

Палеонтологический журнал, 2022, № 5, стр. 29-36
Первая находка Bactritoidea (Cephalopoda) в ассельско-сакмарских отложениях рифа Шахтау (Башкортостан)
А. Ю. Щедухин *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: d_alsch2017@mail.ru
Поступила в редакцию 09.03.2022
После доработки 21.03.2022
Принята к публикации 25.03.2022
- EDN: QTWCVD
- DOI: 10.31857/S0031031X22050117
Аннотация
В статье кратко обсуждается эволюция взглядов на таксономическое положение бактритоидей внутри класса цефалопод. Рассмотрена история установления рода Ctenobactrites и входящих в него видов. Изученный бактрит отнесен к виду C. costatus Shimansky, 1951 и является первой находкой бактритоидей в асселько-сакмарском интервале карьера Шахтау. Кроме того, эта раковина превосходит по своим размерам всех известных представителей этого подкласса.
ВВЕДЕНИЕ
Бактритоидеи – подкласс головоногих моллюсков, существовавших c силура до позднего триаса (Mackenzie, 1966). Их характерными особенностями является небольшая прямая или согнутая раковина с узким краевым сифоном и обособленным протоконхом (Шиманский, 1962). Согласно современным представлениям, именно от бактритоидей в раннем девоне произошли Ammonoidea, а в раннем карбоне – Coleoidea (Kröger et al., 2011). Расцвет этой группы, по-видимому, пришелся на каменноугольный период. Находки бактритов известны на разных континентах – в Европе (Kröger et al., 2005), Азии (Шиманский, 1954; Niko et al., 1997), Африке (Pohle, Klug, 2018), Северной и Южной Америке (Mackenzie, 1966; Mapes, 1979; Cichowolski, Rustan, 2017), Австралии (Becker, 2000). Их положение в системе цефалопод до сих пор обсуждается. В разное время бактриты рассматривались в рангах отряда, надотряда и подкласса (Шиманский, 1951; Erben, 1964; Шевырев, 2005). Принадлежность этой группы к одному из высших таксонов головоногих моллюсков также вызывает дискуссии. Бактритов включали в состав наутилоидей, аммоноидей и колеоидей в ранге отряда, а иногда и семейства (Друщиц, Шиманский, 1982). Мы поддерживаем точку зрения отечественных исследователей (Barskov, Shimansky, 1994; Шевырев, 2005), которыми было обосновано выделение бактритоидей в ранге подкласса, равноценного аммоноидеям и колеоидеям.
Мировая история изучения пермских бактритов довольно скудна на события. Первые находки представителей этой группы в Европе, по-видимому, принадлежат Г. Джеммелляро (Gemmellaro, 1889). Он описал из пермских отложений Сицилии два экземпляра с прямой раковиной и поперечно-ребристой скульптурой под названием Orthoceras oblique-sulcatum. Другие три экземпляра с узким краевым сифоном и скульптурой из продольных ребер были описаны как другой новый вид – O. paternoi. В монографии (Gemmellaro, 1889) приводятся данные измерений длины сохранившейся части раковин, их поперечного сечения и апикального угла. Рисунки к работе выполнены достаточно хорошо, но, поскольку не дано соотношений основных параметров, только по изображениям трудно судить о пропорциях раковин.
В Северной Америке первый достоверный представитель бактритоидей был описан А. Миллером и У. Янгквистом (Miller, Youngquist, 1949) из средней и верхней перми Мексики. В конце 70-х гг. XX в. была опубликована большая специальная работа по каменноугольным и пермским бактритам Северной Америки (Mapes, 1979). Эта монография значительно расширила наши знания о позднепалеозойских бактритах. В ней было описано множество как известных ранее, так и новых видов, но только два из них (Bactrites elcapitanensis Mapes, 1979 и B. mexicanus Miller et Youngquist, 1949) – из нижнепермских отложений (США, Мексика). Р. Мэйпс подробно рассмотрел ранние стадии онтогенеза и предложил выделить новый отряд Annulobactritida в составе подкласса бактритоидей. При установлении этого отряда Мэйпс основывался на реконструкциях поврежденных апикальных частей раковин некоторых бактритов. Основным отличием аннулобактритид от бактритид он считал форму протоконха: у первых он конический, обособленный небольшим пережимом, а у вторых – сферический (Mapes, 1979).
Самый значительный вклад в исследования пермских бактритоидей с территории СССР внес В.Н. Шиманский. В нескольких работах (Шиманский, 1948, 1951, 1954, 1990, 1993) им было описано не менее 17 новых видов. На основании изучения сборов разных лет Шиманский (1951) первым предложил обособить бактритов от других крупных групп цефалопод и возвести их в ранг отряда. Позднее он изучил и описал строение начальных частей раковин различных палеозойских бактритов, выделив несколько типов протоконхов: сферический, яйцевидный и колпачковидный (Шиманский, 1958). Им было отмечено большое значение этого признака для построения системы бактритоидей, но не было выделено новых групп на данном основании. Новый взгляд на классификацию неаммоноидных цефалопод в целом и бактритов в частности был предложен в “Основах палеонтологии” (Шиманский, 1962). В этой сводке бактриты рассматривались в качестве надотряда с единственным отрядом Bactritida. Из пермских отложений было указано девять родов, большинство из которых были описаны с Южного Урала (Шиманский, 1954). Более поздние исследования пермских бактритов были посвящены описанию новых видов, но существенно подход к их изучению не изменился (Шиманский, 1990). Только в XXI в. было предпринято изучение микроструктуры раковин бактритоидей (Догужаева и др., 2016). Основой для этих исследований послужили коллекции Шиманского из нижней перми Южного Урала. В этой работе авторами на основе различий в строении протоконха подтверждается необходимость обособления бактритов от других крупных групп головоногих моллюсков. Больше специальных исследований пермских бактритоидей до настоящего момента не проводилось. Это может объясняться их редкой встречаемостью, а также малым числом исследователей, занимающихся данной тематикой. Поэтому любая новая информация по этой группе является очень полезной для понимания особенностей эволюции бактритоидей, индивидуального развития и уточнения связей с другими таксонами головоногих моллюсков.
Работа поддержана грантом РНФ № 22-24-00099 “Эволюция сообществ моллюсков раннепермского рифа Шахтау”.
МАТЕРИАЛ И МЕТОДИКА
Изученный в настоящей статье экземпляр происходит из пограничных ассельско-сакмарских отложений раннепермского рифа Шахтау. Шахтау ранее представлял собой одну из четырех отдельно стоящих гор, шиханов, расположенных в Стерлитамакском р-не республики Башкортостан, в 5 км на восток от г. Стерлитамак (рис. 1). В настоящее время это местонахождение превращено в карьер по добыче известняка для нужд содовой промышленности. В раннепермское время цепочка рифов, включающая в себя Шахтау, маркировала границу эпиконтинентального моря и более глубоководной части Уральского пролива.
Рис. 1.
Схема расположения местонахождения (отмечено пятиугольником): а – Уфа и Стерлитамак на карте европейской части России; б – положение карьера Шахтау рядом с г. Стерлитамак; в – схема карьера Шахтау по состоянию на 2021 г.
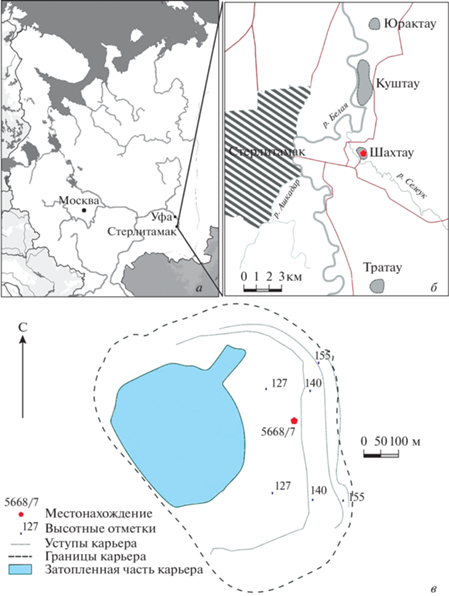
В 2021 г. экспедицией Палеонтологического ин-та им. А.А. Борисяка (ПИН) РАН под руководством А.В. Мазаева проводились полевые работы в карьере Шахтау. Уникальный образец был найден участником экспедиции А.Г. Боевым (Ин-т океанологии им. П.П. Ширшова РАН) и передан нам для изучения. Место находки расположено в восточной части карьера, в основании первого уступа, с абсолютными отметками 127–140 м (рис. 1). Согласно геологическим профилям (Mazaev, 2019), это место соответствует пограничным ассельско-сакмарским отложениям.
Измерения раковины произведены по общепринятым стандартам, применявшимся отечественными исследователями (Шиманский, 1954, 1968). Реконструкция длины недостающей части раковины выполнена по методике, описанной для ордовикских прямораковинных эндоцератоидей (Балашов, 1968). В статье при описании раковины приняты следующие сокращения:
Дс – срединный (сифонный) диаметр раковины; расстояние между дорсальной и вентральной сторонами. Дс – большой срединный диаметр, измеряется ближе к устью; дс – малый срединный диаметр, измеряется ближе к апикальному концу.
Дб – боковой диаметр. Измеряется между латеральными сторонами. Измерения Дб (большой) и дб (малый).
Дл – длина раковины между точками измерений диаметров.
К – число камер, приходящихся на диаметр (срединный диаметр).
Угол А – апикальный угол раковины. Срединный апикальный угол (с) измеряется между дорсальной и вентральной сторонами, боковой (б) – между латеральными сторонами.
Материал хранится в лаб. моллюсков ПИН РАН; экз. ПИН, №№ 5668/7-1, 5668/7-2 (отпечаток).
РЕЗУЛЬТАТЫ ИЗУЧЕНИЯ
Изучение строения раковины позволило установить ее принадлежность к виду Ctenobactrites costatus Shimansky, описанному из верхнесакмарских и артинских отложений Южного Урала (Шиманский, 1951). Раковина C. costatus прямая, очень крупная, умеренно расширяющаяся в латеральном и немного сильнее в дорсо-вентральном направлении (рис. 2, а–г). В поперечном сечении раковина овальная, сжата латерально (Дб/Дс = = 0.9). Боковой апикальный угол составляет 10°, срединный – 8°. Перегородки вогнуты почти на половину высоты камеры. Камеры высокие. На величину, равную дорсо-вентральному диаметру, приходятся 2.5–3 камеры. Поверхность раковины покрыта очень тонкими изогнутыми ребрышками (ширина около 0.1 мм), которые разделены широкими промежутками (до 2 мм). На вентральной стороне ребрышки образуют слабый выступ в сторону устья, переходящий в широкий синус на латеральных сторонах. Ребрышки на дорсальной стороне направлены вперед, в вершине образуют узкий, но неглубокий синус, осложненный отчетливо выраженным килем (рис. 2, д). Ребрышки сходятся на киле под тупым углом. На одну камеру приходятся 11 ребер. Перегородочная линия прямая, перпендикулярная к продольной оси раковины. Сифон краевой, узкий, плотно прилегает к вентральной стенке раковины. На вентральной стороне в месте контакта сифона со стенкой раковины расположена узкая, практически прямоугольная лопасть.
Рис. 2.
Раковина Ctenobactrites costatus Shimansky (×1): a–г – экз. ПИН, № 5668/7-1: a – вид со стороны перегородки, вентральная сторона сверху; б – дорсальная сторона; в – вентральная сторона; г – латеральная сторона, вентральная сторона справа; д – экз. ПИН, № 5668/7-2, отпечаток скульптуры, устье расположено с левой стороны, стрелкой отмечен дорсальный гребень; Башкортостан, Стерлитамакский р-н, карьер Шахтау; верхнеассельские-нижнесакмарские известняки.

В настоящее время известно 10 видов, относимых разными авторами к роду Ctenobactrites. От большинства из них C. costatus отличается тонкими изогнутыми ребрышками на поверхности раковины, которые значительно уже разделяющих их промежутков. От C. shindensis с Памира отличается узкими ребрышками и наличием киля на дорсальной стороне. Провести сравнение между нашим экземпляром и сицилийским C. obliquesulcatus не представляется возможным из-за отсутствия необходимых измерений у последнего. Можно только отметить, что у сицилийского вида, судя по изображению и описанию, отсутствует киль на дорсальной стенке раковины. От голотипа C. obliquesulcatus наш экземпляр отличается бóльшим углом расширения и немного бóльшей высотой камер. Эти отличия мы связываем с различными стадиями онтогенеза, поскольку голотип в поперечнике имеет 26.3 мм, в то время как наш экземпляр достигает 51.8 мм.
Из четырех сохранившихся камер предпоследняя ниже предыдущей, а последняя, более высокая, по-видимому, являлась жилой камерой. Уменьшение размеров камер фрагмокона непосредственно перед жилой камерой является признаком завершения роста моллюска. На этом основании можно сделать предположение, что раковина принадлежала взрослой особи.
Примерная длина недостающей части раковины была восстановлена по методике З.Г. Балашова (1968). Согласно расчетам, для дорсо-вентральной проекции примерная длина раковины могла составлять 322 мм, а для латеральной – 367 мм. Такая длина раковины была характерна для девонских–каменноугольных ортоцератоидей или некоторых ордовикских эндоцератоидей (Балашов, 1968). Следует отметить, что эта методика позволяет установить только длину утраченной апикальной части раковины. Длину недостающей части жилой камеры без каких-либо отпечатков или ее фрагментов установить невозможно. Предположительно, целиком раковина могла достигать около 400 мм в длину. Если это действительно так, то C. costatus принадлежал к наиболее крупным из известных прямораковинных цефалопод пермского периода (см. табл. 1, 2). В настоящее время вид C. costatus известен из ранней перми – ассельских, сакмарских и нижнеартинских отложений Южного Урала (Шиманский, 1954).
ОБСУЖДЕНИЕ
Род Ctenobactrites впервые был установлен Шиманским (1951). В этой работе был кратко охарактеризован вид C. costatus из стерлитамакского горизонта сакмарского яруса Казахстана и помещен в новое семейство Ctenobactritidae, с указанием скульптуры из поперечных ребер как основной критерий для его выделения. Через несколько лет, в монографии по прямораковинным цефалоподам Южного Урала, Шиманский (1954) более подробно описал C. costatus и установил новый вид C. mirus. Отдельно были упомянуты находки бактритов из пермских отложений Сицилии. Orthoceras oblique-sulcatum Gemmellaro был перенесен в род Ctenobactrites под названием C. obliquesulcatus. Другой сицилийский вид, Orthoceras paternoi, Шиманский (1954) предложил отнести к новому роду Sicilioceras, который он также включил в семейство Ctenobactritidae. Однако в более поздних работах Шиманского (1962, 1968) в составе семейства этот род не упоминается. В конце 50-х гг. прошлого века зарубежными исследователями была признана самостоятельность рода Ctenobactrites, но не семейства Ctenobactritidae. Свою точку зрения по этому вопросу высказывал Х. Эрбен (Erben, 1960). По его мнению, выделение этого семейства на основании одной лишь скульптуры недостаточно обосновано. В качестве примеров он привел роды, входящие в состав Bactritidae и обладающие сходной продольной или поперечной скульптурой. Таким образом, два рода, входившие в это семейство, были перенесены Эрбеном в семейство Bactritidae. Эту точку зрения приняли и последующие авторы. С этим мнением согласился и Шиманский (1993). В 1958 г. он описал несколько протоконхов бактритоидей. Среди них был описан фрагмент раковины из каменноугольных (башкирских–серпуховских) отложений Южного Урала, который автор отнес к Ctenobactrites sp. Эта раковина характеризовалась отсутствием сферической первой камеры, была достаточно крупной и имела коническую форму нескольких первых камер (Шиманский, 1958). В сводке “Treatise …” по неаммоноидным головоногим моллюскам Эрбен (Erben, 1964) предположил, что если найденная Шиманским эмбриональная камера действительно принадлежит роду Ctenobactrites, то она нарушает диагноз всей группы, и ктенобактриты должны быть исключены из бактритоидей. По мнению Мэйпса (Mapes, 1979), эта эмбриональная часть раковины Ctenobactrites sp., наряду с другими описанными им фрагментами раковин, относится к новому отряду Annulobactritida. Позднее Шиманский (1990) высказывал сомнения о принадлежности этой раковины к роду Ctenobactrites, полагая, что дальнейшие находки помогут разобраться в этом вопросе. Еще один вид Ctenobactrites, C. inhonorus Shimansky, был описан из Домбарских холмов (визе) Казахстана (Шиманский, 1968). Мэйпс (Mapes, 1979) описал три новых вида, принадлежащих Ctenobactrites. Два из них – C. lesliensis и C. collinsi. Третий вид описан в открытой номенклатуре из-за плохой его сохранности. C. collinsi был описан Мэйпсом по материалам А. Миллера и А. Анклсбея (Miller, Unklesbay, 1947), в их работе он первоначально был отнесен к роду Bactrites. В 1982 г. группой исследователей (Sturgeon et al., 1982) была произведена ревизия некоторых таксонов цефалопод из Северной Америки. Сравнение C. collinsi (Miller et Unklesbay) с материалом Ф. Мика (Meek, 1871; цит. по: Mapes, 1979) позволило свести этот вид в синонимию вида C. isogramma (Meek), ранее относимого к роду Orthoceras. Последние работы по роду Ctenobactrites вышли в 90-е годы XX в. Из артинских отложений Таджикистана был описан новый вид C. shindensis (Шиманский, 1993). Кроме этого, находки из каменноугольных отложений, не определенные до вида, но относимые к ктенобактритидам (Ctenobactrites sp.), были описаны японскими исследователями (Niko et al., 1991, 1997). Более поздних работ, в которых фигурировал бы род Ctenobactrites, обнаружить не удалось.
Полевые работы в Шахтау проводятся с 2015 г. Изучение коллекций из этого карьера позволило установить в ассельско–сакмарском интервале лишь несколько видов прямораковинных Pseudorthoceratida (Leonova, Shchedukhin, 2020), но бактриты в этих сборах отсутствовали. Мы связывали это с их небольшими размерами – известные виды из других шиханов в поперечнике не превышали 7 мм. Для большинства пермских южноуральских бактритов поперечное сечение в среднем составляет примерно 17 мм (Шиманский, 1954). Поэтому недавнее обнаружение раковины со срединным диаметром в 51.8 мм стало полной неожиданностью. В табл. 2 приводится сравнение размеров самых крупных представителей родов Ctenobactrites и Bactrites из позднего палеозоя. Как можно видеть, большие размеры (более 30 мм в диаметре) были известны преимущественно для каменноугольных бактритид. До настоящего момента самым крупным бактритом являлся Bactrites peytonensis Mapes, 1979 из миссисипия США.
Таблица 2.
Таблица измерений диаметров наиболее крупных позднепалеозойских бактритов
| Роды | Виды | Распространение | Диаметры в мм |
|---|---|---|---|
| Ctenobactrites | C. costatus Shimansky, 1951 | Нижняя пермь. Россия, Башкортостан | 51.8 |
| C. isogramma (Meek, 1871) | Пенсильваний. Северная Америка | 43 | |
| C. sp. (Niko et al., 1997) | Нижний карбон. Китай | ~43 | |
| C. sp. (Mapes, 1979) | Пенсильваний. США, Техас | 40 | |
| C. inhonorus Shimansky, 1968 | Нижний карбон. Казахстан | 40 | |
| C. shindensis Shimansky, 1993 | Нижняя пермь. Таджикистан, Памир | ~40 | |
| C. sp. (Niko et al., 1991) | Средний карбон. Япония, о. Хонсю | 34 | |
| C. obliquesulcatus (Gemmellaro, 1889) | Нижняя (?) пермь. Сицилия | 34 | |
| C. lesliensis Mapes, 1979 | Миссисипий. Северная Америка | 28 | |
| C. mirus Shimansky, 1954 | Нижняя пермь. Казахстан | 18.5 | |
| Bactrites | B. peytonensis Mapes, 1979 | Миссисипий. США, Арканзас | 50 |
| B. cherokeensis Miller et Owen, 1934 | Пенсильваний. США, Миссури | 42 | |
| B. gaitherensis Gordon, 1965 | Пенсильваний. США, Оклахома | 40 | |
| B. elcapitanensis Mapes, 1979 | Нижняя пермь. США, Техас | 34 | |
| B. finisensis Mapes, 1979 | Пенсильваний. США, Техас | 33 |
Изучение бактритоидей следует продолжать. Уже сейчас становится ясным, что наши знания о бактритоидеях верхнего палеозоя едва ли можно считать полными. Представители рода Ctenobactrites были широко распространены как в каменноугольное, так и раннепермское время. Новая находка в Шахтау позволила уточнить диагноз вида, изучить более поздние стадии онтогенеза C. costatus и расширить интервал его существования до позднеассельского–раннесакмарского времени.
Список литературы
Балашов З.Г. Эндоцератоидеи ордовика СССР. Л.: Изд-во ЛГУ, 1968. 278 с.
Догужаева Л.А., Богословская М.Ф., Журавлева Ф.А. Взгляды В.Н. Шиманского на филогенетическое значение бактритов и их дальнейшее развитие на материалах верхнего карбона и нижней перми Южного Урала в работах учеников и соратников // Золотой век российской малакологии. Сб. трудов Всеросс. научной конф., посвященной 100-летию со дня рождения проф. Виктора Николаевича Шиманского / Ред. Барсков И.С. и др. Москва–Саратов: ПИН РАН – СГТУ, 2016. С. 127–133.
Друщиц В.В., Шиманский В.Н. Основные вопросы систематики наружнораковинных головоногих // Бюлл. МОИП. Отд. геол. 1982. Т. 57. № 6. С. 94–107.
Шевырев А.А. Макросистема цефалопод: Исторический обзор, современное состояние и нерешенные проблемы. 1. Основные особенности и общая классификация головоногих моллюсков // Палеонтол. журн. 2005. № 6. С. 33–42.
Шиманский В.Н. Некоторые новые ортоцераконы из артинских отложений Южного Урала // Докл. АН СССР. 1948. Т. 60. № 1. С. 153–163.
Шиманский В.Н. К вопросу об эволюции верхнепалеозойских прямых головоногих // Докл. АН СССР. 1951. Т. 79. № 5. С. 867–870.
Шиманский В.Н. Прямые наутилоидеи и бактритоидеи сакмарского и артинского ярусов Южного Урала. М.: Изд-во АН СССР, 1954. 156 с. (Тр. Палеонтол. ин-та АН СССР. Т. 44).
Шиманский В.Н. О протоконхе бактритоидей // Докл. АН СССР. 1958. Т. 122. № 4. С. 702–705.
Шиманский В.Н. Надотряд Bactritoidea // Основы палеонтологии. Моллюски. Головоногие I / Ред. Руженцев В.Е. М.: Изд-во АН СССР, 1962. С. 229–238.
Шиманский В.Н. Каменноугольные Orthoceratida, Oncoceratida, Actinoceratida и Bactritida. М.: Наука, 1968. 151 с. (Тр. Палеонтол. ин-та АН СССР. Т. 117).
Шиманский В.Н. Новые находки ископаемых раковин бактритоидей в пермских отложениях Южного Урала // Ископаемые цефалоподы. М.: Наука, 1990. С. 66–70 (Тр. Палеонтол. ин-та АН СССР. Т. 243).
Шиманский В.Н. Пермские бактритоидеи Памира // Палеонтол. журн. 1993. № 3. С. 120–124.
Barskov I.S., Shimansky V.N. The morphological diversity and systematics of the Bactritoids (Cephalopoda) // Paleontol. J. 1994. V. 28. № 1A. P. 134–142.
Becker R.T. Paleobiogeographic relationships and diversity of Upper Devonian ammonoids from Western Australia // Rec. W. Australian Mus. 2000. Suppl. № 58. P. 358–401.
Cichowolski M., Rustán J.J. First report of Devonian bactritids (Cephalopoda) from South America. Paleobiogeographic and biostratigraphic implications // J. Paleontol. 2017. V. 91. № 3. P. 417–433. https://doi.org/10.1017/jpa.2017.17
Erben H.K. Primitive Ammonoidea aus dem Unterdevon Frankreichs und Deutschlands // N. Jb. Geol. Paläontol. Abh. 1960. Bd 110. № 1. S. 1–128.
Erben H.K. Bactritoidea // Treatise on Invertebrate Paleontology. Pt K. Mollusca 3. Lawrence: Univ. Kansas Press, 1964. P. 491–505.
Gemmellaro G.G. La fauna dei calcari con Fusulina della valle del fiume Sosio nella province di Palermo // Giorn. Sci. Natur. Econ. Palermo. 1889. № 19. P. 97–182.
Kröger B., Jakob V., Fuchs D. Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules // BioEssays. 2011. V. 33. № 8. P. 602–613. https://doi.org/10.1002/bies.201100001
Kröger B., Klug C., Mapes R.H. Soft-tissue attachments in orthocerid and bactritid cephalopods from the Early and Middle Devonian of Germany and Morocco // Acta Palaeontol. Polon. 2005. V. 50. № 2. P. 329–342.
Leonova T.B., Shchedukhin A.Yu. Asselian-Sakmarian nautiloids of the Shakh-Tau Reef (Bashkortostan) // Paleontol. J. 2020. V. 54. № 10. P. 1113–1134.
Mackenzie G. An Upper Triassic bactritoid cephalopod from California // J. Paleontol. 1966. V. 40. № 5. P. 1220–1222.
Mapes R. Carboniferous and Permian Bactritoidea (Cephalopoda) in North America // Univ. Kansas Paleontol. Contrib. 1979. Art. 64. 75 p.
Mazaev A.V. Lower Permian gastropods of Shakhtau (Asselian-Sakmarian boundary beds, Southern Cisuralia) // Paleontol. J. 2019. V. 53. № 12. P. 1237–1345. https://doi.org/10.1134/S0031030119120025
Miller A.K., Unklesbay A.G. The cephalopod fauna of the Conemaugh series in western Pennsylvania // Ann. Carnegie Mus. 1947. V. 30. P. 319–330.
Miller A.K., Youngquist W. American Permian nautiloids // Mem. Geol. Soc. Amer. 1949. № 41. P. 33–34.
Niko S., Nishida T., Kyuma Y. Middle Carboniferous Bactritoidea (Mollusca, Cephalopoda) from the Akiyoshi limestone group, Yamaguchi prefecture (Molluscan paleontology of the Akiyoshi limestone group-x) // Trans. Proc. Soc. Japan, N.S. 1991. № 161. P. 714–719.
Niko S., Nishida T., Kyuma Y. Moscovian (Carboniferous) orthoconic cephalopods from Guizhou and Guangxi, South China // Paleontol. Res. 1997. V. 1. № 2. P. 100–109.
Pohle A., Klug C. Early and middle Devonian cephalopods from Hamar Laghdad (Tafilalt, Morocco) and remarks on epicoles and cameral deposits // N. Jb. Geol. Paläontol. Abh. 2018. V. 290. № 1–3. P. 203–240. https://doi.org/10.1127/njgpa/2018/0776
Sturgeon M.T., Windle D.L., Mapes R.H., Hoare R.D. New and revised taxa of Pennsylvanian cephalopods in Ohio and West Virginia // J. Paleontol. 1982. V. 56. № 6. P. 1453–1479.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал