

Палеонтологический журнал, 2022, № 5, стр. 103-110
Новый вид скумбрий (Scomber, Scombroidei) из тархана (низы среднего миоцена) Северо-Западного Кавказа
А. Ф. Банников a, *, И. Г. Еребакан a, b, **
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
b Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: aban@paleo.ru
** E-mail: erebakan.ivan@mail.ru
Поступила в редакцию 06.04.2022
После доработки 11.04.2022
Принята к публикации 12.04.2022
- EDN: HYEVSO
- DOI: 10.31857/S0031031X22050051
Аннотация
Из тархана (низы среднего миоцена) Северо-Западного Кавказа (Краснодарский край, р. Пшеха) по нескольким скелетам описана скумбрия Scomber collettei sp. nov. (Percomorpha, Scombroidei, Scombridae). Это первая достоверная находка ископаемого представителя рода Scomber в тарханском региоярусе. Новый вид отличается от более древних позднеолигоценовых–раннемиоценовых скумбрий Восточного Паратетиса – S. cubanicus и S. gnarus – иным соотношением туловищных и хвостовых позвонков (14 + 17 вместо 15 + 16) и более разделенными спинными плавниками. S. collettei sp. nov. также характеризуется относительно крупными зубами и lacrimale.
ВВЕДЕНИЕ
Семейство Scombridae (скумбриевые) представлено эпипелагическими морскими преимущественно хищными рыбами с веретеновидным либо немного сжатым с боков телом и тонким хвостовым стеблем. Семейство Scombridae традиционно помещают в подотряд Scombroidei отряда окунеобразных (Perciformes) (Берг, 1940; Collette et al., 1984; Банников, 1985, 2010; Nelson, 2006 и др.). В последнее время, однако, на смену традиционной классификации организмов зачастую приходит формальная кладистическая систематика, построенная на компьютерной обработке молекулярных данных и часто противоречащая морфологическим свидетельствам. В кладистической систематике скумбриевидным придается ранг отряда (Wiley, Johnson, 2010; Near et al., 2012, 2013; Betancur-R. et al., 2013, 2017; Nelson et al., 2016 и др.) в “отделе (division) Percomorphacea” (Wiley, Johnson, 2010), “кладе Pelagia” (Miya et al., 2013) или “серии Pelagiaria” (Betancur-R. et al., 2017). По данным молекулярной систематики (Betancur-R. et al., 2017), отряд Scombriformes включает в себя 17 семейств, в т.ч. и семейство Scombridae. К семейству скумбриевых относят около 34 родов, включая вымершие (Банников, 2010, 2020б). Первые представители скумбриевых встречаются уже в позднем палеоцене, но их основное время диверсификации приходится на олигоцен и миоцен (Банников, 2010).
Исходя из морфологического сходства, принято делить семейство Scombridae на несколько подсемейств (Starks, 1910; Банников, 1985, 2010), либо триб Scombrini (настоящие скумбрии), Scomberomorini (королевские макрели), Sardini (пеламиды), Thunnini (тунцы) (Collette, Nauen, 1983; Collette et al., 1984 и др.). Систематика, построенная на молекулярных данных, противоречит подобной классификации, почти все трибы рассматриваются как парафилетические (Collette et al., 2001). Исходя из некоторых современных данных, единственной непарафилитической группой скумбриевых считается лишь триба Scombrini, включающая в себя род Scomber и сестринский род Rastrelliger (Santini et al., 2013). К скумбриям рода Scomber относят от двух (Fraser-Brunner, 1950) до четырех (Collette, 1999; Froese, Pauly, 2019) современных видов и около 11 ископаемых видов: S. antiquus Heckel in Heckel et Kner, 1861; S. calabrensis Landini et Bannikov, 1983; S. caucasicus (Bogatshov, 1933); S. cubanicus Daniltshenko, 1960; S. gnarus Bannikov, 1979; S. nomurai Niino, 1951; S. priscus Kramberger-Gorjanović, 1882; S. saadii Arambourg, 1967; S. sanctaemonicae (Jordan, 1919); S. susedanus Steindachner, 1860 и S. voitestii Paucă, 1929, не считая не определенные до вида экземпляры ископаемых Scomber (напр., Nazarkin, Bannikov, 2014). Кроме того, несколько миоценовых форм скумбриевых, описанных с территории бывшей Югославии (Хорватия, Словения) как относящиеся к роду Auxis (Kramberger-Gorjanović, 1882; Gorjanović-Kramberger, 1895; Anđelković, 1989), на самом деле должны быть отнесены к Scomber (Банников, 1985; Nam et al., 2021). Большинство из перечисленных выше видов скумбрий, кроме позднеэоценового иранского S. saadii, миоценовых японского S. nomurai и калифорнийского S. sanctaemonicae, а также плиоценового средиземноморского S. calabrensis, обитали в олигоцене–среднем миоцене в Центральном и Восточном Паратетисе. До недавнего времени Scombridae не были известны из морских тарханско-чокракских (низы среднего миоцена) отложений Восточного Паратетиса, однако в ходе полевых работ Палеонтологического ин-та им. А.А. Борисяка (ПИН) РАН (2018–2021 гг.) на местонахождении тарханских рыб на северо-западе Кавказа (Краснодарский край, р. Пшеха) были обнаружены первые скумбриевые. Изучение находок показало, что они представляют пеламиду Sarda sp. и новый вид скумбрий Scomber collettei sp. nov., описание которого приведено ниже. Местонахождение на р. Пшеха является самым богатым из достоверно тарханских местонахождений рыб Восточного Паратетиса (Банников, 2020а).
При всей противоречивости современной систематики подотряда Scombroidei, парафилетического по некоторым данным молекулярной систематики, семейство Scombridae обычно считается монофилетическим. Хотя эволюционные взаимоотношения таксонов более низкого ранга внутри семейства пока остаются под вопросом, триба Scombrini выглядит монофилетической как по морфологическим признакам (Банников, 1985, 2010; Monsch, Bannikov, 2012), так и по данным молекулярной систематики (Miya, 2013; Santini et al., 2013; Friedman et al., 2019).
По сравнению с другими трибами (или подсемействами) скумбриевых, Scombrini выглядят несколько генерализованными, имея меньшее число позвонков (всегда 31, против 32–64 у скумбриевых других триб) и две гипуральные пластинки (кроме Grammatorcynus, у которого они срастаются). В отличие от эволюционно продвинутых Sardini и Thunnini, у Scombrini нет костного киля на хвостовом стебле и “корсета” из увеличенных чешуй. Два современных рода Scombrini, Scomber и Rastrelliger, образуют сестринскую пару. В отличие от эоцен–олигоценового Auxides (=Scombrosarda) и современного Grammatorcynus, у Scomber и Rastrelliger спинные плавники не слиты, а широко разделены. Перешедший к планктоноядности род Rastrelliger отличается от Scomber более высоким и сжатым с боков телом, более мелкими зубами в челюстях и их отсутствием на vomer и palatinum, а также сильным развитием жаберных тычинок.
МАТЕРИАЛ И МЕТОДЫ
Материалом для работы послужили девять отпечатков скелета рыб стандартной длиной тела (SL) 12.5–>90 мм из тонкослоистых глин аргунских слоев тарханского региояруса, собранных на правом берегу р. Пшеха (Краснодарский край) выше ст. Ширванской. Пиритизация, в разной степени выраженная в образовании обильных фрамбоидов сульфида железа по органическим тканям скелетов скумбрий, свидетельствует о периодическом формировании бескислородного придонного слоя в данном бассейне (Liu et al., 2019).
Образцы изучались с помощью бинокулярных микроскопов Leica M165C, МБС-9 и Микромед MC2 ZOOM 1CR, а также сканирующего микроскопа TESCAN VEGA в ПИН РАН. В качестве сравнительного материала использовались образцы ранее описанных ископаемых видов скумбриевых рыб из колл. ПИН РАН, а также остеологические коллекции и рентгеноснимки скелетов современных скумбрий. Фотографии выполнены С.В. Багировым (ПИН РАН) (рис. 1 и табл. X , см. вклейку) и Р.А. Ракитовым (ПИН РАН) (рис. 2). Замечания рецензента М.В. Назаркина позволили улучшить текст статьи.
Рис. 1.
Scomber collettei sp. nov., полные скелеты: а – паратип ПИН, № 5599/7; б – паратип ПИН, № 5599/8; Краснодарский край, Апшеронский р-н, р. Пшеха выше ст. Ширванской; низы среднего миоцена, тархан. Масштабная линейка – 1 см.

Рис. 2.
Scomber collettei sp. nov.: а – экз. ПИН, № 5599/10, передняя часть нижней челюсти, СЭМ; б – экз. ПИН, № 5599/11, передняя часть черепа малька, СЭМ. Масштабная линейка – 0.5 мм.

Исследование выполнено за счет гранта Российского научного фонда № 22-27-00162, https://rscf.ru/project/22-27-00162/. Полевые раскопки 2019–2021 гг. производились при финансовой поддержке грантов PalSIRP Sepkoski (США).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО SCOMBRIDAE RAFINESQUE, 1815
Род Scomber Linnaeus, 1758
Scomber collettei Bannikov et Erebakan, sp. nov.
Табл. X , фиг. 1, 2
Название вида – в честь известного эксперта по современным скумбриевым Б. Коллетта (Национальная служба морского рыболовства, Смитсоновский ин-т, Вашингтон, США).
Голотип – ПИН, № 5599/6, двусторонний отпечаток скелета без хвостового плавника и передней части черепа (табл. X , фиг. 1); Краснодарский край, Апшеронский р-н, правый берег р. Пшеха выше ст. Ширванской (44.368442° N; 39.796874° E); низы среднего миоцена, тарханский региоярус.
Описание (рис. 1–3). Тело удлиненное, веретеновидное, с тонким, коротким хвостовым стеблем; вероятно, почти не сжатое с боков (на что косвенно указывает захоронение крыши черепа паратипа ПИН, № 5599/8 не латерально, а в дорсо-вентральном положении; рис. 1, б). Максимальная высота тела совпадает с высотой головы. Длина головы (HL) 3–3.1 раза укладывается в стандартной длине тела (SL) и в 1.6–1.7 раза превосходит наибольшую высоту тела. HL примерно соответствует длине 14 передних хвостовых позвонков. Глаза относительно крупные, горизонтальный диаметр орбиты почти равен длине рыла и в 1.4 раза меньше заглазничного расстояния. Длина рыла примерно равна длине четырех передних хвостовых позвонков.
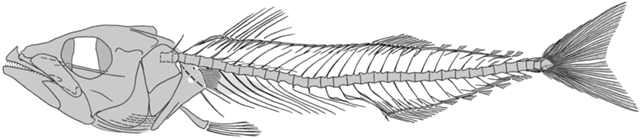
Рис. 3.
Scomber collettei sp. nov., реконструкция скелета по голотипу и паратипам, чешуя не показана.
Голова коническая, со слегка заостренным рылом. Высота головы в 1.6–1.7 раза уступает ее длине. Передний конец нижней челюсти слегка выступает вперед. Окостенение костей черепа относительно слабое (рис. 2, а). Нейрокраний довольно низкий; тонкий, почти прямой парасфеноид проецируется в нижней половине орбиты. Гребень supraoccipitale низкий и относительно короткий. Frontalia в дорсо-вентральном аспекте относительно широкие над орбитой, плавно сужаются кпереди; при этом их сейсмосенсорные каналы слегка расходятся (рис. 1, б). Frontale (возможно, и pterosphenoideum) выступают вниз в орбиту. Этмоидный район черепа невысокий, удлиненный. Кости ушного отдела черепа не различимы; отолит сохранился лишь у паратипа ПИН, № 5599/8, где видна только его наружная выпуклая сторона, а данных о sulcus acusticus нет. Окостенения склеротики занимают всю орбиту, состоят из передней и задней полукруглых частей. Подглазничные кости не видны, кроме lacrimalia, смещенных из их естественного положения на паратипе ПИН, № 5599/9 (табл. X , фиг. 2). Lacrimale довольно плотное, округлое сзади и тупо приостренное антеровентрально, со слегка вогнутым нижним краем. Lacrimale крупное, его длина лишь едва уступает длине верхней челюсти. Высота lacrimale втрое меньше его длины.
Рот умеренно крупный, конечный; нижнечелюстное сочленение расположено примерно под серединой орбиты. Рот явно не выдвижной: кости обеих ветвей верхней челюсти, смещенных из их естественного положения на паратипе ПИН, № 5599/9, находятся в сочленении. Восходящий отросток praemaxillare умеренной высоты, слит с сочленовным отростком. Альвеолярная ветвь praemaxillare почти прямая, постепенно утоньшается каудально, несет ряд из примерно 30 зубов. Зубы конические, крупные относительно других представителей рода Scomber, почти прямые, с немного загнутой внутрь вершиной (рис. 2). Maxillare довольно широкое, на трети своей длины немного загнуто задним концом вниз. Задний конец maxillare расширен книзу в форме лопатки. Нижняя челюсть невысокая, ее высота в 3.5–3.7 раза уступает длине. Длина нижней челюсти составляет около 0.58 длины головы и превышает длину семи последних туловищных позвонков. Симфизная часть нижней челюсти невысокая, слегка отогнута книзу, но без подбородочного выступа. Dentale V-образное, в его заднюю вырезку входит angulo-articulare. Верхний задний отросток dentale примерно одной длины с нижним, но значительно его у́же. Зубы на dentale однорядные, сходны с верхнечелюстными по форме и размеру; их не менее 25 у наиболее крупных экземпляров. Angulo-articulare по площади немного меньше, чем dentale; имеется промежуток между верхними отростками этих костей. Сочленовный отросток аngulo-articulare короткий, retroarticulare не различимо. Hyomandibulare имеет форму перевернутой буквы L, с удлиненным передним мыщелком (для сочленения со sphenoticum); ось кости очень слабо наклонена вперед от вертикали. Quadratum относительно широкое, субтреугольное, с небольшим сочленовным мыщелком. От palatinum различима лишь передняя часть на паратипе ПИН, № 5599/9, где кость выявляет крепкий отросток для сочленения с maxillare. Жаберная крышка умеренно широкая. Praeoperculum довольно крупное, плоское, полулунной формы: выпуклое сзади, вогнутое спереди. Нижняя ветвь предкрышки шире и немного короче верхней ветви. Передненижний край кости образует с передневерхним краем угол около 120°. Свободный край предкрышки ровный у крупных экземпляров, у мальков этот район сохранился плохо. Operculum крупное, плоское, широкое; его нижний край приострен, а в постеродорсальной части имеется небольшая выемка. Кости гиоидного комплекса и жаберного аппарата сохранились плохо; точное число тонких radii branchiostegi неизвестно. На нескольких экземплярах частично сохранились минерализованные жаберные лепестки и тычинки, а на голотипе – также тонкие, заостренные, слегка изогнутые глоточные зубы.
Позвонков 31, из них 14 – туловищные и 17 – каудальные. Длина туловищной части позвоночника в 1.11–1.23 раза короче хвостовой его части. Линия позвоночника слегка изогнута, приподнята спереди. Тела позвонков продольно вытянутые в латеральном аспекте, пережатые посередине, несут по латеральному гребню с каждой стороны. Презигапофизы развиты сильнее постзигапофизов. Большинство остистых отростков очень тонкие и относительно длинные, прямые или слегка изогнутые. Из невральных отростков наименьший угол наклона назад имеют таковые передних хвостовых позвонков. Гемальный отросток первого хвостового позвонка слегка утолщен и изогнут в средней части. Парапофизов не менее четырех, они удлиняются назад в ряду. Два последних парапофиза длинные, изогнутые посередине и с расширенной медиально дистальной частью. Ребра тонкие и длинные, сильно отклонены назад, передние и средние из них почти доходят до брюшного края тела; два последних ребра короткие. Ребра начинаются со второго позвонка. Тонкие epineuralia различимы у оснований невральных отростков туловищных позвонков.
Хвостовой скелет почти полностью скрыт гипуростегией (раздвоенными основаниями лучей хвостового плавника). Тела позвонков хвостового стебля не несут костного латерального киля. Второй преуральный позвонок укорочен. Гипуральная пластинка подразделена на эпаксиальную и гипаксиальную части, разделенные швом. Хвостовой плавник относительно небольшой, глубоко вырезанный, состоит из 17 главных лучей (I8–7I). Верхние и нижние дополнительные лучи относительно многочисленные, их по девять с каждой стороны.
Posttemporale – плоская кость с двумя относительно тонкими передними отростками, связывающими грудной пояс с нейрокранием. Cleithrum крупное, почти прямое, с загнутым вперед верхним концом. Границы coracoideum, scapula и пекторальных radialia различимы плохо. Postcleithrum тонкое, длинное, ребровидное, заканчивается у вентрального края тела позади тазовых костей. Грудной плавник среднего размера, его полная длина неясна. Точное число лучей в грудном плавнике неизвестно, но их не менее 18–20. Основание грудного плавника расположено высоко, сразу под линией позвоночника, против четвертого–пятого позвонка.
Тазовые кости узкие, клиновидные, ориентированы под углом около 45° к оси тела. Брюшные плавники умеренно длинные, прикрепляются под грудными, каждый состоит из слабой колючки и пяти мягких лучей.
Спинные плавники широко разделены – на расстояние, равное длине шести противолежащих позвонков, и в 1.4 раза превышающее длину основания первого спинного плавника. Первый спинной плавник начинается над пятым позвонком; в нем девять тонких колючек, третья или четвертая из которых самая длинная (в 1.6–1.7 раза длиннее первой колючки). Четыре последних шипа быстро укорачиваются. Высота плавника превышает длину его основания, но существенно меньше высоты тела под ним. Первая колючка сверхштатная на переднем птеригиофоре. Птеригиофоров первого спинного плавника восемь; они узкие, клиновидные, с косо ориентированной проксимальной частью. Девять свободных, не несущих лучей птеригиофоров расположены между спинными плавниками, образуя непрерывный ряд дорсальных птеригиофоров.
Второй спинной плавник начинается на вертикали второго-третьего хвостового позвонка, в нем один жесткий и 11 довольно коротких мягких лучей. Лучи расположены очень тесно, в задней части плавника несколько более разрежены. Передний мягкий луч самый длинный, каудально лучи укорачиваются. Высота второго спинного плавника не превышает длину его основания. Дорсальных дополнительных плавничков шесть. Птеригиофоры второго спинного плавника очень тонкие. В каудальном направлении птеригиофоры постепенно укорачиваются. Межневральные промежутки под вторым спинным плавником обычно вмещают проксимальные концы двух птеригиофоров каждый. У птеригиофоров дополнительных плавничков дистальная часть сопоставима по длине с проксимальной, при этом кзади птеригиофоры распрямляются.
Анальный плавник сходен со вторым спинным по форме и размерам, но немного сдвинут назад относительно начала второго спинного. Длина его основания уступает длине четырех противолежащих позвонков. В анальном плавнике две колючки (передняя, короткая, но крепкая – сверхштатная) и 11 мягких лучей. Передний ветвящийся луч самый длинный, каудально лучи укорачиваются. Вентральных дополнительных плавничков шесть. Птеригиофоры анального плавника очень тонкие, сходны по форме с противолежащими птеригиофорами второго спинного плавника и также укорачиваются спереди назад. Первый анальный птеригиофор слегка вогнут спереди. Проксимальные концы двух–трех птеригиофоров входят в межгемальные промежутки над анальным плавником. Птеригиофоры нижних дополнительных плавничков сходны с противолежащими птеригиофорами верхних дополнительных плавничков.
Чешуя очень мелкая, тонкая, циклоидная, несет circuli, образует сплошной покров на теле. Боковая линия не прослеживается.
Следов первоначального рисунка пигментации на теле или плавниках достоверно не выявлено, но пигментация брюшины хорошо выражена.
Размеры. SL целых экземпляров – 12.5–>90 мм. В процентах к SL паратипа ПИН, № 5599/7 (SL 77 мм): длина головы – 32.5; максимальная высота тела – 20; минимальная высота тела – 3.5; первое предорсальное расстояние – 38; второе предорсальное расстояние – 65; преанальное расстояние – 68; высота первого спинного плавника – 13; длина второго шипа анального плавника – 2.5; длина брюшного плавника – 10; длина наибольших лучей хвостового плавника – 13; горизонтальный диаметр орбиты – 9; предглазничное расстояние – 9.5; заглазничное расстояние – 14; длина нижней челюсти – 18.5.
Возрастные изменения. У мальков относительная длина головы существенно больше, чем у взрослых рыб (2.7 раза укладывается в SL), а голова более высокая; с возрастом уменьшается относительная длина зубов, а их число увеличивается.
Сравнение. Неполнота сведений о морфологии некоторых миоценовых видов Scomber [S. antiquus, S. sanctaemonicae и S. caucasicus, а также виды, первоначально описанные (Kramberger-Gorjanović, 1882; Gorjanović-Kramberger, 1895) в составе рода Auxis] до дополнительного их изучения не позволяет адекватно сравнить их с новым видом. S. collettei sp. nov. отличается от многих современных и ископаемых видов рода Scomber соотношением числа позвонков в туловищном и хвостовом отделах (14 + 17 против 13 + 18 у S. scombrus и S. saadii, и 15 + 16 у S. cubanicus, S. gnarus, S. nomurai и S. priscus) (см. табл. 1). От большинства скумбрий, имеющих по пять верхних и нижних дополнительных плавничков (кроме S. saadii, S. voitestii и S. cubanicus), новый вид отличается наличием шести дополнительных плавничков сверху и снизу. Девять колючек в первом спинном плавнике, как у S. collettei sp. nov., у скумбрий бывает довольно редко – обычно их 10 или больше, реже восемь или семь (у S. voitestii и S. calabrensis). Спинные плавники нового вида разделены сильнее, чем у большинства других видов, примерно как у S. scombrus и S. calabrensis; свободных птеригиофоров между спинными плавниками у других видов обычно менее девяти. У S. collettei sp. nov. отчетливо различимы два шипа в анальном плавнике (как у S. voitestii, S. cubanicus и S. gnarus), тогда как у большинства других видов имеется лишь одна анальная колючка.
Таблица 1.
Меристические признаки современных и большинства известных ископаемых (отмечены †) видов скумбрий (по: Kramberger-Gorjanović, 1882; Niino, 1951; Arambourg, 1967; Landini, Bannikov, 1983; Банников, 1985; Froese, Pauly, 2019)
| Название таксона | Число позвонков в туловищном и хвостовом отделах | ID | IID | A |
|---|---|---|---|---|
| Scomber scombrus L. | 13 + 18 | X–XIV | I, 12–13 + 5 | I, 12–13 + 5 |
| S. colias Gmelin | 14 + 17 | IX–X | I, 12 + 5 | I, 12–14 + 5 |
| S. japonicus Houttuyn | 14 + 17 | IX–X | I, 12 + 5 | I, 12–14 + 5 |
| S. australasicus Cuvier | 14 + 17 | X–XIII | 12 + 5 | 12 + 5 |
| †S. saadii Arambourg | 13 + 18 | VIII–X | ? | 12–13 + 6–7 |
| †S. voitestii Paucă | 14 + 17 | VIII | I, 10–11 + 6 | II, 11–13 + 6 |
| †S. cubanicus Daniltshenko | 15 + 16 | IX–X | I, 9–10 + 6 | II, 10-11 + 6 |
| †S. nomurai Niino | 15 + 16 | XI | I, 13 + 5 | I, 11 + 5 |
| †S. gnarus Bannikov | 15 + 16 | XI–XII | I, 11 + 5 | II, 10–11 + 5 |
| †S. priscus Kramberger-Gorjanović | 15? + 16 | X–XI | 10–12 + 5 | 12 + 5 |
| †S. susedanus Steindachner | 14? + 17 | X–XI | 11 + 5 | 12 + 5 |
| †S. calabrensis Landini et Bannikov | 14 + 17 | VII | 12 + 5 | I, 12 + 5 |
| †S. collettei sp. nov | 14 + 17 | IX | I, 11 + 6 | II, 11 + 6 |
Замечания. В фауне рыб тарханского комплекса с р. Пшеха пока не отмечено эласмобранхий, а из костистых рыб доминируют либо таксоны мелкого размера (светящаяся рыба Vinciguerria merklini Dan.), либо мальки других видов (Банников, 2020а). Сходную по размерному составу среднеэоценовую ихтиофауну Месселя (ФРГ) Н. Миклих назвал “детским садом” рыб (Micklich, 2002). Тарханские скумбриевые представлены как мальками, так и взрослыми рыбами небольшого размера (молодью?). Причем если у нового вида Scomber преобладают юные взрослые экземпляры, то хищники-пеламиды Sarda sp. известны почти исключительно по малькам.
Материал. 9 экз. из типового местонахождения, колл. ПИН, № 5599.
Список литературы
Банников А.Ф. Ископаемые скумбриевые СССР // Тр. Палеонтол. ин-та АН СССР. 1985. Т. 210. 111 с.
Банников А.Ф. Ископаемые позвоночные России и сопредельных стран. Ископаемые колючеперые рыбы (Teleostei, Acanthopterygii). М.: ГЕОС, 2010. LXI + 244 с.
Банников А.Ф. Новое местонахождение тарханских (нижний–средний миоцен) костистых рыб на Северо-Западном Кавказе // Биогеография и эволюционные процессы. Матер. LXVI сессии Палеонтол. об-ва при РАН. СПб.: ВСЕГЕИ, 2020а. С. 214–216.
Банников А.Ф. Новый род и вид скумбриевых рыб (Perciformes, Scombroidei, Scombridae) из нижнего олигоцена Кавказа // Палеонтол. журн. 2020б. № 1. С. 56–65.
Берг Л.С. Система рыбообразных и рыб, ныне живущих и ископаемых // Ежегодн. Зоол. музея АН СССР. 1940. Т. 5. С. 87–517.
Anđelković J.S. Tertiary fishes of Yugoslavia. A stratigraphic-paleontologic-paleoecological study // Palaeontol. Jugosl. 1989. Sv. 38. S. 1–121.
Arambourg C. Résultats scientifiques de la Mission C. Arambourg en Syrie et en Iran (1938–1939). II. Les poissons oligocènes de l’Iran // Notes Mém. Moyen-Orient. 1967. T. 8. P. 9–210.
Betancur-R.R., Broughton R.E., Wiley E.O. et al. The tree of life and a new classification of bony fishes // PLOS Currents Tree of Life. 2013. P. 1–41.
Betancur-R.R., Wiley E.O., Arratia G. et al. Phylogenetic classification of bony fishes // BMC Evol. Biol. 2017. 17:162. P. 1–40. https://doi.org/10.1186/s12862-017-0958-3
Collette B.B. Mackerels, molecules, and morphology // Proc. 5th Indo-Pac. Fish Conf., Nouméa, 1997 / Ed. Séret B., Sire J.-Y. P.: Soc. Fr. Ichtyol., 1999. P. 149–164.
Collette B.B., Nauen C.E. FAO species catalogue. Vol. 2. Scombrids of the World. An annotated and illustrated catalogue of tunas, mackerels, bonitos, and related species known to date. Rome: FAO, 1983. VII + 137 p. (FAO Fish. Synop. № 125. V. 2).
Collette B.B., Potthoff T., Richards W.J. et al. Scombroidei: development and relationships // Ontogeny and systematics of fishes / Eds. Moser H.G., Richards W.J., Cohen D.M. et al. Lawrence: Allen Press, 1984. P. 591–620 (Amer. Soc. Ichthyol. Herpetol. Spec. Publ. № 1).
Collette B.B., Reeb C.A., Block B.A. Systematics of the tunas and mackerels (Scombridae) // Tuna: Physiology, Ecology, and Evolution / Eds. Block B.A., Stevens E.D. San Diego: Acad. Press, 2001. P. 1–33. (Ser. Fish Physiol. V. 19).
Fraser-Brunner A. The fishes of the family Scombridae // Ann. Mag. Natur. Hist. Ser. 12. 1950. V. 3. № 26. P. 131–163.
Friedman M., Feilich K.L., Beckett H.T. et al. A phylogenomic framework for pelagiarian fishes (Acanthomorpha: Percomorpha) highlights mosaic radiation in the open ocean // Proc. Roy. Soc. B. 2019. V. 286: 20191502. https://doi.org/10.1098/rspb.2019.1502
Froese R., Pauly D. (eds.). FishBase. 2019. World Wide Web electronic publication. www.fishbase.org, version (12/2019).
Gorjanović-Kramberger D. Fosilne ribe Komena, Mrzleka, Hvara i M. Libanona i dodatak o oligocenskim ribama Tüffera, Zagora i Trifalja // Djela Jugosl. Akad. Znan. Umjetn. 1895. V. 16. S. 1–67.
Kramberger-Gorjanović D. Die jungtertiäre Fischfauna Croatiens // Beitr. Paläontol. Geol. Österr.-Ungarns Orients. 1882. Bd 2. S. 86–136.
Landini W., Bannikov A.F. Scomber calabrensis (Osteichthyes, Scombridae), nuova specie del Plioceno italiano // Atti Soc. Toscana Sci. Natur. Mem. Ser. A. 1983. V. 90. P. 191–198.
Liu Z., Chen D., Zhang J. et al. Pyrite morphology as an indicator of paleoredox conditions and shale gas content of the Longmaxi and Wufeng shales in the middle Yangtze area, South China // Minerals. 2019. V. 9. № 7: 428. https://doi.org/10.3390/min9070428
Micklich N. The fish fauna of Messel Pit: a nursery school? // Cour. Forsch.-Inst. Senckenb. 2002. Bd 237. P. 97–127.
Miya M., Friedman M., Satoh T.P. et al. Evolutionary origin of the Scombridae (tunas and mackerels): Members of a Paleogene adaptive radiation with 14 other pelagic fish families // PLoS One. 2013. V. 8. № 9. P. e73535: 1–19.
Monsch K.A., Bannikov A.F. New taxonomic synopses and revision of the scombroid fishes (Scombroidei, Perciformes), including billfishes, from the Cenozoic of territories of the former USSR // Earth and Environ. Sci. Trans. Roy. Soc. Edinburgh. 2012 (2011). V. 102. P. 253–300.
Nam G.-S., Nazarkin M.V., Bannikov A.F. First discovery of the genus Auxis (Actinopterygii: Scombridae) in the Neogene of South Korea // Boll. Soc. Paleontol. Ital. 2021. V. 60. № 1. P. 61–67. https://doi.org/10.4435/BSPI.2021.05
Nazarkin M.V., Bannikov A.F. Fossil mackerel (Actinopterygii: Scombridae: Scomber) from the Neogene of south-western Sakhalin, Russia // Zoosyst. Ross. 2014. V. 23. № 1. P. 158–163.
Near T.J., Dornburg A., Eytan R.I. et al. Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes // Proc. Natl. Acad. Sci. USA. 2013. V. 110. P. 12738–12743.
Near T.J., Eytan R.I., Dornburg A. et al. Resolution of ray-finned fish phylogeny and timing of diversification // Proc. Natl. Acad. Sci. USA. 2012. V. 109. P. 13698–13703.
Nelson J.S. Fishes of the World. 4th ed. Hoboken, NJ: Wiley, 2006. 601 p.
Nelson J.S., Grande T.C., Wilson M.V.H. Fishes of the World. 5th ed. Hoboken, NJ: Wiley, 2016. 707 p.
Niino H. Description of some fossil fishes and prawns from Japan // J. Tokio Coll. Fish. 1951. V. 38. № 1. P. 47–58.
Santini F., Carnevale G., Sorenson L. First molecular scombrid timetree (Percomorpha: Scombridae) shows recent radiation of tunas following invasion of pelagic habitat // Ital. J. Zool. 2013. V. 80. P. 210–221.
Starks E.C. The osteology and mutual relationships of the fishes belonging to the family Scombridae // J. Morphol. 1910. V. 21. P. 77–100.
Wiley E.O., Johnson G.D. A teleost classification based on monophyletic groups // Origin and Phylogenetic Interrelationships of Teleosts / Eds Nelson J.S., Schultze H.-P., Wilson M.V.H. Munich: Verl. Dr. F. Pfeil, 2010. P. 123–182.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал