

Российская археология, 2023, № 1, стр. 53-69
ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ ЖИТЕЛЕЙ ЦЕНТРАЛЬНОГО ПРЕДКАВКАЗЬЯ в I тыс. до н.э. – I тыс. н.э. ПО ДАННЫМ МИТОХОНДРИАЛЬНОЙ ДНК
Д. С. Коробов 1, *, Е. С. Булыгина 2, **, Н. В. Слободова 2, 3, ***, Ф. С. Шарко 2, ****, А. В. Недолужко 4, *****
1 Институт археологии РАН
Москва, Россия
2 Национальный исследовательский центр “Курчатовский институт”
Москва, Россия
3 Национальный исследовательский университет “Высшая школа экономики”
Москва, Росссия
4 Европейский Университет в Санкт-Петербурге
Санкт-Петербург, Россия
* E-mail: dkorobov@mail.ru
** E-mail: eugenia.bulygina@gmail.com
*** E-mail: nv.slobodova@gmail.com
**** E-mail: fedosic@gmail.com
***** E-mail: nedoluzhko@gmail.com
Поступила в редакцию 29.06.2022
После доработки 12.09.2022
Принята к публикации 10.11.2022
- EDN: MBYQLR
- DOI: 10.31857/S0869606323010129
Аннотация
В статье впервые анализируется генетическое разнообразие населения Центрального Предкавказья в эпоху позднего бронзового – раннего железного веков и раннего средневековья. Авторами было проанализировано 120 образцов из 10 могильников кобанской и аланской культуры, а также представителей среднесарматской культуры и северокавказского населения сарматского времени. Получены сведения об изменчивости митохондриальной ДНК (мт-ДНК) 71 индивида. Анализ собранной и опубликованной ранее информации позволяет заключить, что для проанализированных популяций характерно значительное разнообразие мт-ДНК с преобладанием западно-евразийских митохондриальных гаплогрупп. Восточно-евразийские линии наследования прослеживаются в небольшом количестве у населения кобанской культуры (5%), а также у сарматского (14%) и аланского населения (11%). Наиболее распространенными линиями западно-евразийского происхождения являются различные варианты гаплогрупп H, U и N. При этом на всех могильниках наблюдается существенное разнообразие гаплогрупп мт-ДНК, кроме могильника Кич-Малка II, где на протяжении периода с VII в. до н.э. до начала VI в. н.э. фиксируется преемственность индивидов по материнской линии, представленная гаплогруппой H2a2a1, исследования которой требуют более детального подхода с использованием глубокого секвенирования. Сопоставление проанализированной выборки с опубликованными ранее данными о генетическом портрете северокавказского населения раннего и среднего бронзового века позволяет сделать вывод о достаточно позднем проникновении представителей восточно-евразийского кластера гаплогрупп мт-ДНК в Центральное Предкавказье, вероятно, связанное с контактами населения кобанской культуры с кочевым степным населением.
С глубокой древности Кавказ служил своеобразным мостом между Европой и Азией, через который проходили пути миграции многочисленных человеческих сообществ. Это предопределило чрезвычайное этническое, лингвистическое и культурное разнообразие кавказского региона. В последние годы население Кавказа стало своеобразной моделью изучения подобного историко-культурного и генетического разнообразия, в том числе с помощью методов анализа древней ДНК (Sokolov et al., 2016; De Barros Damgaard, Marchi et al., 2018; Wang et al., 2019; Boulygina et al., 2020).
Среди проблем происхождения многочисленных кавказских этносов особое внимание уделяется вопросу об этногенезе северокавказских алан и связанной с ним проблеме “аланского наследия” (Шнирельман, 2006; Афанасьев и др., 2015; Афанасьев, Коробов, 2018; Коробов, 2019). Не вдаваясь в подробности всестороннего освещения этого достаточно сложного вопроса, отметим лишь существующие гипотезы на происхождение аланской культуры с точки зрения археологии.
Аланская археологическая культура вот уже более 130 лет связывается большинством специалистов с широко распространенным на Северном Кавказе обрядом захоронения в катакомбных могильниках, который практиковался около полутора тысяч лет, со II по XIV вв. (Кузнецов, 1962; Ковалевская, 2005. С. 151–166). Подробная история изучения аланских катакомб и обзор существующих точек зрения на их этническую интерпретацию были обобщены в недавно вышедшей монографии С.Н. Савенко (Савенко, 2017. С. 18–42). В настоящее время существуют две основные гипотезы возникновения на Северном Кавказе во II в. н.э. этого яркого обряда, представленного, прежде всего, подкурганными катакомбными захоронениями, сопровождавшими так называемые земляные городища (Малашев, 2007; Габуев, Малашев, 2009. С. 144–150). Согласно первой из них, обряд погребения в Т-образных катакомбах имеет истоки в местных культурах северокавказского населения, испытывавшего значительное влияние ираноязычных кочевников, начиная с раннесарматского времени и особенно в среднесарматскую эпоху (Абрамова, 1995; Габуев, Малашев, 2009. С. 144–162; Малашев, 2016. С. 59–61; 2021; Малашев, Маслов, 2021). Согласно другой гипотезе, подкурганный обряд в катакомбах был целиком привнесен на Северный Кавказ в ходе миграционных процессов переселения носителей аланской археологической культуры из Средней Азии, где выискиваются аналоги данному обряду в древностях джетыасарской и кенкольской культур (Габуев, 1999; 2021; Габуев, Малашев, 2009. С. 106–114). Подробное сопоставление этих древностей с катакомбами северокавказских алан, однако, не позволяет прийти к подобному выводу (Малашев, Торгоев, 2018).
В последнее время получены первые данные о генетическом разнообразии носителей аланской археологической культуры, которые соотносятся с погребенными в катакомбных могильниках Северного Кавказа и Среднего Дона (Афанасьев и др., 2015; Афанасьев, 2018; De Barros Damgaard, Marchi et al., 2018; Коробов, 2019. С. 112–136). Генетическое разнообразие мужской части аланского населения в области Y-хромосомы, передающейся исключительно по мужской линии наследования, демонстрирует, что большинство из погребенных в катакомбных захоронениях I тыс. н.э., как на Северном Кавказе, так и на Среднем Дону, являются обладателями гаплогрупп, характерных для современного северокавказского населения (G2а1, J1 и J2). При этом около трети образцов ДНК относятся к гаплогруппам R1a и Q1b, которые широко встречаются у ираноязычных кочевников Евразии (De Barros Damgaard, Marchi et al., 2018). Таким образом, представляется очевидным смешанный характер аланского населения Северного Кавказа, начиная с его самого раннего этапа (II–IV вв.), в формировании которого принимало активное участие местное субстратное население при безусловном вкладе ираноязычных кочевников, вероятно, близких к среднесарматской культуре.
Для правильного понимания процесса формирования генофонда алан особо актуально изучение генофонда представителей кобанской культуры и их возможных потомков – населения северокавказских равнин и предгорий сарматского времени – как потенциально возможных обладателей тех специфических генетических характеристик кавказского субстрата, которые были зафиксированы у носителей аланской культуры II–XIV вв. (Афанасьев, 2018). Проблема генетического разнообразия местного населения Северного Кавказа, предшествующего появлению алан на равнинах Центрального Предкавказья, еще далека от своего решения. Однако первые шаги были недавно сделаны в ходе анализа древней ДНК представителей кобанской культуры, который проводился в Лаборатории палео- и этногенетики НИЦ “Курчатовский институт” (Boulygina et al., 2020). По результатам исследования 14 образцов ДНК погребенных в могильниках Клин-Яр III и Заюково-3 были получены данные о пяти вариантах Y-хромосомы, из которых два относились к гаплогруппе G2a1. Таким образом, впервые выстраивается линия генетической преемственности некоторых представителей мужской части населения северокавказских равнин и предгорий, начиная с эпохи раннего железного века вплоть до современности.
Митохондриальная ДНК, которая наследуется по материнской линии, не несет такой же разрешающей способности по сравнению с Y-хромосомой (Балановский, 2015. С. 212, 213). Как правило, оценка вклада ее разнообразия в генетический портрет древнего и средневекового населения не позволяет выйти за пределы крупных регионов. Для территории Евразии в данном случае речь идет о характерных особенностях м-т‑ДНК для населения западной и восточной ее части. Тем не менее эта информация представляется весьма важной и интересной при анализе крупных серий образцов, характеризующих население тех или иных территорий Евразии в диахронном ключе (Unterlander et al., 2017). В нашем случае особенно интересно сравнить полученные результаты с имеющимися данными о мт-ДНК носителей сарматских культур, недавно опубликованными коллективом антропологов, археологов и палеогенетиков (Пилипенко и др., 2020).
Картина генетического разнообразия северокавказского населения по мт-ДНК решалась с привлечением широкой серии образцов из могильников, оставленных населением нескольких культурно-хронологических этапов (кобанская культура; носители культурно-хронологических групп Чегем-Манаскент и Подкумок-Хумара, которое далее обозначается нами как северокавказское население сарматского времени; представители среднесарматской культуры; материалы из подкурганных и грунтовых катакомбных могильников Центрального Предкавказья, соотносимых с аланским населением).
Таким образом, в ходе палеогенетического анализа, проведенного в НИЦ “Курчатовский институт”, нами получено значительное количество данных по изменчивости мт-ДНК. Всего было проанализировано 120 образцов из 10 могильников. Сохранность антропологического материала не всегда позволяла извлечь генетический материал, пригодный для амплификации и последующего секвенирования участка D-петли мт‑ДНК (гипервариабильный регион 1, HVR1), поэтому в результате были получены данные по нуклеотидной последовательности HVR1 участка мт-ДНК только для 71 индивида, погребения которых обнаружены в девяти некрополях (рис. 1). Среди них проанализировано 19 образцов представителей кобанской культуры IX–V вв. до н.э., 10 образцов оседлого северокавказского населения первых веков н.э., 6 образцов кочевых носителей среднесарматской культуры того же периода, а также 36 образцов аланского населения преимущественно из захоронений раннего этапа аланской культуры III–IV вв. н.э. и раннего средневековья (вторая пол. V – нач. VI в. н.э.) (табл. 1).
Рис. 1.
Могильники Центрального Предкавказья с проанализированными образцами мт-ДНК: 1 – Клин-Яр III; 2 – Кич-Малка II; 3 – Верхний Куркужин; 4 – Заюково-3; 5 – Киевский I; 6 – Октябрьский I; 7 – Братские 1-е курганы; 8 – Кошкельдинский II; 9 – Айгурский 2.
Fig. 1. Burial grounds of the Central Ciscaucasia with analyzed mtDNA samples: 1 – Klin-Yar III; 2 – Kich-Malka II; 3 – Upper Kurkuzhin; 4 – Zayukovo-3; 5 – Kievsky I; 6 – Oktyabrsky I; 7 – Bratskiye first mounds; 8 – Koshkeldinsky II; 9 – Aygursky 2
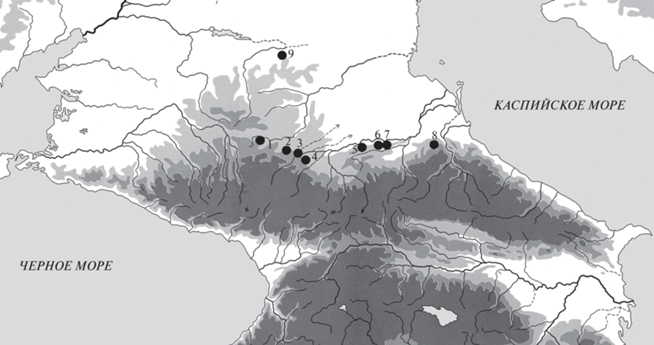
Таблица 1.
Сводные данные об итогах палеогенетического анализа образцов проведенного исследования Table 1. Summarized results of the palaeogenetic analysis of the study samples
| Лаборатор-ный номер | Могильник | Кург./ погр. | Культура | Датировка | Пол гене-тический | мт-ДНК | Y-хромо-сома | Автор раскопок |
|---|---|---|---|---|---|---|---|---|
| AL_1 | Клин-Яр III | 353 | Кобанская | IX–VII вв. до н.э. | XX | H20a | – | А.Б. Белинский, Г. Харке |
| AL_2 | –“– | 355 | –“– | Кон. VIII – 1 пол. VII в.до н.э. | XY | J1c+16261 | R1b | –“– |
| AL_6 | Заюково-3 | 71 | –“– | VIII–VII вв. до н.э. | XX | N | – | А.А. Кадиева, С.В. Демиденко |
| AL_7 | –“– | 72 | –“– | V в. до н.э. | XY | U5a1a1h | G2a1a1a2 | –“– |
| AL_8 | –“– | 79 | –“– | VIII–VII вв. до н.э. | XY | HV1 | D1a2a1~ | –“– |
| AL_9 | –“– | 80 | –“– | VIII–VII вв. до н.э. | XY | T1a | G2a1a | –“– |
| AL_10 | –“– | 81 | –“– | VIII–VII вв. до н.э. | XX | H1e+16129 | – | –“– |
| AL_11 | –“– | 82/1 | –“– | VI–V вв. до н.э. | XY | W5a | R1b1a1b | –“– |
| AL_12 | –“– | 82 | –“– | VI–V вв. до н.э. | XX | R6+16129, H1e+16129, D4a (из-за двух вариабельных позиций) | – | –“– |
| AL_13 | –“– | 91 | Сарматское время | II–III вв. н.э. | XY | H14b1, M3, U4a2b (из-за двух вариабельных позиций) | R1a~ | –“– |
| AL_14 | –“– | 95 | Кобанская | VIII–V вв. до н.э. | XX | R6 | – | –“– |
| AL_15 | –“– | 105 | –“– | VI–V вв. до н.э. | XX | I1 | – | –“– |
| AL_16 | Кич-Малка II | 29/1 | Сарматское время | I–III вв. н.э. | XY | H2a2a1 | Неопр. | Е.Е. Васильева |
| AL_17 | –“– | 29/2 | –“– | I–III вв. н.э. | XX | H2a2a1 | – | –“– |
| AL_18 | –“– | 33/1 | Аланская | 2 пол. V – нач. VI в. н.э. | XY | R8a1a1d | Неопр. | –“– |
| AL_19 | –“– | 33/2 | –“– | 2 пол. V – нач. VI в. н.э. | XX | H2a2a1h | – | –“– |
| AL_20 | –“– | 28 | –“– | 2 пол. V – нач. VI в. н.э. | XX | H2a2a1 | – | –“– |
| AL_21 | –“– | 32 | Кобанская | VII–VI вв. до н.э. | XY | N | Неопр. | –“– |
| AL_22 | –“– | 34/1 | Аланская | 2 пол. V – нач. VI в. н.э. | XY | H1bv1 | Неопр. | –“– |
| AL_23 | –“– | 34/2 | –“– | 2 пол. V – нач. VI в. н.э. | XX | H2a2a1 | – | –“– |
| AL_24 | –“– | 24 | Кобанская | VII–VI вв. до н.э. | XY | Z1a | Неопр. | –“– |
| AL_25 | –“– | 20 | –“– | VII–VI вв. до н.э. | XY | H1bv1 | –“– | –“– |
| AL_26 | –“– | 10 | –“– | VII–VI вв. до н.э. | XY | H2a2a1 | –“– | –“– |
| AL_27 | –“– | 26 | –“– | VII–VI вв. до н.э. | XX | HV0 | – | –“– |
| AL_28 | –“– | 25 | –“– | VII–VI вв. до н.э. | XX | H2a2a1 | – | –“– |
| AL_29 | –“– | 31 | Сарматское время | I–III вв. н.э. | XX | U6a1a | – | –“– |
| AL_30 | –“– | 21 | Кобанская | VII–VI вв. до н.э. | XX | H2a2a1 | – | –“– |
| AL_31 | –“– | 22 | Сарматское время | I–III вв. н.э. | XX | N1b1 | – | –“– |
| AL_32 | Братские 1-е курганы | 12-1 | Аланская | 1 пол. III в. н.э. | ? | N | Неопр. | В.Ю. Малашев |
| AL_33 | –“– | 38 | –“– | Сер. III в. н.э. | XY | HV0 | –“– | –“– |
| AL_34 | –“– | 44/2 | –“– | 2 пол. III в. н.э. | XX? | N | –“– | –“– |
| AL_35 | –“– | 69 | –“– | Сер. III в. н.э. | XX? | N | –“– | –“– |
| AL_36 | –“– | 59 | –“– | 1 пол. III в. н.э. | ? | HV0 | –“– | –“– |
| AL_37 | –“– | 64 | –“– | 1 пол. III в. н.э.? | ? | H1bv1 | –“– | –“– |
| AL_39 | –“– | 1400 | –“– | 1 пол. III в. н.э.? | XY? | V7a / R8a1a1b | –“– | –“– |
| AL_40 | –“– | 1402 | –“– | 1 пол. III в. н.э.? | XY? | H2a2a1 | –“– | –“– |
| AL_41 | –“– | 1436 | –“– | 1 пол. – сер. III в. н.э.? | ? | H14b1 | –“–. | –“– |
| AL_42 | –“– | 1451 | –“– | 3 четв. IV в. н.э.? | ? | N | –“–. | –“– |
| AL_43 | –“– | 1418/2 | –“– | 1 пол. – сер. III в. н.э. | XY? | G1b | –“– | –“– |
| AL_45 | –“– | 1389 | –“– | 1 пол. III в. н.э. | XY? | U5a | –“– | –“– |
| AL_46 | –“– | 1352/2 | –“– | 1 пол. III в. н.э. | XY? | H2a2a1 | –“– | –“– |
| AL_47 | –“– | 1373 | –“– | 1 пол. III в. н.э. | ? | H1bt / N | –“– | –“– |
| AL_48 | –“– | 1474/2 | –“– | 2–3-четв. IV в. н.э. | ? | H14b1 / T2i2 | –“– | –“– |
| AL_49 | –“– | 1456 | –“– | 2–3 четв. IV в. н.э. | ? | T1a | –“– | –“– |
| AL_50 | –“– | 1387 | –“– | 1 пол. III в. н.э.? | ? | N | –“– | –“– |
| AL_51 | –“– | 1392 | –“– | 1 пол. III в. н.э. | ? | HV4b | –“– | –“– |
| AL_52 | –“– | 1396 | –“– | 1 пол. III в. н.э.? | XY | N | –“– | –“– |
| AL_53 | –“– | 1374 | –“– | 1 пол. III в. н.э.? | XX? | T1a | –“– | –“– |
| AL_54 | –“– | 1352/2 | –“– | 1 пол. III в. н.э.? | XY? | M34a1 / N10 | –“– | –“– |
| AL_56 | –“– | 44/1 | –“– | 2 пол. III в. н.э. | ? | HV19 | –“– | –“– |
| AL_57 | –“– | 58 | –“– | 2 пол. III в. н.э. | ? | E1a1b3 | –“– | –“– |
| AL_58 | Кошкель-динский II | 51 | Сарматское время | II – нач. III в. н.э. | XY | J | –“– | Т.Е. Прокофьева |
| AL_61 | –“– | 64 | –“– | II – нач. III в. н.э. | XX | U7 | – | –“– |
| AL_63 | –“– | 52 | –“– | II – нач. III в. н.э. | XX | H13a1a1d | – | –“– |
| AL_66 | –“– | 20 | –“– | II – нач. III в. н.э. | XX | H1e1a5 | – | –“– |
| AL_67 | –“– | 45А | –“– | II – нач. III в. н.э. | XX? | T | Неопр. | –“– |
| AL_69 | Айгурский 2 | 5-1/1 | Среднесарматская | I – нач. II в. н.э | XX | W | – | В.А. Бабенко |
| AL_70 | –“– | 5-1/2 | –“– | I – нач. II в. н.э | XX | W | – | –“– |
| AL_72 | –“– | 10-1 | –“– | I – нач. II в. н.э | XY | H15a1b | Неопр. | –“– |
| AL_75 | –“– | 14-1/2 | –“– | I – нач. II в. н.э | XX? | H2a2b | –“– | –“– |
| AL_76 | –“– | 14-1/3 | –“– | I – нач. II в. н.э | XY | H2a2b | –“– | –“– |
| AL_77 | –“– | 15-1/1 | –“– | I – нач. II в. н.э | XY | H2a+152 | –“– | –“– |
| AL_86 | Верхний Куркужин | 50 | Кобанская | Нач. – сер. VII в. до н.э. | XX | U5a | – | Ю.В. Марченко |
| AL_89 | Октябрьский I | 19/1 | Аланская | 2 пол. III в. н.э. | XY | U7 | Неопр. | В.Ю. Малашев |
| AL_91 | –“– | 23 | –“– | 2 пол. III в. н.э. | XY | H2a+152 16311/H2a2a1 | –“– | –“– |
| AL_92 | –“– | 797 | –“– | 1 пол. III в. н.э. | XY | H5 | –“– | –“– |
| AL_94 | –“– | 807 | –“– | Сер. III в. н.э. | XY | T1a | –“– | –“– |
| AL_101 | –“– | 854/2 | –“– | 2 пол. IV в. н.э. | XY | I1 | –“– | –“– |
| AL_110 | Киевский I | 769 | –“– | 2 пол. IV в. н.э. | XY | U4a2h1 | –“– | –“– |
| AL_111 | –“– | 725 | –“– | 2 пол. IV в. н.э. | XY | M24b | –“– | –“– |
| AL_115 | –“– | 786 | –“– | 2 пол. IV в. н.э. | XY | M6 | –“– | –“– |
Кобанские образцы происходят из погребений могильников Клин-Яр III, Заюково-3, Кич-Малка II и Верхний Куркужин (Белинский, Дударев, 2015. С. 242–244; Belinskij, Harke, 2018. P. 9–19, 284–286; Кадиева, Демиденко, 2017; Кадиева, 2021; Васильева, 2009; 2010; Марченко, 2018). Это сравнительно близко расположенные памятники, происходящие с территории Кисловодской котловины и прилегающей к ней с востока территории бассейна рек Малки и Баксана с притоками. Расположение могильников с погребениями северокавказского населения сарматского времени гораздо шире в пространстве. Половина из них происходит с упоминаемых выше могильников Заюково-3 и Кич-Малка II; вторая половина относится к недавно открытому могильнику Кошкельдинский II на территории равнинной Чечни (Прокофьева, 2018). Среднесарматские погребения, участвующие в анализе, были раскопаны на курганном могильнике Айгурский 2 на территории Ставропольского края (Бабенко, Березин, 2009). Наконец, большинство аланских погребений происходит с недавно исследованных курганных могильников III–IV вв. н.э. Братские 1‑е курганы, Октябрьский I и Киевский I (Малашев и др., 2018; 2020) (рис. 1). Кроме того, в анализе фигурируют материалы пяти грунтовых катакомбных захоронений могильника Кич-Малка II эпохи раннего средневековья (Васильева, 2012; Васильева, Ахмедов, 2015).
Работы по выделению древней ДНК из антропологического материала (зубы) представителей кобанской и аланской культуры, а также представителей среднесарматской культуры и северокавказского населения сарматского времени проводились в специально оборудованном комплексе чистых помещений с соблюдением всех необходимых для подобных экспериментов условий. В частности, были использованы комплекты спецодежды для чистых помещений, проводилась обработка рабочих поверхностей и приборов реагентом DNAAWAY (Thermo Fisher Scientific, США) и ультрафиолетом, на всех стадиях эксперимента присутствовали контрольные образцы, кроме того, анализировалась ДНК у всех сотрудников, осуществлявших работы в палеогенетической лаборатории. Результаты показали, что гаплогруппы мт-ДНК, выявленные у проанализированных образцов, не совпадали с таковыми у исследователей.
Выделение ДНК из зубов проводили по стандартной методике (Orlando et al., 2011) с некоторыми модификациями, включая двукратное уменьшение реакционного объема во время депротеинизации костной муки. Для амплификации HVR1 участка D-петли мт-ДНК использовали праймерные системы, предложенные ранее (Sampietro et al., 2005) и разработанные авторами исследования. Последовательность полученных ПЦР-фрагментов (длиной от 138 до 210 пар нуклеотидов), покрывающих HVR1 участок D-петли мт-ДНК, прочитывали, используя секвенатор ABI 3730xl (ThermoFisher Scientific, США). ДНК последовательности HVR1 участка D-петли депонированы в международную базу данных NCBI под номерами SAMN30702111–SAMN30702181 (проект PRJNA797283). Нуклеотидные последовательности анализировали и выравнивали на D‑петлю кембриджской эталонной последовательности мт-ДНК (NC_012920.1, rCRS) при помощи программы BioEdit. Программа mtDNAprofiler позволила получить список нуклеотидных отличий от референса, который затем был использован для определения митохондриальных гаплотипов людей, используя HaploGrep2 (Weissensteiner et al., 2016).
Сеть гаплотипов для последовательностей HVR1 образцов из данного исследования, а также образцов из данного региона и исторического периода, опубликованных ранее (табл. 2), была построена с использованием программы Pegas в статистической среде R.
Таблица 2.
Образцы из ранее опубликованных работ, задействованные для построения дерева гаплотипов, используя гипервариабельный участок митохондриального генома (HVR1) Table 2. Samples from previously published works used to construct a haplotype tree with the hypervariable region of mitochondrial genome (HVR1)
| Маркиров- ка образца | Происхожде- ние образца | Эпоха/архео- логическая культура | Митохон- дриальная гаплогруппа | Метод | Ссылка |
|---|---|---|---|---|---|
| SH_1 | Адыгея, Шушук 75, погр. 2, образец совпадает с образцом SH_5 | Средняя бронза | H1a | Секвенирование HVR1 | Erlikh et al., 2021 |
| SH_5 | Адыгея, Шушук 75, погр. 2, образец совпадает с образцом SH_1 | –“– | H1a | –“– | –“– |
| SH_4 | Адыгея, Шушук 75, погр. 2 | –“– | H1a | –“– | –“– |
| SH_2 | Адыгея, Шушук 75, погр. 2 | –“– | H1a | –“– | –“– |
| SH_3 | Адыгея, Шушук 75, погр. 2 | –“– | H1a | –“– | –“– |
| SH_6 | Адыгея, Шушук 75, погр. 2 | –“– | H1a | –“– | –“– |
| SH_7 | Адыгея, Шушук 42, погр. 1 | –“– | H2a2a1 | –“– | –“– |
| SH_9 | Адыгея, Шушук 42, погр. 1 | –“– | H5 | –“– | –“– |
| SH_12 | Адыгея, Шушук 49, погр. 1 | –“– | H1aj1 | –“– | –“– |
| PG2004 | Ставропольский край, Прогресс-2, кург. 4, погр. 9 | Энеолит | H2 | Полногеномное секвени-рование | Wang et al., 2019 |
| LYG001 | Ставропольский край, Лысогорская 6, кург. 3 погр. 4 | Средняя бронза/ северокавказская | H13a1a2 | –“– | –“– |
| AY2003 | Ставропольский край, Айгурский-2, кург. 22, погр. 9 | Ранняя бронза/ майкопская | H2a1 | –“– | –“– |
| I2051 | Краснодарский край, Марченкова гора, дольмен 13 | Поздняя бронза/ дольменная | H6a1a2a | –“– | –“– |
| DA144 | Ростовская обл., Чеботарев V, кург. 6, погр. 1 | РЖВ/среднесарматская | H28 | –“– | De Barros Dam-gaard, Marchi et al., 2018 |
| DA162 | Северная Осетия, Бесланский, кат. 439 | РЖВ/аланская | H13a2c | –“– | –“– |
| DA164 | Северная Осетия, Змейский, кат. 182, погр. 2 | Средневековье/ аланская | H5 | –“– | –“– |
| DA191 | Венгрия, Тисасолош-Чаланьсег, погр. 19 | РЖВ/скифская | H2a2 | –“– | –“– |
| DA194 | Венгрия, Шандорфальва-Эперьеш, погр. 118 | –“– | H2a2a1 | –“– | –“– |
| DA195 | Венгрия, Шандорфальва-Эперьеш, погр. 125 | –“– | H+16311 | - “ - | –“– |
| DA197 | Венгрия, Шандорфальва-Эперьеш, погр. 168 | –“– | H2a2a | - “ - | –“– |
| DA198 | Венгрия, Шандорфальва-Эперьеш, погр. 143 | –“– | H7a1 | - “ - | –“– |
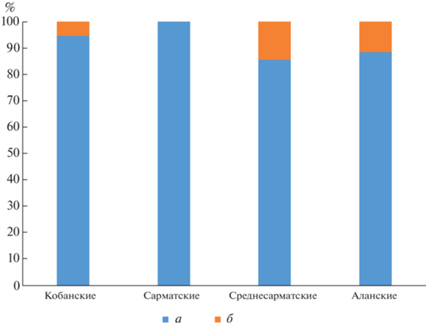
Суммируя полученные нами результаты с опубликованными ранее сведениями о мт-ДНК представителей аланского и сарматского населения (De Barros Damgaard, Marchi et al., 2018), можно довести число исследованных образцов до 90, что позволяет увидеть следующие тенденции. Прежде всего, следует отметить, что 91% выделенных гаплогрупп митохондриальной ДНК предсказуемо относится к западно-евразийским гаплогруппам (82 из 90), на долю восточно-евразийских приходится 9%. Однако распределение этих гаплогрупп по популяциям разных культурно-хронологических периодов неравномерно. Так, среди 19 проанализированных представителей кобанской культуры вклад единственной отмеченной восточно-евразийской гаплогруппы Z составляет около 5% (табл. 3; рис. 2). Среди северокавказского населения сарматской эпохи восточно-евразийские гаплогруппы не отмечены. Все 10 индивидов обладали западно-евразийскими гаплогруппами митохондриальной ДНК. Вклад представителей восточно-евразийских популяций по материнской линии в генетическое разнообразие кочевников среднесарматской культуры был несколько выше, чем у кобанского населения – он прослеживается у двух индивидов из 14, что составляет около 14%. В обоих случаях речь идет об одном субкладе гаплогруппы А, отмеченной у погребенных в курганном могильнике Несветай II на Нижнем Дону (De Barros Damgaard, Marchi et al., 2018. Supplementary Table 8 , образцы DA136 и DA141). Близкие результаты в процентном отношении получены у самой многочисленной из проанализированных групп населения – представителей аланской культуры, в основном раннего ее этапа, где восточно-евразийские гаплогруппы отмечены у 5 из 47 индивидов (11%). При этом здесь имеется некоторое разнообразие – в единичных случаях встречены гаплогруппы E и G, в трех случаях – гаплогруппа M (табл. 3). Все они относятся к наиболее ранним из проанализированных подкурганным погребениям III–IV вв. н.э. могильников Киевский I и Братские 1-е курганы. Таким образом, ощутимый вклад женского населения Восточной Евразии в формирование северокавказских алан представляется очевидным, а близкие в процентном отношении, хотя и единичные случаи встречаемости восточно-евразийских гаплогрупп митохондриальной ДНК у среднесарматского населения степей Предкавказья могут быть неслучайными.
Таблица 3.
Распределение гаплогрупп митохондриальной ДНК по объединенной выборке погребений разных культурно-хронологических групп (проанализированных авторским коллективом и опубликованных в: de Barros Damgaard, Marchi et al., 2018) Table 3. Distribution of mitochondrial DNA haplogroups in the combined sample of burials belonging to different cultural and chronological groups (analyzed by the authors and published in: de Barros Damgaard, Marchi et al., 2018)
| Гаплогруппы | Кобанская культура | Сарматское время | Среднесарматская культура | Аланская культура | Всего | % |
|---|---|---|---|---|---|---|
| Западно-евразийские гаплогруппы | ||||||
| H | 6 | 5 | 5 | 14 | 30 | 33.3 |
| HV | 2 | 0 | 0 | 5 | 7 | 7.8 |
| I | 1 | 0 | 1 | 2 | 4 | 4.4 |
| J | 1 | 1 | 1 | 2 | 5 | 5.6 |
| N | 2 | 1 | 0 | 6 | 9 | 10.0 |
| R | 2 | 0 | 0 | 1 | 3 | 3.3 |
| T | 1 | 1 | 0 | 3 | 5 | 5.6 |
| U | 2 | 2 | 3 | 6 | 13 | 14.4 |
| V | 0 | 0 | 0 | 1 | 1 | 1.1 |
| W | 1 | 0 | 2 | 1 | 4 | 4.4 |
| X | 0 | 0 | 0 | 1 | 1 | 1.1 |
| Восточно-евразийские гаплогруппы | ||||||
| A | 0 | 0 | 2 | 0 | 2 | 2.2 |
| E | 0 | 0 | 0 | 1 | 1 | 1.1 |
| G | 0 | 0 | 0 | 1 | 1 | 1.1 |
| M | 0 | 0 | 0 | 3 | 3 | 3.3 |
| Z | 1 | 0 | 0 | 0 | 1 | 1.1 |
Рис. 2.
Процентное распределение исследованных образцов по западно-евразийским (а) и восточно-евразийским (б) субкластерам мт-ДНК.
Fig. 2. Percentage distribution of the studied samples by the West Eurasian (а) and East Eurasian (б) mtDNA subclusters
Разумеется, преждевременно говорить о непосредственном вкладе восточно-евразийских популяций в формирование как среднесарматского, так и аланского населения, без более глубокого анализа субкладов мт-ДНК, поскольку рассматриваемые субклады могли проникнуть на территории степного и равнинного Предкавказья задолго до появления здесь среднесарматского или аланского населения, например, в скифскую эпоху (Пилипенко и др., 2020. С. 27). На востоке Евразии некоторые рассматриваемые субклады мт-ДНК появились значительно раньше. Так, субклад Z1a, отмеченный у представителя кобанской культуры, прослежен у энеолитического населения ботайской культуры на территории Казахстана, а субклад А + 152 + 16362, прослеженный у погребенных в могильнике среднесарматской культуры Несветай II, обнаружен в Южной Сибири в эпоху энеолита и раннего бронзового века (De Barros Damgaard, Martiniano et al., 2018).
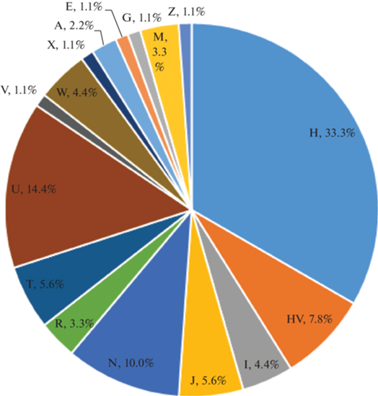
Среди западно-евразийских гаплогрупп мт‑ДНК около трети относится к наиболее распространенной на европейской территории гаплогруппе H (33.3%). С учетом предковой для нее гаплогруппы HV, эта доля может быть увеличена до 41.1%. Далее по значимости стоит гаплогруппа U, к которой относится 14.4% участвовавших в анализе индивидов, и гаплогруппа N – 10%. По 4.4–5.6% приходится на гаплогруппы I, J, T и W; остальные гаплогруппы (R, V и X) встречены в единичных случаях (табл. 3; рис. 3).
Рис. 3.
Процентное распределение исследованных образцов по гаплогруппам мт-ДНК.
Fig. 3. Percentage distribution of the studied samples by mtDNA haplogroups
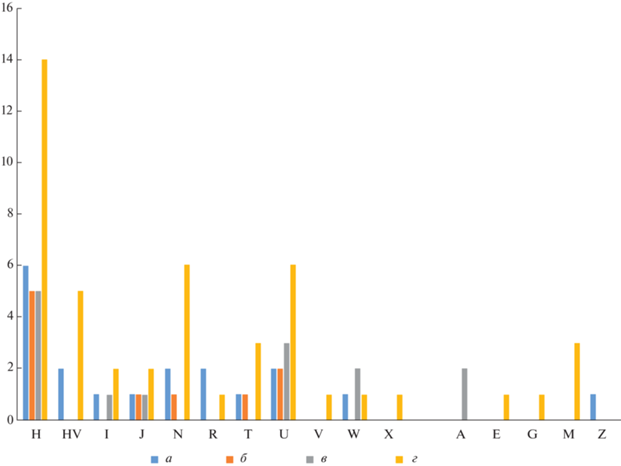
Как правило, на одном могильнике присутствует значительное разнообразие гаплогрупп мт-ДНК (табл. 1; рис. 4). Исключение наблюдается на могильнике Кич-Малка II, где в восьми погребениях отмечено присутствие гаплогруппы H2a2a1, которая отмечена у представителей трех культурно-хронологических групп населения – кобанского (3), сарматского времени (2) и аланского (3). Субклад H2a, в том числе его разновидность H2a2a1, встречался также в двух аланских погребениях могильника Братские 1-е курганы, трех среднесарматских погребениях Айгурского 2-го курганного могильника и в одном аланском захоронении в подкурганной катакомбе могильника Октябрьский I (табл. 1). Таким образом, данный субклад присутствует у 14 индивидов (это чуть менее половины от всех проанализированных индивидов с гаплогруппой H), из которых три относятся к кобанской культуре (18.8%), два – к сарматскому времени (12.5%), три – к среднесарматскому населению (18.8%) и шесть – к аланскому (37.5%). Такая устойчивость во времени в передаче материнской мт-ДНК может говорить о преемственности населения, по крайней мере, в его женской части, как минимум для ряда археологических памятников (прежде всего, могильника Кич-Малка II) и, возможно, для формирования рассматриваемых культур в целом. В то же время важно отметить ограниченную разрешающую способность использованного метода, поскольку анализ ряда образцов (в основном из могильника Кич-Малка II) продемонстрировал полное совпадение нуклеотидной последовательности использованного HVR1 региона с таковым у кембриджской эталонной последовательности мт-ДНК, что может предполагать потенциальное единое происхождение по материнской линии, но требует использования методов глубокого секвенирования генома.
Рис. 4.
Диахронное распределение исследованных образцов по гаплогруппам мт-ДНК (а – кобанские, б – сарматского времени, в – среднесарматские, г – аланские).
Fig. 4. Diachronic distribution of the studied samples by mtDNA haplogroups (а – Koban, б – Sarmatian period, в – Middle Sarmatian, г – Alan)
Некоторые из выявленных субкладов имеют ближневосточное происхождение и нередко встречаются на Северном Кавказе (Пилипенко и др., 2020. С. 26). К ним относятся, например, субклады H20a и U7, встреченные в единичных случаях в захоронениях кобанской культуры (Клин-Яр III), северокавказского населения сарматского времени (Кошкельдинский II) и ранних алан (Октябрьский I) (табл. 1).
С этой точки зрения любопытно сравнить данные о мт-ДНК анализируемой здесь выборки с недавно опубликованными сведениями о генетическом портрете населения степей и предгорий Северного Кавказа в эпоху ранней и средней бронзы (Wang et al., 2019). Следует отметить, что среди 55 проанализированных погребений с известными сведениями о мт-ДНК носителей восточно-евразийских линий материнского наследования встречено не было. Западно-евразийские линии представлены наиболее широко вариантами гаплогруппы U (23, или 41.8%), R (8, или 14.5%) и T (7, или 12.7%). Присутствуют также минорные варианты гаплогрупп H, HV, I, J, K, W и X (Wang et al., 2019. Supplementary Data 1). Кроме того, разнообразные варианты гаплогруппы H описаны для захоронений среднего бронзового века археологического комплекса Шушук (Республика Адыгея). Более того, анализ пяти индивидов из захоронения 75 предполагает их потенциальное родство по материнской линии (E-rlikh et al., 2021). Таким образом, очевидно, что доминирование материнской гаплогруппы U в раннем и среднем бронзовом веке, которая наблюдается у представителей куро-аракской, майкопской, новосвободненской, ямной, катакомбной, северокавказской и дольменной культур, сменяется в позднем бронзовом веке на преобладание наследования мт-ДНК по линии гаплогруппы H, отмеченной в более раннее время в единичных случаях.
Сравнительный анализ последовательностей HVR1 из образцов, несущих гаплогруппу H, представленных в данной и ранее опубликованных работах, указывает на их значительную близость (рис. 5). В то же время все представленные здесь признаки потенциального родства являются лишь предварительными и требуют более детального и сложного исследования с использованием анализа полного митохондриального и ядерного геномов.
Рис. 5.
Филогенетическая сеть гаплотипов гипервариабельного региона 1 мт-ДНК (HVR1) у представителей археологических культур бронзового и железного веков на Северном Кавказе, несущих гаплогруппу H. Условные обозначения: Поперечные линии указывают на количество отличительных замен в ДНК-последовательности HVR1 между субгаплотипами. Субгаплотипы, совпадающие у нескольких образцов, отмечены буквами: А – (AL_16, AL_20, AL_23, AL_26, AL_28, AL_30, AL_40, AL_46, AL_47, LYG001, PG2004); B – (AL_33, AL_36); C – (DA191, DA4, SH_9); D – (SH_1, SH_5); E – (DA144, SH_12); F – (AL_22, AL_25, AL_37); G – (AL_77, DA195); H – (AL_13, AL_41, AL_48, AL_51); I – (AL_75, AL_76); J – (AL_17, AY2003). Маркировка археологических образцов: а– кобанская культура; б – сарматское время; в – аланская культура; г – среднесарматская культура; д – образцы из могильника Шушук (Адыгея); е – другие образцы из ранее опубликованных работ. Расшифровка происхождения образцов представлена в табл. 2.
Fig. 5. Phylogenetic network of haplotypes of the mtDNA hypervariable region 1 (HVR1) in haplogroup H carriers of the Bronze and Iron Age archaeological cultures of the North Caucasus
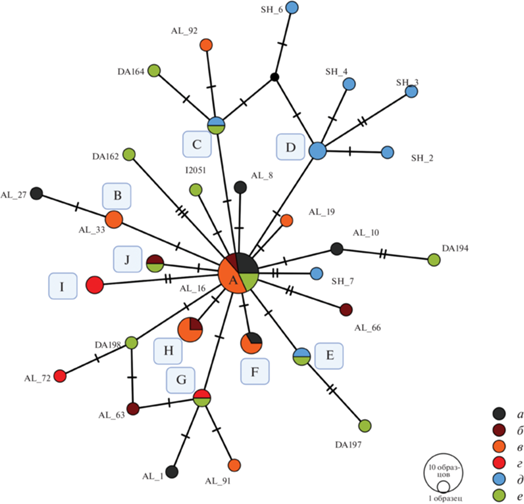
Различные варианты гаплогруппы H доминируют и в проанализированных коллективом исследователей захоронениях сарматского населения Нижнего Поволжья, где они составляют в совокупности 22.6%. Далее по частоте встречаемости стоят субкластеры гаплогруппы Т (17.7%) и U (около 10%) (Пилипенко и др., 2020. С. 24). В целом следует отметить близкий характер генетического разнообразия мт-ДНК у анализируемого нами населения с данными о сарматах Нижнего Поволжья. Среди последнего также присутствует относительно небольшое количество представителей восточно-евразийских линий наследования мт-ДНК, представленных вариантами гаплогрупп A, C, F и G, тогда как от 80 до 90% проанализированных индивидов относились к западно-евразийским вариантам мт-ДНК (Пилипенко и др., 2020. С. 24. Рис. 3). Учитывая упомянутое выше наблюдение об отсутствии прослеженного влияния восточно-евразийских линий наследования мт-ДНК у северокавказского населения степи и предгорий в эпоху ранней и средней бронзы, можно предположить, что данное влияние появляется в позднем бронзовом – раннем железном веке и может быть получено в ходе контактов представителей кобанской культуры с кочевым степным населением.
Исследование генетического разнообразия северокавказского населения древности и средневековья делает свои первые шаги, поэтому полученные нами выводы следует рассматривать как сугубо предварительные. Дальнейшее накопление данных о генетическом портрете как представителей аланской культуры, так и их предшественников поможет уточнить и, возможно, пересмотреть уже имеющиеся представления о путях возникновения и развития аланского этноса.
Авторы выражают благодарность руководителям археологических раскопок, давшим возможность использовать материалы из захоронений: А.Б. Белинскому, Г. Харке, А.А. Кадиевой, С.В. Демиденко, В.Ю. Малашеву, Е.Е. Васильевой, Т.Е. Прокофьевой, В.А. Бабенко, Ю.В. Марченко. Анализ антропологических материалов и отбор образцов производились А.П. Бужиловой, М.Б. Медниковой, М.В. Добровольской, Т.Ю. Шведчиковой, С.Ю. Фризеном, И.К. Решетовой, Е.В. Перервой, Е.Ф. Батиевой, Е.В. Пугачевой. Статья подготовлена в рамках выполнения темы НИР ИА РАН “Междисциплинарный подход в изучении становления и развития древних и средневековых антропогенных экосистем” (№ НИОКТР 122011200264-9). Генетическое исследование образцов было поддержано грантом РФФИ № 18-00-00399.
Список литературы
Абрамова М.П. Катакомбные могильники III–V вв. н.э. центральных районов Северного Кавказа // Аланы: история и культура. Владикавказ: Северо-Осетинский ин-т гуманитар. исслед., 1995 (Alanica; III). С. 65–78.
Афанасьев Г.Е. Некоторые дополнения к исторической интерпретации новых генетических исследований сармато-аланских образцов // Кавказ в системе культурных связей Евразии в древности и средневековье. XXX Крупновские чтения по археологии Северного Кавказа: материалы Междунар. науч. конф. / Отв. ред. У.Ю. Кочкаров. Карачаевск: Карачаево‑Черкесский гос. ун-т, 2018. С. 284–289.
Афанасьев Г.Е., Добровольская М.В., Коробов Д.С., Решетова И.К. Новые археологические, антропологические и генетические аспекты в изучении донских алан // Краткие сообщения Института археологии. 2015. Вып. 237. С. 64–79.
Афанасьев Г.Е., Коробов Д.С. Северокавказские аланы по данным палеогенетики // Этногенез и этническая история народов Кавказа: сб. материалов I Междунар. нахского науч. конгр. (г. Грозный. 11–12 сентября 2018 г.) / Отв. ред. Ш.А. Гапуров, С.С. Магамадов. Грозный: Грозненский рабочий, 2018. С. 180–191.
Бабенко В.А., Березин Я.Б. Сарматские погребения могильников Айгурский 2 и Барханчак 2 (северное Ставрополье) // Материалы по изучению историко-культурного наследия Северного Кавказа. Вып. IX. Археология, краеведение / Отв. ред. А.Б. Белинский. Ставрополь: Наследие, 2009. С. 279–320.
Балановский О.П. Генофонд Европы. М.: Т-во науч. изданий КМК, 2015. 354 с.
Белинский А.Б., Дударев С.Л. Могильник Клин-Яр III и его место среди древностей Кавказа и Юго-Восточной Европы начала эпохи раннего железа. Ставрополь: Дизайн-студия Б, 2015. 446 с.
Васильева Е.Е. Могильник Кичмалка II – новый памятник кобанской культуры на Северном Кавказе // Лавровский сборник: материалы XXXIII Среднеазиатско-Кавказских чтений, 2008–2009 гг. Этнология, история, археология, культурология. К столетию со дня рождения Л.П. Лаврова. СПб.: Музей антропологии и этнографии РАН, 2009. С. 42–44.
Васильева Е.Е. Исследования кобанского могильника Кичмалка II в Кабардино-Балкарии // Проблемы хронологии и периодизации археологических памятников и культур Северного Кавказа. XXVI Крупновские чтения по археологии Северного Кавказа: тез. докл. Междунар. науч. конф. / Отв. ред. М.Б. Мужухоев. Магас: Пилигрим, 2010. С. 80–82.
Васильева Е.Е. Погребальный комплекс аланской культуры на Северном Кавказе. Вторая половина V – первая половина VI века н.э. // Кочевники Евразии на пути к империи: Из собрания Государственного Эрмитажа: каталог выставки. СПб.: Славия, 2012. С. 176–179.
Васильева Е.Е., Ахмедов И.Р. Новое погребение аланской знати постгуннского времени из Кабардино-Балкарии // Социальная стратификация населения Кавказа в конце античности и начале средневековья: археологические данные: материалы междунар. науч. конф. (Сухум, 31 мая – 5 июня 2015 г.) / Отв. ред. А.В. Мастыкова. М.: ИА РАН, 2015. С. 13–16.
Габуев Т.А. Ранняя история алан (по данным письменных источников). Владикавказ: Иристон, 1999. 148 с.
Габуев Т.А. О дискуссионной проблеме сложения и распространения раннеаланской культуры на Северном Кавказе // Эпоха всадников на Северном Кавказе: к 90-летию Веры Борисовны Ковалевской / Отв. ред. З.Х. Албегова, Д.С. Коробов, А.В. Мастыкова. М.: ИА РАН, 2021. С. 143–150.
Габуев Т.А., Малашев В.Ю. Памятники ранних алан центральных районов Северного Кавказа. М.: Т-АУС, 2009. 468 с.
Кадиева А.А. Погребальный обряд населения Баксанского ущелья предскифского времени (по материалам могильника Заюково-3) // Эпоха всадников на Северном Кавказе: к 90-летию Веры Борисовны Ковалевской / Отв. ред. З.Х. Албегова, Д.С. Коробов, А.В. Мастыкова. М.: ИА РАН, 2021. С. 111–124.
Кадиева А.А., Демиденко С.В. Раскопки комплекса археологических памятников близ селения Заюково (Кабардино-Балкарская Республика) // Вестник Российского фонда фундаментальных исследований. Гуманитарные и общественные науки. 2017. № 2 (87). С. 164–171.
Ковалевская В.Б. Кавказ – скифы, сарматы, аланы. I тыс. до н.э. – I тыс. н.э. М.: ИА РАН, 2005. 398 с.
Коробов Д.С. Аланы Северного Кавказа: этнос, археология, палеогенетика. М.; СПб.: Нестор-История, 2019. 156 с.
Кузнецов В.А. Аланские племена Северного Кавказа. М.: Изд-во АН СССР, 1962 (Материалы и исследования по археологии СССР; № 106). 164 с.
Малашев В.Ю. Культурная ситуация в центральных районах Северного Кавказа во II–IV вв. н.э. // Три четверти века. Д.В. Деопику – друзья и ученики / Отв. ред. Н.Н. Бектимирова. М.: Памятники исторической мысли, 2007. С. 487–501.
Малашев В.Ю. Памятники среднесарматской культуры северокавказских степей и их традиции в курганных могильниках северо-восточного Кавказа второй половины II – середины V в. М.: ИА РАН, 2016. 202 с.
Малашев В.Ю. Памятники типа Подкумок-Хумара // Эпоха всадников на Северном Кавказе: к 90-летию Веры Борисовны Ковалевской / Отв. ред. З.Х. Албегова, Д.С. Коробов, А.В. Мастыкова. М.: ИА РАН, 2021. С. 125–142.
Малашев В.Ю., Магомедов Р.Г., Дзуцев Ф.С., Мамаев Х.М., Кривошеев М.В. Охранно-спасательные исследования могильника “Братские 1-е курганы” на территории Чеченской Республики в 2018 г. // История, археология и этнография Кавказа. 2018. Т. 14. № 4. С. 195–206.
Малашев В.Ю., Магомедов Р.Г., Дзуцев Ф.С., Мамаев Х.М., Кадзаева З.П. Охранно-спасательные исследования могильников раннего этапа аланской культуры на Среднем Тереке Октябрьский I и Киевский I в Моздокском районе Республики Северная Осетия-Алания в 2019 г. // История, археология и этнография Кавказа. 2020. Т. 16. № 2. С. 439–460.
Малашев В.Ю., Маслов В.Е. Курганы-кладбища центральных и восточных районов Северного Кавказа III в. до н.э. – начала (первой половины) II в. н.э. (памятники типа Чегем-Манаскент) // Нижневолжский археологический вестник. 2021. Т. 20. № 2. С. 81–132.
Малашев В.Ю., Торгоев А.И. Т-образные катакомбы сарматского времени Северного Кавказа и Средней Азии // Российская археология. 2018. № 4. С. 36–52.
Марченко Ю.В. Отчет Северокавказской археологической экспедиции Государственного Эрмитажа о проведении раскопок грунтового могильника в селе Верхний Куркужин Кабардино-Балкарской республики. 2018 // Архив Института археологии РАН. Р-1. № 62231, 62232.
Пилипенко А.С., Черданцев С.В., Трапезов Р.О., Томи-лин М.А., Балабанова М.А., Пристяжнюк М.С., Журавлев А.А. К вопросу о генетическом составе сарматского населения Нижнего Поволжья (данные палеогенетики) // Вестник Волгоградского государственного университета. Серия 4: История. Регионоведение. Международные отношения. 2020. Т. 25. № 4. С. 17–50.
Прокофьева Т.Е. Отчет об археологических раскопках грунтового могильника “Кошкельдинский II” в Гудермесском районе Чеченской Республики в 2018 г. // Архив Института археологии РАН. Р-1. № 70341, 70342.
Савенко С.Н. Характеристика социального развития аланского общества Северного Кавказа по материалам катакомбных могильников X–XII вв. н.э. Пятигорск; Казань: Казанская недвижимость, 2017. 384 с.
Шнирельман В.А. Быть аланами: интеллектуалы и политика на Северном Кавказе в XX веке. М.: Новое литературное обозрение, 2006. 696 с.
De Barros Damgaard P., Marchi N. et al. 137 ancient human genomes from across the Eurasian steppes [Электронный ресурс] // Nature. 2018. № 557. P. 369–374. URL: https://www.nature.com/articles/s41586-018-0094-2 (дата обращения: 13.11.2022).
De Barros Damgaard P., Martiniano R. et al. The first horse herders and the impact of early Bronze Age steppe expansions into Asia [Электронный ресурс] // Science. 2018. Vol. 360. 7711. URL: https://www.science.org/doi/10.1126/science.aar7711 (дата обращения: 13.11.2022).
Belinskij A.B., Härke H. Ritual, society and population at Klin-Yar (North Caucasus). Excavations 1994–1996 in the Iron Age to early medieval cemetery. Bonn: Dr. Rudolf Habelt GmbH, 2018 (Archäologie in Eurasien; 36). 443 p.
Boulygina E. et al. Mitochondrial and Y-chromosome diversity of the prehistoric Koban culture of the North Caucasus [Электронный ресурс] // Journal of Archaeological Science: Reports. 2020. Vol. 31. 102357. URL: https: //www.sciencedirect.com/science/article/abs/pii/S2352409X20301486 (дата обращения: 13.11.2022).
Erlikh V.R. et al. Potential maternal kinship among humans from the Northern Caucasus “post-dolmen” burials [Электронный ресурс] // Journal of Archaeological Science: Reports. 2021. Vol. 39. 103198. URL: https://www.sciencedirect.com/science/article/pii/S2352409X21004107 (дата обращения: 13.11.2022).
Orlando L. et al. True single-molecule DNA sequencing of a Pleistocene horse bone [Электронный ресурс] // Genome Research. 2011. Vol. 21. № 10. P. 1705–1719. URL: https://genome.cshlp.org/content/21/10/1705 (дата обращения: 13.11.2022).
Sampietro M.L. et al. The genetics of the pre-Roman Iberian Peninsula: A mtDNA study of ancient Iberians [Электронный ресурс] // Annals of Human Genetics. 2005. Vol. 69. № 5. P. 535–548. URL: https://onlinelibrary.wiley.com/doi/10.1111/j.1529-8817.2005.00194.x (дата обращения: 13.11.2022).
Sokolov A.S. et al. Six complete mitochondrial genomes from Early Bronze Age humans in the North Caucasus [Электронный ресурс] // Journal of Archaeological Science. 2016. Vol. 73. P. 138–144. URL: https://www.sciencedirect.com/science/article/abs/pii/S0305440316301091 (дата обращения: 13.11.2022).
Unterlander M. et al. Ancestry, Demography, and Descendants of Iron Age Nomads of the Eurasian Steppe [Электронный ресурс] // Nature Communications. 2017. Vol. 8. 14615. URL: https://www.nature.com/articles/ncomms14615 (дата обращения: 13.11.2022).
Wang C.C. et al. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions [Электронный ресурс] // Nature Communications. 2019. Vol. 10. 590. URL: https://www.nature.com/articles/s41467-018-08220-8 (дата обращения: 12.11.2022).
Weissensteiner H. et al. HaploGrep 2: mitochondrial haplogroup classification in the era of high-throughput sequencing [Электронный ресурс] // Nucleic Acids Research. 2016. Vol. 44, W1. P. W58–W63. URL: https://academic.oup.com/nar/article/44/W1/W58/2499296 (дата обращения: 13.11.2022).
Дополнительные материалы отсутствуют.
Инструменты
Российская археология