичных полос такие же по форме и размерам, как и клетки верхнего эпидермиса, с прямыми антиклинальными и ровными периклинальными стенками. На ширину устьичной полосы приходится 1–3 устьица в узких листьях (табл. II, фиг. 1, 3 ) и до 4 устьиц в широких листьях (табл. II, фиг. 9 ). Устьица в устьичных полосах расположены достаточно редко, не образуют рядов, ориентированы продольно. Устьичные комплексы моноциклические, чаще удлиненной формы. Большинство побочных клеток устьиц не имеют папилл, в редких случаях встречаются небольшие проксимальные папиллы (табл. II, фиг. 10, 11 ). В полярных частях устьиц видны выросты замыкающих клеток (табл. II, фиг. 13 ). Длина устьиц с этими выростами 70–76 мкм, длина замыкающих клеток без выростов 34–65 мкм, ширина каждой – 10–13 мкм. После мацерации сохраняются наружные стенки, иногда также апертурные и эпидермальные, реже внутренние стенки замыкающих клеток (табл. II, фиг. 8, 12, 13 ). Эпидермальные стенки замыкающих клеток часто подогнуты в сторону апертуры и видны на световом микроскопе в виде двух узких латеральных полумесяцев (табл. II, фиг. 1, 7, 10, 13 ).
Сравнение и замечания. Согласно сделанным Долуденко описаниям (Вахрамеев, Долуденко, 1961), листья P. longifolia немного шире (3–5 мм) листьев P. angustifolia (1.5–3 мм). Ревизия образцов показала, что среди листьев, описанных Долуденко как P. longifolia, встречаются экземпляры с шириной 2–4 мм. В соответствии с шириной листьев количество жилок тоже варьирует, при этом количество жилок на 5 мм ширины листа остается постоянным. Таким образом, существенных морфологических отличий между этими двумя видами мы не обнаружили.
В диагнозе P. longifolia указано, что устьица собраны в полосы, однако в описании вида отмечается, что устьица разбросаны по всей нижней поверхности, не образуя полос и рядов (Вахрамеев, Долуденко, 1961, с. 115–116). Позднее, описывая под этим же видовым названием остатки листьев из Иркутского бассейна, Долуденко и Рассказова (1972) отметили расположение устьиц в полосах. Изучив эпидермальное строение типовых экземпляров P. longifolia, мы обнаружили, что устьица на нижней поверхности листьев расположены в полосах, хотя местами эти полосы выражены нечетко, сливаясь друг с другом (табл. II, фиг. 6 ). Согласно приведенным Долуденко описаниям, P. angustifolia отличается от P. longifolia наличием редких папилл на побочных клетках устьиц. Однако наше исследование показало, что редкие папиллы встречаются и на побочных клетках устьиц листьев, ранее описанных как P. longifolia. На основании сходства морфологических и эпидермальных признаков экземпляры, описанные Долуденко как P. longifolia, рассматриваются в составе вида P. angustifolia.
Красилов (1972) включил в состав Pseudotorellia angustifolia описанный Долуденко вид P. ensiformis (Вахрамеев, Долуденко, 1961), добавив в описание P. angustifolia следующие эпидермальные признаки: наличие срединных кутикулярных утолщений (= валиков) на периклинальных стенках клеток верхней поверхности листа и частое присутствие папилл на побочных клетках устьиц. Изучив P. ensiformis, мы нашли ряд существенных как морфологических, так и эпидермальных отличий от P. angustifolia. Листья P. ensiformis заметно отличаются от P. angustifolia ланцетовидной формой, бόльшей шириной и почти в 10 раз меньшей длиной. В отличие от P. angustifolia, клетки верхнего эпидермиса P. ensiformis дифференцированы на узкие удлиненные клетки, расположенные над жилками, и широкие как удлиненные, так и короткие, расположенные между ними. Кроме того, P. angustifolia отличается от P. ensiformis отсутствием кутикулярных валиков на эпидермальных клетках. Полученные нами данные подтверждают мнение Долуденко о том, что буреинские ланцетовидные листья следует выделить в отдельный вид, но, принимая во внимание то, что эпидермальное строение типовых экземпляров P. ensiformis из Усть-Балея неизвестно, мы выделяем новый вид P. doludenkoae sp. nov. (см. описание ниже).
Pseudotorellia angaranica (Киричкова и др., 2020) из Иркутского бассейна по морфологическим и эпидермальным признакам сходна с P. angustifolia из Буреинского бассейна, на основании чего мы рассматриваем этот иркутский вид как младший синоним P. angustifolia.
В целом Pseudotorellia angustifolia характеризуется узкими и длинными линейными листьями; узкими удлиненными клетками как верхнего, так и нижнего эпидермиса; отсутствием кутикулярных утолщений и папилл на основных эпидермальных клетках; наличием редких небольших проксимальных папилл на некоторых побочных клетках устьиц.
Распространение. Дубликанская свита (титон–берриас) Буреинского бассейна (Хабаровский край), переясловская (нижняя юра) и камалинская (средняя юра) свиты Канского бассейна (Восточная Сибирь), тюменская свита (байос) площади Южно-Часельская (Ямало-Ненецкий автономный округ), наунанская свита (келловей–оксфорд) площади Западно-Останинская (Томская область), присаянская свита (аален–байос) Иркутского бассейна (Восточная Сибирь).
Материал. Правый берег р. Бурея, Катаевская протока, ГИН РАН, экз. № 3318/120-26 (более 8 листьев и их фрагментов), экз. № 3318/120-27 (5 фрагментов листьев); пос. Чегдомын (шахта № 3), ГИН РАН, экз. № 3318/9-38 (более 15 листьев и их фрагментов), дубликанская свита (титон–берриас).
Pseudotorellia crassifolia Doludenko, emend. Nosova, Kostina et Bugdaeva
Табл. I , фиг. 7, 9, 10 ; табл. III
Pseudotorellia crassifolia (Prynada) Doludenko: Вахрамеев, Долуденко, 1961, с. 114, табл. LVI , фиг. 4, 5 .
Pseudotorellia pulchella auct. non (Heer) Vassilevskaja, 1959: Вахрамеев, Долуденко, 1961, с. 113, табл. LVI , фиг. 6, 7 .
Голотип – ГИН РАН, № 3318/58-3; Дальний Восток России, правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд; Вахрамеев, Долуденко, 1961, с. 113, табл. LVI , фиг. 4 ; обозначен здесь, табл. I , фиг. 7 .
Emended diagnosis. Leaves oblanceolate, with their widest part at about the middle or slightly closer to the apex, apex rounded or obtuse; base cuneate. Leaves with 5–6 veins per 5 mm. Leaves hypostomatic. Cells of upper and lower epidermis tetra- to polygonal, elongate to short across intercostal zones and elongate across costal zone; periclinal cell walls smooth. Anticlinal epidermal cell walls of both leaf surfaces straight. Stomata assembled in bands. Stomatal complexes monocyclic, sometimes dicyclic. Subsidiary cells without papillae.
Описание. Листья обратноланцетные с тупой или округлой верхушкой (табл. I , фиг. 7, 9, 10 ). Длина листьев 30–45 мм, ширина у основания 1–1.5 мм, в верхней трети – 4–12 мм. Число жилок в широкой части листа колеблется от 8 до 15 (5–6 жилок на 5 мм ширины листовой пластинки). Между жилками располагаются смоляные тяжи шириной 28–60 мкм (табл. III, фиг. 3 ). Наибольшая сохранившаяся длина тяжа – 2300 мкм (табл. III, фиг. 8 ). Кутикула листьев тонкая.
Листья гипостоматные. По форме эпидермальных клеток костальные (над жилками) и интеркостальные (между жилками) зоны на верхней поверхности листа выражены не всегда четко и примерно равны по ширине (табл. III, фиг. 1). Зоны над жилками образованы рядами четырех- и многоугольных удлиненных эпидермальных клеток как широких (35–46 × 93–115 мкм), так и узких (20–28 × 100–167 мкм). В зонах между жилками клетки эпидермиса четырех- и многоугольные (табл. III, фиг. 2 ), как короткие (30–89 × 45–92 мкм), так и удлиненные (34–56 × 70–114 мкм). Антиклинальные стенки всех клеток верхнего эпидермиса прямые, углы между ними острые, реже закругленные. Периклинальные стенки клеток ровные.
Нижний эпидермис состоит из чередующихся безустьичных и устьичных полос, соответствующих костальным и интеркостальным зонам (табл. III, фиг. 4 ). Основные эпидермальные клетки четырех- и многоугольные, в безустьичных зонах (табл. III, фиг. 5 ) удлиненные (20–45 × 109–140 мкм), в устьичных полосах – как короткие (37–41 × 57–65 мкм), так и удлиненные (25–37 × 99–128 мкм). Антиклинальные стенки клеток прямые, углы между ними острые, реже закругленные. Периклинальные стенки клеток ровные. В пределах полосы устьица расположены очень неравномерно, ориентированы продольно. На ширину полосы приходится три–пять (редко до семи) устьиц (табл. III, фиг. 4 ). Устьичные комплексы моноциклические (табл. III, фиг. 9 ), иногда неполно дициклические (табл. III, фиг. 10 ). Побочных клеток устьиц пять–шесть, две из них полярные; папиллы на побочных клетках отсутствуют. Замыкающие клетки погруженные, крыловидной формы (табл. III, фиг. 7, 10 ). Длина замыкающих клеток 26–47 мкм, ширина каждой – 8–20 мкм, их полярные выросты не видны. После мацерации сохраняются наружные стенки, а также часто апертурные и эпидермальные (табл. III, фиг. 9, 11, 12 ), реже внутренние стенки замыкающих клеток (табл. III, фиг. 12 ).
Сравнение и замечания. Долуденко описала комбинацию P. crassifolia (Prynada) Doludenko, указав в синонимике В.Д. Принаду как автора видового названия: “Feildenia crassifolia Prynada, Принада, 1939 в кн. Херасков и др.” (Вахрамеев, Долуденко, 1961, с. 114). Однако в работе Хераскова и др. (1939) приведены только списки видов без их описаний и изображений. F. crassifolia была описана Принадой в неопубликованном геологическом отчете в 1937 г., а значит, этот вид является невалидным. Поскольку Долуденко указала тип вида, ошибочно обозначив его как неотип, и привела видовой диагноз, опубликованный таксон P. crassifolia (Вахрамеев, Долуденко, 1961) является валидным. За голотип в настоящей работе принят образец, указанный Долуденко в качестве неотипа.
Изучено также эпидермальное строение буреинских листьев, описанных как P. pulchella, и обнаружено, что по всем признакам они сходны с P. crassifolia. Поскольку типовой материал P. crassifolia происходит из Буреинского бассейна, в отличие от P. pulchella, буреинские листья описаны под видовым названием P. crassifolia.
P. crassifolia значительно отличается от P. angustifolia обратноланцетовидной формой листьев, в 2–4 раза большей шириной и в 2 раза меньшей длиной. В отличие от P. crassifolia, эпидермальные клетки обеих поверхностей листьев P. angustifolia узкие и сильно удлиненные, устьичные полосы более узкие, устьичные комплексы узкие моноциклические, а замыкающие клетки крупнее.
Распространение. Верхняя подсвита талынджанской свиты (оксфорд) Буреинского бассейна (Хабаровский край).
Материал. Правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд; ГИН РАН, экз. № 3318/58-3 (три фрагмента листьев), экз. № 3318/57-22 (более 18 листьев и их фрагментов).
Pseudotorellia doludenkoae Nosova, Kostina et Bugdaeva, sp. nov.
Табл. I , фиг. 5, 6, 8 ; табл. IV, V
Pseudotorellia ensiformis (Heer) Doludenko: Вахрамеев, Долуденко, 1961, с. 111, табл. LV , фиг. 1–8 ; табл. LVI , фиг. 1–3 .
Pseudotorellia angustifolia Doludenko: Красилов, 1972, с. 58, pro parte, табл. XXI , фиг. 3, 5 .
Название вида в честь палеоботаника М.П. Долуденко.
Голотип – ФНЦ Биоразнообразия ДВО РАН, № 515/284a, Дальний Восток России, правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд; Красилов, 1972, табл. XXI , фиг. 5 ; обозначен здесь, табл. I , фиг. 5 а, 6.
Diagnosis. Leaves oblanceolate, with their widest part at about the middle or slightly closer to the apex. Leaf apex rounded or obtuse; base cuneate. Leaves with 5–6 veins per 5 mm in widest part. Leaves hypostomatic. Cells of upper and lower epidermis tetra-, rarely pentagonal; elongate to short across intercostal zones and elongate across costal zones. Some cells of the upper epidermis and the most cells of the lower epidermis with longitudinal cuticular ridges, anticlinal cell walls of both leaf surfaces straight. Stomata assembled in narrow bands. Subsidiary cells without papillae, very rarely with small papillae.
Описание. Листья обратноланцетные, c тупой или округлой верхушкой (табл. I , фиг. 5, 6, 8 ). Длина листьев 38–65 мм, ширина у основания 1–2 мм, в верхней трети – 6–13 мм. В средней части листа 8–10 жилок (5–6 жилок на 5 мм ширины листовой пластинки). Между жилками располагаются смоляные тяжи шириной 59–80 мкм (табл. V, фиг. 6, 11 ).
Листья гипостоматные. Верхний эпидермис сложен четырех- и пятиугольными клетками, как короткими (30–50 × 36–115 мкм), так и узкими удлиненными (18–30 × 110–170 мкм), расположенными преимущественно над жилками (табл. IV, фиг. 2, 4, 5 ; табл. V, фиг. 7 ). Углы эпидермальных клеток закругленные, реже острые; антиклинальные стенки прямые. Клетки верхнего эпидермиса около сгиба листа имеют кутикулярные валики (табл. V, фиг. 1), остальные эпидермальные клетки верхней поверхности – с ровными периклинальными стенками (табл. IV, фиг. 2, 4, 5 ).
Нижний эпидермис состоит из чередующихся устьичных (130–310 мкм шириной) и безустьичных (215–400 мкм шириной) полос (табл. IV, фиг. 1, 3, 6 ; табл. V, фиг. 3, 11 ). Клетки безустьичных полос четырех- и пятиугольные, узкие удлиненные (10–30 × 110–195 мкм), в устьичных полосах клетки как удлиненные, так и короткие (30–55 × 40–156 мкм); антиклинальные стенки прямые. Большинство клеток имеют продольный кутикулярный валик, иногда прослеживающийся из одной клетки в другую (табл. IV, фиг. 6 ; табл. V, фиг. 1, 5 ). Валики от узких (табл. IV, фиг. 3, 7 ; табл. V, фиг. 1) до широких (табл. IV, фиг. 1, 6 ; табл. V, фиг. 5 ), расположены в центре клетки (табл. IV, фиг. 3, 7, 8 ; табл. V, фиг. 1, 4 ) или смещены к одной из продольных антиклинальных стенок (табл. IV, фиг. 6 ; табл. V, фиг. 5 ). Количество устьиц в устьичной полосе от 2 до 5 (табл. IV, фиг. 1, 3, 6 ; табл. V, фиг. 1, 3, 11 ). Устьица в устьичных полосах не образуют четких продолжительных рядов, ориентированы продольно. Устьичные комплексы моноциклические, редко неполно дициклические. Большинство устьичных комплексов без папилл на побочных клетках (табл. IV, фиг. 3, 8 ; табл. V, фиг. 1, 4, 5, 10 ), редко некоторые побочные клетки имеют небольшие проксимальные папиллы (табл. IV, фиг. 1, 6 ; табл. V, фиг. 2 ). В полярных частях устьиц видны выросты в виде хвоста ласточки или Т-образные (табл. IV, фиг. 8 ; табл. V, фиг. 2 ). Длина устьиц с этими выростами 65–90 мкм, длина замыкающих клеток без выростов 38–63 мкм, ширина каждой – 10–18 мкм. После мацерации сохраняются наружные стенки, а также иногда и внутренние стенки замыкающих клеток (табл. IV, фиг. 8 ; табл. V, фиг. 5, 8, 9 ).
Сравнение и замечания. Комбинация Pseudotorellia ensiformis была предложена Долуденко на основе сходства морфологических признаков буреинского материала с Podozamites ensiformis, описанным Геером из юрских отложений местонахождения Усть-Балей в Иркутском бассейне (Heer, 1876). Однако, в отличие от иркутских листьев, для буреинских было изучено их эпидермальное строение (Вахрамеев, Долуденко, 1961). Позже Долуденко и Рассказова отметили, что отождествление буреинских экземпляров Pseudotorellia ensiformis с иркутскими было неверным, поскольку эпидермальные признаки иркутских листьев неизвестны (Долуденко, Рассказова, 1972, с. 21), и предложили относить к P. ensiformis листья с неизвестными эпидермальными признаками, имеющие такие же морфологические характеристики, как и выбранный ими лектотип из Усть-Балея. В результате листья из Буреинского бассейна, описанные Долуденко как P. ensiformis, мы выделяем в новый вид.
По морфологии P. doludenkoae сходна с P. crassifolia (см. описание выше), но существенно отличается по эпидермальным признакам. Кутикула листьев нового вида заметно толще кутикулы P. crassifolia. Кроме этого, P. doludenkoae существенно отличается от P. crassifolia наличием кутикулярных валиков на периклинальных стенках эпидермальных клеток, а также бόльшим размером устьиц.
P. doludenkoae отличается от P. angustifolia ланцетовидной формой листьев, их большей шириной и меньшей длиной. В отличие от P. angustifolia, клетки верхнего эпидермиса P. doludenkoae дифференцированы по форме на узкие удлиненные, расположенные над жилками, и широкие как удлиненные, так и короткие, расположенные между полосами узких клеток. Кроме того, новый вид отличается от P. angustifolia наличием кутикулярных валиков на эпидермальных клетках.
По форме и размерам листьев, а также по расположению устьиц в полосах, наличию кутикулярных валиков на основных эпидермальных клетках и отсутствию папилл на побочных клетках большинства устьиц P. doludenkoae сходна с P. grojecensis Reymanówna из нижней юры Польши (Reymanówna, 1963), P. krassilovii Bugdaeva из нижнего мела Южного Приморья (Bugdaeva, Markevich, 2009) и P. retusa Bose et Manum из нижнего мела Шпицбергена (Bose, Manum, 1990; Pott et al., 2016).
P. grojecensis отличается от P. doludenkoae своеобразным строением верхнего эпидермиса, сложенного из “пачек” четырехугольных клеток одинаковой длины, ориентированных то вдоль, то поперек оси листа.
В отличие от нового вида, у P. krassilovii кутикулярные утолщения на периклинальных стенках основных эпидермальных клеток образованы скоплениями небольших уплощенных папилл. Кроме этого, у P. krassilovii ширина устьичных полос в три раза шире безустьичных, в то время как у P. doludenkoae безустьичные полосы равны или немного шире устьичных полос.
P. retusa отличается от P. doludenkoae меньшими размерами листьев (3–5 × 10–19 мм), зубчатым краем листовой пластинки, большей шириной устьичных полос (до 6 устьиц на ширину полосы) и слабоизвилистыми антиклинальными стенками некоторых эпидермальных клеток.
Распространение. Верхняя подсвита талынджанской свиты (оксфорд) и дубликанская свита (титон–берриас) Буреинского бассейна (Хабаровский край).
Материал. Правый берег р. Бурея, ниже устья р. Умальта; верхняя подсвита талынджанской свиты, оксфорд; ФНЦ Биоразнообразия ДВО РАН, экз. № 515/272 (более 10 листьев и их фрагментов), экз. № 515/284 (четыре листа); ГИН РАН, экз. №№ 3318/52-3, 3318/52-4, 3318/54-9; Катаевская протока; дубликанская свита, титон–берриас; ГИН РАН, экз. №№ 3318/118-1, 3318/ 118-2, 3318/120-25.
ЗАКЛЮЧЕНИЕ
Отдельные листья Pseudotorellia и их фрагменты по морфологическим признакам сходны с некоторыми листьями Phoenicopsis Heer, Eretmophyllum Thomas, Podozamites Braun, а также с фрагментами листьев Baiera Braun и Sphenobaiera Florin, поэтому их определение должно основываться не только на морфологии, но и на строении эпидермиса. Эпидермальные признаки Pseudotorellia позволяют достаточно четко отличить этот род от других ископаемых таксонов, также они являются решающими и при выделении видов, поскольку сходные по морфологии листья Pseudotorellia часто имеют различное строение эпидермиса. Это подтверждают и наши исследования остатков Pseudotorellia из Б-уреинского бассейна, где у одинаковых по форме и размерам листьев P. crassifolia и P. doludenkoae sp. nov. эпидермальное строение совершенно разное. У большинства листьев Pseudotorellia кутикула достаточно толстая, устойчивая к химической мацерации, что позволяет детально изучать их эпидермальное строение и определять систематическую принадлежность не только целых листьев, но и их фрагментов, а также дисперсных кутикул. Это, в свою очередь, указывает на стратиграфическую значимость рода Pseudotorellia, особенно при изучении скважинного материала и углей, где сохранность растительных остатков не позволяет с уверенностью судить об их морфологии.
Систематический состав рода Pseudotorellia, а также стратиграфическая и географическая приуроченность видов неоднократно уточнялись разными авторами (Bugdaeva, 1999; Киричкова, Носова, 2009; Horiuchi, Uemura, 2017; Nosova, Golovneva, 2018). В результате проведенной нами ревизии число видов Pseudotorellia в Буреинском бассейне сократилось до трех, описанных как по морфологическим, так и по эпидермальным признакам (табл. 1).
Таблица 1.
Результаты ревизии видового состава рода Pseudotorellia в Буреинском бассейне
| Вахрамеев, Долуденко, 1961 | Красилов, 1972 | Результаты ревизии (данная статья) |
|---|---|---|
| P. angustifolia Doludenko | P. angustifolia Doludenko | P. angustifolia Doludenko |
| P. longifolia Doludenko | P. longifolia Doludenko | |
| P. pulchella (Heer) Vassilevskaja | P. (Eretmophyllum?) pulchella (Heer) Vassilevskaja | P. crassifolia Doludenko |
| P. crassifolia (Prynada) Doludenko | ||
| P. ensiformis (Heer) Doludenko | P. angustifolia Doludenko | P. doludenkoae sp. nov. |
Остатки Pseudotorellia angustifolia были обнаружены в дубликанской свите, P. crassifolia – в талынджанской, а ископаемые листья P. doludenkoae sp. nov. встречаются как в талынджанской, так и в дубликанской свите. В дальнейшем эти виды могут выступать в роли коррелятивных таксонов при внутрибассейновой и межрегиональной корреляции континентальных толщ.
Была проведена ревизия всех находок листьев, описанных ранее как Pseudotorellia angustifolia, и в состав этого вида были включены только те из них, у которых эпидермальные признаки сходны с таковыми изученных нами типовых листьев из Буреинского бассейна. На основе полученных данных можно сделать вывод, что вид P. angustifolia имел достаточно широкое распространение в юре и раннем мелу Сибири и Дальнего Востока. Помимо титон–берриаса Буреинского бассейна, этот вид известен из нижне- и среднеюрских отложений Канского бассейна (Костина, 2004), из среднеюрских отложений Ямало-Ненецкого автономного округа, средне-верхнеюрских отложений Томской области (Киричкова и др., 2005) и из среднеюрских отложений Иркутского бассейна (Киричкова и др., 2020). По-видимому, P. angustifolia появился в раннеюрскую эпоху на территории современной Восточной Сибири (Канский бассейн), в средней юре его ареал расширился как на северо-запад (Томская область и Ямало-Ненецкий автономный округ), где вид сохранился местами (Томская область) до поздней юры, так и на восток – до Иркутской области и Хабаровского края (Буреинский бассейн), где он сохранился до раннего мела (берриас). Возможно, подобный эволюционный стазис P. angustifolia был обусловлен существованием растения в одних и тех же условиях теплого и влажного палеоклимата данных территорий.
Благодарности. Работа выполнена на оборудовании центра коллективного пользования “Клеточные и молекулярные технологии изучения растений и грибов” БИН РАН (Санкт-Петербург), а также на оборудовании центра коллективного пользования ФНЦ Биоразнообразия ДВО РАН (Владивосток). Авторы благодарны Александру Борисовичу Доуэльду (Национальный институт карпологии (Гертнеровский Институт), Москва) за советы в решении вопросов синонимики и Максиму Алексеевичу Афонину (ФНЦ Биоразнообразия ДВО РАН, Владивосток) за помощь в поиске литературы.
Источники финансирования. Работа выполнена при поддержке гранта РФФИ № 20-04-00355 и в рамках госзаданий БИН РАН (тема № AAAA-A19-119021190031-8), ГИН РАН (тема № 0135-2019-0045) и ФНЦ Биоразнообразия ДВО РАН (тема № АААА-А17-117062710083-0).
Список литературы
Анойкин В.И. Государственная геологическая карта Российской Федерации. Масштаб 1 : 200 000. (Изд. 2-ое). Серия Буреинская. Лист M-53-VIII (Чегдомын). Объяснительная записка. СПб.: Картфабрика ВСЕГЕИ, 2003. 123 с. +5 вкл.
Василевская Н.Д. Голосеменные растения из угленосных отложений Сангарского района (Ленский угленосный бассейн) // Сборник статей по палеонтологии и биостратиграфии. Л.: НИИГА, 1959. С. 49–76.
Вахрамеев В.А., Долуденко М.П. Верхнеюрская и нижнемеловая флора Буреинского бассейна и ее значение для стратиграфии // Тр. ГИН АН СССР. 1961. Вып. 54. 132 с.
Вахрамеев В.А., Лебедев Е.Л. Палеоботаническая характеристика и возраст угленосных верхнемезозойских отложений Дальнего Востока (междуречье Амура и Уды) // Изв. АН СССР. Сер. геол. 1967. № 2. С. 120–133.
Головнева Л.Б., Носова Н.В. Альб-сеноманская флора Западной Сибири. СПб.: Марафон, 2012. 436 с.
Государственная геологическая карта Российской Федерации. Масштаб 1 : 1 000 000 (третье поколение). Серия Дальневосточная. Лист М-53 – Хабаровск. Объяснительная записка. СПб.: Картфабрика ВСЕГЕИ, 2009. 376 с.
Давыдова Т.П., Гольдштейн Ц.Л. Литологические исследования в Буреинском бассейне. М.: Госгеолиздат, 1949. 306 с.
Долуденко М.П., Рассказова Е.С. Гинкговые и чекановскиевые Иркутского бассейна // Мезозойские растения (гинкговые и чекановскиевые) Восточной Сибири. М.: Наука, 1972. С. 7–43.
Киричкова А.И., Носова Н.В. Род Pseudotorellia Florin (Ginkgoales): таксономический и стратиграфический аспекты // Стратиграфия. Геол. корреляция. 2009. Т. 17. № 6. С. 59–76.
Киричкова А.И., Костина Е.И., Быстрицкая Л.И. Фитостратиграфия и флора юрских отложений Западной Сибири. СПб.: Недра, 2005. 378 с.
Киричкова А.И., Носова Н.В., Костина Е.И., Ярошенко О.П. Континентальная юра Иркутского угленосного бассейна. СПб.: ВСЕГЕИ, 2020. 288 с.
Костина Е.И. Юрская флора Канского угленосного бассейна. М.: ГЕОС, 2004. 165 с.
Красилов В.А. Мезозойская флора реки Буреи (Ginkgoales и Czekanowskiales). М.: Наука, 1972. 151 с.
Красилов В.А. Материалы по стратиграфии и палеофлористике угленосной толщи Буреинского бассейна // Ископаемые флоры и фитостратиграфия Дальнего Востока. Владивосток: ДВНЦ АН СССР, 1973. С. 28–51.
Лебедев Е.Л., Паракецов К.В. О границе юры и мела в континентальных отложениях Дальнего Востока // Изв. АН СССР. Сер. геол. 1975. № 4. С. 124–133.
Маркевич В.С. Меловая палинофлора севера Восточной Азии. Владивосток: Дальнаука, 1995. 200 с.
Маркевич В.С., Бугдаева Е.В. Палинологическое обоснование возраста пограничных отложений юры и мела в Буреинском бассейне (Российский Дальний Восток) // Тихоокеанская геология. 2009. Т. 28. № 3. С. 90–99.
Маркевич В.С., Бугдаева Е.В. Позднеюрско-раннемеловые растения-углеобразователи Буреинского бассейна (российский Дальний Восток) // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 3. С. 14–30.
Петрук Н.Н., Волкова Ю.Р., Шилова М.Н. и др. Государственная геологическая карта Российской Федерации, масштаб 1 : 1 000 000 (третье поколение). Серия Дальневосточная. Лист М-52 – Благовещенск. Объяснительная записка. СПб.: ВСЕГЕИ, 2012. 496 с.
Решения IV Межведомственного регионального стратиграфического совещания по докембрию и фанерозою юга Дальнего Востока и Восточного Забайкалья, Хабаровск, 1990. Объяснительная записка к стратиграфическим схемам. Хабаровск: ХГГГП, 1994. 124 с.
Станиславский Ф.А. Новый род Toretzia из верхнего триаса Донбасса и его отношение к родам порядка Ginkgoales // Палеонтол. журн. 1973. Т. 1. С. 88–96.
Херасков Н.П., Давыдова Т.Н., Крашенинников Г.Ф., Пенинский Д.Д. Геология Буреинского бассейна. М.–Л.: ГОНТИ, 1939. 173 с. (Тр. Всесоюзного научно.-исслед. ин-та минерального сырья. Вып. 149).
Bose M.N., Manum S.B. Mesozoic conifer leaves with “Sciadopitys-like” stomatal distribution. A re-evaluation based on fossils from Spitsbergen, Greenland and Baffin Island // Norsk Polarinst. Skrifter. 1990. V. 192. P. 1–81.
Bugdaeva E.V. The history of the genus Pseudotorellia Florin (Pseudotorelliaceae, Ginkgoales) // Paleontol. J. 1999. V. 33. № 5. P. 566–576.
Bugdaeva E.V., Markevich V.S. The coal-forming plants of Rhabdopissites in the Lipovtsy Coal Field (Lower Cretaceous of Southern Primorye) // Paleontol. J. 2009. V. 43. № 10. P. 1217–1229.
Chen F., Meng X.Y., Ren S.Q., Wu C.L. The Early Cretaceous Flora of Fuxin Basin and Tiefa Basin, Liaoning Province. Beijing: Geol. Publishing House, 1988. 180 p. (in Chinese).
Dong C., Zhou Z., Zhang, B., Wang Y., Shi G. Umaltolepis and associated Pseudotorellia leaves from the Middle Jurassic of Yima in Henan Province, Central China // Rev. Palaeobot. Palynol. 2019. V. 271. P. 104–111.
Florin R. Die fossilen Ginkgophyten von Franz-Joseph-Land nebst Erorterung uber vermeintliche Cordaitales mesozoischen Alters. I. Spezieller Teil // Palaeontogr. Abt. B. 1936. Bd. 81. P. 71–173.
Gordenko N.V. Vladimariales ordo nov. (Gymnospermae) from the Middle Jurassic (Kursk Region, European Russia) // Paleontol. J. 2010. V. 44. № 10. P. 1281–1307.
Heer O. Beitrage zur Jura-Flora Ostsibiriens und des Amurlandes // Mem. l’Acad. Imperiale Sci. St. Petersbourg. 1876. Ser. VII. T. XXII. № 12. 122 p. (also Flora fossilis arctica. Zurich. V. 4).
Herrera F., Shi G., Ichinnorov N., Takahashi M., Bugdaeva E.V., Herendeen P.S., Crane P.R. The presumed ginkgophyte Umaltolepis has seedbearing structures resembling those of Peltaspermales and Umkomasiales // Proc. Nat. Acad. Sci. 2017. V. 114. P. E2385–E2391.
Horiuchi J., Uemura K. Paleocene occurrence of Pseudotorellia Florin (Ginkgoales) from Northeast Japan and the Meso-Cenozoic history of Pseudotorellia and Torellia // Rev. Palaeobot. Palynol. 2017. V. 246. P. 146–160.
Nosova N. Female reproductive structures of Umaltolepis Krassilov and associated short shoots, buds and leaves of Pseudotorellia Florin from the Middle Jurassic of Angren, Uzbekistan // Rev. Palaeobot. Palynol. 2020. V. 281. https://doi.org/10.1016/j.revpalbo.2020.104266
Nosova N. Female reproductive structures of Umaltolepis Krassilov and associated leaves of Pseudotorellia Florin from the Middle Jurassic of East Siberia, Russia // Rev. Palaeobot. Palynol. 2021. V. 289.https://doi.org/10.1016/j.revpalbo.2021.104412
Nosova N., Golovneva L. Phoenicopsis (Leptostrobales) and Pseudotorellia (Ginkgoales) from the Cretaceous of North Asia // Cretaceous Res. 2018. V. 86. P. 149–162.
Pott C., van der Burgh J., van Konijnenburg-van Cittert J.H.A. New ginkgophytes from the Upper Triassic-Lower Cretaceous of Spitsbergen and Edgeøya (Svalbard, Arctic Norway): the history of Ginkgoales on Svalbard // Int. J. Plant Sci. 2016. V. 177. P. 175–197.
Reymanówna M. The Jurassic flora from Grojec near Cracow in Poland, part 1 // Acta Palaeobot. 1963. V. 4. P. 9–48.
Schweitzer H.J., Kirchner M. Die Rhäto-Jurassischen Floren des Iran und Afghanistans. 8. Ginkgophyta // Palaeontographica. 1995. B. 237. P. 1–58.
Shi G., Crane P.R., Herendeen P.S., Ichinnorov N., Takahashi M., Herrera F. Diversity and homologies of corystosperm seed-bearing structures from the Early Cretaceous of Mongolia // J. Systematic Palaeontol. 2019. V. 17. № 12. P. 997–1029.
Wang Z.Q. Plant kingdom. Palaeontological Atlas of North China II Mesozoic. Tianjin Institute of Geology and Mineral Resources. Beijing: Geological Publishing House, 1984. P. 223–296 (in Chinese).
Watson J. A revision of the English wealden flora I: Charales-Ginkgoales // Bull. Br. Mus. Nat. Hist. (Geol.). 1969. V. 17. № 5. P. 209–254.
Zhou Z.Y. Phylogeny and evolutionary trends of Mesozoic ginkgoaleans: a preliminary assessment // Rev. Palaeobot. Palynol. 1991. V. 68. P. 203–216.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция