

Палеонтологический журнал, 2022, № 4, стр. 59-72
Муравьи (Hymenoptera: Formicidae) из местонахождений Амгу и Великая Кема (Дальний Восток, Россия)
К. С. Перфильева *
Московский государственный университет имени М.В. Ломоносова
119991 Москва, Россия
* E-mail: ksenperf@mail.ru
Поступила в редакцию 15.09.2021
После доработки 10.01.2022
Принята к публикации 28.02.2022
- EDN: VCPJPT
- DOI: 10.31857/S0031031X22040080
Аннотация
В коллекции Палеонтологического ин-та им. А.А. Борисяка РАН имеются отпечатки изолированных крыльев муравьев из кайнозойских местонахождений с территории России, собранные в разное время. Отпечатки из местонахождений Амгу (один отпечаток переднего крыла) и Великая Кема (16 крыльев) [Дальний Восток, нижний олигоцен–начало среднего миоцена (?)] описаны впервые и принадлежат представителям Dolichoderinae (Emplastus amguensis sp. nov., E. obliquus sp. nov., E. similis sp. nov., Technomyrmex kemaensis sp. nov.; 6 экз.), Formicinae (Camponotus parvus sp. nov., Lasius vulgaris sp. nov.; 4 экз.), Myrmicinae (Paraphaenogster ussuriensis sp. nov.; 3 экз.), а также Formicidae incertae sedis (4 экз.). Отмечается одновременное присутствие элементов мирмекокомплексов умеренного климата и тропических теплолюбивых видов, а также значительная доля дендрофильных видов.
ВВЕДЕНИЕ
В коллекции лаборатории артропод Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН), собранной в разное время из разных географических точек, имеются отпечатки изолированных крыльев Formicidae. Поскольку для рецентных муравьев крыловые признаки практически не применяются в качестве таксономических, а их изменчивость довольно слабо изучена, то описание отпечатков отдельных крыльев и определение их таксономической принадлежности является довольно сложной задачей. Наши оригинальные исследования и обобщение информации других авторов позволяют утверждать, что крылья муравьев имеют не меньшую таксономическую ценность, чем для других хорошо летающих групп перепончатокрылых (Brown, Nutting, 1949; Перфильева, 2000, 2010, 2011, 2021). Поэтому нам представляется актуальным и адекватным описание имеющихся отпечатков изолированных крыльев муравьев для предварительной оценки таксономического состава мирмекофаун местонахождений, где таковые встречаются.
МАТЕРИАЛ И МЕТОДЫ
В настоящем исследовании описываются один отпечаток переднего крыла из местонахождения Амгу (Приморский край, Татарский пролив, р. Амгу, Восточный Сихотэ-Алинь) и отпечатки передних и задних крыльев 16 экземпляров из местонахождения Великая Кема (Приморский край, Тернейский р-н, пос. Великая Кема, р. Кема, Восточный Сихотэ-Алинь), имеющиеся в коллекции лаборатории артропод ПИН. Оценка возраста местонахождений по описанию флористических комплексов неодинакова у разных авторов. Кизинская свита (Великая Кема) в региональной стратиграфической схеме соответствует геохронологическому интервалу поздний олигоцен – средний миоцен, но некоторые исследователи считают эту свиту невалидным стратоном (Павлюткин и др., 2014). Флористический комплекс соответствует так называемой “энгельгардиевой” флоре, которую долгое время относили к среднему миоцену. Однако наиболее поздние исследования флористических комплексов «удревнили» датировки местонахождений Амгу и Великая Кема до раннего олигоцена. Мы приводим обе точки зрения, а в описаниях указываем новейшие датировки.
Согласно флористическому анализу Д.А. Лопатиной (2004), возраст местонахождения Амгу оценивается как конец позднего олигоцена–начало раннего миоцена, местонахождение Великая Кема – вторая половина раннего–начало среднего миоцена (рис. 1). Ранее из этого региона было описано богатое отпечатками муравьев местонахождение Большая Светловодная позднеэоценового возраста (рис. 1) (Dlussky et al., 2015). По данным Лопатиной, климат и флористическая обстановка в этом регионе заметно менялись во временном промежутке эоцен–средний миоцен. Флора Большой Светловодной соответствует смешанным мезофитным лесам с доминированием листопадных широколиственных родов с высокими значениями термофильности и коэффициента влажности флоры (субтропические и тропические растительные компоненты преобладают), что указывает на довольно высокие среднегодовые температуры: +10…+13°С. Похолодание и иссушение климата на рубеже эоцена–олигоцена в этой области довольно ярко выражено и отражается в замещении формальных родов цветковых на современные. Более поздняя флора Амгу (поздний олигоцен–ранний миоцен) отличается доминированием хвойных, значительную роль играли также таксодиевые, с примесью ильмовых, березовых. По данным эколого-флористического анализа Лопатиной, климат был менее жарким (среднегодовые температуры +3…+10°С), чем в Светловодной: похолодание на рубеже олигоцена–миоцена еще более ярко выражено, чем на границе эоцена–олигоцена. Снижение роли гигро- и гидрофитов и увеличение доли мезофитов говорит о заметном понижении влажности. Местонахождение Великая Кема относится ко времени проявления миоценового температурного оптимума (конец раннего–начало среднего миоцена). Для него характерны хвойно-широколиственные леса с заметной ролью бука, таксодиевых и разнообразных вечнозеленых. Значительна доля субтропических и тропических элементов растительности (хурма, магнолия, конский каштан). Прослеживается сходство растительности миоценового термального оптимума (Великая Кема) с лесами этого региона в период позднего эоцена–раннего олигоцена (Большая Светловодная), что соответствует климатическим условиям со среднегодовыми температурами +10… +13°С (Лопатина, 2004).
Рис. 1.
Расположение изученных местонахождений: 1 – Амгу, 2 – Большая Светловодная, 3 – Великая Кема (по: Лопатинa, 2004).
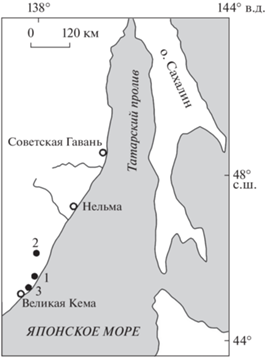
В относительно недавних работах палеоботаников местонахождения Амгу и Великая Кема (Ключ Тихий) признаются раннеолигоценовыми (рюпель), при этом флористический состав этих местонахождений несколько различен (Павлюткин и др., 2014; Bondarenko et al., 2019). Несмотря на существенную разницу в датировках местонахождений у Лопатиной и приведенных авторов, оценки флористической обстановки сопоставимы у всех авторов. Флористический анализ макрофоссилий О.В. Бондаренко с соавт. (Bondarenko et al., 2019) предполагает наличие широколиственного листопадного леса с высокой долей хвойных (до 25%) и участием вечнозеленых субтропических компонентов. Флора Амгу имеет большее участие вечнозеленых субтропических растений (18% против 8.3% в Ключе Тихом), и авторы относят ее к переходному экотону: от широколиственного теплоумеренного к субтропическому смешанному лесу. Широколиственный листопадный лес Великой Кемы отнесен авторами к теплоумеренному типу растительности.
Таким образом, все авторы отмечают наличие в период образования отложений в местонахождениях Амгу и Великая Кема мезофитного широколиственного листопадного леса с высокой долей хвойных и существенным участием субтропических вечнозеленых компонентов. Представляется, что флора носила характер современных горных субтропических лесов без участия равнинных элементов (отсутствие трав). Климат в это время характеризовался довольно высокими температурами летнего периода со значительным понижением зимних, т.е., наблюдается увеличение сезонности по отношению к эоценовому периоду.
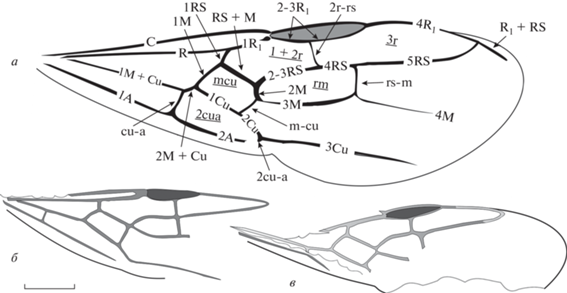
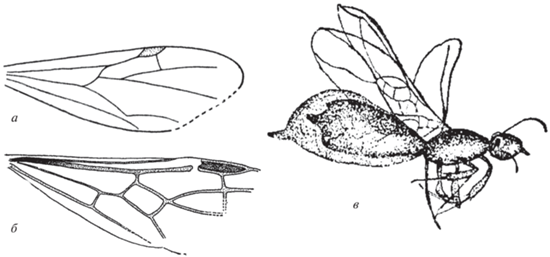
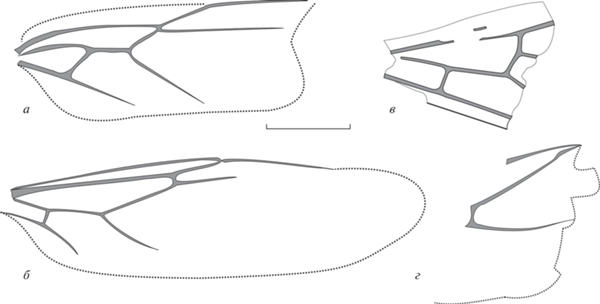
Номенклатура жилкования переднего крыла муравьев согласно А.П. Расницыну (1980) представлена на рис. 2, а. Сокращения названий используемых в тексте измерений и расшифровка индексов: FWL – длина переднего крыла (forewing length); FWPt – расстояние между основаниями крыла и птеростигмы; Icu = [1Cu + (2M + + Cu)]/1Cu – отражает положение поперечной жилки cu-a относительно медиокубитальной (mcu) ячейки; Icua = [(1M + Cu) + (2M + + Cu)]/(1M + Cu) – отражает положение поперечной жилки cu-a относительно основания крыла; I(mcu) = (высота ячейки mcu)/(длина средней линии ячейки mcu) – отражает относительную высоту медиокубитальной ячейки [I(mcu)=1 указывает на близость формы ячейки к квадрату; >1 соответствует высокой и короткой ячейке, <1 – низкой и длинной]. Индекс I(mcu) адекватно применять только для ячеек, в которых RS+M приблизительно параллельна 1Cu – трапециевидных, прямоугольных, квадратных, типа параллелограмма. Индекс радиальных ячеек (Ir) – соотношение длин ячеек 1+2r и 3r.
Рис. 2.
а – Номенклатура жилкования переднего крыла муравьев (по: Расницын, 1980); б – Emplastus amguensis sp. nov., голотип ПИН, № 3135/1, переднее крыло (зеркальное отражение); Амгу; в – E. obliquus sp. nov., голотип ПИН, № 3136/9, переднее крыло (зеркальное отражение); Великая Кема. Масштабная линейка – 1 мм.
ПАЛЕОНТОЛОГИЧЕСКОЕ ОПИСАНИЕ
СЕМЕЙСТВО FORMICIDAE LATREILLE, 1809
ПОДСЕМЕЙСТВО DOLICHODERINAE FOREL, 1878
Род (формальный) Emplastus Donisthorpe, 1920
Типовой вид – Dolichoderus britannicus Cockerell, 1915 (младший синоним: Emplastus emeryi Donisthorpe, 1920).
Диагноз (для крыльев). Отпечатки ископаемых муравьев с признаками принадлежности к подсемейству Dolichoderinae, сохранность которых не достаточна для описания нового или отнесения к известному ортотаксону, но имеющие определенный набор признаков. Передние крылья с тремя замкнутыми дистальными ячейками: 3r, rm и mcu. Вершина ячейки 3r лежит на краю крыла, отрезки 1RS и 2r-rs отходят от радиальной жилки под прямым углом или с небольшим уклоном. Медиокубитальная ячейка среднего размера, ее дистальный край не достигает уровня основания птеростигмы. Icu>1.45, Icua>1.2.
Видовой состав. На настоящий момент было описано 12 ископаемых видов (каталог AntWeb, август 2021). Из верхнего эоцена Дальнего Востока: E. macrops Dlussky, Rasnitsyn et Perfilieva, 2015, E. elongatus Dlussky, Rasnitsyn et Perfilieva, 2015, E. biamoensis (Perfilieva et Rasnitsyn, 2015); Великобритании: E. britannicus (Cockerell, 1915), E. hypolithus (Cockerell, 1915), E. gurnetensis (Donisthorpe, 1920), E. kozlovi Dlussky et Perfilieva, 2014; из миоценовых отложений Хорватии – E. miocenicus (Dlussky et Putyatina, 2014), E. antiquus (Mayr, 1867), E. dubius (Dlussky et Putyatina, 2014), E. haueri (Mayr, 1867), E. ocellus (Heer, 1849).
Замечания. Сходный тип жилкования имеют Aneuretinae, однако в крыльях их рецентных представителей ячейка 3r относительно большей длины по отношению к другим ячейкам и длине всего крыла, чем у долиходерин. Принадлежность крыла с подобным жилкованием к Aneuretinae на данный момент рационально трактовать только в сочетании с иными признаками тела, по причине того, что изменчивость жилкования целого подсемейства по одному (современному) виду оценить невозможно. Тип жилкования, представленный в этом формальном роде, может встречаться у представителей Liometopum, Dolichoderus, Azteca, Technomyrmex, Tapinoma, но жилкование крыльев представителей данных родов отличается разнообразной формой медиокубитальной ячейки, соотношением длин ячеек 1 + 2r и 3r и/или наличием участка 4RS в ячейке rm (когда rs-m расположена значительно дистальнее 2r-rs).
Жилкование крыльев Dolichoderus довольно изменчиво: имеются крылья от наиболее полного типа с пятиугольной радиомедиальной ячейкой и выраженными участками 2M и 4RS до треугольной ячейки rm со стебельком – участком 2RS + M; существенно варьируют форма и относительные размеры ячейки mcu. Необходимы дополнительные исследования для выявления общих особенностей жилкования, характерных для представителей этого рода.
Сходен с Emplastus тип жилкования у некоторых Myrmicinae. Однако современные мирмицины с подобным типом жилкования (Pheidole, Messor, Myrmica, Pogonomyrmex, Aphaenogaster), помимо обычно (но не всегда) незамкнутой ячейки 3r, имеют отличительные особенности, позволяющие отнести отпечатки их крыльев к ортотаксонам. Поэтому описание ископаемых мирмицин по отпечаткам тел муравьев с крыльями будет способствовать корректному отнесению отпечатков крыльев этого формального рода к Myrmicinae.
Emplastus amguensis Perfilieva, sp. nov.
Табл. X , фиг. 1 (см. вклейку)
Название вида от местонахождения отпечатка у р. Амгу.
Голотип – ПИН, № 3135/1, отпечаток переднего крыла; Приморский край, Татарский пролив, Тернейский р-н, пр. берег р. Кудья, притока р. Амгу; нижний олигоцен, гранатненская толща.
Описание (рис. 2, б). Птеростигма удлиненная. Радиомедиальная ячейка треугольная без выраженного участка 2M и без стебелька, либо стебелек экстремально короткий, практически не выражен; соотношение длины ячейки к ширине составляет около 2.5. Отрезки 2r-rs и rs-m находятся на одной прямой. Медиокубитальная ячейка средних размеров трапециевидная, m-cu и 1M почти одинаковой длины (m-cu > 1M в 1.1 раза), 1Cu в 1.3 раза длиннее RS+M. Ячейка mcu удлиненная, ее дистальный конец не достигает уровня птеростигмы. Вторая кубитоанальная ячейка не сформирована. Соотношение длин радиальных ячеек Ir = 0.8. Icu ~ 1.5, Icua ~ 1.3, I(mcu) ~ 0.5.
Размеры в мм. FWL~7.0. FWPt = 3.5, длина ячейки 3r = 2, длина ячейки rm = 1.1.
Сравнение. Треугольная без стебелька радиомедиальная ячейка имеется у трех видов этого рода: E. kozlovi (FWL 7.8–8.8 мм у самцов), E. haueri (FWL 5.4 мм), E. miocenicus (FWL 10.1). Последний, помимо значительно больших размеров, отличается от нового вида прямоугольной формой медиокубитальной ячейки. У E. haueri относительно короткая ячейка 3r и меньший размер крыла. Наибольшее сходство имеется с E. kozlovi из эоценовых мергелей Бембриджа, но относительно более длинная медиокубитальная ячейка у E. amguensis sp. nov. [I(mcu) 0.7 и 0.5 соответственно] имеет иное соотношение длин 1M и m-cu. Размеры крыльев у трех видов этого рода из Большой Светловодной несколько меньше, чем у нового вида (Dlussky et al., 2015).
Замечания. Жилкование крыла свидетельствует о принадлежности к Dolichoderinae. Хотя по отдельности признаки жилкования E. amguensis sp. nov. и E. kozlovi встречаются у современных представителей нескольких родов долиходерин, определенное сочетание признаков жилкования у этих отпечатков уникально [треугольная rm, отсутствие 4RS, трапециевидная mcu, соотношение 1M и m-cu, I(mcu)]. Наиболее сходное жилкование имеют некоторые современные Tapinoma (напр., Specimen CASENT0109245, онлайн каталог AntWeb.org), но при этом иная форма медиокубитальной ячейки и меньшее значение индекса радиальных ячеек – Ir = 0.5. Также можно отметить большое сходство с крылом Aneuretinae, однако, как отмечено выше, в отсутствие отпечатка тела, мы не рассматриваем этот вариант.
Материал. Голотип.
Emplastus obliquus Perfilieva, sp. nov.
Табл. X , фиг. 2
Название вида obliquus лат. – косой, по характерной форме медиокубитальной ячейки.
Голотип – ПИН, № 3136/9, отпечаток переднего крыла; Приморский край, Тернейский р-н, пос. Великая Кема; нижний олигоцен, кизинская свита.
Описание (рис. 2, в). Птеростигма удлиненная. Радиомедиальная ячейка четырехугольная без стебелька, имеется очень короткий участок 2M, ее длина примерно в 1.9 раз больше ширины. Отрезок rs-m расположен дистальнее 2r-rs, так что длины 4RS и 2r-rs одинаковы. Медиокубитальная ячейка средних размеров типа параллелограмма, так что противоположные стороны параллельны и почти равны по длине: 1M > m-cu в 1.1 раза; 1Cu и (RS+M)+2M одинаковой длины. Ячейка mcu удлиненная, ее дистальный конец не достигает уровня птеростигмы. Вторая кубитоанальная ячейка не сформирована. Ir = 0.7. Icu ~ 1.6, Icua ~ 1.3, I(mcu) ~ 0.6.
Размеры в мм. FWL 6.7–7.0. FWPt ~ 3.3, длина ячейки 3r = 1.9, длина ячейки rm = 0.96.
Сравнение. Размеры крыльев у трех видов этого рода из Большой Светловодной несколько меньше, чем у нового вида, а также этот отпечаток значительно отличается от всех известных Emplastus по форме медиокубитальной ячейки.
Замечания. Особенностью нового вида является форма медиокубитальной ячейки, что отражено в ее названии. Наибольшее сходство, в том числе по форме ячейки mcu, наблюдается с жилкованием крыла Britaneuretus anglicus (Cockerell, 1915) (Antropov et al., 2014). Похожее жилкование имеют некоторые Pachycondyla, но важные отличия, а именно: проксимальное положение m-cu (дистального края ячейки mcu) относительно птеростигмы и cu-a, расположенная на удалении от ячейки mcu, обуславливают принадлежность крыла к Dolichoderinae (либо к вымершим родам Aneuretinae, подобно Britaneuretus).
Материал. Голотип.
Emplastus similis Perfilieva, sp. nov.
Табл. X , фиг. 3
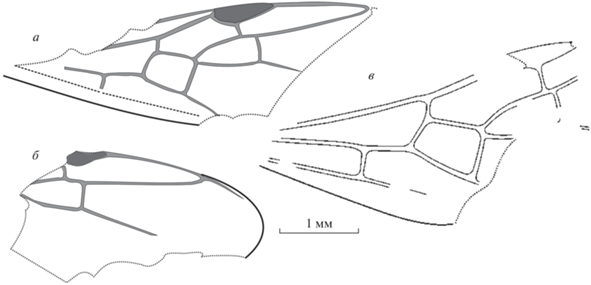
Рис. 3.
Виды Emplastus: а, б – E. similis sp. nov., передние крылья; Великая Кема: а – голотип ПИН, № 3136/10, б – экз. ПИН, № 3136/11 (зеркальное отражение); в – E. hypolithus (Cockerell, 1915), голотип USNM 61411; Бембридж. Масштабная линейка – 1 мм.
Название вида similis лат. – подобный, похожий.
Голотип – ПИН, № 3136/10, отпечаток переднего крыла; Приморский край, Тернейский р-н, пос. Великая Кема; нижний олигоцен, кизинская свита.
Описание (рис. 3, а, б). Птеростигма округлая. Ячейка 3r замкнута, ее вершина расположена близко к краю крыла, имеется выраженный участок R1 + RS. Радиомедиальная ячейка (rm) без стебелька, имеется небольшая зона контакта с ячейкой mcu (отрезок 2M очень короткий), ее длина в 2.5 раза больше ширины. Поперечная 2r‑rs слегка наклонена нижним концом к вершине крыла. Ячейка mcu почти прямоугольная, средних размеров, 1M равна m-cu и в 2 раза длиннее 1RS. Отрезок rs-m расположен значительно дистальнее 2r-rs, но проксимальнее уровня вершины птеростигмы, так что 4RS хорошо выражен. Поперечная cu-a расположена на удалении от ячейки mcu. Вторая кубитоанальная ячейка не сформирована. Ir = 0.63, Icu = 1.3, Icua ~ 1.4, I(mcu) ~ 0.8.
Размеры в мм. Голотип: FWL ~ 4.8, FWPt = 2.8, длина ячейки 3r = 1.7, длина ячейки rm = 0.8.
Сравнение. Крыло по размерам и деталям жилкования очень сходно с крылом голотипа Emplastus hypolithus USNM 61411, описанного из эоценового местонахождения Бембридж, Великобритания (рис. 3, в). Небольшие отличия наблюдаются по размерам крыла, в относительной длине участков 2M + Cu и 4RS. Учитывая некоторые отличия в жилковании и в силу временного интервала, разделяющего находки, возможно описание нового вида.
Замечания. Особенность описанного жилкования – в сочетании признаков, имеющихся в крыльях разных современных родов долиходерин, но не встречающихся совместно: относительно небольшой размер крыла, длина m-cu несколько больше 1M, имеется выраженный отрезок 4RS, поэтому отнесение крыла к ортотаксону пока не может быть нами обосновано.
Материал. Голотип (переднее крыло) и отпечаток вершинной части крыла ПИН, № 3136/11.
Род Technomyrmex Mayr, 1872
Типовой вид –Technomyrmex strenuous Mayr, 1872; по монотипии.
Диагноз (для крыльев). Передние крылья с замкнутыми дистальными ячейками: 3r, rm, mcu. Однако маленькие самцы некоторых видов могут иметь редукцию дистальных ячеек rm и mcu, в этом случае сохраняются хорошо выраженные изгибы RS + M (в месте отхождения редуцированной m-cu) и свободной ветви M (в месте, где гипотетическая rs-m переходит в 4M). Радиомедиальная ячейка может быть четырехугольной, так что отрезок 2M хорошо выражен, однако не менее чем вдвое короче RS + M, или треугольной с очень коротким стебельком (т.е. отрезок 2RS + M короткий). Отличительная особенность рода — относительно маленькая прямоугольной формы медиокубитальная ячейка. Вторая кубитоанальная ячейка не сформирована. Заднее крыло у мелких видов с сильно редуцированным жилкованием, без свободной M, часто все дистальные свободные ветви представлены спектральными жилками, либо редуцированы.
Видовой состав. Род насчитывает 94 современных вида (каталог AntWeb, август 2021), представители которого встречаются на всех континентах. Наибольшее разнообразие видов наблюдается в Старом Свете: Африке и на юге Азии, и лишь четыре вида встречаются в Новом Свете, два из которых, по-видимому, являются завезенными человеком. Населяют влажные леса теплого климата, гнезда в почве, либо на деревьях. К настоящему моменту были описаны четыре ископаемых вида: два из доминиканского янтаря (Brandão et al., 1999): T. caritatis Brandão et al., 1999 (рабочий) и T. hispaniolae (Wilson, 1985); T. deletus Emery, 1891 из сицилийского янтаря (рабочий) и T. septentrionalis Zhang, 1989 (самец?) из миоцена Китая (формация Шанвонг, Шаньдун).
Technomyrmex kemaensis Perfilieva, sp. nov.
Табл. X , фиг. 4, 5
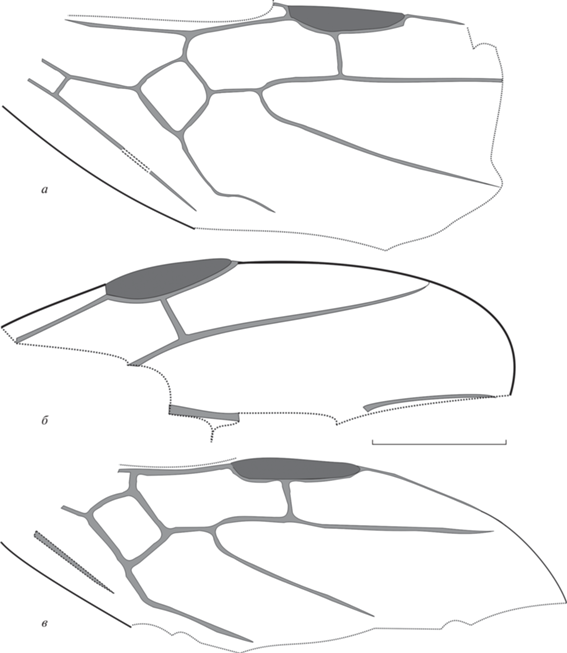
Рис. 4.
Technomyrmex kemaensis sp. nov., переднее крыло; Великая Кема: а – экз. ПИН, № 3136/6, б – голотип ПИН, № 3136/14. Масштабная линейка – 1 мм.
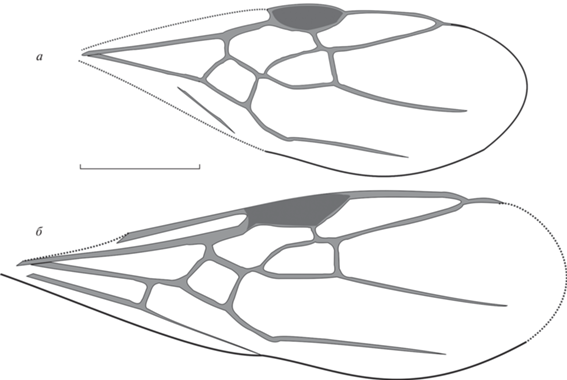
Рис. 5.
Ископаемые экземпляры, выведенные из Camponotus: а – Myrmecites ullrichi comb. nov. (Camponotus ullrichi, из: Bachmayer, 1960, рис. 8), б – Shanwangella palaeoptera (Camponotus palaeopterus, из: Zhang, 1989, рис. 307 ) , в – Camponotus (?) tokunagai (из: Naora, 1933, рис. 19 ).
Название вида от местонахождения отпечатка у р. Кемы.
Голотип – ПИН, № 3136/14, отпечаток переднего крыла; Приморский край, Тернейский р-н, пос. Великая Кема; нижний олигоцен, кизинская свита.
Описание (рис. 4). Крыло с относительно большой, округлой птеростигмой. Ячейка 3r замкнута, ее вершина расположена на краю крыла, участок R1 + RS расположен по краю крыла. Радиомедиальная ячейка на коротком стебельке, так что участок 2RS + M составляет не более одной четверти длины ячейки rm. Длина ячейки rm в 2.0–2.3 раза больше ширины. Отрезок 2-3RS изогнут. Поперечная 2r-rs слегка наклонена нижним концом к вершине крыла. Медиокубитальная ячейка относительно маленькая, форма практически квадратная; 1M в 2 раза длиннее 1RS. Отрезок rs-m расположен значительно дистальнее 2r-rs, но проксимальнее уровня вершины птеростигмы, так что 4RS хорошо выражен. Поперечная cu-a расположена недалеко от ячейки mcu (у отпечатка № 3136/6 cu-a не видна). Ir = 0.7, Icu = 1.7, Icua ~ 1.3, I(mcu) ~ 0.8.
Размеры в мм. Голотип: FWL ~ 4.8, FWPt = 2.1, длина ячейки 3r = 1.3, 1M = m-cu = = RS + M = 0.3, 2M + Cu = 0.3, 1M + Cu = 1, 1Cu = = 0.5. Экз. ПИН, № 3136/6: FWL ~ 3.7, FWPt = 1.6, длина ячейки 3r = 1, 1M = m-cu = RS + M = 0.3, 1‑2M + Cu = 0.9, 1Cu = 0.4.
Сравнение. Отпечатки крыльев имеются только у китайского T. septentrionalis, который значительно крупнее (FWL = 6.7 мм).
Материал. Голотип и прямой и обратный отпечатки переднего крыла ПИН, № 3136/6.
ПОДСЕМЕЙСТВО FORMICINAE LATREILLE, 1809
Род Camponotus Mayr, 1861
Типовой вид – Formica ligniperda Latreille, 1802, по последующему обозначению Bingham, 1903.
Диагноз (для крыльев). Передние крылья с замкнутой ячейкой 3r. Ячейка 1 + 2r среднего размера, не длиннее 3r. Ячейки радиомедиальная и медиокубитальная отсутствуют. 1RS и 1M составляют единую прямую линию и приблизительно равной длины. RS + M, ограничивающая ячейку 1 + 2r, плавно выгнута, практически не имеет S‑образного изгиба. Вторая кубитоанальная ячейка не сформирована.
Видовой состав. Род насчитывает более 1000 современных видов (1045 видов, каталог AntWeb, август 2021), встречающихся на всех континентах. Наибольшее разнообразие видов наблюдается в гумидных бореальных экосистемах. К настоящему моменту описан 31 ископаемый вид с эоцена по миоцен в местонахождениях Северной Америки, Европы и Китая: эоцен: C. cockerelli (Donisthorpe, 1920) (Великобритания), C. mengei Mayr, 1868 (балтийский янтарь), Северная Америка – C. fuscipennis Carpenter, 1930, C. microcephalus Carpenter, 1930, C. petrifactus Carpenter, 1930, C. vetus Scudder, 1877, и Китай – C. tokunagai Naora, 1933 (янтарь). Олигоцен: C. vehemens Förster, 1891 (Германия), Франция – C. theobaldi Özdikmen, 2010, C. compactus Förster, 1891, C. longiventris Théobald, 1937, C. penninervis Théobald, 1937. 10 миоценовых видов из Китая: C. ambon Zhang, 1989, C. ampullosus Zhang, 1989, C. curviansatus Zhang, 1989, C. gracilis Zhang, 1989, C. longus Zhang, 1989, C. microthoracus Zhang, 1989, C. palaeopterus (Zhang, 1989), C. pictus Zhang, Sun et Zhang, 1994, C. plenus Zhang, 1989 и C. shanwangensis Hong, 1984; четыре вида из местонахождений Германии и Швейцарии: C. obesus Piton, 1935, C. lignitus (Germar, 1837), C. heracleus (Heer, 1849), C. oeningensis (Heer, 1849), по одному из Франции, Хорватии, Чехии, Австрии и Северной Америки соответственно, – C. crozei Riou, 1999, C. induratus (Heer, 1849), C. novotnyi Samsinák, 1967, C. ullrichi Bachmayer, 1960, Paleosminthurus juliae Pierce et Gibron, 1962 (перенесен в Camponotus: Najt, 1987).
Замечания. Из рода Camponotus следует исключить C. ullrichi Bachmayer, 1960. Схематичный рисунок крыла и недостаточно хорошего качества фотоизображение в оригинальной публикации не позволяют определить однозначно таксономическую принадлежность отпечатка крыла, однако незамкнутая 3r и проксимальное положение ответвления свободной продольной ветви M относительно 2r-rs и птеростигмы, взаимное под углом расположение 1RS и 1M, отсутствие наклона 2r-rs и другие особенности указывают на ошибочность описания этого крыла как Camponotus (рис. 5, а) (Bachmayer, 1960). Наиболее вероятно его положение в подсемействе Myrmicinae, поэтому здесь переносим в формальный род неясного положения – Myrmecites ullrichi, comb. nov.
Изображение и рисунок миоценового C. palaeopterus (Zhang, 1989) вполне отчетливы и детальны для того, чтобы исключить принадлежность этого муравья к роду Camponotus: из-за наличия радиомедиальной ячейки очевидна его принадлежность к Dolichoderinae (рис. 5, б). Мы считаем, что первоначальное таксономическое положение этого экземпляра (Zhang, 1989) необходимо восстановить – Shanwangella palaeoptera Zhang, 1989.
Весьма сомнительна принадлежность C. tokunagai Naora, 1933: авторский рисунок крыла не соответствует не только жилкованию Camponotus, но не сопоставим с любым типом жилкования крыльев муравьев (рис. 5, в). Также есть сомнения в отнесении C. microcephalus Carpenter, 1930 к этому роду, поскольку на рисунке крыла 1RS находится под углом к 1M, однако эти утверждения требуют дополнительного исследования материала.
Camponotus parvus Perfilieva, sp. nov.
Табл. X , фиг. 6
Рис. 6.
Camponotus parvus sp. nov., голотип ПИН, № 3136/359, переднее крыло (зеркальное отражение); Великая Кема. Масштабная линейка – 1 мм.
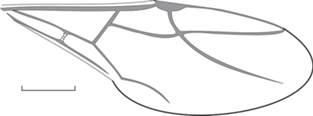
Название вида parvus лат. – маленький.
Голотип – ПИН, № 3136/359, прямой и обратный отпечатки переднего крыла самца (?); Приморский край, Тернейский р-н, пос. Великая Кема; нижний олигоцен, кизинская свита.
Описание (рис. 6). Птеростигма маленькая, округлая. Ячейка 3r приблизительно в 1.6 раза длиннее 1 + 2r. Отрезок 1RS приблизительно в 1.9 раза короче 1M. RS + M, 5RS плавно выгнуты. Ir = 0.63, Icua = 1.5
Размеры в мм. FWL ~ 6; FWPt = 2.9; длина ячейки 3r = 2.2.
Сравнение. Сопоставление признаков отпечатков крыльев Camponotus, описанных на данный момент, малопродуктивно. Во-первых, большинство из 28 ископаемых видов (т.е. без трех отпечатков, выведенных из Camponotus в настоящей работе) описаны по самкам. С учетом выраженного полового диморфизма по размерам половых особей в этом роде сложно сопоставить даже размеры самцов и самок из разных местонахождений. Поскольку длина крыла самца C. parvus sp. nov. составляет 6 мм, то можно считать, что размеры самок этого вида могут быть 8–16 мм. Во-вторых, стандарты описания и рисунков большинства видов, описанных в начале прошлого века, не позволяют сравнивать детали жилкования, которые являются важными для таксономических целей. Учитывая эти особенности, наше сравнение показывает, что среди известных ископаемых представителей Camponotus имеются три вида с относительно длинной ячейкой 3r (Ir около 0.6): C. curviansatus (самка, FWL = 10 мм), C. longus (самка, FWL = 11.8 мм) и C. gracilis (самец, FWL = 10.4 мм). Однако последний значительно крупнее C. parvus sp. nov. Исключить синонимию C. parvus sp. nov. с одной из двух самок указанных двух видов без дополнительного исследования образцов невозможно.
Замечания. В исследовании крыльев современных представителей Formicinae было показано, что самцы имеют относительно более широкие крылья, чем самки этого подсемейства, вне зависимости от абсолютных размеров особей (Перфильева, 2007, 2021). Исходя из этих данных, мы предполагаем, что отпечаток крыла C. parvus sp. nov. принадлежит самцу.
Материал. Голотип.
Род Lasius Fabricius, 1804
Типовой вид – Formica nigra (L., 1758), по последующему обозначению Bingham, 1903.
Диагноз (для крыльев). Передние крылья с двумя дистальными ячейками: 3r и mcu. Радиомедиальная ячейка отсутствует. Ячейка mcu у мелких самцов может отсутствовать за счет редукции поперечной m-cu. Ячейки 1 + 2r и 3r среднего размера, последняя обычно относительно длиннее первой. Ячейка mcu трапециевидная, обычно средних размеров, иногда относительно маленькая; 1M и m-cu приблизительно равной длины, 1Cu несколько длиннее отрезка 1RS + M. Отрезок 2RS + M, как правило, плавно выгнут, отсутствует S-образный изгиб. 2r-rs прямая и явно наклонена нижним концом к вершине крыла, 5RS и свободная M выходят из одной точки, при этом изгиб последней заметно больше, чем у 5RS.
Видовой состав. На настоящий момент насчитывается 127 современных видов Lasius, обитающих в голарктической области, и 22 ископаемых вида (каталог AntWeb, август 2021). Два вида из эоцена США: L. glom LaPolla et Greenwalt, 2015, L. peritulus (Cockerell, 1927). Четыре ископаемых вида из балтийского янтаря: L. punctulatus Mayr, 1868, L. pumilus Mayr, 1868, L. schiefferdeckeri Mayr, 1868, L. nemorivagus Wheeler, 1915 (Dlussky, 2011). В олигоценовых отложениях Франции найден L. epicentrus Théobald, 1937 (самец). Миоценовые представители рода описаны из разных местонахождений: Хорватия – L. occultatus (Heer, 1949), L. globularis (Heer, 1949), L. longaevus (Heer, 1949), L. ophthalmicus (Heer, 1949), L. anthracinus (Heer, 1867), L. longipennis (Heer, 1849) (Dlussky, Putyatina, 2014); Китай – L. inflatus (Zhang, 1989) L. truncatus Zhang, 1989 (самка), L. validus Zhang, 1989, L. mordicus Zhang, 1989 (Zhang, 1989); Россия – L. vetulus Dlussky, 1981, L. tertiarius Zalessky, 1949; Польша – L. oblongus Assmann, 1870; Франция – L. crispus Théobald, 1935, а также из плиоценовых отложений L. chambonensis Théobald, 1935.
Замечания. Один или оба вида из верхнеэоценовых отложений о-ва Уайт, описанные в формальном таксоне, Leucotaphus donisthorpei Dlussky et Perfilieva, 2014 и L. gurnetensis (Cockerell, 1915), также могут принадлежать Lasius (Antropov et al., 2014).
Lasius vulgaris Perfilieva, sp. nov.
Табл. X , фиг. 7, 8
Рис. 7.
Lasius vulgaris sp. nov., переднее крыло: а – голотип ПИН, № 3136/3, самка (?), б – экз. ПИН, № 3136/8, самец (?), в – экз. ПИН, № 3136/7, самец (?) (зеркальное отражение); Великая Кема. Масштабная линейка – 1 мм.
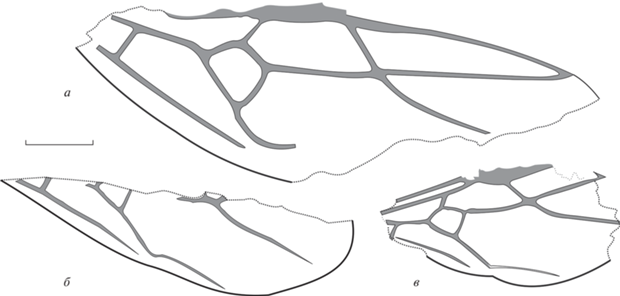
Рис. 8.
Paraphaenogaster ussuriensis sp. nov., переднее крыло: а – голотип ПИН, № 3136/1, б – экз. ПИН, № 3136/362 (зеркальное отражение), в – экз. ПИН, № 3136/4; Великая Кема. Масштабная линейка – 1 мм.
Название вида vulgaris лат. – типичный, обыкновенный.
Голотип – ПИН, № 3136/3, прямой и обратный отпечатки переднего крыла самки (?); Приморский край, Тернейский р-н, пос. Великая Кема; нижний олигоцен, кизинская свита.
Описание (рис. 7). Ячейки 1 + 2r и 3r среднего размера, последняя относительно длиннее первой. Ячейка mcu трапециевидная, средних размеров; 1M и m-cu приблизительно равной длины, 1Cu приблизительно в 1.9 раза длиннее отрезка 1RS + M; отрезки 2RS + M и 5RS слабо выгнуты – почти прямые линии, без изгибов. 2r‑rs прямая и явно наклонная. Ir = 0.6, I(mcu) = = 0.8, Icu ~ 2–2.2. Icua рассчитать не удается, т.к. основание крыла не сохранилось.
Размеры в мм. FWL (самка, голотип) ~9–9.5 (длина видимой части отпечатка от cu-a до вершины ячейки 3r около 6.6), FWL (самцы, экз. № 3136/7 и 8) ~5–5.4 (длина видимой части отпечатка от cu-a до вершины ячейки 3r около 3.2).
Сравнение. Размеры крыльев большинства описанных ископаемых видов Lasius и двух видов Leucotaphus отличаются от описанного вида. Сходные размеры крыльев у олигоценового L. epicentrus, однако рисунок крыла недостаточно детальный для анализа. Особый интерес представляет сравнение с другими миоценовыми видами подходящего размерного класса: L. anthracinus и L. occultatus имеют более высокую ячейку mcu, у L. ophthalmicus cu-a расположена значительно дальше от ячейки mcu, L. validus и L. chambonensis имеют относительно более короткую ячейку 3r. Китайский L. truncatus без крыльев, поэтому сравнение невозможно. Leucotaphus donisthorpei отличается относительно маленькой ячейкой mcu.
Замечания. Судя по пропорциям крыльев, более маленькие крылья принадлежат самцам, а крупное — самке. Остальные признаки жилкования очень сходны, поэтому мы считаем возможным объединение трех отпечатков в один вид. Абсолютные размеры крыльев вычислены по реконструкциям, выполненным на основании анализа формы крыльев современных Lasius.
Материал. Помимо голотипа, экз. ПИН, № 3136/7 – отпечаток (прямой и обратный) центральной части переднего крыла самца (?) и экз. ПИН, № 3136/8 – неполный отпечаток нижней части переднего крыла самца (?).
ПОДСЕМЕЙСТВО MYRMICINAE LEPELETIER DE SAINT-FARGEAU, 1835
Род (формальный) Paraphaenogaster Dlussky, 1981
Типовой вид – Paraphaenogaster microphthalma Dlussky, 1981, миоцен Северного Кавказа.
Диагноз (для крыльев). Передние крылья с редуцированной ячейкой rm. Третья радиальная ячейка обычно открыта: свободная ветвь RS длинная, не доходит до R1, но в редких случаях они соединяются, так что 3r оказывается замкнутой. Поперечная 2r-rs строго вертикальная по отношению к нижнему краю птеростигмы, либо слегка наклонена нижним концом к основанию крыла, относительно короткая. Радиомедиальная ячейка отсутствует за счет полной редукции поперечной rs-m. Отрезки 2RS + M и 3RS хорошо выражены и, таким образом, свободная ветвь M ответвляется значительно проксимальнее 2r-rs. Ячейка mcu трапециевидной формы, среднего размера. Вторая кубитоанальная ячейка не сформирована.
Видовой состав. Описано 11 ископаемых видов с верхнего эоцена до среднего миоцена. Из верхнего эоцена Великобритании известен P. hooleyana Dlussky et Perfilieva, 2014 (о. Уайт, Бембридж). Шесть видов из верхнего олигоцена Германии (Энспель): P. bizeri Jessen, 2020, P. freihauti Jessen, 2020, P. loosi Jessen, 2020, P. schindleri Jessen, 2020, P. wettlauferi Jessen, 2020 и P. wuttkei Jessen, 2020. Из миоцена Китая: P. dumetorum (Lin, 1982). Из нижнего миоцена Хорватии (Радобой): P. tertiaria (Heer, 1849) и P. jurinei (Heer, 1849). Из среднего миоцена Ставропольского края (Россия, Вишневая балка): P. microphthalma Dlussky, 1981.
Замечания. Целесообразно рассматривать представителей этого рода, включая в анализ три миоценовых вида из Китая, которые отнесены к Aphaenogaster: A. lapidescens Zhang, 1989, A. paludosa Zhang, 1989 и A. shanwangensis (Hong, 1984). P. hooleyana Dlussky et Perfilieva, 2014 имеет наиболее уклоняющееся жилкование крыла среди всех остальных видов, а именно: относительно короткий участок 3RS. По нашему мнению, формальный характер рода требует пересмотра на основании довольно репрезентативного к настоящему моменту палеонтологического материала, включающего самцов, крылатых самок и рабочих особей и, соответственно, следует пересмотреть его состав. Эта работа планируется на ближайшее будущее.
Paraphaenogaster ussuriensis Perfilieva, sp. nov.
Табл. X , фиг. 9
Рис. 9.
Отпечатки, отнесенные к Formicidae inc. sed.: а – экз. ПИН, № 3136/2, заднее крыло; б – экз. ПИН, № 3136/5, заднее крыло; в – экз. ПИН, № 3136/12 (зеркальное отражение); г – экз. ПИН, № 3136/13; Великая Кема. Масштабная линейка – 1 мм.
Название вида – от исторического названия географического региона – Уссурийский край.
Голотип – ПИН, № 3136/1, прямой и обратный отпечатки переднего крыла; Приморский край, Тернейский р-н, пос. Великая Кема; нижний олигоцен, кизинская свита.
Описание (рис. 8). Птеростигма продолговатая. Вершина ячейки 3r плохо видна, поэтому сложно определить, замкнута она или нет. Медиокубитальная ячейка средних размеров, трапециевидная, длины 1M и m-cu равны, отрезок 1Cu в 1.1–1.4 раза длиннее 1RS + M. Поперечная жилка 2r-rs вертикальная. Свободная ветвь M отходит от 2RS + M значительно проксимальнее 2r-rs, так что длина 2RS + M приблизительно в 1.7 раза короче 3RS. Icu = 2.1, I(mcu) = 1.
Размеры в мм. Голотип: FWL ~ 5, 1R1 ~ 0.7, 1Cu ~ 0.5.
Сравнение. Крылья у нового вида по размерам меньше, чем у всех описанных на настоящий момент (за исключением P. hooleyana, имеющего уникальное жилкование, отличающее его от всех иных Paraphaenogaster, см. выше). Также можно отметить специфическое для нового вида соотношение разных отрезков жилок (форма mcu, соотношение 2RS + M/3RS и др.).
Замечания. Три неполных отпечатка крыла, отнесенные нами к этому виду, имеют почти одинаковые размеры и очень сходны по деталям жилкования.
Материал. Помимо голотипа, экз. ПИН, № 3136/4, прямой и обратный отпечатки переднего крыла без проксимальной части, и экз. ПИН, № 3136/362, дистальная часть крыла.
* * *
Прямые и обратные почти полные отпечатки двух задних крыльев разных видов муравьев из Великой Кемы, экз. ПИН, №№ 3136/2 и 5 (табл. X , фиг. 10, 11 ), определены как Formicidae inc. sed.; длина обоих крыльев около 5.2 мм (рис. 9, а, б). Оба экземпляра с редуцированной свободной ветвью M. В крыле экз. № 3136/2 rs-m относительно короткая, ее длина приблизительно равна длине участка 2M + Cu, жилка 1RS не выражена, cu-a относительно длинная (около 0.4 мм) (см. рис. 9, а) (табл. X , фиг. 10 ). В крыле экз. № 3136/5 rs-m относительно длинная, ее длина приблизительно в 1.5 раза больше участка 2M + Cu, жилка 1RS хорошо выражена, cu-a относительно короткая (около 0.2 мм) (рис. 9, б; табл. X , фиг. 11 ).
Отпечаток основания переднего крыла экз. ПИН, № 3136/12: по положению cu-a относительно основания крыла и ячейки mcu, крыло, очевидно, имеет медиокубитальную ячейку и может принадлежать представителям Formicinae, Dolichoderinae или Myrmicinae (рис. 9, в). FWL ~ 4.8 мм, длина видимой части отпечатка крыла около 2 мм.
Отпечаток вершинной части крыла экз. ПИН, № 3136/13 (вершина 3r плохо просматривается): скорее всего, крыло не принадлежит представителю Formicinae (рис. 9, г). FWL ~ 4 мм, длина видимой части отпечатка крыла около 1.7 мм.
ОБСУЖДЕНИЕ
Учитывая небольшое число отпечатков муравьев, имеющихся из местонахождений Амгу и Великая Кема, можно сделать лишь предварительный анализ особенностей этих находок. Одновременное присутствие элементов мирмекокомплексов умеренного климата и тропических теплолюбивых видов показывает, что ситуация, описываемая “дилеммой Вилера” и обнаруженная в богатом местонахождении этого региона в позднем эоцене (Большая Светловодная), наблюдается здесь и в раннем олигоцене, хотя и с другим таксономическим составом. Так, в позднеэоценовой Светловодной это были Formica и, по-видимому, Lasius против теплолюбивых Gesomyrmex и Myrmeciinae, а в местонахождении Великая Кема, соответственно, Lasius и Technomyrmex. Представители Technomyrmex (до нашей находки) появляются в палеонтологической летописи в позднем олигоцене (cицилийский янтарь), и очевидно, что этот теплолюбивый представитель мирмекофауны, в настоящее время распространенный в тропических и субтропических областях Старого Света (в Новом Свете завезенные человеком виды), раньше имел гораздо более широкий ареал (сицилийский янтарь, Великая Кема, доминиканский янтарь, Шанвонг). Наши данные соответствуют представленному флористическому заключению о более теплом климате в этой местности в раннем олигоцене, чем в настоящее время.
Можно отметить высокое видовое разнообразие ориктоценоза: 14 отпечатков передних крыльев соотносятся с семью разными видами трех подсемейств. Интересно значительное число особей (семь экз.) и преобладание по таксономическому разнообразию (пять видов) муравьев, связанных с древесным образом жизни – все виды Dolichoderinae и Camponotus, на фоне меньшей доли мирмицин, представленных одним видом Paraphaenogaester, и при полном отсутствии геобионтов и стратобионтов понероморф. Такое соотношение жизненных форм характерно для захоронения в смолах, а в осадочных отложениях встречается только в североамериканских местонахождениях Грин-Ривер (ранний эоцен) и Флориссант (терминальный эоцен). Высокая доля дендробионтов подтверждает представление палеоботаников о выраженном лесном характере флоры Амгу и Великой Кемы без примеси равнинных компонентов, однако отсутствие понероморф можно объяснить пока только небольшим числом находок муравьев из этих местонахождений.
Подсемейство Myrmicinae представлено в описанных здесь местонахождениях единственным видом формального рода Paraphaenogaster. Виды этого рода встречаются в ориктоценозах Европы с верхнего эоцена до миоцена, а также есть в миоценовом метонахождении Шанвонг (Китай). Причем их представленность часто очень высока: в позднеэоценовом Бембридже мирмицин очень мало (менее 1% отпечатков), но три четверти всех отпечатков мирмицин принадлежат этому роду; шесть видов Paraphaenogaster описано из позднеолигоценового местонахождения Энспель (Германия); 80% всех мирмицин, которые составляют около трети всех отпечатков, наблюдаются в позднеолигоценовом Ротте (Германия). Все Myrmicinae (составляющие 40% отпечатков) в миоценовой Вишневой балке (Ставрополь) представлены отпечатками нескольких видов этого рода, а в среднемиоценовом Шанвонг четыре из пяти отпечатков мирмицин принадлежат одному виду Paraphaenogaster. При этом в богатом местонахождении позднего эоцена Дальнего Востока (Большая Светловодная) среди отпечатков мирмицин, представляющих 21% всех отпечатков, нет Paraphaenogaster. По-видимому, присутствие этого рода следует трактовать как более позднее «приобретение» местной мирмекофауны. Таким образом, по имеющимся данным следует сделать предварительный вывод о большем сходстве мирмекофаун местонахождений Великая Кема и китайского Шанвонг, чем с географически более близким, но более древним позднеэоценовым местонахождением Большая Светловодная.
* * *
Автор признательна сотрудникам лаб. артропод ПИН РАН и лично А.П. Расницыну за предоставленный материал и помощь при работе с ним; С.Н. Лысенкову и В.Г. Гринькову (МГУ) за помощь при работе с текстом. Работа выполнена в рамках научного проекта государственного задания МГУ 04-1-21 №121031600198-2.
Список литературы
Лопатина Д.А. Растительность и климат эоцена–миоцена восточного Сихотэ-Алиня по данным палеоботанического анализа // Тихоокеанск. геол. 2004. Т. 23. № 3. С. 98–112.
Павлюткин Б.И., Чекрыжов И.Ю., Петренко Т.И. Геология и флоры нижнего олигоцена Приморья. Владивосток: Дальнаука, 2014. 236 с.
Перфильева К.С. Аномалии крыльев у половых особей муравьев (Hymenoptera, Formicidae) с разной стратегией брачного поведения // Зоол. журн. 2000. Т. 79. № 11. С. 1311–1312.
Перфильева К.С. Характеристика изменчивости количественных признаков крыльев на примере некоторых видов муравьев (Hymenoptera, Formicidae) // Успехи совр. биол. 2007. Т. 127. № 2. С. 147–156.
Перфильева К.С. Тенденции в эволюции жилкования крыльев муравьев (Hymenoptera, Formicidae) // Зоол. журн. 2010. Т. 89. № 8. С. 965–977.
Перфильева К.С. Новые данные по морфологии крыльев меловых муравьев Sphecomyrminae (Hymenoptera: Formicidae) // Палеонтол. журн. 2011. № 3. С. 38–45.
Перфильева К.С. Распространение и дифференциация по отпечаткам крыльев ископаемых видов Oecophylla (Hymenoptera: Formicidae) // Палеонтол. журн. 2021. № 1. С. 80–93.
Расницын А.П. Происхождение и эволюция перепончатокрылых насекомых. М.: Наука, 1980. 192 с. (Тр. Палеонтол. ин-та АН СССР. Т. 174).
Antropov A.V., Belokobylskij S.A., Compton S.G. et al. The wasps, bees and ants (Insecta: Vespida = Hymenoptera) from the Insect Limestone (Late Eocene) of the Isle of Wight, UK // Earth Envir. Sci. Trans. Roy. Soc. Edinb. 2014. V. 104. № 3–4. P. 335–446.
AntWeb. Version 8.64.2. California Academy of Science, online at https://www.antweb.org. Accessed 13 August 2021.
Bachmayer F. Insektenreste aus den Congerienschichten (Pannon) von Brunn-Vösendorf (südl. von Wien) Niederösterreich // Sitzungsber. Österr. Akad. Wiss. Math.-Naturwiss. Kl., Abt. I. 1960. Bd 169. S. 11–16.
Bondarenko O.V., Blokhina N.I., Utescher T. Major plant biome changes in the Primorye Region (Far East of Russia) during the Paleogene // Bot. Pacif. 2019. V. 8. № 1. P. 3–18.
Brandão C.R.F., Baroni Urbani C., Wagensberg J. et al. New Technomyrmex in Dominican amber (Hymenoptera: Formicidae), with a reappraisal of Dolichoderinae phylogeny // Entomol. Scandinavica. 1999. V. 29. P. 411–428.
Brown W.L., Nutting W. Wing venation and the phylogeny of the Formicidae // Trans. Amer. Entomol. Soc. 1949 (1950). V. 75. № 3–4. P. 113–132.
Dlussky G.M. The ants of the genus Lasius Fabricius (Hymenoptera, Formicidae) from the Late Eocene European ambers // Vestn. Zool. 2011. V. 45. № 3. P. 209–222.
Dlussky G.M., Putyatina T.S. Early Miocene ants (Hymenoptera, Formicidae) from Radoboj, Croatia // N. Jb. Geol. Paläontol. Abh. 2014. V. 272. № 3. P. 237–285.
Dlussky G.M., Rasnitsyn A.P., Perfilieva K.S. The ants (Hymenoptera: Formicidae) of Bol’shaya Svetlovodnaya (Late Eocene of Sikhote-Alin, Russian Far East) // Caucasian Entomol. Bull. 2015. V. 11. № 1. P. 131–152.
Najt J. Le Collembole fossile Paleosminthurus juliae est un Hyménoptère // Rev. Fr. Entomol. (N. Sér.). 1987. V. 9. P. 152–154.
Zhang J. Fossil insects from Shanwang, Shandong, China. Jinan: Shandong Sci. and Technol. Publ. House, 1989. 459 p. [In Chinese].
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал