

Палеонтологический журнал, 2022, № 3, стр. 35-46
Эволюция жуков подотряда Polyphaga (Insecta: Coleoptera) в поздней перми и раннем–среднем триасе
Е. В. Ян a, *, О. Д. Стрельникова a, **
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: yan-e@mail.ru
** E-mail: ol.strelnikova@mail.ru
Поступила в редакцию 07.05.2021
После доработки 27.08.2021
Принята к публикации 25.10.2021
- EDN: KUVOEA
- DOI: 10.31857/S0031031X22030175
Аннотация
Даны краткая история изучения ранних этапов эволюции Polyphaga, а также на основе опубликованных данных разобраны причины их эволюционного успеха, позволившие Polyphaga занять доминирующее положение среди подотрядов Coleoptera с раннего мела и по настоящее время. Проанализирована морфология предков высших подотрядов жуков (семейства Ademosynidae и родов Archosyne и Ponomarenkium) из средней и поздней перми и триасовых предков (семейства Peltosynidae), с выделением ключевых особенностей их строения. Приведена оценка молекулярно-генетических и палеонтологических прогностических идей, предсказывавших облик и предполагаемый образ жизни древнейших Coleoptera. Показано, что базальные Polyphaga дают широкий спектр видов наравне с другими группами ранга инфраотряда среди насекомых с полным превращением.
ВВЕДЕНИЕ
Жуки подотряда Polyphaga являются одним из наиболее ярких примеров огромного биоразнообразия среди живых организмов. Представители Polyphaga начали доминировать в сообществах Coleoptera с мела (Пономаренко, 1983; Ponomarenko, 2003), и по числу видов и численности остаются абсолютно доминирующей группой среди современных жуков. Почти все современные жуки, с которыми человек сталкивается в природе, принадлежат к этому подотряду (за исключением жужелиц Carabidae, водных жуков плавунцов и вертячек Dytiscidae, Gyrinidae). Polyphaga объединяет более 380 тыс. видов (это число предполагается гораздо выше за счет неописанных пока видов; см., напр., Stork et al., 2015), что составляет 90% всех жуков, почти 40% насекомых и около 25% видов живых организмов на Земле (Oberprieler et al., 2007; Ślipiński et al., 2011; Costello et al., 2012).
Подобный эволюционный успех Polyphaga объясняют несколькими причинами. Первой из них называют тесную связь представителей подотряда с широко распространенными и разнообразными кормовыми субстратами и местообитаниями, в первую очередь – с покрытосеменными растениями (Angiospermae), древесиной (McKenna et al., 2015, 2019), растительной подстилкой (McKenna, 2016), а также с различными грибами (Robertson et al., 2015). Ключом к усвоению такой малопитательной и сложной для переваривания пищи [примерно половина всех растительноядных насекомых – жуки (Farrell, 1998)], как растительные ткани, стали геномные инновации, передавшиеся путем горизонтального переноса генов (horizontal gene transfer, HGT) от бактерий и грибов и позволившие жукам (в первую очередь это касается Buprestoidea и Phytophaga) вырабатывать энзимы для самостоятельного переваривания растительных клеточных стенок (т.н. plant cell wall-degrading enzymes, PCWDEs), без участия симбиотических бактерий (Calderón-Cortés et al., 2010; Kirsch et al., 2014).
Среди других причин необычного многообразия называют относительно раннее время появления жуков в геологической летописи (ассельский век, около 298 млн л.н.), давшее им достаточно времени на диверсификацию, а также высокую скорость видообразования при общих низких темпах вымирания (имеются в виду таксоны уровня семейства и выше), связываемых с механизмами экологического оппортунизма (Crowson, 1981; Farrell, 1998; Grimaldi, Engel, 2005; Hunt et al., 2007; McKenna, Farrell, 2009; McKenna et al., 2015). Экологический оппортунизм в этом ключе понимается как способность входить в новые адаптивные зоны (приближаясь по этому признаку к Hymenoptera и Diptera) путем развития ключевых эволюционных инноваций (Crowson, 1981; Lawrence, Newton, 1982; Beutel, 1997; Grimaldi, Engel, 2005). Таким образом, считается, что жуки используют уже существующие преадаптации, такие как, например, стратегии питания для освоения новых ниш (Leschen, 1993; McKenna, Farrell, 2006; Leschen, Buckley, 2007), примером может служить диверсификация и активное расселение покрытосеменных растений, образовавших растительные сообщества нового типа в мелу, и последовавшая за этим вспышка разнообразия у связанных с ними групп жуков, в первую очередь из инфраотряда Cucujiformia (см. Farrell, 1998; Bernhardt, 2000; McKenna, Farrell, 2009).
Граница перми и триаса 251 млн л.н. стала одним из таких ключевых этапов в эволюции жуков, поскольку данные палеонтологии, а также молекулярной генетики указывают на то, что именно в это время впервые появляются предки Polyphaga (Ponomarenko, 2003; Пономаренко, 2004; McKenna et al., 2015, 2019; Yan et al., 2017, 2018). В настоящее время до конца не ясно, как выглядели древнейшие представители Polyphaga, в чем заключались ранние этапы их эволюции и филогенетические отношения этой группы с оставшимися тремя рецентными подотрядами: Archostemata, Myxophaga и Adephaga (тем не менее, продемонстрирована монофилетичность всех четырех групп, см.: McKenna et al., 2015). К настоящему моменту уже накоплен большой объем информации по признакам как взрослых жуков, так и их личинок (особо стоит отметить стафилиноидную личинку Angarolarva Ponomarenko, 1985 из нижней–средней юры Сибири, местонахождения Усть-Балей, и триасовую Trialarva Prokin et Bashkuev, 2020 из верхнего триаса Германии, как примеры базально организованной личинки Polyphaga), а также молекулярные данные по большинству современных семейств (см. Lawrence et al., 2011; Bocak et al., 2014; McKenna et al., 2015). Таксономическое положение Polyphaga среди остальных трех подотрядов остается неоднозначным, и уже опубликованы работы со всеми возможными вариантами расхождения ветвей (Kukalová-Peck, Lawrence, 1993, 2004; Hunt et al., 2007; Beutel et al., 2008; Misof et al., 2014; McKenna et al., 2015). Подобные полномасштабные исследования, призванные разрешить крупный таксономический вопрос, демонстрируют высокую эффективность при выяснении филогенетических отношений на уровне видов и родов, но теряют разрешающую способность прямо пропорционально повышению ранга изучаемых таксонов (McKenna et al., 2015; McKenna, 2016), что, вероятно, связано с пока еще недостаточной выборкой задействованных таксонов, и будет решено в будущем. Получается, что первый из двух масштабных биологических вопросов Polyphaga, заключающийся в выяснении их филогенетических связей с другими подотрядами, решается в основном при помощи кладистического анализа больших объемов закодированной информации о морфологических признаках, а также методами молекулярной генетики. Тогда как решение второй задачи – кем были ранние Polyphaga, от кого они произошли, через какие эволюционные этапы они прошли на пути к становлению современных таксонов – почти целиком опирается на результаты палеонтологических исследований.
МАТЕРИАЛ И МЕТОДЫ
Сухие и смоченные 95% этанолом образцы просматривались под бинокуляром Leica М165С. Фотографии выполнены при помощи камеры Leica DFC 420, смонтированной на упомянутый бинокуляр, в приложении Leica Application Suite Version 3.4.1. Результирующие изображения получены с помощью Helicon Focus 5.1.28. Векторные рисунки по фотографиям и реконструкции выполнены в Corel Draw X8. Трехмерные модели .fbx выполнены в ZBrush 2020 по изображениям из соответствующих работ, результирующий рендер получен в Substance Painter версии 2020.2.2.
Условные обозначения: сплошные линии – границы склеритов и частей тела, борозды на склеритах; пунктирные линии – неотчетливые границы склеритов, частей тела и борозд, места повреждения отпечатка, места наложения одних частей тела на другие; точка-тире – бугры и рельеф поверхности склеритов. Длина всех масштабных линеек 1 мм. Классификация Coleoptera принимается по Дж. Лоуренсу (Lawrence et al., 2011). Обсуждаемая информация взята из опубликованных работ, поэтому данные об ископаемом материале специально не обсуждаются.
ИСТОРИЯ ИЗУЧЕНИЯ ДРЕВНЕЙШИХ POLYPHAGA И POLYPHAGA-ПОДОБНЫХ ФОРМ
Ранние гипотезы о строении, образе жизни и путях эволюции предковых Polyphaga выдвигались задолго до обнаружения реальных палеонтологических свидетельств (Crowson, 1960) и базировались на анализе морфологии современных групп жуков (т.е., такие базальные полифаги оказывались комбинацией из плезиоморфных состояний всех возможных морфологических признаков).
Среди предполагаемых предковых признаков указывались такие, чья проверка на палеонтологическом материале оказывается практически невозможной:
– потеря питающих клеток в фолликулах яичников;
– криптонефрические мальпигиевы сосуды;
– особенности строения метэндостернита (которые теоретически возможно наблюдать на отпечатках в качестве структур, проступающих через наружные покровы, если кутикула достаточно тонкая и позволяет видеть элементы эндоскелета, но это очень редкое явление).
Среди признаков редких, но подлежащих проверке на материале исключительно хорошей сохранности, указывались:
– потеря поперечных жилок на заднем крыле;
– нефункциональные дыхальца на восьмом сегменте брюшка;
– эдеагус с тегменом, схожим с таковым у Cucujoidea (или его производными).
Одним из немногих признаков, который надежно работал на ископаемом материале, был тип строения задних тазиков, которые не должны были иметь вырезки на заднем крае бедренной покрышки. Предполагалось (Crowson, 1960), что ранние Polyphaga были мелкими или очень мелкими жуками, подобными современным Myxophaga или Clambidae (Polyphaga: Scirtiformia).
На первый взгляд, подобные рассуждения хорошо соотносятся с находками вероятных предков Polyphaga на пермо-триасовой границе в местонахождении Бабий Камень (Волков, 2012) или рассматриваемыми ниже пермскими миниатюрными жуками, однако сходство это лишь поверхностное, связанное с мелкими размерами отпечатков (табл. V , фиг. 3–7 ; см. вклейку). В настоящее время большинство признаков, указанных в качестве “плезиоморфий базальных Polyphaga”, рассматриваются как продвинутые, специализированные признаки, например, криптонефрия является апоморфией Cucujiformia (см. Lawrence, Newton, 1982). Согласно схеме эволюции Polyphaga от очень мелких Myxophaga, предложенной Р. Кроусоном (Crowson, 1960), от них произошли некие “протополифаги”, которые, в свою очередь, дали сразу три эволюционных ветви: стафилиноидную (соответствовавшую современному инфраотряду Staphylinoidea), эуцинетоидную (Scirtoidea) и дерместоидную (Bostrichoidea). Предполагалось, что все три группы появляются в конце триаса или начале юры, а в позднем мелу вместе с покрытосеменными растениями возникают и наиболее продвинутые надсемейства. Сам процесс эволюции подотряда описывался как длительный и постепенный, с последовательным появлением все более продвинутых групп (Crowson, 1960, 1975, 1981).
По мере накопления фактического палеонтологического материала стало понятно, что личинки Polyphaga, эволюция которых считалась еще одним ключевым фактором эволюционного успеха подотряда, крайне редки в ископаемом состоянии в связи со своим скрытым образом жизни, часто непосредственно внутри питательного субстрата, и не смогут стать надежным свидетельством эволюционных преобразований Polyphaga на преимагинальных этапах. Еще одним препятствием в изучении стала общая тенденция Polyphaga к редукции тех или иных морфологических структур: исчезновению швов, уменьшению числа мышц и жилок заднего крыла, а, значит, связанная с этим высокая вероятность независимого приобретения одного и того же состояния признака (Kasap, Crowson, 1975; Пономаренко, 1983; Lawrence et al., 2011). Последующие работы только усилили недоверие к признакам строения тела, поскольку раннее отделение Polyphaga от всех остальных подотрядов указывало на то, что, например, упрощение торакальной скелето-мускульной системы имело место независимо у не-архостематных подотрядов, особенно у Myxophaga (Beutel, Haas, 2000; Friedrich et al., 2009). Следствием таких особенностей строения стали повышенные требования к степени сохранности изучаемого материала. Даже для проверки такого “несложного для наружного наблюдения” признака, как криптоплеврия, т.е. превращения проплевры переднегруди во внутреннюю структуру, обеспечивающую площадь для прикрепления мышц (одного из основных признаков, отличающих Polyphaga от прочих жуков), необходим материал весьма хорошей сохранности, встречающийся крайне редко. Наиболее распространенные остатки жуков – изолированные надкрылья – оказались практически невостребованными для палеонтологических исследований, т. к. их строение является неспецифичным и могло быть очень сходным у представителей разных семейств, тогда как у отдельных видов внутри одного рода разительно отличаться (Пономаренко, 1983).
Упомянутые теоретические рассуждения о ранних Polyphaga, которые выглядели как некие Myxophaga-подобные жуки (Crowson, 1960), подверглись критике, ведь оказывалось, что если представить себе гипотетического предка Polyphaga как собрание плезиоморфных состояний всех имеющихся у взрослого жука признаков, то он будет очень похож на представителя вымершего семейства Ademosynidae (Coleoptera: Archostemata), и лишь его проплевры (рис. 1, б, пр) будут полностью втянуты внутрь тела (Пономаренко, 1983).
Рис. 1.
Реконструкции облика пермских и триасовых жуков по опубликованным данным: а – Sylvacoleodes admirandus Ponomarenko, 1969; б – Dolichosyne sulcata Ponomarenko, 1969, из: Yan et al. (2017), с изменениями; в – Archosyne permiana Ponomarenko, Yan et Huang, 2014, из: Yan et al. (2017), с изменениями; г – Ponomarenkium belmonthense (Yan et al., 2017), из: Yan et al. (2017), с изменениями; д – Peltosyne triassica Ponomarenko, 1977, из: Yan et al. (2018), с изменениями. Обозначения: ан3 – метанэпистерн; пар – параноталии; пр – проплевра (также залита светло-серым; темно-серым отмечены открытые сзади впадины передних тазиков); со.г – субокулярный гребень; эп – эпиплевра. Темно-серым цветом отмечены открытые сзади впадины передних тазиков.
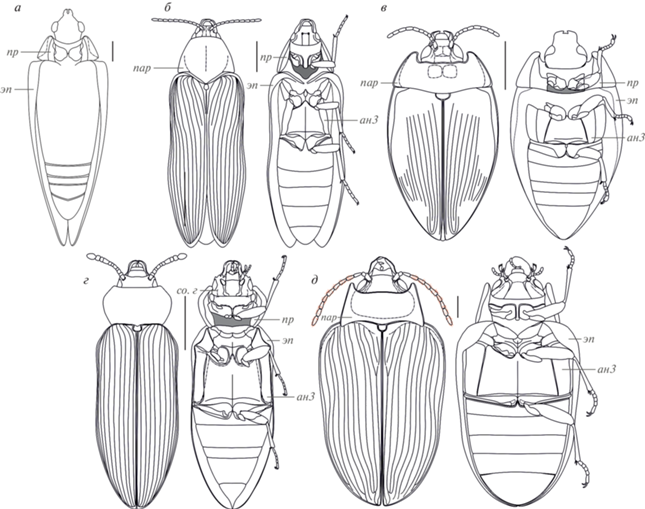
За последующие 30 лет с момента публикации исследования Кроусона (1960) удалось накопить достаточно ископаемого материала для построения предварительного облика древнейших Polyphaga (Lawrence, Newton, 1982; Пономаренко, 1983). Общими признаками предковой формы в обеих работах стали:
– наличие почти гомономных антенномеров;
– полностью или частично втянутые внутрь переднегруди проплевры;
– поперечные передние тазики, вырезанные сзади;
– поперечный метавентрит с отчетливым продольным и поперечным (паракоксальным) швами.
В основном полагающаяся на палеонтологические свидетельства концепция А.Г. Пономаренко (1983) изображала предковые формы Polyphaga схожими со Scirtiformia или отчасти со Staphyliniformia. Выбор этих двух групп можно объяснить устойчивыми представлениями о сциртоидах как о наиболее базальных Polyphaga, ввиду наличия у них наружных проплевр (как, напр., у Decliniidae), мембранозного сочленения мезо- и метавентрита и т.д.). В то же время, у стафилинид сохраняются такие нечастые для жуков признаки, как парные простые глазки (есть свидетельства даже о тройных глазках у единичной находки с Японских островов, однако это может быть морфологической абберацией, см.: Naomi, 1987), что “вписывалось” в гипотетический план строения древнейших Polyphaga. Из признаков указывались:
– Y-образный шов на голове (являющийся не настоящим швом, т.е. местом втягивания кутикулы внутрь тела, а линочной линией);
– мандибулы с отчетливой молярной частью и отчлененной простекой;
– наружные про- и мезотрохантины;
– надкрылья с десятью точечными бороздами и пришовной бороздкой;
– короткий мезовентрит без швов;
– мезококсальные впадины, не закрытые с боков метанэпистернами;
– метакоксы, идущие вдоль всего заднего края метавентрита и продолжающиеся вбок, отделяя брюшко от метанэпистернов;
– мембранозный второй брюшной стернит, закрытый метакоксами.
При этом отмечалась невозможность происхождения всего таксономического многообразия Polyphaga от миниатюризованных жуков, поскольку в таком случае необходимо допустить вторичное развитие сложных морфологических структур (таких как, например, жилкование крыльев Hydrophilidae, практически не несущее следов редукции), в обход закона Долло. Таким образом, по представлениям Пономаренко (1983), взрослый представитель древнейших Polyphaga оказывался короткоживущим насекомым, скорее всего, не питающимся, с чем и связывались многочисленные редукции в его строении. Все задачи по питанию возлагались на скрыто живущую личинку, обитавшую среди или внутри гниющей растительности, во влажных местообитаниях и являющуюся детрито- или мицетофагом.
Анализ находок из верхней перми и триаса позволил выявить группы-кандидаты на роль базальных Polyphaga. Одной из таких групп стало семейство Ademosynidae (рис. 1, б), описанное в подотряде Archostemata (Пономаренко, 1968). Представители адемосинид обладали нетипичными для Archostemata надкрыльями с точечными бороздами; некоторые исследователи предлагали считать их наиболее базальными (stem-group) Polyphaga (Crowson, 1981; Lawrence, 1999; Grimaldi, Engel, 2005), отмечая, тем не менее, их необычные, уплощенные тела с ярко выраженными параноталиями, наличие наружных проплевр и габитуальное сходство с уже упомянутыми Scirtidae. Наличие отчетливых паранотальных выростов (рис. 1, пар) и общая уплощенная форма пронотума рассматривались как серьезный аргумент в пользу примитивности Ademosynidae. Современные базальные полифаги, действительно, обычно имеют уплощенную переднегрудь с боковыми выростами, тогда как у большинства таксонов выросты сокращаются до ребра на боковой стороне переднегруди или исчезают полностью. Жуки без параноталий или продольного ребра на переднегруди практически не встречаются среди мезозойских форм, даже в таких группах, как Curculionidae, где округлая в сечении переднегрудь встречается почти у всех современных форм (Пономаренко, 1983).
Пономаренко (1977) не считал Ademosynidae непосредственным предком Polyphaga, указывая на их явственные проплевры и триасовый возраст; он предлагал обратить внимание на еще один монотипический род – Peltosyne Ponomarenko, 1977 (рис. 1, д) из того же триасового местонахождения, где были найдены адемосиниды Dolichosyne (Ponomarenko, 1969). Peltosyne, на первый взгляд, походил на некий “генерализованный” образ жука, сочетая в своем строении плезиоморфные состояния большинства диагностических признаков, однако, уже обладал внутренними проплеврами. Тем не менее, на момент описания дальнейшее изучение Peltosyne не было продолжено ввиду очень ограниченной типовой серии (три образца), и род получил статус “Polyphaga incertae sedis”.
В дальнейшем указанные группы были переизучены и переописаны. Причем для Ademosynidae подтвердилось первоначальное их определение как Archostemata, которые независимо развили множество признаков, сходных с Polyphaga; таким образом, адемосиниды стали ярким примером параллельной эволюции, приведшей к образованию тупиковой ветви (все они вымерли в начале мела). Peltosyne, наоборот, были выделены в отдельное семейство базальных Polyphaga. Также впервые был реконструирован облик предковых форм подотряда с основой на версии разных исследователей (Yan et al., 2017, рис. 8 ; Ян и др., 2018).
Проходящие параллельно исследования нуклеотидных генов по большей части согласуются с упомянутыми палеонтологическими находками, утверждая, что “высшие жуки” (т.е. представители подотрядов Adephaga и Polyphaga) появляются в поздней перми (253 млн л.н.), и только две филогенетические линии переживают позднепермское массовое вымирание. Представители всех четырех подотрядов жуков фиксируются в палеонтологической летописи к концу триаса (Пономаренко, 1983; Ponomarenko, 2002; Papier et al., 2005; McKenna et al., 2015). Временем разделения Polyphaga на современные инфраотряды (т.е., на Scirtiformia, Staphyliniformia, Scarabaeformia, Elateriformia, Derodontiformia, Bostrichiformia и Cucujiformia) также указан конец триаса или, самое позднее, начало юры. Следует отметить, что это пока лишь теоретические предсказания, и палеонтологические свидетельства для всех указанных таксонов известны из намного более молодых отложений. Самые продвинутые семейства возникают в мелу (Grimaldi, Engel, 2005; Hunt et al., 2007; McKenna, Farrell, 2009; McKenna et al., 2015, 2019).
ПЕРМСКИЕ ПРЕДСТАВИТЕЛИ ЖЕСТКОКРЫЛЫХ SENSU STRICTO
Поздняя пермь была временем расцвета Archostemata, а также близких к ним базальных групп жуков (т.н. stem-group Coleoptera). Совокупность таксономических групп, за исключением Tshekardocoleidae (рис. 1, а), Permocupedidae и Rhombocoleidae, понимается в настоящей работе как “Coleoptera sensu stricto” за вычетом базальных групп. Возникновение первых Polyphaga на границе перми и триаса предсказывалось задолго до получения данных по молекулярной филогении (Hunt et al., 2007; McKenna et al., 2019), на основе изучения изолированных надкрылий с рядами точечных борозд и их процентного соотношения с надкрыльями иного строения в различных местонахождениях (Пономаренко, 1983, 2016; Ponomarenko, 2003). Надкрылья с продольными рядами точечных бороздок, лишенные бугорчатой скульптуры, ячей и видимых жилок (т.н. “основных”, т.е. гомологичных крыловым жилкам), относят к семейству Permosynidae. Сходство в названии с Ademosynidae не случайно. Как уже было сказано, вымершие аберрантные Archostemata, такие как Ademosynidae, обладали надкрыльями именно такого типа (в современной фауне среди Archostemata надкрылья с точками известны только у Jurodidae). Тем не менее, массовость надкрылий с подобной организацией, особенно в местонахождениях Восточной Европы (напр., Аристово, конец верхней перми, чансингский век, самые низы вятки, около 252 млн л.н.), указывает на возможность того, что в действительности, по крайней мере часть из них, могла принадлежать и представителям Adephaga и Polyphaga. Как уже было сказано, неспецифичность строения надкрылий большинства жуков (не считая Archostemata) оставляет большой простор для спекуляций на тему об их таксономической принадлежности (поскольку для введения в практику анализа изолированных надкрылий требуется накопление больших объемов знаний по находкам из множества местонахождений. Тем не менее, это не является невозможным, стоит лишь взглянуть на методы изучения плейстоценовых Coleoptera), поэтому не подвергался сомнению лишь факт смены таксономического состава Coleoptera близ пермо-триасовой границы.
Если исключить Ademosynidae из кандидатов в предковые Polyphaga, наиболее древним местонахождением, из которого известны Polyphaga-подобные жуки, является местонахождение Иньпин на юге Китая (провинция Анхой) с остатками насекомых, найденными в одноименной свите. Возраст свиты Иньпин неоднократно пересматривался и изменялся от терминальной средней до начала поздней перми (см. Пономаренко и др., 2014). В настоящее время принято считать свиту Иньпин терминальной средней пермью (кэпитенский век, около 261 млн л.н., см.: Huang et al., 2007; Lin et al., 2010). Из темно-серых алевролитов Иньпин известно сразу несколько жуков, которые могут иметь отношение к высшим подотрядам Coleoptera. Первый из них – монотипический род Archosyne Ponomarenko, Yan et Huang, 2014.
Описанная по единственному отпечатку (полному, но достаточно плохо сохранившемуся экземпляру, что является обычным для палеозойских остатков) A. permiana Ponomarenko, Yan et Huang, 2014 (рис. 1, в) изначально была отнесена к Ademosynidae на основании наличия наружных проплевр и способа закрывания средних тазиковых впадин с участием передних краев метанэпистернов (рис. 1, в, ан3), что сделало ее древнейшим представителем семейства. Archosyne – очень маленький жук (длина тела около 4.3 мм), по-видимому, с сильно уплощенным телом и прочными покровами, несущими парные бугры на голове в местах прикрепления усиков и у переднего края пронотума. Еще одной особенностью Archosyne является сильно поперечный пронотум, почти втрое превышающий ширину головы. При этом уплощенные параноталии оказываются относительно узкими. Особенностью надкрылий было то, что они, скорее всего, лежали “плашмя” сверху на теле таким образом, что эпиплевра (рис. 1, эп) не прикрывала тело с боков (иначе пришлось бы предположить, что она была необычайно широкой). Единственный похожий пример такой организации сцепления между эпиплеврой и телом встречается у Tshekardocoleidae (см. Пономаренко, 1969, рис. 29, 30 ), относящихся к древнейшим (т.н. stem-group) предкам Coleoptera (рис. 1, а). Анализ вышеупомянутых признаков вынудил в более поздних работах исключить Archosyne из состава Ademosynidae (Yan et al., 2017). Среди других признаков, указывающих на необходимость ее отделения от Ademosynidae:
– наличие выступающих вперед передних углов пронотума;
– прямой передний край пронотума;
– интеркоксальный отросток пронотума более короткий, чем прококсы;
– наличие смежных мезококс;
– уплощенные основания надкрылий.
Archosyne получила статус “Coleoptera incertae sedis” до момента появления новых находок, потенциально способных уточнить ее связь с Polyphaga или с какой-то иной группой жуков. Archosyne являлась единственной находкой Coleoptera s. s. из Иньпин, получившей формальное описание. Однако, из того же местонахождения известны отпечатки двух других необычных жуков. У одного из них сходный с Archosyne размер (до 5 мм), но гораздо более удлиненное тело с прогнатной головой, почти не скрытой основанием пронотума, выступающими вперед мандибулами и крупными глазами. Габитуально такой жук очень близок к тому, что впоследствии был описан из Австралийского позднепермского (татарский век) местонахождения Бельмонт (Yan et al., 2017; Ян и др., 2018). Второй неописанный жук с вальковатым телом, гладкими покровами, надкрыльями без борозд и бугорков и поперечной прогнатной головой не имел отношения к Polyphaga, однако предварительно был определен как принадлежащий семейству Triaplidae (А.Г. Пономаренко, личн. сообщ.), считающемуся представителями базальных Adephaga. Жуки такого типа неизвестны из одновозрастных местонахождений европейской части России и Сибири, и появляются лишь в терминальной перми (чансингский век), делая, таким образом, Иньпин уникальным по фаунистическому комплексу жуков.
Следующей ключевой для понимания позднепермского этапа эволюции жуков современного типа является находка Ponomarenkium Yan et al., 2018 (рис. 1, г; табл. V , фиг. 1, 2 ), известного из немного более молодых отложений местонахождения Бельмонт, расположенного к северу от Сиднея (Новый Южный Уэльс, поздняя пермь, татарский век). В палеоэкологических реконструкциях территория, занятая местонахождением, интерпретируется как система болот и соединяющих их водных потоков. Находки насекомых Бельмонта происходят из тонких кремнистых сланцев свиты Коудас Бэй со средней толщиной в 30 см, выходящих на поверхность на площади около 20 км. Местонахождение образовалось в результате захоронения одновременно большого количества насекомых под оседающим после извержения вулкана пеплом (т.н. snapshot kill), что позволяет моделировать условия среды на основе процентного соотношения разных групп (Beattie, 2007). Ископаемый энтомокомплекс Бельмонт известен достаточно давно, с XIX в. (Knight, 1950), отсюда собраны несколько тыс. отпечатков насекомых из 11 отрядов, среди которых по количеству обнаруженных остатков преобладают Protelytroptera, Hemiptera, Mecoptera и Coleoptera (Riek, 1968). Последние составляют почти 20% от всех находок (Beattie, 2007). Однако, как и в случае с Иньпин, очень немногие отпечатки получили в итоге формальное описание, в основном это – изолированные надкрылья (Tillyard, 1924). Наиболее многочисленная группа среди жуков – Permosynidae. Также обнаружены Permocupedidae, Rhombocoleidae, Taldycupedidae, Asiocoleidae и Schizocoleidae (Yan et al., 2013).
Обладая сравнимыми с Archosyne размерами тела (4.5 мм), похожей прогнатной головой и надкрыльями с продольными бороздами (в качестве одного из важнейших элитральных признаков указывалась организация борозд на вершинах надкрылий; А.Г. Пономаренко, перс. сообщ.), Ponomarenkium, тем не менее, мало чем отличался от современных жуков (например, уже упомянутых в качестве примера возможных древнейших Polyphaga Scirtoidea, а точнее, Decliniidae). Он уже лишен сильно поперечного, имеющего форму плоского щита пронотума (а также и параноталий). Надкрылья, судя по всему, плотно соприкасаются с телом, подразумевая наличие запирающего механизма между эпиплеврой и тораксом (скорее всего, брюшко Ponomarenkium все еще не участвовало в фиксации надкрылий и формировании полноценной субэлитральной полости), тогда как надкрылья Archosyne фиксировались на теле лишь в районе плеч (самое основание надкрылья). Тем не менее, среди плезиоморфных признаков указываются: наличие субокулярных гребней на голове (рис. 1, г, со.г), сохранение обоих, продольного и поперечного (паракоксального) швов на мезо- и метавентрите, а также очень необычная, коническая форма пятичленикового брюшка с сильно заостренным, суженным последним стернитом. Несмотря на наличие проплевры (хотя она была заметно ýже, чем у Archosyne), у жука из Бельмонта уже отсутствовали метатрохантины, а метанэпистерны не принимали участия в закрывании с боков мезококсальных впадин. Покровы тела лишены примитивной бугристой скульптуры (табл. V , фиг. 1, 2 ) и были гладкими, либо слабопунктированными. Также отсутствуют бугры на голове и пронотуме (Yan et al., 2017). Без отнесения Ponomarenkium к одному из четырех подотрядов, авторы указывали, что этот таксон мог бы принадлежать как базальным представителям Polyphaga, так и Myxophaga. Однако, у него нет характерной черты последних – широкого, часто мембранозного щелевидного контакта между мезо- и метавентритом (см. Beutel, Haas, 2000). Еще одним важным событием для систематики древнейших жуков современного облика, связанным с обнаружением Ponomarenkium, стало выделение отдельного семейства для подобных жуков (Yan et al., 2017).
Ранние прогнозы о происхождении Polyphaga от форм, близких к Scirtoidea или Derodontidae, не подтверждаются реальными палеонтологическими находками пермских жуков, рассмотренными выше. По молекулярным данным, базальное разделение между Scirtoidea + Derodontidae и всеми остальными надсемействами Polyphaga произошло предположительно в триасе, примерно 229 млн л.н. (McKenna et al., 2015, 2019). По морфологическим признакам эти группы также противопоставляются всем остальным Polyphaga (см., напр., Lawrence et al., 2011), однако это, скорее всего, связано с наличием в матрице признаков наружных проплевр. К апоморфиям семейства можно отнести:
– очень узкий межтазиковый отросток простернума, фактически оставляющий передние тазики соприкасающимися;
– мезовентрит без ямки для вкладывания межтазикового отростка простернума;
– внутренние задние трохантины;
– апикальный брюшной стернит с заостренной, шиповидной вершиной.
При этом находки ископаемых Scirtoidea появляются не ранее юры и мела (Hong, 1995; Kaddumi, 2007; Kirejtshuk, Ponomarenko, 2010), так же как и юрских Derodontidae (Cai et al., 2014).
Среди возможных кормовых субстратов и мест обитания как для древнейших жуков, таких как Tshekardocoleidae, так и для пермо-триасовых предков высших подотрядов, указана древесина. Обычно подразумевают влажную, обильно пронизанную грибными гифами древесину, возможно, уже мертвых деревьев (Пономаренко, 1969; Ponomarenko, 2003; Grimaldi, Engel, 2005; Наугольных, Пономаренко, 2010). Однако существующие уже довольно давно представления о подобном образе жизни жуков во многом являются следствиями применения принципа актуализма, когда на базальных Coleoptera проецируют образ жизни современных Cupedidae и Ommatidae (Coleoptera: Archostemata), у которых древогрызущими являются только личинки (Hörnschemeyer, Yavorskaya, 2016; Hörnschemeyer, Beutel, 2016), тогда как для взрослых Cupedidae указывается питание пыльцой (Crowson, 1962). О находке ископаемых следов питания жуков древесиной сообщается из поздней перми Китая (чансингский век, около 254–252 млн л.н.). Из отложений, близких по возрасту тем, откуда был описан Ponomarenkium, происходят следы питания камбием и древесиной хвойных. Повреждение представляет собой уже очень сложную структуру с материнским ходом, прогрызенным через кору в камбий, где была организована яйцевая камера, от которой в одном направлении отходит гребенчатая последовательность личиночных ходов, заканчивающихся выходным отверстием. В современной фауне похожие повреждения оставляют специализированные ксилофаги и ксилобионты из инфраотрядов Bostrichiformia и Cucujiformia. Подобный пример заботы о потомстве оказывается одним из самых древних у жуков (Feng et al., 2017; см. также Feng et al., 2019, с примерами еще более древних повреждений древесины, не связанных, впрочем, с деятельностью Polyphaga). Подобные находки могут косвенно указывать на правильность предположений, упомянутых выше, о питании древесиной как о ключевом факторе, ставшем залогом невероятного разнообразия Polyphaga. Если принять это предположение, тогда питание древесиной также привело в будущем к развитию таких специализированных форм растительноядности, как минирование листьев и заражение растений грибами, специально принесенными насекомым в только что прогрызенный ход, что характерно для некоторых современных Platypodinae и Scolytinae (Johnson et al., 2018; McKenna et al., 2019).
ТРИАСОВЫЕ ПРЕДСТАВИТЕЛИ POLYPHAGA
Хотя считается, что кризисные события на границе перми и триаса не только не сказались отрицательно на эволюционном успехе жуков, но и послужили триггером к возникновению и расцвету современных подотрядов, находок, приходящихся на ранний триас, крайне мало. Это объясняется общим падением численности многих таксонов и их временному исчезновению из тафономического окна, а также распространением грубозернистых осадков, усложнявших сохранение отпечатков (Пономаренко, 2016).
Одно из немногих известных местонахождений данного возраста – Бабий Камень, расположенное в Кемеровской области, где в кедровских слоях мальцевской свиты было найдено необычайно большое количество остатков жуков (около 80 экз. к 2001 г. и несколько сотен на настоящий момент) и, что особенно важно, многие из которых представлены отпечатками целых тел (Ponomarenko, 2003; Yan et al., 2018). Несмотря на то, что возраст свиты до сих пор обсуждается и разными авторами интерпретируется как поздняя пермь (Садовников, Орлова, 1994; Lozovsky, Korchagin, 2013; Садовников, 2016) или ранний триас (Kozur, Weems, 2011), находки оттуда, несомненно, имеют ключевое значение для понимания ранней эволюции высших подотрядов жуков. Среди пока неописанных образцов (табл. V , фиг. 3–7 ) имеются мелкие (до 5 мм длиной) жесткокрылые удлиненно-овальной формы (немногим более чем вдвое длиннее своей ширины), обладающие крупными (слегка короче пронотума) головами, узким простернумом и близко расположенными средними тазиками. Их параноталии довольно узкие, что требует дополнительной проверки, однако на данный момент не удается обнаружить у них следы внешних проплевр. Перечисленные особенности (а также относительно хорошая для нижнего триаса степень сохранности) делают находки из местонахождения Бабий Камень очень перспективными для понимания становления Polyphaga в период активной перестройки экосистем, тем более что из этого же местонахождения описан самый ранний представитель второго по величине подотряда жуков, Adephaga (Yan et al., 2018).
Только к началу позднего триаса (карний, около 225 млн л. н.) появляются несомненные представители базальных Polyphaga – вымершее семейство Peltosynidae (рис. 1, д). Эти жуки разительно отличаются от всех более ранних претендентов на роль древнейших Polyphaga своими довольно крупными размерами, до 15 мм. Т.е., пока не объясненная тенденция на миниатюризацию, свойственная пермским и раннетриасовым жукам, проходит. Будучи непохожими ни на Scirtoidea, ни на Derodontoidea, и являясь скорее неким генерализованным представителем Coleoptera, Peltosynidae демонстрируют уже полностью внутреннюю проплевру, массивный грызущий ротовой аппарат, укороченный, полностью разделяющий передние тазики простернум, также продольный и поперечный швы на метавентрите и, что не встречается у современных Polyphaga, на мезовентрите. Для них также характерен слабо модифицированный и считающийся исходным для Polyphaga трехлопастной эдеагус. Оригинальное помещение Peltosyne в Polyphaga (Пономаренко, 1977) подтверждается отсутствием у них наружной плевры, открытыми сзади впадинами передних тазиков и метанэпистернами, не участвующими в образовании впадин средних тазиков. Последний признак также отличает Peltosynidae от Ademosynidae. Признак, объединяющий их со всеми “неархостематными” подотрядами – отсутствие наружного метатрохантина. Присутствие молы на мандибулах отдаляет Peltosyne от Archostemata и Adephaga и сближает с Polyphaga и Myxophaga. Относительно уплощенное тело, грубая скульптура покровов (лишенная при этом бугорков), крупные выступающие мандибулы с тупыми вершинами и мощным мезальным зубцом, и борозды для вкладывания антенн указывают на вероятную связь Peltosynidae с древесиной и подтверждают предположения о ксилобионтности и ксиломицетофагии, как об исходном для Polyphaga образе жизни. Многочисленные хвойные, птеридоспермы, цикадофиты и кордаиты, произраставшие по берегам речного оазиса Джайляучо (Buchwitz, 2011; Franeck et al., 2012), окруженного ксероморфными долинами с теплым континентальным климатом и сезонными изменениями температуры (Сикстель, 1962, 1965; Пономаренко, 1969), предоставляли бы в таком случае микроареалы для жизни и питания Peltosynidae. Можно предположить, что похожие жуки были среди первых насекомых-опылителей покрытосеменных и голосеменных, а полинофагия могла стать промежуточной диетой между генерализованными детритофагией, микофагией и сапрофагией (древнейшие способы питания, согласно Ponomarenko, 2003, для stem-group Coleoptera, появившиеся задолго до возникновения тесной связи между жуками и покрытосеменными растениями) и специализированной растительноядностью, как у современных Phytophaga (Rainford et al., 2015; Peris et al., 2017; Cai et al., 2018; Liu et al., 2018; McKenna et al., 2019).
Хотя пермский этап становления Polyphaga до сих пор остается наиболее загадочным, а таксоны, приведенные в качестве примеров, имеют статус incertae sedis или Coleoptera s. s., их связь с высшими подотрядами жуков сложно отрицать. Даже несмотря на то, что в оригинальном исследовании (Yan et al., 2017) Ponomarenkium указан как возможный предок и для Polyphaga, и для Myxophaga, в действительности реальные свидетельства ранних этапов эволюции миксофаг появляются лишь в триасе (Fikáček et al., 2019), причем уже демонстрируют сильное сходство с одним из современных семейств.
Можно предположить, что накопление палеонтологических данных приведет к переносу Ponomarenkiidae в один из существующих подотрядов, как это уже произошло с Peltosyne. Переходные формы между базальными жуками stem-group и Polyphaga следует искать в пограничных пермо-триасовых местонахождениях, таких как Бабий Камень, а также Бор-Тологой, Караунгир, Недуброво и Тихвинское.
Список литературы
Волков А.Н. Ископаемые жуки местонахождения Бабий Камень (пермь-триас Кузбасса) // Современная палеонтология: классические и новейшие методы – 2012. М.: ПИН РАН, 2012. С. 49–53.
Пономаренко А.Г. Жуки-архостематы юры Каратау // Юрские насекомые Каратау / Ред. Родендорф Б.Б. М.: Наука, 1968. С. 118–138.
Пономаренко А.Г. Историческое развитие жесткокрылых – архостемат. М.: Наука, 1969. 240 с. (Тр. Палеонтол. ин-та АН СССР. Т. 125).
Пономаренко А.Г. Состав и экологическая характеристика мезозойских жесткокрылых. Подотряд Adephaga. Инфраотряд Staphyliformia // Мезозойские жесткокрылые / Ред. Родендорф Б.Б. М.: Наука, 1977. С. 8–119 (Тр. Палеонтол. ин-та АН СССР. Т. 161).
Пономаренко А.Г. Историческое развитие жесткокрылых насекомых. Дисс. … докт. биол. наук. М., 1983. 413 с.
Пономаренко А.Г. Жуки (Insecta, Coleoptera) поздней перми–раннего триаса // Палеонтол. журн. 2004. № 2. С. 185–196.
Пономаренко А.Г. Насекомые во временных окрестностях пермо-триасового кризиса // Палеонтол. журн. 2016. № 2. С. 68–81.
Наугольных С.В., Пономаренко А.Г. Предполагаемые следы питания жуков в древесине кониферофита из казанского яруса Прикамья // Палеонтол. журн. 2010. № 4. С. 105–110.
Пономаренко А.Г., Ян Е.В., Хуан Д. Новые жуки (Coleoptera) из терминальной средней перми Китая // Палеонтол. журн. 2014. № 2. С. 90–98.
Садовников Г.Н. Эволюция биома траппового плато средней Сибири // Палеонтол. журн. 2016. № 5. С. 87–99.
Садовников Г.Н., Орлова Э.Ф. Таймырский ярус – терминальный ярус континентальной перми // Докл. Акад. наук. 1994. Т. 338. № 5. С. 658–661.
Сикстель T.A. Флора поздней перми и раннего триаса южной Ферганы // Стратиграфия и палеонтология Узбекистана и сопределенных районов. Т. 1. Ташкент: Изд-во Акад. наук УзССР, 1962. С. 284–414.
Сикстель T.A. Триасовые высшие растения Советской Азии // Тез. докл. к Межвед. совещ. по континентальным отложениям мезозоя и кайнозоя Советской Азии и их биостратиграфии (3–5 февраля 1965 г.). Л., 1965. С. 19–20.
Ян Е.В., Бейтель Р.Г., Битти Р., Лоуренс Дж.Ф. Ponomarenkium gen. nov., новое название для позднепермского рода Ponomarenkia Yan et al., 2017 (Insecta: Coleoptera) // Палеонтол. журн. 2018. № 2. С. 110.
Beattie R.G. The geological setting and palaeoenvironmental and palaeoecological reconstructions of the Upper Permian insect beds at Belmont, New South Wales, Australia // Afr. Invertebr. 2007. V. 48. P. 41–57.
Bernhardt P. Convergent evolution and adaptive radiation of beetle-pollinated angiosperms // Plant Syst. Evol. 2000. V. 222. P. 293–320.
Beutel R.G. Über Phylogenese und Evolution der Coleoptera (Insecta), insbesondere der Adephaga // Verhandl. Naturwiss. Ver. Hamburg (NF). 1997. V. 31. S. 1–164.
Beutel R.G., Ge S.Q., Hörnschemeyer T. On the head morphology of Tetraphalerus, the phylogeny of Archostemata and the basal branching events in Coleoptera // Cladistics. 2008. V. 24. P. 270–298. https://doi.org/10.1111/j.1096-0031.2007.00186.x
Beutel R.G., Haas F. Phylogenetic relationships of the suborders of Coleoptera (Insecta) // Cladistics. 2000. V. 16. P. 103–141. https://doi.org/10.1006/clad.1999.0124
Bocak L., Barton C., Crampton-Platt A. et al. Building the Coleoptera tree-of-life for >8000 species: composition of public DNA data and fit with Linnaean classification // Syst. Entomol. 2014. V. 39. P. 97–110. https://doi.org/10.1111/syen.12037
Booth R.G., Cox M.L., Madge R.B. Guides to Insects of Importance to Man. 3. Coleoptera. L.: Intern. Inst. of Entomology (C.A.B. International), 1990. 384 p.
Buchwitz M. Taxonomy, phylogeny, and palaeobiology of the Madygen Tetrapod Fauna // Unpubl. thesis. Freiberg, TU Bergakademie, 2011.
Cai C., Escalona H.E., Li L. et al. Beetle pollination of cycads in the Mesozoic // Current Biol. 2018. V. 28. P. 1–7. https://doi.org/10.1016/j.cub.2018.06.036
Cai C.Y., Lawrence J.F., Ślipiński A., Huang D.Y. First fossil tooth-necked fungus beetle (Coleoptera: Derodontidae): Juropeltastica sinica gen. n. sp. n. from the Middle Jurassic of China // Eur. J. Entomol. 2014. V. 111. P. 299–302.
Calderón-Cortés N., Watanabe H., Cano-Camacho H. et al. cDNA cloning, homology modelling and evolutionary insights into novel endogenous cellulases of the borer beetle Oncideres albomarginata chamela (Cerambycidae) // Insect Mol. Biol. 2010. V. 19. P. 323–336. https://doi.org/10.1111/j.1365-2583.2010.00991.x
Costello M.J., Wilson S., Houlding B. Predicting total global species richness using rates of species description and estimates of taxonomic effort // Syst. Biol. 2012. V. 61. P. 871–883. https://doi.org/10.1093/sysbio/syr080
Crowson R.A. The phylogeny of Coleoptera // Ann. Rev. Entomol. 1960. V. 5. P. 111–134.
Crowson R.A. Observations on the beetle family Cupedidae, with description of two new fossil forms and a key to the recent genera // Ann. Mag. Natur. Hist. 13 ser. 1962. V. 51. P. 147–157. https://doi.org/10.1080/00222936208651227
Crowson R.A. The evolutionary history of Coleoptera as documented by fossil and comparative evidence // Atti del Congr. Nazionale Italiano di Entomologia, Sassari – 20–25 Maggio 1974. Firenze, 1975. P. 47–90.
Crowson R.A. The Biology of the Coleoptera. L., N.Y., Toronto et al.: Acad. Press, 1981. 802 p.
Farrell B.D. “Inordinate fondness” explained: Why are there so many beetles? // Science. 1998. V. 281. P. 555–559. https://doi.org/10.1126/science.281.5376.555
Feng Z., Bertling M., Noll R. et al. Beetle borings in wood with host response in early Permian conifers from Germany // Paläontol. Z. 2019. Bd 93. № 3. P. 409–421. https://doi.org/10.1007/s12542-019-00476-9
Feng Z., Wang J., Rößler R. et al. Late Permian wood-borings reveal an intricate network of ecological relationships // Nature Commun. 2017. V. 8. № 556. P. 1–6. https://doi.org/10.1038/s41467-017-00696-0
Fikáček M., Beutel R.G., Cai C. et al. Reliable placement of beetle fossils via phylogenetic analyses – Triassic Leehermania as a case study (Staphylinidae or Myxophaga?) // Syst. Entomol. 2019. V. 45. P. 175–187. https://doi.org/10.1111/syen.12386
Franeck F., Schneider J.W., Fischer J. et al. Mircovertebrate remains from the non-marine Triassic Madygen Formation of Kyrgyzstan // Thesis of Centenary Meeting of the Paläontologische Gesellschaft: Programme, abstracts, and field guides (24–29.09.2012). Berlin: Alfred-Wegener Stiftung, 2012. V. 3. P. 57–58.
Friedrich F., Farrell B.D., Beutel R.G. The thoracic morphology of Archostemata and the relationships of the extant suborders of Coleoptera (Hexapoda) // Cladistics. 2009. V. 25. P. 1–37.
Grimaldi D., Engel M.S. Evolution of the Insects. Cambridge: Cambridge Univ. Press, 2005. 755 p.
Hong Y.C. Fossil insects of the southern Ordos Basin // Acta Geol. Gansu. 1995. V. 4. № 1. P. 1–13.
Hörnschemeyer T., Beutel R.G. Ommatidae Sharp & Miur, 1912 // Handbook of Zoology, Vol. IV: Arthropoda: Insecta. Pt 38 Coleoptera, Beetles, Vol. 3: Morphology and Systematics (Phytophaga) / Eds. Leschen R.A.B., Beutel R.G. Berlin: Walter de Gruyter, 2016. P. 52–57.
Hörnschemeyer T., Yavorskaya M. Cupeididae Laporte, 1836 // Handbook of Zoology, Vol. IV: Arthropoda: Insecta. Pt 38 Coleoptera, Beetles, Vol. 3: Morphology and Systematics (Phytophaga) / Eds. Leschen R.A.B., Beutel R.G. Berlin: Walter de Gruyter, 2016. P. 44–48.
Huang D.Y., Nel A., Lin Q.B., Dong F.B. The first Glosselytrodea (Insecta) from the latest Middle Permian of Anhui Province, China // Bull. Soc. Entomol. France. 2007. V. 112. № 2. P. 179–182.
Hunt T., Bergsten J., Levkanicova Z. et al. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation // Science. 2007. V. 318. P. 1913–1916. https://doi.org/10.1126/science.1146954
Johnson A.J., McKenna D.D., Jordal B.H. et al. Phylogenomics clarifies repeated evolutionary origins of inbreeding and fungus farming in bark beetles (Curculionidae, Scolytinae) // Mol. Phylog. Evol. 2018. V. 127. P. 229–238.https://doi.org/10.1016/j.ympev.2018.05.028
Kaddumi H.F. Amber of Jordan. The Oldest Prehistoric Insects in Fossilized Resin. Amman: Eternal River Mus. of Natur. History, 2007. 298 p.
Kasap H., Crowson R.A. A comparative anatomical study of Elateriformia and Dascilloidea // Trans. R. Entomol. Soc. London. 1975. V. 126. P. 441–495. https://doi.org/10.1111/j.1365-2311.1975.tb00858.x
Kirejtshuk A.G., Ponomarenko A.G. A new coleopterous family Mesocinetidae fam. nov. (Coleoptera: Scirtoidea) from Late Mesozoic and notes on fossil remains from Shar-Teg (Upper Jurassic, South-Western Mongolia) // Zoosyst. Ross. 2010. V. 19. № 2. P. 301–325.
Kirsch R., Gramzow L., Theißen G. et al. Horizontal gene transfer and functional diversification of plant cell wall degrading polygalacturonases: Key events in the evolution of herbivory in beetles // Insect Biochem. and Mol. Biol. 2014. V. 52. P. 33–50. https://doi.org/10.1016/j.ibmb.2014.06.008
Knight O. Le M. Fossil insect beds of Belmont, NSW // Rec. Austral. Museum. 1950. V. 22. P. 251–254.
Kozur H.W., Weems R.E. Detailed correlation and age of continental late Changhsingian and earliest Triassic beds: implications for the role of the Siberian Trap in the Permian-Triassic biotic crisis // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2011. V. 308. P. 22–40. https://doi.org/10.1016/j.palaeo.2011.02.020
Kukalová-Peck J., Lawrence J.F. Evolution of the hind wing in Coleoptera // Canad. Entomol. 1993. V. 125. P. 181–258. https://doi.org/10.4039/Ent125181-2
Kukalová-Peck J., Lawrence J.F. Relationships among coleopteran suborders and major endoneopteran lineages: evidence from hind wing characters // Eur. J. Entomol. 2004. V. 101. P. 95–144.
Lawrence J.F. The Australian Ommatidae (Coleoptera: Archostemata): New species, larva and discussion of relationships // Invertebr. Taxonomy. 1999. V. 13. P. 369–390.
Lawrence J.F., Newton A.F. Evolution and classification of beetles // Ann. Rev. Ecol. and Syst. 1982. V. 13. P. 261–290. https://doi.org/10.1146/annurev.es.13.110182.001401
Lawrence J.F., Ślipiński A., Seago A.E. et al. Phylogeny of the Coleoptera based on morphological characters of adults and larvae // Ann. Zoologici (Warszawa). 2011. V. 61. № 1. P. 1–217. https://doi.org/10.3161/000345411X576725
Leschen R.A.B. Evolutionary patterns of feeding in selected Staphylinoidea (Coleoptera): shifts among food textures // Functional Morphology of Insect Feeding / Eds. Schaefer C.W., Leschen R.A.B. Lanham, MD: Th. Say Publ. in Entomol.; Entomol. Soc. America, 1993. P. 59–104.
Leschen R.A.B., Buckley T.R. Multistate characters and diet shifts: evolution of Erotylidae (Coleoptera) // Syst. Biol. 2007. V. 56. P. 97–112. https://doi.org/10.1080/10635150701211844
Lin Q.B., Nel A., Huang D.Y. The first agetopanorpine mecopteroid insect from Middle Permian of China (Insecta: Mecoptera: Permochoristidae) // Ann. Soc. Entomol. France (N.S.). 2010. V. 46. № 1–2. P. 62–66. https://doi.org/10.1080/00379271.2010.10697639
Liu Z., Ślipiński A., Lawrence J.F. et al. Palaeoboganium gen. nov. from the Middle Jurassic of China (Coleoptera: Cucujoidea: Boganiidae): The first cycad pollinators? // J. Syst. Palaeontol. 2018. V. 16. P. 351–360. https://doi.org/10.1080/14772019.2017.1304459
Lozovsky V., Korchagin O. The Permian period ended with the impact of a “Siberia” comet on earth // Bull. New Mexico Mus. Natur. Hist. and Sci. 2013. V. 60. P. 224–229.
McKenna D.D. Molecular phylogenetics and evolution of Coleoptera // Handbook of Zoology, Vol. IV: Arthropoda: Insecta. Pt 38 Coleoptera, Beetles, Vol. 3: Morphology and Systematics (Phytophaga) / Eds. Leschen R.A.B, Beutel R.G. Berlin: Walter de Gruyter, 2016. P. 1–11.
McKenna D.D., Farrell B.D. Tropical forests are both evolutionary cradles and museums of leaf beetle diversity // Proc. Nat. Acad. Sci. USA. 2006. V. 103. P. 1047–1051. https://doi.org/10.1073/pnas.0602712103
McKenna D.D., Farrell B.D. Beetles (Coleoptera) // The Timetree of Life / Eds. Hedges S.B., Kumar S. Oxford: Oxford Univ. Press, 2009. P. 278–289.
McKenna D.D., Shin S., Ahrens D. et al. The evolution and genomic basis of beetle diversity // Proc. Nat. Acad. Sci. U.S.A. 2019. V. 116. № 49. P. 24729–24737. https://doi.org/10.1073/pnas.1909655116
McKenna D.D., Wild A.L., Kanda K. et al. The beetle tree of life reveals Coleoptera survived end Permian mass extinction to diversify during the Cretaceous terrestrial revolution // Syst. Entomol. 2015. V. 40. P. 835–880. https://doi.org/10.1111/syen.12132
Misof M., Liu S., Meusemann K. et al. Phylogenomics resolves the timing and pattern of insect evolution // Science. 2014. V. 346. P. 763–767.
Naomi S.I. Comparative morphology of the Staphylinidae and the allied groups (Coleoptera, Staphylinoidea) // Jap. J. Entomol. 1987. V. 55. P. 450–458.
Oberprieler R.G., Marvaldi A.E., Anderson R.S. Weevils, weevils, weevils everywhere // Zootaxa. 2007. V. 1668. № 1. P. 491–520. https://doi.org/10.11646/zootaxa.1668.1.24
Papier F., Nel A., Grauvogel-Stamm L., Gall J.C. La diversité des Coleoptera (Insecta) du Trias dans le nordest de la France // Geodiversitas. 2005. V. 27. № 2. P. 181–199.
Peris D., Pérez-de la Fuente R., Peñalver E. et al. False blister beetles and the expansion of Gymnosperm-insect pollination modes before Angiosperm dominance // Curr. Biol. 2017. V. 27. P. 897–904. https://doi.org/10.1016/j.cub.2017.02.009
Ponomarenko A.G. Superorder Scarabaeida Laicharting, 1781. Order Coleoptera Linne, 1758. The beetles // History of Insects / Eds. Rasnitsyn A.P., Quicke D.L.J. Dordrecht: Kluwer Academic, 2002. P. 167–176.
Ponomarenko A.G. Ecological evolution of beetles (Insecta: Coleoptera) // Acta Zool. Cracov. 2003. V. 46 (suppl. – Fossil Insects). P. 319–328.
Rainford J.L., Mayhew P.J. Diet evolution and clade richness in Hexapoda: A phylogenetic study of higher taxa // Amer. Natur. 2015. V. 186. P. 777–791. https://doi.org/10.1086/683461
Riek E.F. Undescribed fossil insects from the Upper Permian of Belmont, New South Wales (with an appendix listing the described species) // Rec. Austral. Museum. 1968. V. 27. P. 303–310.
Robertson J.A., Ślipiński A., Moulton M. et al. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia) // Syst. Entomol. 2015. V. 40. P. 745–778. https://doi.org/10.1111/syen.12138
Ślipiński S.A., Leschen R.A.B., Lawrence J.F. Order Coleoptera Linnaeus, 1758 // Animal Biodiversity: an Outline of Higher-level Classification and Survey of Taxonomic Richness / Ed. Zhang Z.-Q. Auckland: Magnolia Press, 2011. P. 203–208 (Zootaxa. V. 3148).
Stork N.E., McBroom J., Gely C., Hamilton A.J. New approaches narrow global species estimates for beetles, insects, and terrestrial arthropods // Proc. Nat. Acad. Sci. USA. 2015. V. 112. P. 7519–7523. https://doi.org/10.1073/pnas.1502408112
Tillyard R.J. Upper Permian Coleoptera and a new order from the Belmont beds, New South Wales // Proc. Linn. Soc. New South Wales. 1924. V. 49. P. 429–435.
Wiegmann B.M., Trautwein M.D., Winkler I.S. et al. Episodic radiations in the fly tree of life // Proc. Nat. Acad. Sci. USA. 2011. V. 108. P. 5690–5695. https://doi.org/10.1073/pnas.1012675108
Yan E.V., Beutel R.G., Lawrence J.F. Whirling in the late Permian: ancestral Gyrinidae show early radiation of beetles before Permian-Triassic mass extinction // BMC Evol. Biol. 2018. V. 18:33. P. 1–10. https://doi.org/10.1186/s12862-018-1139-8
Yan E.V., Lawrence J.F., Beattie R., Beutel R.G. At the dawn of the great rise: Ponomarenkia belmonthensis (Insecta: Coleoptera), a remarkable new Later Permian beetle from the Southern Hemisphere // J. Syst. Palaeontol. 2017. V. 16. № 7. P. 611–619. https://doi.org/10.1080/14772019.2017.1343259
Yan E.V., Ponomarenko A.G., Beattie R., Wang B. Beetle body fossils and new elytra from the Upper Permian of New South Wales // 6th Intern. Congr. on Fossil Insects, Arthropods and Amber. Lebanon, Byblos, 2013. P. 31.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал