

Российская археология, 2022, № 4, стр. 63-76
Археологический навоз как источник данных о системе питания сельскохозяйственных животных на Северном Кавказе во II–IV вв. н.э. (по материалам Киевского городища)
А. Ю. Сергеев 1, *, А. Н. Бабенко 1, **, Д. А. Куприянов 1, ***, Д. С. Коробов 1, ****
1 Институт археологии РАН
Москва, Россия
* E-mail: alexarchbot@yandex.ru
** E-mail: mnemosina_a@mail.ru
*** E-mail: dmitriykupriyanov1994@yandex.ru
**** E-mail: dkorobov@mail.ru
Поступила в редакцию 05.03.2022
После доработки 05.03.2022
Принята к публикации 14.06.2022
- EDN: PEESDD
- DOI: 10.31857/S0869606322040146
Аннотация
Остатки навоза, сохраняющиеся в культурном слое, могут нести уникальную информацию о содержании и питании домашних животных, хозяйственной деятельности населения и природных условиях. В статье рассматриваются результаты комплексного археоботанического анализа образцов из навозной прослойки культурного слоя раннеаланского Киевского городища II–IV вв. н.э. (Северный Кавказ). Изучение растительных макро- (зерна, семена, мякина и древесный уголь) и микроостатков (споры, пыльца и непыльцевые палиноморфы) позволило установить тафономию и видовую принадлежность навоза (конский навоз сохранился in situ после сгорания), определить осенне-зимний период накопления навоза и систему кормления лошадей в зимний период. Животные содержались на подножном корме с фуражной подкормкой. Кроме того, наличие культурных злаков и их мякины в образце говорит о выращивании раннеаланским населением проса, пшеницы и ячменя в округе городища.
Важным носителем информации о корме сельскохозяйственных животных в древности являются остатки навоза, обнаруженные непосредственно в археологических контекстах. Несмотря на то что навоз сравнительно быстро разлагается, он все же имеет возможность сохраняться в переувлажненных условиях (Долгих, Александровский, 2010; Bleicher, 2016), под скальными навесами (Delhon et al., 2008; Mlekuz, 2009; Бабенко и др., 2018) или в загонах для скота в аридных зонах (Shahack-Gross, 2011). При отсутствии консервирующих факторов навоз разлагается, а его остатки фиксируются стратиграфически или планиграфически в виде небольших прослоек, линз или пятен темно-коричневого цвета. Известны случаи обнаружения отдельных пометин или их фрагментов (см., например: Wilson, 1979; Akeret, Rentzel, 2001). Чаще всего сохранению навоза способствует огонь (Mlekuz, 2009; Spengler, 2019). В основном же археологи имеют дело с “сухим” культурным слоем, где остатки навоза попадают в состав мусора, рассеиваются в процессе каждодневной деятельности людей, не оставляя видимых следов (Miller, Smart, 1984; Сергеев, Лебедева, 2018).
Изучение древнего навоза позволяет решать ряд вопросов: реконструкция питания животных (см., например: Akeret, Jaсomet, 1997), локальной палеоэкологической обстановки (Carrion et al., 2000; Бабенко и др., 2018), особенностей систем скотоводства, земледелия и землепользования (см., например: Akeret, Rentzel, 2001, Bleicher, 2016), использования растительных ресурсов населением (Fuks, Dunseth, 2021), масштабов применения навоза в качестве топлива (Mlekuz, 2009), сезонности стоянок и заготовок кормов (Akeret, Jaсomet, 1997; Karg, 1998). Роль ботанического направления при изучении археологического навоза сложно переоценить (см., например: Wilson, 1979; Derreumaux, 2005).
В 2019 г. при работах на Киевском городище была зафиксирована органогенная прослойка, ставшая предметом настоящего исследования. Цель данной работы – комплексный анализ образцов навоза как источника информации по реконструкции системы кормления и содержания скота населением Киевского городища II–IV вв. н.э.
Киевское городище находится в Моздокском районе РСО-Алания у с. Киевское, на правом коренном берегу р. Терек. Площадь укрепленной части городища достигает 6.5 га. Оно выделяется в рельефе, представляя собой выступающий в северном направлении мыс треугольной формы, ограниченный по периметру крутыми склонами со следами эскарпирования (рис. 1). Визуально прослеживаются цитадель и две площадки, отделенные двумя рвами. Территория городища с юго-восточной стороны сильно разрушена карьерами по добыче глины и силосными ямами. На второй площадке городища устроена одна из таких ям в виде глубокой траншеи длиной 29 м. В 2019 г. экспедицией Института археологии РАН (рук. Д.С. Коробов) в северном борту траншеи проведена зачистка, в ходе которой зафиксированы восемь хозяйственных ям и участки культурного слоя (разрез 1) (рис. 2). Из ям и прослоек на разной глубине отобран материал для археоботанического анализа: 22 пробы для изучения макроостатков и 57 – для палинологического анализа.
Рис. 1.
Топографический план Киевского городища, 2019 г. Стрелкой обозначено месторасположение разреза 1. Условные обозначения: а – деревья; б – ямы; в – ЛЭП; г – ров; д – протейхизма; е – траншея; ж – канал; з – карьер; и – проселок; к – шоссе; л – шурф 1; м – поле; н – кустарник.
Fig. 1. Topographic plan of the Kievskoye fortified settlement in 2019. The arrow indicates the location of cross-section 1
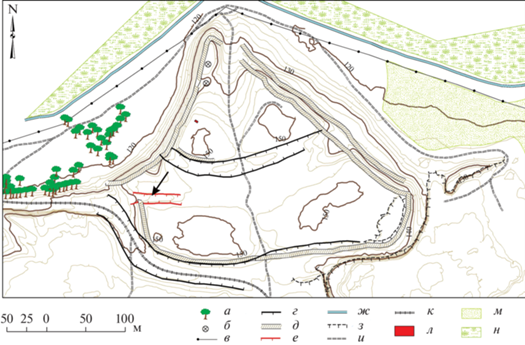
Рис. 2.
Ортофотография разреза 1, полученная методом фотограмметрии. Стрелкой обозначена прослойка навоза в культурном слое восточной части разреза 1.
Fig. 2. Orthophoto of cross-section 1 obtained by photogrammetry. The arrow indicates the dung streak in the cultural layer of the eastern part of cross-section 1

В данной работе рассматриваются результаты изучения образцов из темной серо-коричневой прослойки, расположенной в восточной части профиля разреза 1 на глубине 3.1 м от поверхности на отметках 27.0–29.2 м по базовой линии разреза (рис. 2). Судя по расположению в нижней части культурного слоя, данная прослойка была сформирована на раннем этапе существования городища.
Для изучения растительных макроостатков (зерна, семена, мякина и древесный уголь) из интересующей нас прослойки отобран один образец (№ ан. 3359), а для палинологического анализа – два.
Угли и другие макроостатки выделялись по единой методике. Объем пробы (№ ан. 3359) составил 5 л – половину стандартного объема (Лебедева, 2016), что связано с плотностью и небольшой мощностью (13 см) прослойки. В процессе взятия пробы остатки растений не отмечены, но сам характер отложений напоминал сгоревший навоз – имел слабозаметную тонкослоистую структуру, содержал золу – чем явно отличался от вмещающего слоя. После флотации образца на сите с ячейкой 0.5 мм осталось большое количество нераспавшихся фрагментов навоза, зерен, угля и фрагментов соломы общим объемом 200 мл.
Навозные отложения часто имеют высокую концентрацию растительных макроостатков (см., например: Akeret, Rentzel, 2001; Derreumaux, 2005), поэтому в археоботанике применяется методика учета макроостатков по субпробам (Steiner et al., 2017). В нашем случае были полностью учтены все остатки из фракций крупнее 2 мм, а из фракций 0.5–2 мм исследована только одна четвертая часть, после чего проводились пересчет по всем таксонам и суммирование с фракциями крупнее 2 мм.
Определение породного состава древесного угля выполнялось для фрагментов размером от 4 мм. Частицы углей раскалывались на три части в радиальной, тангенциальной и поперечной проекциях. В большинстве случаев определения осуществлялись до рода. Исключение составляют фрагменты, определенные до подсемейства яблоневых Maloidae и семейства лоховых Elaeagnaceae, представители которого – растения рода лох Elaeagnus и облепиха Hippophaë – не различимы по анатомическому строению древесины.
Палинологическим методом изучено два образца из верхней (№ 2) и нижней (№ 3) частей прослойки. Для сравнения также проанализированы образцы из выше- и нижележащих слоев (№ 1, 4). Пыльца и споры выделялись согласно стандартной сепарационной методике В.П. Гричука (Пыльцевой анализ, 1950. C. 32–35). При микроскопировании учитывались не только зерна пыльцы и споры, но и непыльцевые палиноморфы (микроугли и споры грибов). Для подсчета концентрации использовались таблетки со спорами Lycopodium clavatum (Stockmarr, 1973). При расчете процентной доли пыльцы за 100% принималась сумма пыльцевых зерен древесных и травянистых растений, а доля спор рассчитывалась от суммы подсчитанных пыльцы и спор. Диаграммы построены в программе Tilia 2.6.1 (Grimm, 2019).
Археоботанический анализ11. Изученный образец характеризуется высокой концентрацией макроостатков: в 5-литровом объеме содержалось 10 857 ед., что в несколько раз превышает среднюю концентрацию в слоях из разреза 1. Основу пробы составляют зерно, фрагменты соломы, мякины и семена сорных растений. Практически все компоненты происходят из навоза, что подтверждают оставшиеся включенными в его нерастворившиеся фрагменты зерновки и колосовые чешуи, а также загрязненная частицами органики поверхность большинства зерновок, веточек, специфически деформированные фрагменты мякины.
Бóльшую долю макроостатков составляют культурные растения (63%). На долю определимых зерновок культурных злаков приходится 31.2%, неопределимых фрагментов – 3.1%, мякина составляет 21.7%. Узлы соломы не учтены в общей статистике. Сохранность зерна однородная, может быть оценена в 3.5–4 балла по 5-балльной шкале. Зерновые находки представлены преимущественно просом обыкновенным Panicum miliaceum (78.5%), тогда как ячмень Hordeum vulgare составляет лишь пятую часть. Еще два вида культурных растений зафиксированы в незначительных количествах: пшеница однозернянка Triticum monococcum и лен Linum usitatissimum.
Зерновки проса представлены как выполненными, зрелыми экземплярами, так и щуплыми – невызревшими или недоразвитыми. Последние составляют 60.3% от общего количества проса. Изначально, вероятно, все они были неочищенными. Ячмень, как и просо, был скормлен и позже сгорел в неочищенном состоянии. Ячмень в пробе представлен исключительно пленчатой формой многорядного подвида Hordeum vulgare ssp. vulgare. Наряду с крупными, выполненными, встречены зерновки узких пропорций (рис. 3, 1). Некоторые из них настолько тонкие, что схожи с дикими представителями семейства мятликовые Poaceae. Возможно, как и в случае с просом, они происходят из невызревших колосьев.
Рис. 3.
Карбонизированные зерновки ячменя (1) и колоски однозернянки (2). Масштаб – 1 мм.
Fig. 3. Carbonized barley grains (1) and spikelets of einkorn (2). Scale – 1 mm

Сохранность мякины заметно хуже, чем зерна. Преобладают колосовые остатки ячменя (87.6%), а на однозернянку приходится 6.8%, еще часть их отнесена в общую категорию “пленчатые пшеницы” (3.9%) и к роду Triticum sp. (0.3%). Также обнаружено три фрагмента, видимо, от метелки проса с характерными утолщениями в местах разветвления. Ячмень представлен заметно деформированными остатками колосового стержня (отдельными целыми сегментами (685 ед.) и их многочисленными мелкими фрагментами (1141 ед.)), основаниями колоса (9 ед.) и цветочных чешуй (224 ед.). В колосовых остатках пленчатых пшениц (в основном однозернянки) встречаются колоски разной степени сохранности (42 ед.) (рис. 3, 2), вилочки (94 ед.), основания чешуй (93 ед.) и основания колоса (21 ед.).
Несмотря на то что при разборе пробы в лаборатории отмечено значительное присутствие соломы, по факту учтено всего 26 ее узлов, что не так много по сравнению с аналогичными контекстами (см. Derreumeaux, 2005).
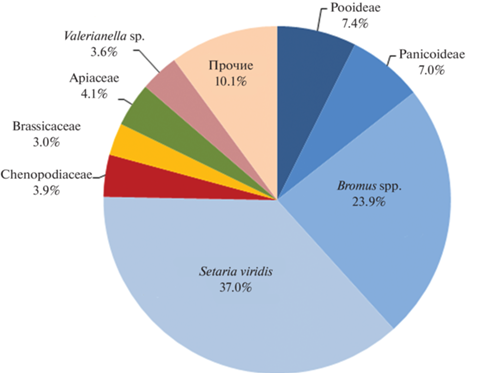
Доля сорных и дикорастущих видов составляет 34.5%; всего определено 42 таксона различного уровня. Среди них три четверти относятся к семейству мятликовые Poaceae (рис. 4), из которых самые массовые щетинник зеленый Setaria viridis (37%) и костер Bromus spp. (24.4%), в основном представленный видом B. arvensis. Щетинник – один из основных засорителей проса, а учитывая его количество и состояние, нет сомнений в том, что здесь он связан именно с урожаем данной культуры. Костер полевой в степной части выступает чаще в качестве рудерального сорняка (Никитин, 1983. С. 107). Остальные семейства совокупно занимают 25%. Преобладают сорные таксоны, а также растения широкого диапазона обитания.
Рис. 4.
Структура дикорастущих и сорных травянистых таксонов.
Fig. 4. Structure of wild and weedy herbaceous taxa
В категории “прочие” учтены макроостатки, которые не были достоверно причислены ни к одной из описанных выше категорий (2.5%). В основном это фрагменты, скорее всего остатки зерновок злаков и, вероятно, коробочек льна, также отмечено несколько фрагментов оболочек плодов, плодоножек и, вероятно, бутонов, не поддающихся таксономической идентификации.
Антракологический анализ. В изучаемом образце отобрано 140 фрагментов древесного угля, из которых удалось определить видовую принадлежность 101 фрагмента. Анализ углей показал присутствие не менее семи таксонов древесных пород, основные из которых дуб Quercus и лох/облепиха Elaeagnus/Hippophaë (рис. 5). Насчитываются сотни фрагментов древесных молодых побегов (менее 4 мм). Из них для таксономических определений оказались годными лишь 12: 1 неопределенный, а 11 принадлежат Elaeagnus/Hippophaë (рис. 6). На поперечных срезах этих побегов видны полностью сформировавшиеся годовые кольца и граница с камбием (рис. 6, 1), что свидетельствует об их срезке во второй половине года. Многие из этих фрагментов обмазаны навозом, что дает основания предполагать, что они также входили в состав корма.
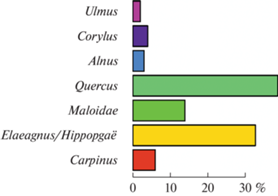
Рис. 6.
Поперечный (1) и радиальный срезы (2) молодого побега Elaeagnus/Hippophaë.
Fig. 6. Transverse (1) and radial sections (2) of a young shoot of Elaeagnus/Hippophaë

Спорово-пыльцевой анализ. Изученные образцы из прослойки (№ 2, 3) от выше- и нижележащих образцов (№ 1, 4) резко отличаются по составу как пыльцевых, так и непыльцевых палиноморф (рис. 7). В спектрах образцов № 2 и 3 выявлено лишь 10 таксонов, тогда как в выше- и нижележащих образцах определено в 2 раза больше. Образцы из прослойки характеризуются отсутствием, а № 1 и 4 – низким содержанием пыльцы древесных пород. Пыльца сосны Pinus, ольхи Alnus и лещины Corylus может переноситься ветром на большие расстояния, поэтому можно говорить лишь о произрастании ивы Salix в окрестностях городища. В образцах № 1 и 4 доминируют мятликовые Poaceae (49–60%). Пыльцевые спектры прослойки относятся к монодоминантным с преобладанием пыльцы астровых, среди которых доля трибы цикориевых Cichorioideae составляет 72–82%. Остальные таксоны (в том числе и пыльца культурных злаков Cerealia t.) представлены единично, за исключением мятликовых (рис. 7, А).
Рис. 7.
Диаграммы распределения пыльцевых (А) и непыльцевых (Б) палиноморф в культурном слое Киевского городища.
Fig. 7. Diagrams of the distribution of pollen (A) and non-pollen (Б) palynomorphs in the cultural layer of the Kievskoye fortified settlement
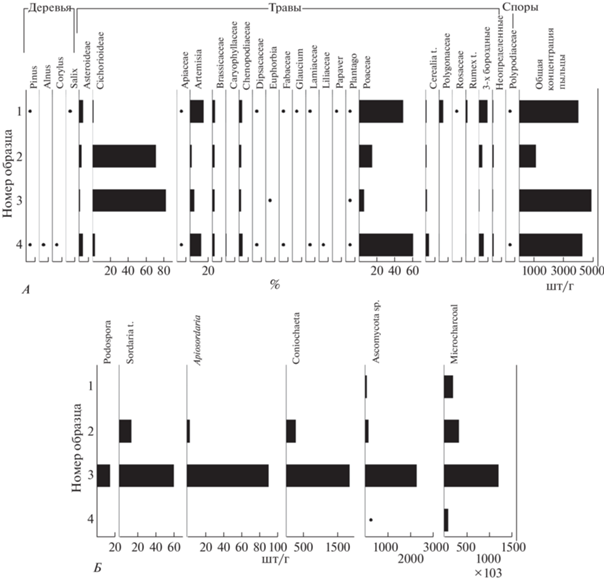
Образцы № 1 и 4 характеризуются низкой концентрацией аскоспор (67 и 8 шт/г соответственно), тогда как в № 3 концентрация аскоспор достигает 4314 шт/г, среди которых определены копрофильные споры Podospora, Sordaria t., Apiosordaria и Coniochaeta. Концентрация микроуглей в этом же образце достигает более миллиона в 1 г грунта. В образце № 2 концентрация аскоспор, как и микроуглей, значительно ниже (447 и 328 800 шт/г соответственно), но эти показатели существенно выше, чем в окружающих образцах (рис. 7, Б).
Итак, спорово-пыльцевые спектры, состав и концентрация макроостатков изучаемой прослойки указывают на их навозное происхождение. Большая доля пыльцы насекомоопыляемых растений и наличие слипшейся в комки пыльцы характерны для спектров экскрементов травоядных (Carrion, 2002; Бабенко и др., 2018; Florenzano, 2019). Кроме того, на навоз указывает и присутствие спор копрофильных грибов (Krug et al., 2004).
Видовая принадлежность навоза. Жвачные животные (коровы, козы, овцы) гораздо тщательнее пережевывают и переваривают корм, поэтому в их навозе почти не фиксируются целые семена и растительные фрагменты крупнее 2 мм, зерно же усваивается почти полностью (см., например: Wallace, Charles, 2013; Spengler, 2019). Навоз жвачных не только слабо насыщен семенами и мякиной, но и беден таксономически (см., например: Akeret, Jaсomet, 1997; Schepers, Van Haaster, 2015). Строение пищеварительной системы у свиней также имеет свои особенности (Аксенова, Осипов, 2013. С. 72–74), в связи с чем и конечные продукты тоже будут иметь иную структуру и состав, зерно они также усваивают почти полностью (80% – см. Черницкая [Электронный ресурс]).
Пищеварительная система лошадей принципиально отличается от жвачных (Аксенова, Осипов, 2013. С. 69–71), поэтому концентрация растительных остатков в конском навозе выше (Anderson, Ertug-Yaras, 1998), он более богат на идентифицируемый растительный материал (Wilson, 1979; Derreumaux, 2005). В результате экспериментального сожжения навоза разных видов животных нами получены образцы, при сравнении с которыми выявлено наибольшее сходство фрагментов навоза из изучаемой прослойки именно с конским22 (рис. 8).
Рис. 8.
Матрицы горелого современного (1) и древнего (2) конского навоза. Масштаб – 1 мм.
Fig. 8. Matrices of burnt modern (1) and ancient (2) horse dung . Scale – 1 mm

Состав навоза (макроостатки). Состав навоза определяется рядом факторов: доступностью тех или иных видов растений, выбором корма людьми или животными при выпасе, сезоном, особенностями пищеварения, а также внедрением материала, не связанного изначально с навозом. Также необходимо учитывать, что животными бывают случайно съедены древесина, уголь, кости, острые шипы, даже керамика и ядовитые части растений (Wilson, 1979; Akeret, Rentzel, 2001).
Важно отметить, что почти две трети образца представлено культурными злаками при значительном участии мякины в структуре образца (21.7%). Вероятнее всего, после обмолота просо и ячмень не были отвеяны и отсортированы и хранились в таком виде, т.е. либо какая-то часть, либо даже весь урожай с конкретного поля предназначался для фуража. Возможно и обратное – смесь от разных стадий очистки этой культуры (см. Сергеев, Лебедева, 2021). Второй вариант выглядит более вероятным, поскольку пыльца культурных злаков присутствует единично.
В случае с однозернянкой использовались именно отходы от обмолота этого вида пшеницы, о чем свидетельствуют почти целые колоски, основания колоса и малое количество зерновок. Высокая концентрация вилочек и оснований чешуй (в 7.5 раз больше, чем в среднем в слое) говорит о присутствии в корме и отходов от домашней очистки. Возможно, и отходы обмолота льна использовалась в фураже, что отмечено и на других памятниках (см. Kühn et al., 2013).
Солома злаков могла входить как в состав фуража, так и в состав подстилки в стойле. Несмотря на то что находки узлов соломы не указывают прямо на ее использование, в нашем случае мы вправе предполагать, что именно их невысокая концентрация вероятнее позволяет связать солому с кормом. Овсяная, просяная и ячменная солома – ценная добавка для кормления (см., например: Бобинский, 1836. С. 163; Калоев, 1993. С. 119; Милов, 1998. С. 227).
Как уже было сказано, луговые травянистые виды, которые служат маркерами использования сена или выпаса животных, представлены очень слабо. Бóльшая часть дикорастущих травянистых растений относится к сорнякам (Poaceae, Chenopodiaceae, Brassicaceae), поэтому некоторые из них привнесены в корм с зерновыми культурами, другие могли расти вблизи человеческого жилья, на пустырях, залежах и вдоль дорог, где были съедены животными. Здесь же могли обитать и растения более широкого экологического диапазона: Rosaceae, Fabaceae, Lamiaceae, Valerianella sp. (Косенко, 1970). Луг могут представлять Cynodon dactylon, Plantago sp., Prunella sp., Phleum sp. Их участие в спектре очень незначительно, однако первые три вида обычно невысокие растения, поэтому в составе сена им оказаться сложно. Семена Lappula sp. и Daucus carota покрыты шипиками и могли прицепиться к шерсти животных (см. Fuks, Dunseth, 2021), после чего попали в стойло, где животные подвергались регулярной чистке, оказались на полу и сгорели вместе с навозом.
Сложнее всего связать с исследованным комплексом семена белены Hyoscyamus niger. Алкалоиды всех частей этого растения токсичны для животных (Alizadeh et al., 2014). Семена несъедобных растений могут случайно быть проглочены животными, если они присутствуют в заготовляемых кормах (Stegelmeier, Panter, 2012), а также применяться в ветеринарии (Lempiainen, 1991).
Еще один важный компонент растительного корма – это молодые побеги древесных растений. Наибольшее значение в нашем случае имели лох/облепиха, а также представители подсемейства яблоневые, использовались, вероятно, и ветви дуба. Существует немало свидетельств (начиная с эпохи неолита) применения в холодный период года веточного корма самых разных древесных видов (см., например: Краснов, 1971. С. 124–126; Akeret, Jaсomet, 1997; Милов, 1998. С. 241; Kühn et al., 2013; Bleicher, 2016). Известно применение листового корма в случае бескормицы и на Северном Кавказе (Калоев, 1993. С. 119, 122).
Сезонность и система кормления. Наиболее вероятным сезоном формирования спорово-пыльцевых спектров рассматриваемой прослойки навоза является осень. В условиях равнинных жарких летних условий (Будун, 1994, С. 60), в которых расположено городище, основная часть растений к этому времени отцветает, что объясняет бедные спектры. Однако 50% видов семейства астровых, произрастающих в настоящее время в Моздокском районе, продолжают цвести до сентября или октября (Косенко, 1970. С. 394–453; Галушко, 1980. С. 164–271). Семена выявленных травянистых таксонов имеют растянутый период созревания – от июня до сентября, т.е. также могли присутствовать в осеннем навозе. Этому периоду противоречит только наличие валерианницы, чьи семена созревают уже в мае-июне (Косенко, 1970. С. 369, 370).
Все молодые побеги из навозного слоя принадлежат веткам возрастом 1 год или меньше. С уверенностью можно сказать, что они собраны во второй половине года, судя по тому, что в этих образцах есть выраженный переход от ранней к поздней древесине. Работы по изучению питания копытных показывают, что лошади хоть и менее охотно, но могут включать в свой рацион веточный корм в зимний период (Prieditis, 2002; Cromsigt et al., 2018). Таким образом, результаты исследования (незначительное присутствие семян пастбищных видов, обилие зерна и мякины, преобладание пыльцы астровых на фоне низкого таксономического разнообразия, присутствие веточного корма) указывают на то, что наиболее вероятный период формирования навозной прослойки – осенне-зимний.
Чаще всего фураж применяется по причине отсутствия доступного свежего травянистого корма (Anderson, Ertug-Yaras, 1998). Зерно же используют и для откорма на мясо или усиленного питания животных. В случае лошадей это может быть связано с тяжелой работой или подготовкой к усиленным нагрузкам (см., например: Шаманов, 1972; Калоев, 1993. С. 79). Значительное количество зерна, соломы и мякины в навозе может свидетельствовать и о том, что лошадей содержали обеспеченные хозяева (Калоев, 1993. С. 125).
Просо и ячмень – давно признанные первосортные кормовые злаки (Бобинский, 1836). На Кавказе кормление просом лошадей считается традицией адыгских народов, при этом оно воспринималось и как лекарственное средство (Калоев, 1993. С. 79). Конечно, материал из Киевского ни в коем случае нельзя расценивать как этнический маркер, а преобладание проса скорее связано со сходными агроэкологическими условиями предгорных равнин разных частей Северного Кавказа. Пленчатые пшеницы обычно не задаются лошадям и ослам или только в малых количествах, так как могут быть вредны для них (Anderson, Ertug-Yaras, 1998). Чистое зерно в больших количествах вредно для животных, поэтому для фуража зерно оставляют необработанным или вводят значительное количество мякины и соломы; этот прием известен коневодам издревле (см., например: Бобинский, 1836. С. 163).
Если культурные злаки представляли часть фуража, то источник дикорастущих видов – заготовленные корма (ветки и сено) или растения, поедаемые животными при выпасе, – трудно установить по археоботаническим данным. Однако преобладание среди семян рудеральных и универсальных дикорастущих растений может указывать на содержание животных на подножном корме (наряду с подкормкой фуражом). При этом поиск растений происходил в основном на антропогенно измененных участках. В случае поедания животными заготовленного сена следовало бы ожидать более богатый спектр луговых трав как в составе пыльцевых спектров, так и в представленности семян.
Заготовка луговых трав была ограничена, вероятно, и в связи с отсутствием кос. Несмотря на существующее представление, что косы в Восточной Европе распространяются со II–V вв. (Краснов, 1971. С. 124), на памятниках раннего этапа аланской культуры (II–IV вв.) находки кос пока неизвестны. Имеется единичный фрагмент железной косы на поселении VIII–IX вв. Козьи Скалы возле Пятигорска (Кузнецов, Рудницкий, 1998. С. 304. Рис. 11 , 10). Многочисленные фрагменты кос известны на памятниках салтово-маяцкой археологической культуры Среднего и Нижнего Дона (Михеев, 1985. С. 29–32; Колода, Горбаненко, 2010. С. 38), некоторые из которых оставлены представителями аланского этноса. Кроме того, существует устойчивое представление о том, что название косы у некоторых народов Северного Кавказа восходит к аланской терминологии и является, таким образом, одним из элементов аланского культурного наследия (Калоев, 1981. С. 222). Однако очевидно, что само появление этого инструмента относится уже к эпохе раннего средневековья, что фиксируется также у германских народов (см., например: Henning, 1985).
Относительно высокое разнообразие углей и небольшое количество фрагментов молодых древесных побегов могут также указывать скорее на самостоятельное поедание их животными, а не выступать свидетельством целенаправленной заготовки корма. На территории Европы содержание животных в зимний период в большей степени на подножном корму практиковалось, вероятно, долгое время – с неолита до железного века, а в некоторых регионах и до XVIII в. (Hejcman et al., 2014). На Кавказе еще в XIX в. часто практиковали свободный выпас животных в течение всего года (см. Калоев, 1993. С. 125).
Аммиан Марцеллин писал, что аланы много внимания уделяют заботе о лошадях (Алемань, 2003. С. 71). Рекомендации по поддержанию боевых/рабочих коней в хорошей форме, схожие с современными, можно найти у римских авторов (см., например: Wilson, 1979). Сопоставление их с пропорциями основных компонентов фуража из исследованного слоя показывает, что жители Киевского городища во II–IV вв. понимали, как правильно кормить лошадей.
Производство и распределение зерновых. Навозный слой из Киевского выводит нас и на ряд других тем. Во-первых, наличие значительного количества мякины всех трех видов злаков в корме говорит о том, что коневладельцы имели прямой доступ к этому ресурсу. Это дает возможность предполагать, что выращивание трех культур – ячменя, проса, пшеницы – осуществлялось в ближней округе городища, так как не очищенное от отходов обмолота и сорняков зерно нецелесообразно перевозить на большие расстояния. Во-вторых, наши исследования показали, что просом аланы “делились” со своими лошадьми, зерно пшеницы потребляли сами, а вот пищевой статус ячменя могут показать только дальнейшие исследования.
Тафономия и происхождение прослойки. Не вызывает сомнений факт единовременного сгорания обнаруженных остатков – слой довольно однородный, без разрывов. Период, в течение которого накапливался навоз, неизвестен, но есть основания предполагать, что сгорание произошло довольно быстро. Изучение лежалого навоза показывает, что в нем довольно быстро появляются насекомые, в первую очередь мухи. Оболочки их куколок сохраняются в образцах даже после сожжения. В навозе из Киевского городища таких остатков нет. Кроме того, при долгом экспонировании на открытом воздухе остатков навоза из-за постоянного “пыльцевого дождя” их спорово-пыльцевые спектры в большей степени отражали бы окружающую растительность (Ершова и др., 2017). В то же время эти экофакты могут быть связаны с зимним периодом, что не противоречит нашей основной аргументации. В позднеосенний-зимний период в воздухе практически нет пыльцы, так как нет цветущих растений, поэтому можно предположить, что навоз мог накапливаться в течение этого периода, и был сожжен и перекрыт культурным слоем до наступления весны, когда распускаются весеннецветущие растения. О коротком периоде между отложением и сгоранием говорит и отсутствие следов порчи зерен грызунами. Навоз не подвергался растаптыванию и перемешиванию, так как в слое сохранились хрупкие карбонизированные макроостатки, отсутствует керамика, а находки костей редки. О сохранении навоза in situ говорит и тот факт, что в верхней части изучаемой прослойки концентрация аскоспор, микроугля и пыльцы значительно меньше, чем в нижней. Это может быть связано с лучшим прогоранием в верхней части навозного слоя.
Таким образом, навоз не только быстро сгорел, но и был сразу перекрыт слоем грунта. Нельзя сказать наверняка, имеем ли мы дело с намеренным выжиганием при санитарной чистке стойла, или же с пожаром, уничтожившим стойло.
В заключение отметим следующее. Комплексное изучение навозной прослойки, сохранившейся в культурном слое Киевского городища, показало уникальность и высокий информационный потенциал такого рода материала. Хорошая сохранность хрупких растительных макроостатков и распределение концентрации пыльцевых и непыльцевых палиноморф свидетельствуют об отсутствии перемешивания слоя после сгорания. Схожесть навозной матрицы с экспериментальными образцами и высокая доля крупных растительных остатков указывают на принадлежность исследованного навоза лошади.
По преобладанию в пыльцевых спектрах астровых, большой доле зерновых и наличию молодых веток в составе корма удалось установить осенне-зимнее формирование прослойки и, следовательно, реконструировать систему кормления лошадей в зимний период на раннем этапе существования городища. Разнообразие древесных пород при их небольшом количестве и наличие семян рудеральных растений, таксономически бедные пыльцевые спектры скорее указывают на отсутствие заготовки сена и веточного корма для животных. Вероятнее всего животные содержались на подножном корме. В связи со скудностью доступного естественного корма зимой лошадей подкармливали зерном.
Таким образом, изучение конского навоза позволяет получить сведения о содержании животных, а также говорить и о производстве зерновых (проса, пшеницы и ячменя) в округе городища во II–IV вв. н.э.
Авторы выражают большую благодарность Е.Ю. Лебедевой за консультации на разных этапах настоящего исследования.
Статья подготовлена в рамках выполнения темы НИР ИА РАН “Междисциплинарный подход в изучении становления и развития древних и средневековых антропогенных экосистем” (№ НИОКТР 122011200264-9).
Список литературы
Аксенова В.М., Осипов А.П. Физиология системы пищеварения: учебное пособие. Пермь: Пермская гос. сельхоз. акад., 2013. 104 с.
Алемань А. Аланы в древних и средневековых письменных источниках. М.: Менеджер, 2003. 608 с.
Бабенко А.Н., Сергеев А.Ю., Коробов Д.С. Зоогенные (пометные) отложения как источник информации для комплексной реконструкции истории экосистем // Археология и естественные науки в изучении культурного слоя объектов археологического наследия: материалы междисциплин. науч. конф. / Ред. Д.С. Коробов и др. Пущино: КМК, 2018. С. 23–31.
Бобинский И. Краткая иппология и курс верховой езды. СПб.: Тип. Христиана Гинца, 1836. 219 с.
Будун А.С. Природа, природные ресурсы Северной Осетии и их охрана. Владикавказ: Рекламно-издат. агентство, 1994. 254 с.
Галушко А.И. Флора Северного Кавказа: определитель. Т. 3. Ростов-на-Дону: Изд-во Ростовского ун-та, 1980. 328 с.
Долгих А.В., Александровский А.Л. Почвы и культурный слой Великого Новгорода // Почвоведение. 2010. № 5. С. 515–526.
Ершова Е.Г., Бакуменко В.О., Воронцов Т.П., Гонча-ров М.М., Клименко М.С., Куликов Н., Ревокато-ва Д.П., Селезнева Е.М., Фариш Н.Р., Фетисова Е.С., Яковенко Е.П. Спорово-пыльцевые спектры современного и средневекового конского навоза // Материалы Всероссийской междисциплинарной научной конференции с международным участием “Палеопочвы, палеоэкология, палеоэкономика”. Пущино: КМК, 2017. С. 70–74.
Калоев Б.А. Земледелие народов Северного Кавказа. М.: Наука, 1981. 248 с.
Калоев Б.А. Скотоводство народов Северного Кавказа (с древнейших времен до начала XX века). М.: Наука, 1993. 231 с.
Колода В.В., Горбаненко С.А. Сельское хозяйство носителей салтовской культуры в лесостепной зоне. Киев: Ин-т археологии Нац. акад. наук Украины, 2010. 216 с.
Косенко И.С. Определитель высших растений Северо-Западного Кавказа и Предкавказья. М.: Колос, 1970. 614 с.
Краснов Ю.А. Раннее земледелие и животноводство в лесной полосе Восточной Европы. М.: Наука, 1971. 168 с.
Кузнецов В.А., Рудницкий Р.Р. Поселение “Козьи скалы” у горы Бештау // Материалы по изучению историко-культурного наследия Северного Кавказа. Вып. 1. Археология / Отв. ред. А.Б. Белинский. Ставрополь: Наследие, 1998. С. 297–332.
Лебедева Е.Ю. Археоботаника: методы исследований и интерпретация результатов // Междисциплинарная интеграция в археологии (по материалам лекций для аспирантов и молодых ученых). М.: ИА РАН, 2016. С. 118–146.
Милов Л.В. Великорусский пахарь и особенности российского исторического процесса. М.: РОССПЭН, 1998. 573 с.
Михеев М.К. Подонье в составе Хазарского каганата. Харьков: Вища школа, 1985. 148 с.
Никитин В.В. Сорные растения флоры СССР. Л.: Наука, 1983. 454 с.
Пыльцевой анализ. М.: Госгеолиздат, 1950. 540 с.
Сергеев А.Ю., Лебедева Е.Ю. Сырец как источник растительных макроостатков в культурном слое археологических памятников // Археология и естественные науки в изучении культурного слоя объектов археологического наследия: материалы междисциплин. науч. конф. / Ред. Д.С. Коробов и др. -Пущино: КМК, 2018. С. 148–154.
Сергеев А.Ю., Лебедева Е.Ю. Сорные растения из “фуражных” скоплений Болгарского городища // Аналитические исследования лаборатории естественнонаучных методов. Вып. 5. М.: ИА РАН, 2021. С. 285–307.
Черницкая М. Как и чем кормить свиней: основы кормления, рацион и методы откорма [Электронный ресурс] // Ferma.expert. URL: https://ferma.expert/jivotnie/svini/uxod/kormlenie-sviney/ (дата обращения: 26.11.2021).
Шаманов И.М. Скотоводство и хозяйственный быт карачаевцев в XIX – нач. XX в. // Кавказский этнографический сборник. Т. V. М.: Наука, 1972. С. 67–97.
Яворская Л.В. Предварительные результаты археозоологического исследования остеологической коллекции из шурфа № 1 поселения Киевское, исследованное в 2019 году // Архив Института археологии РАН. Р-1. Б/н. Л. 79–88. Коробов Д.С. Отчет об археологической разведке на городище Киевское в Моздокском районе РСО-Алания в 2019 г.
Akeret Ö., Jacomet S. Analysis of plant macrofossils in goat/sheep faeces from the Neolithic lake shore settlement of Horgen Scheller – an indication of prehistoric transhumance? // Vegetation History and Archaeobotany. 1997. V. 6. P. 235–239.
Akeret Ö., Rentzel P. Micromorphology and Plant Macrofossil Analysis of Cattle Dung from the Neolithic Lake Shore Settlement of Arbon Bleiche 3 // Geoarchaeology: An International Journal, 2001. V. 16. Iss. 6. P. 687–700.
Alizadeh A., Moshiri M., Alizadeh J., Balali-Mood M. Black henbane and its toxicity – a descriptive review // Avicenna Journal of Phytomedicine, 2014. V. 4. № 5. P. 297–311.
Anderson S., Ertug-Yaras F. Fuel Fodder and Faeces: An Ethnographic and Botanical Study of Dung Fuel Use in Central Anatolia // Environmental Archaeology. 1998. V. 1. Iss. 1. P. 99–109.
Bleicher N. Pastoralism, Landscape and Woodland Use in the Late Neolithic in Southern Germany // Agricultural and pastoral landscapes in pre-industrial society. Oxford: Oxbow Books, 2016 (Early agricultural remnants and technical heritage; vol. 3). P. 239–252.
Carrion J.S. A taphonomic study of modern pollen assemblages from dung and surface sediments in arid environments of Spain // Review of Palaeobotany and Palynology. 2002. V. 120. Iss. 3–4. P. 217–232.
Carrion J.S., Scott L., Huffman T., Dreyer C. Pollen analysis of Iron Age cow dung in southern Africa // Vegetation History and Archaeobotany. 2000. V. 9. № 4. P. 239–249.
Cromsigt J.P.G.M., Kemp Y.J.M., Rodriguez E., Kivit H. Rewilding Europe’s large grazer community: how functionally diverse are the diets of European bison, cattle, and horses? // Restoration Ecology. 2018. V. 26. Iss. 5. P. 891–899.
Delhon C., Martin L., Argant J., Thiébault S. Shepherds and plants in the Alps: multi-proxy archaeobotanical analysis of neolithic dung from “La Grande Rivoire” (Isère, France) // Journal of Archaeological Science. 2008. V. 35. Iss. 11. P. 2937–2952.
Derreumaux M. How to detect fodder and litter? A case study from the Roman site “Le Marais de Dourges”, France // Vegetation History and Archaeobotany. 2005. V. 14. № 4. P. 373–385.
Florenzano A. The History of Pastoral Activities in S. Italy Inferred from Palynology: A Long-Term Perspective to Support Biodiversity Awareness // Sustainability. 2019. V. 11. Iss. 2. P. 404–424.
Fuks D., Dunseth Z. Dung in the dumps: what we can learn from multi‑proxy studies of archaeological dung pellets // Vegetation History and Archaeobotany. 2021. V. 30. № 1. P. 137–153.
Grimm E.C. TILIA 2.6.1 version (computer software) [Электронный ресурс]. Illinois State Museum, 2019.
Hejcman M., Hejcmanová P., Stejskalová M., Pavlu V. Nutritive value of winter-collected annual twigs of main European woody species, mistletoe and ivy and its possible consequences for winter foddering of livestock in prehistory // Holocene. 2014. V. 24. Iss. 6. P. 659–667.
Henning J. Zur Datierung von Werkzeug- und Agrargeratfunden im germanischen Landnahmegebiet zwischen Rhein und oberer Donau. Der Hortfund von Osterburken // Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz. Bd. 32. Mainz, 1985. S. 570–594.
Karg S. Winter- and Spring-foddering of Sheep/Goat in the Bronze Age Site of Fiave- Carera, Northern Italy // Environmental Archaeology. 1998. V. 1. Iss. 1. P. 87–94.
Krug J.C., Benny G.L., Keller H.W. Coprophilous Fungi // Biodiversity of Fungi: Inventory and Monitoring Methods. Burlington, MA: Elsevier Academic Press, 2004. P. 467–500.
Kühn M., Maier U., Herbig C., Ismail-Meyer K., Le Bailly M., Wick L. Methods for the examination of cattle, sheep and goat dung in prehistoric wetland settlements with examples of the sites Alleshausen-Täschenwiesen and Alleshausen-Grundwiesen (around cal 2900 BC) at Lake Federsee, south-west Germany // Environmental Archaeology. 2013. V. 18. Iss. 1. P. 43–57.
Lempiainen T. Past occurrence of Hyoscyamus niger L. (Solanaceae) in Finland according to the macrofossil finds // Annales Botanici Fennici. 1991. V. 28. № 4. P. 261–272.
Miller N., Smart T.L. Intentional burning of dung as fuel: A mechanism for the incorporation of charred seeds into the archeological record // Journal of Ethnobiology. 1984. V. 4. № 1. P. 15–28.
Mlekuz D. The materiality of dung: the manipulation of dung in Neolithic Mediterranean caves // Documenta Praehistorica. 2009. V. 36. P. 219–225.
Prieditis A. Impact of Wild Horses Herd on Vegetation at Lake Pape, Latvia // Acta Zoologica Lituanica. 2002. V. 12. Iss. 4. P. 392–396.
Shahack-Gross R. Herbivorous livestock dung: formation, taphonomy, methods for identification, and archaeological significance // Journal of Archaeological Science. 2011. V. 38. Iss. 2. P. 205–218.
Shepers M., Van Haaster H. Dung matters: An experimental study into the effectiveness of using dung from hay-fed livestock to reconstruct local vegetation // Environmental Archaeology. 2015. V. 20. Iss. 1. P. 66–81.
Spengler III R.N. Dung burning in the archaeobotanical record of West Asia: where are we now? // Vegetation History and Archaeobotany. 2019. V. 28. № 3. P. 215–227.
Stegelmeier B.L., Panter K.E. Poisonous Plants and Plant Toxins That Are Likely to Contaminate Hay and Other Prepared Feeds in the Western United States // Rangelands. 2012. V. 34. № 2. P. 2–11.
teiner B.L., Antolín F., Vach W., Jacomet S. Subsampling of large-volume samples in waterlogged sediments. A time-saving strategy or a source of error? // Review of Palaeobotany and Palynology. 2017. V. 245. P. 10–27
Stockmarr J. Determination of spore concentration with an electronic particle counter // Danmarks Geologiske Undersøgelse Årbog. 1972. København, 1973. P. 87–89.
Wallace M., Charles M. What goes in does not always come out: The impact of the ruminant digestive system of sheep on plant material, and its importance for the interpretation of dung-derived archaeobotanical assemblages // Environmental Archaeology. 2013. V. 18. Iss. 1. P. 18–30.
Wilson D.G. Horse Dung from Roman Lancaster: A Botanical Report // Archaeo-Physika. 1979. Bd. 8. S. 331–350.
Дополнительные материалы отсутствуют.
Инструменты
Российская археология