

Палеонтологический журнал, 2022, № 4, стр. 73-76
Новый вид апионин рода Baltoconapium Legalov, 2020 (Coleoptera: Brentidae) из балтийского янтаря
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
b Алтайский государственный университет
656049 Барнаул, Россия
c Томский государственный университет
634050 Томск, Россия
* E-mail: fossilweevils@gmail.com
Поступила в редакцию 09.11.2021
После доработки 10.12.2021
Принята к публикации 10.12.2021
- EDN: SNKULD
- DOI: 10.31857/S0031031X22040031
Аннотация
Описан новый вид жука-апиониды Baltoconapium vilhelmseni sp. nov. из балтийского янтаря. Новый вид отличается от единственного входящего в этот род B. anderseni (Voss, 1972) более узким телом, менее выпуклыми надкрыльями с зеленоватым блеском, едва выпуклыми промежутками надкрылий, более длинной головотрубкой и более мелкими размерами.
ВВЕДЕНИЕ
Триба Piezotrachelini – всесветно распространенная группа подсемейства Apioninae (семейство Brentidae), включающая в себя 24 рода, два из которых – полностью вымершие (Alonso-Zarazaga, Lyal, 1999; Legalov, 2015, 2020b). В ископаемом состоянии известно три представителя трибы: Electrapion kuntzeni (Wagner, 1924), Conapium alleni Legalov, 2012 и Baltoconapium anderseni (Voss, 1972). Все они найдены в позднеэоценовом балтийском янтаре (Legalov, 2020a). Первые два описаны из янтаря, добытого в Калининградской обл. (Россия), а последний – из янтаря, собранного в Дании (Wagner, 1924; Voss, 1972; Legalov, 2012, 2020a). В виде отпечатков, а также в миоценовых янтарях Piezotrachelini пока не обнаружены (Legalov, 2015; Poinar, Legalov, 2015).
В настоящей работе описывается четвертый ископаемый представитель трибы Piezotrachelini – новый вид рода Baltoconapium из эоценового балтийского янтаря.
МАТЕРИАЛ И МЕТОДЫ
Исследованный образец происходит из балтийского янтаря, добытого в окрестностях пос. Янтарный, расположенного на побережье Балтийского моря в Калининградской обл. (Россия). Продуцентом балтийского янтаря, вероятно, был один из видов рода Sciadopitys семейства Sciadopityaceae (Sadowski et al., 2016). Отложения прусской свиты, в которых найден образец, датируются диапазоном от бартонского яруса, средний эоцен (Bukejs et al., 2019), до приабонского яруса, поздний эоцен (Perkovsky et al., 2007).
Типовой материал хранится в коллекции Ин-та систематики и экологии животных (ИСЭЖ) СО РАН, Новосибирск.
Описание, промеры и фотографии были сделаны при помощи бинокулярного микроскопа Zeiss Stemi 2000-C. Для обработки иллюстраций использовался Adobe Photoshop версии 21.1.3.
* * *
Автор благодарит Л. Вильгельмсена (L.B. Vilhelmsen) (Копенгаген) за возможность изучения голотипа Apion anderseni Voss, 1972 из коллекций Зоологического музея Копенгагенского ун-та (Zoological Museum, University of Copenhagen).
ТАКСОНОМИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО BRENTIDAE BILLBERG, 1820
ПОДСЕМЕЙСТВО APIONINAE SCHOENHERR, 1823
Триба Piezotrachelini Voss, 1959
Род Baltoconapium Legalov, 2020
Baltoconapium vilhelmseni Legalov, sp. nov.
Табл. XI , фиг. 1, 2 (см. вклейку)
Рис. 1.
Baltoconapium vilhelmseni sp. nov., голотип ИСЭЖ СО РАН, BA2021/6, общий вид: а – латерально, слева; б – дорсально; балтийский янтарь, поздний эоцен. Длина масштабного отрезка 0.5 мм.
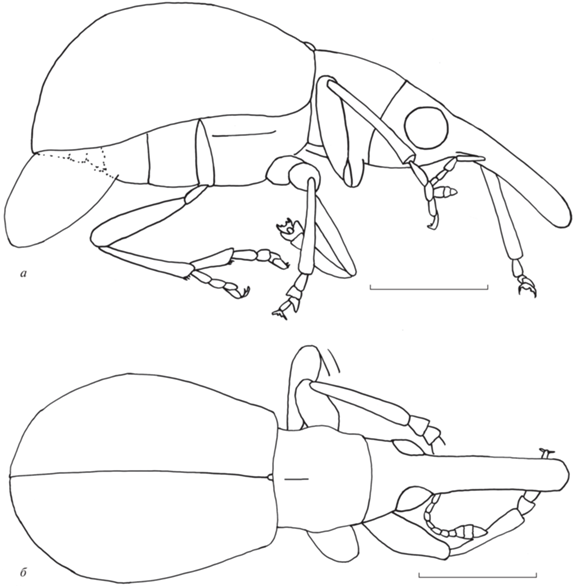
Название вида – в честь Л. Вильгельмсена (L.B. Vilhelmsen), Копенгаген.
Голотип – ИСЭЖ СО РАН, BA2021/6; балтийский янтарь, поздний эоцен.
Описание (рис. 1). Тело темно-коричневое, почти голое. Усики и ноги красно-коричневые. Головотрубка длинная, почти цилиндрическая, слабо изогнутая, немного расширяющаяся на вершине, мелко и густо пунктированная. Длина головотрубки в 1.4 раза больше длины переднеспинки, в 3.9 раза больше ширины на вершине, в 4.6 раза больше ширины на середине и в 4.4 раза больше ширины на основании. Усиковые бороздки явственные, глубокие, направлены под глаза, но не достигают их. Глаза округленные, явственно выступают из контура головы, грубо фасеточные. Лоб довольно широкий, слабовыпуклый, мелко и редко пунктированный. Ширина лба составляет 0.8 ширины головотрубки в основании. Длина висков составляет 0.5 длины глаза. Темя уплощенное, почти гладкое. Усики не коленчатые, прикреплены в первой трети головотрубки. Стволик короткий, не достигает глаза. Жгутик 7‑члениковый. Членики усика с первого по восьмой конические. Длина первого членика в 3.9 раза больше его ширины на вершине. Длина второго членика в 2.0 раза больше его ширины на вершине и равна половине длины стволика. Ширина второго членика составляет около 0.9 ширины стволика. Длина третьего членика в 2.1 раза больше его ширины на вершине и составляет 0.7 длины второго членика. Ширина третьего членика составляет 0.8 ширины второго членика. Длина четвертого членика в 1.9 раза больше его ширины на вершине и составляет около 0.9 длины третьего членика. Ширина четвертого членика немного меньше ширины третьего членика. Четвертый–шестой членики равной длины. Длина пятого членика в 1.8 раза больше его ширины на вершине. Ширина пятого членика немного больше ширины четвертого членика. Длина шестого членика в 1.5 раза больше его ширины на вершине. Ширина шестого членика в 1.2 раза больше ширины пятого членика. Длина седьмого членика в 1.2 раза больше его ширины на вершине и немного больше длины шестого членика. Ширина седьмого членика в 1.1 раза больше ширины шестого членика. Длина восьмого членика составляет 0.8 его ширины на вершине и 0.9 длины седьмого членика. Ширина восьмого членика в 1.2 раза больше ширины седьмого членика. Булава крупная, компактная, с явственными швами между члениками. Длина девятого членика составляет 0.6 его ширины на вершине и в 1.3 раза превышает длину восьмого членика. Ширина девятого членика в 1.7 раза больше ширины восьмого членика. Длина десятого членика составляет 0.7 его ширины на вершине и в 1.2 раза больше длины девятого членика. Ширина десятого членика в 1.1 раза больше ширины девятого членика. Длина 11-го членика в 1.3 раза больше его ширины на вершине и в 1.5 раза больше длины десятого членика. Ширина 11-го членика составляет 0.9 ширины десятого членика. Переднеспинка колоколовидная, густо и крупно пунктированная, с небольшой продольной серединной бороздкой перед щитком. Длина переднеспинки равна ширине на вершине, составляет 0.8 ширины на середине и слегка у́же ширины на основании. Промежутки между точками мелко шагренированные, равные или у́же диаметра точек. Диск переднеспинки слабо уплощенный, с очень тонкими и редкими волосками, направленными косо к середине. Бока слабо округленные. Ширина основания переднеспинки составляет 0.8 ширины основания надкрылий. Щиток явственный, почти прямоугольный, едва выступает над надкрыльями. Надкрылья явственно выпуклые, почти грушевидные, с зеленоватым блеском. Плечи выпуклые. Длина надкрылий почти в 2.8 раза больше длины переднеспинки, вдвое больше ширины на основании, в 1.2 раза больше ширины на середине и в 2.2 раза больше ширины в вершинной четверти. Бороздки надкрылий явственные, глубокие и широкие. Десятая бороздка отсутствует. Промежутки широкие, едва выпуклые, редко пунктированные. Ширина промежутков в 2.0–3.5 раза больше ширины бороздок. Седьмой промежуток надкрылий без длинной хеты. Переднегрудь без заглазничных лопастей, пунктированная. Пре- и посткоксальные части переднегруди очень короткие. Тазиковые впадины переднегруди соединенные, расположены на ее середине. Длина посткоксальной части больше длины прекоксальной части и составляет 0.5 длины переднего тазика. Тазиковые впадины среднегруди разделенные. Заднегрудь уплощенная, пунктированная, ее длина в 2.2 раза больше длины заднего тазика. Эпистерны заднегруди узкие. Брюшко выпуклое, пунктированное. Первый и второй вентриты удлиненные и сросшиеся. Длина первого вентрита в 1.8 раза больше длины заднего тазика. Третий и четвертый вентриты короткие и свободные, расположенные в другой плоскости по отношению к предыдущим вентритам. Длина третьего вентрита составляет 0.4 длины второго вентрита. Длина четвертого вентрита равна длине третьего вентрита. Длина пятого вентрита в 1.8 раза больше длины четвертого вентрита. Ноги длинные. Передние тазики конические. Вертлуги крупные, удлиненные. Бедра булавовидные, не соединяются с тазиками, без зубцов. Длина переднего и заднего бедер в 2.9 раза больше ширины на середине. Голени узкие и длинные, без мукро. Длина передней голени в 7.3 раза больше ширины на вершине. Длина задней голени в 4.7 раза больше ширины на вершине. Лапки длинные. Первый и второй членики лапок широко-трапециевидные. Третий членик лапок двухлопастный. Коготки широко расставленные, с зубцами. Длина первого членика передних лапок в 1.2 раза больше ширины на вершине. Длина второго членика передних лапок составляет 0.8 ширины на вершине и 0.9 длины первого членика. Ширина второго членика передних лапок в 1.3 раза больше ширины первого членика. Длина третьего членика передних лапок составляет 0.7 ширины на вершине и в 1.2 раза превышает длину второго членика. Ширина третьего членика передних лапок в 1.3 раза больше ширины второго членика. Длина первого членика средних лапок составляет 0.8 ширины на вершине. Длина второго членика средних лапок составляет 0.6 ширины на вершине и немного меньше длины первого членика. Ширина второго членика средних лапок в 1.2 раза больше ширины первого членика. Длина третьего членика средних лапок составляет 0.6 ширины на вершине и немного больше длины второго членика. Ширина третьего членика средних лапок в 1.1 раза больше ширины второго членика. Длина пятого членика средних лапок в 2.6 раза больше ширины на вершине и в 1.3 раза больше длины третьего членика. Ширина пятого членика средних лапок составляет 0.3 ширины третьего членика. Крылья частично видны из-под надкрылий.
Размеры в мм. Длина тела без головотрубки – 1.7; длина головотрубки – 0.55.
Сравнение. Новый вид отличается от B. anderseni (Voss, 1972) более узким телом, менее выпуклыми надкрыльями с зеленоватым блеском, едва выпуклыми промежутками надкрылий, более длинной головотрубкой и более мелкими размерами.
Замечания. Новый вид относится к семейству Brentidae, поскольку характеризуется удлиненными и сросшимися первым и вторым вентритами, а также короткими и свободными третьим и четвертым вентритами, расположенными в другой плоскости по отношению к предыдущим вентритам. Принадлежность нового вида к подсемейству Apioninae подтверждается удлиненными вертлугами, разделяющими бедро и тазик, не коленчатыми усиками, соединенными переднетазиковыми выемками, глубокими усиковыми бороздками и явственным щитком. Почти голое тело, волоски на переднеспинке, направленные косо к середине, отсутствие десятой бороздки надкрылий, явственные швы между члениками булавы и седьмой промежуток надкрылий без длинной хеты позволяют поместить новый вид в трибу Piezotrachelini.
Новый вид отнесен к роду Baltoconapium, поскольку обладает широкими первым и вторым члениками лапок, а также колоколовидной переднеспинкой.
Материал. Голотип.
* * *
Виды трибы Piezotrachelini в своем развитии связаны с растениями семейства бобовых (Alonso-Zarazaga, 1990). Из балтийского янтаря описаны Dalbergia sommerfeldii Casp. и Leguminosites myrtifolius Conw. (Czeczott, 1961). Возможно, Baltoconapium vilhelmseni sp. nov. развивался на одном из этих видов.
Список литературы
Alonso-Zarazaga M.A. Revision of the supraspecific taxa in the Palaearctic Apionidae Schoenherr, 1823 (Coleoptera, Curculionoidea). 2. Subfamily Apioninae Schoenherr, 1823: Introduction, keys and descriptions // Graellsia. 1990. V. 46. P. 19–156.
Alonso-Zarazaga M.A., Lyal C.H.C. A world catalogue of families and genera Curculionoidea (Insecta: Coleoptera) (excepting Scolytidae and Platypodidae). Barcelona: Entomopraxis, 1999. 315 p.
Bukejs A., Alekseev V.I., Pollock D.A. Waidelotinae, a new subfamily of Pyrochroidae (Coleoptera: Tenebrionoidea) from Baltic amber of the Sambian peninsula and the interpretation of Sambian amber stratigraphy, age and location // Zootaxa. 2019. V. 4664. № 2. P. 261–273.
Czeczott H. The flora of the Baltic amber and its age // Prace Muz. Ziemi. 1961. V. 4. P. 119–145.
Legalov A.A. New and little known Apioninae (Coleoptera, Brentidae) in Eocene Baltic amber // Euroasian Entomol. J. 2012. V. 11. № 3. P. 219–222.
Legalov A.A. Fossil Mesozoic and Cenozoic weevils (Coleoptera, Obrienioidea, Curculionoidea) // Paleontol. J. 2015. V. 49. № 13. P. 1442–1513.
Legalov A.A. A review of the Curculionoidea (Coleoptera) from European Eocene ambers // Geosciences. 2020a. V. 10. № 1(16). P. 1–74.
Legalov A.A. Fossil history of Curculionoidea (Coleoptera) from the Paleogene // Geosciences. 2020b. V. 10(9). № 358. P. 1–50.
Perkovsky E.E., Rasnitsyn A.P., Vlaskin A.P., Taraschuk M.V. A comparative analysis of the Baltic and Rovno amber arthropod faunas: representative samples // Afr. Invertebr. 2007. V. 48. P. 229–245.
Poinar G.Jr., Legalov A.A. New Apioninae (Coleoptera: Brentidae) in Dominican amber // Hist. Biol. 2015. V. 27. № 2. P. 134–157.
Sadowski E.-M., Schmidt A.R., Kunzmann L. et al. Sciadopitys cladodes from Eocene Baltic amber // Bot. J. Linn. Soc. 2016. V. 180. № 2. P. 258–268.
Voss E. Einige Rüsselkäfer der Tertiärzeit aus baltischen Bernstein (Coleoptera, Curculionidea) // Steenstupia. 1972. V. 2. P. 167–181.
Wagner H. Ein neues Apion aus dem baltischen Bernstein (Coleoptera, Curculionidae) // Dtsch. Entomol. Z. 1924. Jg. 1924. P. 134–136.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал