

Палеонтологический журнал, 2022, № 3, стр. 61-70
О первой находке Soergelia minor (Artiodactyla, Bovidae) в нижнем плейстоцене пещеры Таврида в Крыму и истории рода Soergelia
И. А. Вислобокова *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: ivisl@paleo.ru
Поступила в редакцию 27.10.2021
После доработки 25.11.2021
Принята к публикации 26.11.2021
- EDN: WIOLKJ
- DOI: 10.31857/S0031031X22030163
Аннотация
Изучение остатков Caprinae (рогового стержня, фрагментов верхней и нижней челюстей), найденных в 2020 г. в нижнеплейстоценовых отложениях карстовой пещеры Таврида в Крыму, показало их принадлежность Soergelia minor Moya-Sola, 1987. Эти ископаемые остатки – первое свидетельство присутствия рода Soergelia Schaub, 1951 в Восточной Европе. Вид S. minor был распространен в раннем плейстоцене на юге Западной Европы, а в Крыму найден впервые. Находки Soergelia в Тавриде важны для уточнения особенностей эволюции и распространения рода.
ВВЕДЕНИЕ
Пещера Таврида у пос. Зуя к востоку от Симферополя – крупное местонахождение остатков плейстоценовых позвоночных, открытое в 2018 г. при прокладке федеральной автотрассы “Таврида”. Пещера находится на Внутренней гряде Крымских гор в междуречье рек Бештерек и Фундуклы. По составу млекопитающих основные костеносные слои относятся к нижнему плейстоцену и имеют возраст 1.8–1.5 млн лет, а фауна сопоставляется с псекупским комплексом Восточной Европы и с поздним виллафранком Западной Европы (Лопатин и др., 2019). В 2020 г. новые интересные находки из основного слоя были сделаны в южном коридоре. В их числе оказались остатки полорогого вымершего рода Soergelia Schaub, 1951.
Род Soergelia относится к подсемейству Caprinae Gill, 1872 семейства Bovidae Gray, 1821 и обычно включается в состав трибы Ovibovini Gray, 1872. Он отличается от всех других родов особенностями положения и строения роговых стержней. Этот род известен по редким находкам из плейстоцена Евразии и Северной Америки (Shaub, 1951; Шер, 1971; Harington, 1980, 1987; Vekua, 1995; Васильев, 2005, 2010, 2011; Бондарев, Голубев, 2018; Боескоров, 2019). В Евразии Soergelia были распространены примерно с 1.8 млн л.н. и в Европе представлены четырьмя видами: S. elisabethae Schaub, 1951 (типовой вид), S. minor Moya-Sola, 1987, S. brigittae Kostopoulos, 1997 и S. intermedia Cregut-Bonnoure et Dimitrijevic, 2006. Диагноcтика видов основывается на размерах и особенностях строения роговых стержней и щечных зубов.
В Европе остатки зоргелий присутствуют в местонахождениях Испании, Франции, Италии, Германии, Нидерландов, Греции, Черногории, Болгарии и Румынии (Radulesco, Samson, 1965; Kahlke, 1963, 1969; De Giuli, Masini, 1983; Kostopoulos, 1997; Kolfschoten, Vervoort-Kerkhoff, 1999; Bukhsianidze, 2005; Crégut-Bonnoure, Dimitrijević, 2006; Fernandez, Crégut, 2007; Martinez-Navarro et al., 2012). На территории Европы к востоку от р. Прут они найдены впервые.
Данные о морфологии этих животных не полны, и многие вопросы таксономии, эволюции, родственных связей и распространения требуют дальнейшего изучения. Остатки из пещеры Таврида представляют особый интерес, так как принадлежат одной из древнейших популяций зоргелий. Изучение материала из Тавриды дает новую важную информацию об истории зоргелий и расширяет представления о биоразнообразии раннеплейстоценовой фауны Тавриды и региона в целом.
Автор выражает благодарность Д.Б. Старцеву (Крымский федеральный ун-т им. В.И. Вернадского), Д.О. Гимранову (Ин-т экологии растений и животных УрО РАН) и А.В. Лаврову (Палеонтологический ин-т им. А.А. Борисяка РАН, ПИН) за сбор материалов в 2020 г., и всем участникам раскопок. Автор благодарит своих коллег Д. Костопулоса (Ун-т Аристотеля, Салоники) за предоставленную возможность знакомства с ископаемыми материалами из Аполлонии-1, Я. ван дер Мада (Национальный музей естественной истории, Мадрид), М. Буксианидзе (Национальный музей Грузии, Тбилиси) и Р.-Д. Кальке (Исследовательский отдел четвертичной палеонтологии Зенкенберга, Веймар) за присланные статьи и фотографии ископаемого материала, а также Р.А. Ракитова (ПИН) за томографию рогового стержня из Тавриды.
ОПИСАНИЕ И СРАВНЕНИЕ МАТЕРИАЛА
СЕМЕЙСТВО BOVIDAE GRAY, 1821
ПОДСЕМЕЙСТВО CAPRINAE GILL, 1872
Род Soergelia Schaub, 1951
Soergelia minor Moya-Sola, 1987
Soergelia minor: Moyá-Solá, 1987, с. 192–207, рис. 8 ; Kostopoulos, 1997, с. 862, рис. 8–21 , табл. 3 ; Crégut-Bonnoure, Dimitrijević, 2006, с. 743; Fernandez, Crégut, 2007, с. 433; Martinez-Navarro et al., 2012, с. 99, рис. 2C.
Голотип – Ин-т палеонтологии им. Мигеля Крусафонта, г. Сабадель, VM-85 Sl N 24-39, череп с роговыми стержнями, изображен в работе (Moyá-Solá, 1987, рис. 8 ); Испания, местонахождение Вента Мицена-2; нижний плейстоцен.
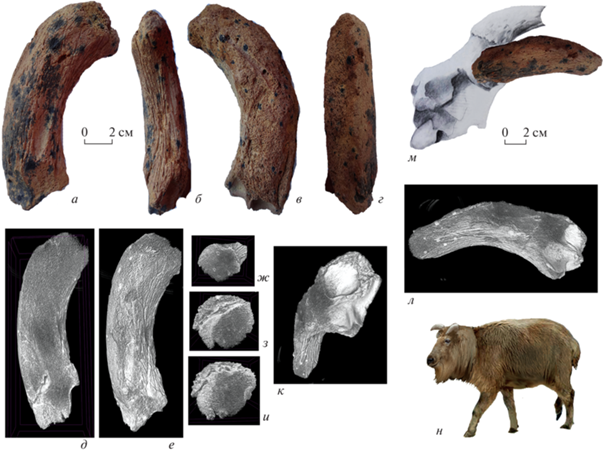
Описание (рис. 1–3). Роговой стержень экз. ПИН, № 5644/107 полный, с частично разрушенной внутренней стороной (рис. 1). Рог был направлен вверх и немного наружу, отогнут вперед и слегка гетеронимно скручен. Роговой стержень не очень массивный, слабо уплощенный, с чуть заметным передневнутренним килем (гребнем) и небольшим продольным желобом на внешней стороне. Контакт рогового стержня с пеньком хорошо виден на задней и наружной стороне образца. Костная ткань рогового стержня довольно плотная, с редкими каналами для крупных сосудов. Синусы лобной кости заходят лишь в основание рога, на высоту 14.8 мм. Поверхность рогового стержня неравномерно покрыта бороздами. Сечение рогового стержня овальное.
Рис. 1.
Soergelia minor Moya-Sola, 1987: а–г – экз. ПИН, № 5644/107, правый роговой стержень с пеньком снаружи (а), спереди (б), изнутри (в), сзади (г); д–и – его томография: продольное сечение (д), снаружи (е), поперечные сечения (ж–и); к, л – ЗD модели: со стороны синусов лобной кости (к), с внутренней стороны (л); м – реконструкция положения рогового стержня (на основе голотипа: по Moyá-Solá, 1987, рис. 8 ); н – реконструкция облика; Крым, пещера Таврида; нижний плейстоцен.
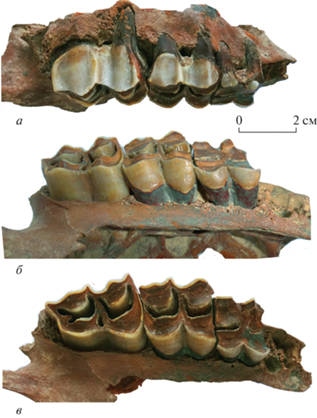
Верхнечелюстная кость экз. ПИН, № 5644/108 обломана сверху и спереди за альвеолой P4 (рис. 2). Коронка М1 стерта почти до основания. Коронки М2 и М3 удлиненные, расширяющиеся к основанию; задние половины коронок длиннее передних; передние лингвальные полулуния более угловатые. На буккальной стороне коронок хорошо развиты парастиль, мезостиль и метастиль, а также столбики паракона и метакона. Парастиль и мезостиль довольно широкие и округлые, умеренно выступающие, они чуть вздуты в основании. Столбики паракона и метакона широкие. На М3 метастиль слегка оттянут кзади, есть хорошо выраженная шпора в задней долинке (марке) и островок, удлиненный букко-лингвально, как у представителей Ovibovini.
Рис. 2.
Soergelia minor Moya-Sola, 1987, экз. ПИН, № 5644/108, правая верхнечелюстная кость с М1–М3: а – с буккальной, б – с лингвальной, в – с окклюзиальной сторон; Крым, пещера Таврида; нижний плейстоцен.
Нижнечелюстная кость экз. ПИН, № 5644/109 с обломанной восходящей ветвью (рис. 3). Тело нижней челюсти слабо вздуто под М3, сужается и понижается к P2; его вентральный край на уровне P2–M3 слабовыпуклый. Длина диастемы от P2 до С немного меньше альвеолярной длины P2–M3. Верхняя поверхность здесь (между P2 и симфизом) округленная, гребень почти не выражен. Резцовая часть тела, с не полностью сохранившимся альвеолярным краем, расширяется кпереди; ее дорсальная (лингвальная) поверхность узкая, длинная, слабовогнутая. Длина симфиза примерно равна альвеолярной длине ряда премоляров. Маленькое, узкое нижнечелюстное отверстие лежит под P2. Более крупное подбородочное отверстие, с коротким сосудистым желобом перед ним, расположено на расстоянии примерно 11 мм позади симфиза.
Рис. 3.
Soergelia minor Moya-Sola, 1987, экз. ПИН, № 5644/109: а–г – правая ветвь нижнечелюстной кости с PЗ–M3: а, г – снаружи, б – изнутри, в – сверху; д–ж – PЗ–M3: д – с буккальной, е – с лингвальной, ж – с окклюзиальной сторон; з – PЗ–P4 с окклюзиальной стороны; Крым, пещера Таврида; нижний плейстоцен.
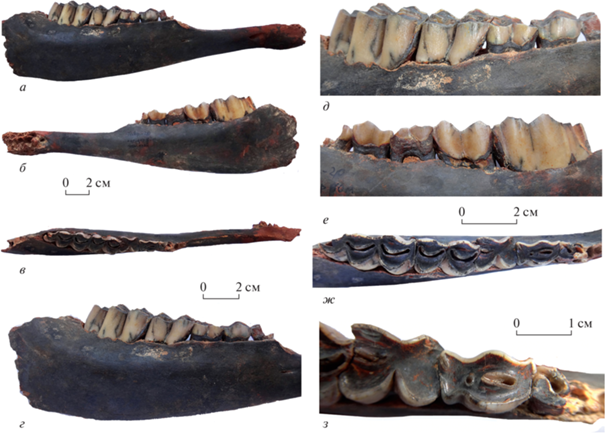
Ряд премоляров сравнительно длинный, его альвеолярная длина чуть больше половины длины ряда моляров. Р3 сохранился лишь частично. Метаконид лингвально уплощенный, с длинным задним крылом, протягивающимся почти до заднего края коронки. Энтоконид занимает косое положение относительно переднезадней оси зуба. Третья долинка расширена и углублена на переднем конце, сужена кзади и открывается на лингвальной стороне у задневнутреннего угла коронки. P4 удлиненный, узкий. Индекс его длины – 58%. Задняя доля коронки P4 длинная, ее длина равна примерно 1/3 длины коронки. Метаконид удлиненный, слабовыпуклый лингвально; крылья метаконида сливаются с параконидом и энтоконидом; при этом на лингвальной стороне желобки между ними широкие. Вторая и третья долинки замкнутые. Tретья долинка маленькая, округлая, лежит ближе к буккальному краю. Энтоконид скошен назад и четвертая долинка, очень узкая и косая, открывалась у задневнутреннего угла коронки. Протоконид очень слабо выпуклый и слегка угловатый, на буккальной стороне он отделен от гипоконида глубокой V-образной бороздкой (буккальной долинкой), почти достигающей третьей долинки на жевательной поверхности. Гипоконид небольшой, угловатый. Моляры удлиненные, без метастилида. Передние половины коронок M2 и M3 длиннее и шире, чем задние. На M1 есть очень слабо выраженный эктостилид. На M2 и M3, у переднего наружного угла коронок, присутствует цингулум. M3 с удлиненной третьей долей, она полуокруглая с буккальной стороны.
Размеры в мм, индексы в %. Экз. ПИН, № 5644/107, роговой стержень: длина (L), по прямой – сa. 112, по большой кривизне – сa. 150; переднезадний и поперечный диаметры основания (DAP × DT) – 45.6 × 35.01; индекс сжатия основания рогового стержня (DT/DAP) – 77.
Экз. ПИН, № 5644/108, верхние моляры (длина × ширина, L × W): M1 – 15.40 × 17.63; M2 – 21.92 × 20.69; M3 – 27.08 × 20.81; длина M1–M3 – 62.36.
Экз. ПИН, № 5644/109, нижняя челюсть: высота тела впереди Р2 – 25.46; его ширина впереди Р2 – 11.72, высота под Р3 – 34.24; ширина под Р3 – 13.24; высота под М3 – 42.63; ширина под М3 – 22.29; высота позади М3 – 50.82; длина диастемы – ca. 86.6; высота в середине диастемы – 17.22; ширина – 11.67; длина симфиза – ca. 40.8; длина P2‒P4 – 38.37; M1–M3 – 70.95; P2–M3 – 107.8; индекс P2–P4/P2–M3 – 36; индекс P2–P4/M1–M3 – 54.
Экз. ПИН, № 5644/109, нижние щечные зубы (L × W) в мм: P4 – 14.97 × 8.68; M1 – 15.89 × 12.68; M2 – 22.78 × 12.74; M3 – 33.22 × 13.88; длина и ширина передней и задней долей P4: La ×Wa – 10.25 × × 8.68; Lp × Wp – 4.74 × 8.02; его индексы Lp/L – 32; Wa/L – 58; Wp/Wa – 92.
Сравнение и замечания. По морфологии, размерам и пропорциям образцы из Тавриды соответствуют S. minor Moya-Sola, 1987 из нижнего плейстоцена Вента Мицены (Испания; MNQ20, 1.3 млн л.н., MmQ-2; Moyá-Solá, 1997, рис. 8–12, 14, 17 , Lamina 3) (рис. 4, 5, табл. 1). Роговой стержень из Тавриды немного мельче, чем у голотипа S. minor – VM-85 Sl N 24-39, хранящегося в Ин-те палеонтологии им. Мигеля Крусафонта в Сабаделе. Но он попадает в диапазон изменчивости роговых стержней S. minor из Вента Мицены, DAP – 41.2–51.9; DT – 32.7–38.5; индекс сжатия – 62–81.1 (m = 74.88, n = 8) (по материалам Moyá-Solá, 1987). По форме, размерам и пневматизации экз. ПИН, № 5644/107 очень похож на роговой стержень VM 82SS (см. Moyá-Solá, 1987, рис. 12 ; L – 155, DAP × DT – 41.2 × 33.4). По морфологии, размерам и пропорциям щечных зубов зоргелия из Тавриды также хорошо соответствует S minor. Строение Р3 и Р4 у экз. ПИН, № 5644/109, и VM-84-eIII E-9-23 идентично, но на образце из Тавриды гипоконид на Р4 лингвально менее округлен и слегка угловатый, что, возможно, говорит о большей архаичности. Индексы длины Р4 и ширины задней доли коронки Р4 у экз. ПИН, № 5644/109 попадают в диапазон этих индексов у S. minor из Вента Мицены, Wa/L – 57–61 (m = 57, n = 11) и Wp/Wa – 82–100 (m = = ?97.5, n = 11) (по материалам Moyá-Solá, 1987).
Рис. 4.
Соотношение параметров роговых стержней у разных видов Soergelia Европы и Кавказа (c использованием данных: Kahlke, 1969; Moyá-Solá, 1987; Kostopoulos, 1997; Bukhsianidze, 2005).
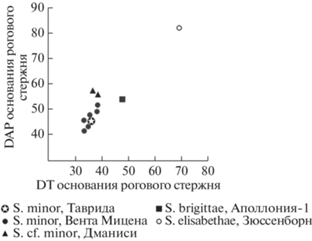
Рис. 5.
Сопоставление строения нижних премоляров Р3 и Р4 у разных видов Soergelia из местонахождений Европы и Кавказа: а – S. minor, экз. ПИН, № 5644/109, пещера Таврида; б, в – S. cf. minor, Дманиси: б – экз. D654, в – экз. D1733; г – S. intermedia, голотип TRL 88/68/1, Трлица; д – S. minor, экз. VM-84-eIII E‑9-23, Вента Мицена; e, ж –S. brigittae, Аполлония-1: е – экз. APL-25, ж – голотип APL-383; з – S. elisabethae, паратип D 377, Зюссенборн (г: по Crégut-Bonnoure, Dimitrijević, 2006, табл. III , фиг. 1a; з: по Schaub, 1951, табл. XII , фиг. 3).
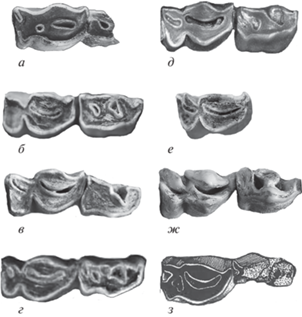
Таблица 1.
Сравнение промеров и индексов нижних зубных рядов видов рода Soergelia*
| Виды | Промеры (мм) и индексы (%) | ||||
|---|---|---|---|---|---|
| P2–P4 | M1–M3 | P2–M3 | P2–P4/P2–M3 | P2–P4/M1–M3 | |
| S. minor, экз. ПИН, № 5644/109, Таврида | 38.37 | 70.95 | 107.8 | 36 | 54 |
| S. minor, экз. VM-84-eIII E-9-23, Вента-Мицена | 43 | 74.5 | 116 | 37.07 | 57.72 |
| S. cf. minor, Дманиси | 49.99 | 80.10–84.42 | 130.28 | 38.37 | 62.41 |
| S. brigittae, Аполлония-1 | 50.3–54 | 92.6–102.5 | 143–156.8 | 34.9 | 53.65 |
| S. intermedia, голотип TRL 88/68/1, Трлица | 46 | 80.5 | 123.5 | 37 | 57 |
| S. elisabethae, паратип D 377, Зюссенборн | 45.8 | 101.6 | 148.8 | 31 | 45 |
Зоргелия из Тавриды проявляет значительное сходство с S. cf. minor из нижнего плейстоцена (1.77 млн л.н., MNQ18) Дманиси, Грузия, в т.ч. по типу и степени моляризации Р3 и Р4, но отличается от последней формы меньшими размерами, менее сжатым основанием рогового стержня и более коротким рядом премоляров (см. Vekua, 1995; Bukhsianidze, 2005). Р3 и Р4 на образцах из Дманиси менее стерты и хорошо видны такие примитивные черты, как: 1) сближенные на жевательной поверхности буккальная и третья лингвальная долинки, 2) скошенное положение энтоконида, и 3) косое положение четвертой долинки, открывавшейся на лингвальной стороне у задневнутреннего угла коронки.
Oт всех других видов зоргелия Тавриды отличается меньшими размерами и некоторыми особенностями зубной системы, в частности, более косым положением энтоконида на Р3 и Р4, а от S. elisabethae, кроме того, более длинной задней долей коронки Р4 и положением четвертой долинки на Р3 и Р4 у задневнутреннего угла, более длинным рядом премоляров, а также особенностями рогового стержня.
От S. brigittae Kostopoulos, 1997 из нижнего плейстоцена (~1 млн л.н, MNQ20) Аполлонии-1, Греция, отличается немного более длинным рядом нижних премоляров, замкнутостью третьей долинки на P4 и строением P3.
Роговой стержень S. brigittae представлен единственным образцом APL-310, небольшой частью лобной кости с основанием рогового стержня; его промеры – DAP × DT – 54 × 47.54; индекс сжатия – 88% (Kostopoulos, 1997, рис. 9G ). Голотип S. brigittae – APL-383, неполная нижняя челюсть с Р2–М3 – хранится в Ун-те Аристотеля в Салониках (Kostopoulos, 1997, рис. 9 С). Диагноз вида включает, в частности, открытую третью долинку и закрытый тригонид на Р4. На Р3 голотипа заднее крыло метаконида короткое, энтостилид и энтоконид длинные, расположены почти перпендикулярно переднезадней оси зуба, и третья долинка открывается на лингвальной стороне коронки.
От S. intermedia Cregut-Bonnoure et Dimitrijevic, 2006 из верхов нижнего–низов среднего плейстоцена Трлицы в Черногории отличается более узкими коронками нижнего зубного ряда, более скошенным назад энтоконидом на P3 и P4, менее моляризованным РЗ, менее вздутыми протоконидом и гипоконидом, а также присутствием небольшого эктостилида (зачаточного) на M1 (примитивные признаки). Кроме того, задняя доля на P4 длиннее и шире, чем у S. intermedia, индексы Lp/L – 22.82% и Wp/Wa – 81% (по данным Crégut-Bonnoure, Dimitrijević, 2006).
Голотип S. intermedia – правая нижнечелюстная кость TRL 88/68/1 – хранится в Ун-те Белграда. Роговые стержни в коллекции отсутствуют. По устному сообщению В. Димитриевич, голотип найден в слое 2 (Layer II) (конец раннего плейстоцена, MmQ3; Crégut-Bonnoure, Dimitrijević, 2006, табл. III , фиг. 1a–1c; MNQ20: Fernandez, Crégut, 2007). Этот слой 2 соответствует слою 5 более поздних работ (начало среднего плейстоцена, MmQ3, MIS 19; Agadzhanyan et al., 2017; Vislobokova et al., 2020).
От типового вида рода – S. elisabethae Schaub, 1951 из среднего плейстоцена Центральной Европы – зоргелия из Тавриды отличается менее массивными, более стройными и более уплощенными в основании роговыми стержнями, более длинным рядом премоляров и строением P4 (см. Schaub, 1951; Kahlke, 1969). На нижней челюсти D 377 S. elisabethae из Зюссенборна, хранящейся в Музее естественной истории в Базеле, коронка P4 короткая и широкая, со сплошной, почти плоской лингвальной стенкой и с сильновыпуклой буккальной стенкой. Задняя стенка энтоконида не скошена и почти перпендикулярна переднезадней оси коронки. На РЗ и Р4 четвертая долинка открывается у заднего края коронки. Задняя доля коронки Р4 сильно укорочена. Индекс длины задней доли Lp/L на Р4 у S. elisabethae вдвое меньше, чем у зоргелии из Тавриды. У паратипа D 377 он равен 14.42%, а у образца из Бад Франкенхаузена W-Frank. 1642 – 16.29% (Crégut-Bonnoure, Dimitrijević, 2006). Реконструкция тела нижнечелюстной кости паратипа D 377 (см. Schaub, 1951, табл. 12 , фиг. 1–3; Kahlke, 1969, рис. 4) искажает строение его передней части, которая, как и у зоргелии из Tавриды, по-видимому, была не очень высокой.
Таким образом, сравнительный анализ показывает, что остатки Soergelia из нижнего плейстоцена пещеры Таврида относятся к S. minor.
Следует отметить, что вид S. minor представлен в типовом местонахождении Вента Мицена-2 в Испании богатым материалом, хорошо обоснован морфологически, и его самостоятельность не вызывает сомнений. Некоторые исследователи предлагают относить к нему и виды S. brigittae и S. intermedia (напр., Martinez-Navarro et al., 2012). Однако эти виды отличаются от S. minor более крупными размерами и особенностями строения зубной системы. Их сведение в синонимику S. minor до получения новых ископаемых материалов и данных об изменчивости не может быть поддержано.
Материал. Правый роговой стержень с пеньком, экз. ПИН, № 5644/107; часть правой верхнечелюстной кости с М1–М3, экз. ПИН, № 5644/108; неполная правая ветвь нижнечелюстной кости с PЗ–M3, экз. ПИН, № 5644/109; Крым, пещера Таврида; нижний плейстоцен.
К ИСТОРИИ ЗОРГЕЛИЙ ЕВРОПЫ
В истории европейских Soergelia прослеживаются три этапа, соответствующих фаунистическим стадиям, выделяемым по крупным млекопитающим: 1) поздневиллафранкский с S. minor (~1.8–1.2 млн л.н.), 2) эпивиллафранкский с S. brigittae и S. intermedia (~1.2–0.8 млн. л.н.) и 3) среднеплейстоценовый (~0.8–0.4 млн л.н.) c S. elisabethaе.
Европейские виды зоргелий имеют разный эволюционный уровень и, по меньшей мере, два из них (S. minor и S elisabethae) пришли в Европу с дисперсионными волнами из Азии: первый – вскоре после палеомагнитного эпизода Олдувей (1.95–1.77 млн л.н.), а второй – в начале среднего плейстоцена, примерно на уровне или вскоре после инверсии Matuyama/Брюнес (0.781 млн л.н.).
Основные направления эволюции Soergelia связаны с усилением адаптаций к жизни в открытых пространствах и более прохладном климате на фоне глобального тренда понижения температуры в плейстоцене. Они отразились в увеличении размеров, развороте рогов в стороны и совершенствовании специализации к грейзингу (grazing), а именно: усилении гипсодонтности, удлинении моляров и сокращении относительной длины ряда премоляров (уменьшение индекса зубных сегментов), усилении моляризации премоляров. Среди ранних Soergelia (S. minor, S. brigittae и S. intermedia) строение рогов пока известно только для вида S. minor. Но все эти виды существенно отличаются от поздних форм меньшей гипсодонтностью, более высокими значениями индексов нижних зубных рядов и особенностями строения P3 и P4, в частности, более длинной задней долей коронки на P4 и положением четвертой долинки на лингвальной поверхности у задневнутреннего угла. К примитивным признакам в строении P4 у ранних Soergelia относятся: узкая коронка, длинная задняя доля коронки, слабовыпуклые метаконид и протоконид, косое положение энтоконида (его лингвальная стенка скошена назад), глубокие лингвальные долинки. При этом в строении зубной системы зоргелий конца раннего плейстоцена, с его нестабильными глобальными климатическими условиями, прослеживается мозаика примитивных и продвинутых признаков и их разное сочетание.
Первый этап, ~1.8–1.2 млн л.н. Зоргелии вида S. minor – самые примитивные и мелкие представители рода. Они имели небольшие, довольно стройные рога, уплощенные в основании и слабо сужающиеся к концу. Рога располагались над глазницами и были направлены вверх, отогнуты вперед, слабо гетеронимно скручены и лишь слегка развернуты в стороны. Древнейшая зоргелия S. cf. minor из Дманиси (1.77 млн л.н.) в Закавказье по морфологии зубной системы почти идентична S. minor из Вента Мицены, но она отличается немного более крупными размерами и такими примитивными признаками, как более уплощенное основание рогового стержня и более высокое значение относительной длины ряда премоляров (Bukhsianidze, 2005).
По большей уплощенности основания рогового стержня и индексу зубных сегментов S. minor из Tавриды более продвинута, чем S. cf. minor из Дманиси (1.77 млн л.н.), и занимает промежуточное положение между этой формой и эпивиллафранкскими S. brigittae и S. intermedia. По степени моляризации P4 и гипсодонтности моляров S. minor из Тавриды также примитивнее, чем S. brigittae и S. intermedia.
Кроме Вента Мицены-2 (1.3 млн л.н.) в Испании (Moyá-Solá, 1987) и пещеры Tаврида (1.8–1.5 млн л.н.) в Крыму, поздневиллафранкские Soergelia представлены в Италии (Монте Арджентарио) (Martinez-Navarro et al., 2012) и, возможно, в Венгрии (Villany 3) (Crégut-Bonnoure, 2002). В Монте Арджентарио найден фрагмент лобной кости с неполным рогом, определенный как Soergelia sp. cf. S. minor (Martinez-Navarro et al., 2012, рис. 2A). Роговой стержень крупнее, чем у S. minor; диаметры его основания DAP и DT (58.5 × 52.1) ближе к таковым у S. brigittae.
Второй этап, ~1.2–0.8 млн л.н. Ранние Soergelia представлены видами S. brigittae из Аполлонии-1 (~1 млн л.н) в Греции и S. intermedia из Трлицы (> 0.8 млн л.н.) в Черногории, известными лишь из их типовых местонахождений (Коstopoulos, 1987; Crégut-Bonnoure, Dimitrijević, 2006). Роговых стержней в Трлице нет, а в Аполлонии-1 найден фрагмент лобной кости с основанием рога. Рог мог быть слегка гетеронимно скручен (Коstopoulos, 1987). Эти виды отличаются от S. minor более крупными размерами и особенностями строения P3 и P4 (см. выше), в частности, менее скошенным положением энтоконида, а друг от друга – некоторыми особенностями специализации к грейзингу (разным сочетанием примитивных и продвинутых признаков в строении зубной системы). По уровню эволюционного развития они занимают промежуточное положение между самыми ранними и среднеплейстоценовыми формами, а по размерам близки к последним.
Кроме того, Soergelia этого этапа, по-видимому, присутствуют в пещере Козарника в Болгарии, Маасвлакте в Нидерландах (Kolfschoten, Vervoort-Kerkhoff, 1999) и пещере Валлонне (~1 млн л.н.) во Франции (Crégut-Bonnoure, 2007), а также отмечены в составе фауны Тетою-3 Румынии (Radulesco, Samson, 2001). В пещере Козарника в нижних слоях (биозона B2-2) найдено четыре зуба S. aff. intermedia, в т.ч. M1 (H14-H15, K/2264, 11b) с довольно высоким эктостилидом, высотой около 1 см (Fernandez, Crégut, 2007, табл. 1, фиг. 7 ). Нижний моляр M3 из Маасвлакта идентифицирован как S. minor (Kolfschoten, Vervoort-Kerkhoff, 1999, рис. 4) и по размерам соответствует этому виду. Но, судя по степени его гипсодонтности, он мог принадлежать более продвинутому виду ранних Soergelia. В Валлоне часть материала, отнесенная к Ammotragus europaeus, по мнению Э. Крегут-Боннур, принадлежит Soergelia (Crégut-Bonnoure, 2007), в частности M3 Val A8 AI19 654 с удлиненным букко-лингвально островком (см. Moullé et al., 2004, рис. 2).
Третий этап, ~0.8–0.4 млн л.н. Наиболее продвинутые формы зоргелий в Европе представлены видом S. elisabethaе – крупными формами, впервые описанными из среднего плейстоцена местонахождений Зюссенборн и Капелленберг в Германии (Schaub, 1951; Kahlke, 1969). Этот вид присутствует в Германии в местонахождении Бад Франкенхаузен, а также, возможно, во Франции (Сейн Сол Белкастел), Италии (Грималди), Румынии (Олтения) и др. (Radulesco, Samson, 1965; Kahlke, 1969; Koenigswald, Heinrich, 1999; Crégut-Bonnoure, 2002, 2005; Crégut-Bonnoure, Dimitrijević, 2006; Kahlke et al., 2011). У S. elisabethaе, в отличие от ранних Soergelia, роговые стержни были сильнее развернуты в стороны и, по-видимому, без гетеронимного скручивания; они массивные в основании и сильно сужающиеся к концу, иногда с заметной вздутостью в основании и средней части (Schaub, 1951; Kahlke, 1969; Bukhsianidze, 2005; Crégut-Bonnoure, 2005). S. elisabethaе хорошо отличается от ранних Soergelia и по более высокой степени гипсодоности и сильной моляризации P4 (см. выше), которые свидетельствуют о довольно высоком уровне специализации этого вида к грейзингу.
Экология и распространение. Все ранние Soergelia Западной Палеарктики были теплолюбивыми, с выраженными адаптациями к грейзингу и, вероятно, в разной степени способные к браузингу. В состав их кормов наряду с травянистыми растениями и травами могли входить побеги, листва и ветви древесных растений и кустарников. Эти зоргелии были, скорее всего, обитателями полуоткрытых и мозаичных ландшафтов (саванноподобных, лесостепных и степных) на склонах гор, в предгорьях и долинах. Все виды встречены в регионах, относящихся к Альпийскому горному поясу, на Пиренейском и Апеннинском полуостровах, Балканах, в Крыму и Закавказье. Рост разнообразия горных обитателей таких регионов в раннем плейстоцене мог быть связан как с разрывом ареалов при значительных глобальных климатических колебаниях и изменениях ландшафтно-климатической обстановки, так и с разновозрастными дисперсионными волнами.
Первые Soergelia, как и Нomo, пришли в восточные районы Западной Палеарктики вместе с дисперсионной волной примерно на уровне палеомагнитного эпизода Олдувей, и постепенно распространились к западу. На Балканах первое появление Soergelia отмечено в Трлице, уровень TRL11–10, 1.8–1.5 млн л.н., Donau/Eburonian, MIS 63–51 (Vislobokova et al., 2020), а на Пиренейском п-ове – позже, 1.5–1.3 млн л.н. В Европе широкое распространение ранних Soergelia связано с дисперсионной волной перед палеомагнитным эпизодом Харамильо, ~1.2 млн л.н. (MIS 36), а среднеплейстоценовых – с дисперсионной волной в начале эльстерского оледенения.
По сравнению с ранними Soergelia, вид S. elisabethae из среднего плейстоцена Европы был лучше адаптирован к прохладному и холодному климату и открытым пространствам (в частности, степным и, возможно, тундростепным). Эти крупные зоргелии (так называемые “степные козы”), по-видимому, вселились в Европу из Восточной Палеарктики с расширением открытых пространств во время эльстерского оледенения; они наиболее широко распространились в холодную стадию MIS 16 (Kahlke, 1995; Von Koenigswald, Heinrich, 1999; Kahlke et al., 2011; van der Made et al., 2017). В фауне Бад Франкенхаузена, которую относят к холодной стадии MIS 12 эльстера, поздние S. elisabethae сосуществовали с первыми шерстистыми носорогами Coelodonta tologoijensis, пришедшими из Азии (Kahlke et al., 2011). Вселение Сoelodonta в Центральную Европу свидетельствует о существовании перигляциальных условий и обширных открытых пространств (см. Kahlke, Lacombat, 2008; Kahlke et al., 2011). Поздние европейские S. elisabethae могли быть автохтонными формами, преадаптированными к обитанию в таких условиях. Но не исключено, что они, как и первые популяции S. elisabethae, проникли в Центральную Европу с дисперсионной волной из Восточной Палеарктики. Крупные формы зоргелий были распространены там с конца раннего плейстоцена. Их остатки найдены в Восточной Сибири (Шер, 1971; Sher, 1986; Боескоров, 2016, 2019) и на юге Западной Сибири (Кожамкулова, Мотузко, 1974; Алексеева, 1980; Васильев, 2005). Наиболее древние формы Soergelia sp. присутствуют в Якутии в раннеолерской фауне, а наиболее молодые найдены в позднеплейстоценовых местонахождениях Красный Яр и Тараданово на юге Западной Сибири в верховьях р. Оби (Васильев, Мартынович, 2007; Васильев, 2010, 2011). S. aff. elisabethae известны из позднеолерской фауны, коррелируемой с галерием Западной Европы (Sher, 1986). Эта зоргелия из Якутии, возможно, конспецифична S. elisabethae из Европы (Боескоров, 2019). Роговые стержни у нее в среднем были более уплощенные, чем у S. elisabethae из Европы (Боескоров, 2019), и это позволяет предполагать, что она была немного примитивнее и могла быть близка к базальным популяциям вида. В раннем и среднем плейстоцене сибирские Soergelia, по-видимому, были интразональными и могли существовать в нескольких ландшафтных зонах (от степных и лесостепных до лесотундровых и тундровых). Такие адаптации позволили им распространиться в Европу и Северную Америку при подходящих глобальных ландшафтно-климатических изменениях.
ЗАКЛЮЧЕНИЕ
Таким образом, исследование позволило дополнить фаунистический список из пещеры Таврида первым представителем трибы Ovibovini, найденным в нижнем плейстоцене Крыма. Исследование nоказало, что Soergelia из Тавриды принадлежала к одной из наиболее ранних популяций европейского вида S. minor, существовавшей в интервале примерно от 1.8 до 1.5 млн л.н. Эта находка расширяет представления о биохронологическом распространении и ареале S. minor, проливает свет на некоторые аспекты эволюции, экологии и распространения рода, а также свидетельствует о большом разнообразии раннеплейстоценовых парнопалых Крыма и фаунистических связях с Западной Европой и Закавказьем.
Список литературы
Алексеева Э.В. Млекопитающие плейстоцена юго-востока Западной Сибири. М.: Наука, 1980. 188 с.
Боескоров Г.Г. Новые данные о распространении и таксономии ископаемой зоргелии (Soergelia sp., Bovidae, Artiodactyla, Mammalia) на территории Якутии // Докл. Акад. наук. Общ. биол. 2016. Т. 469. № 4. С. 508–512.
Боескоров Г.Г. К распространению и таксономии ископаемой Soergelia sp. (Caprinae, Bovidae, Artiodactyla) на территории Якутии // Зоол. журн. 2019. Т. 98. № 10. С. 1148–1155.
Бондарев А.А., Голубев А.С. Новые находки Soergelia (Bovidae, Artiodactyla) на территории Среднего Прииртышья / Эволюция жизни на Земле. Матер. V Междунар. симп. Томск: Изд-во Дом Томского гос. ун-та, 2018. С. 199–201.
Васильев С.К. Зоргелия (Soergelia sp.) в Новосибирском Приобье // Фауны Урала и Западной Сибири в плейстоцене и голоцене. Челябинск: Изд-во Рифей, 2005. С. 81–88.
Васильев С.К. Остатки зоргелии (Soergelia sp.) в позднем плейстоцене Предалтайской равнины // Эволюция жизни на Земле. Матер. IV Междунар. симп. Томск: ТМЛ-Пресс, 2010. С. 537–541.
Васильев С.К. Некоторые итоги изучения Тарадановского аллювиального местонахождения позднеплейстоценовой мегафауны // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. 2011. Т. 17. С. 20–25.
Васильев С.К., Мартынович Н.В. Палеотериологические сборы в Тараданово в 2007 г. и новые находки остатков зоргелии Soergelia sp. // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. 2007. Т. 13. С. 23–28.
Кожамкулова Б.С., Мотузко А.Н. Зоргелия в Казахстанском Прииртышье // Вестн. АН Каз. ССР. 1974. № 7. С. 61–63.
Шер А.В. Млекопитающие и стратиграфия плейстоцена крайнего Северо-Востока СССР и Северной Америки. М.: Наука, 1971. 310 с.
Agadzhanyan A.K., Vislobokova I.A., Shunkov M.V., Ulyanov V.A. Pleistocene mammal fauna of the Trlica locality, Montenegro // Fossil Imprint. 2017. V. 73. № 1–2. P. 93–114.
Bukhsianidze M. The fossil Bovidae of Dmanisi. Doct. Thesis. Ferrara: Univ. Ferrara, 2005. 192 p.
Crégut-Bonnoure E. Les Ovibovini et Caprini (Mammalia, Artiodactyla, Bovidae, Caprinae) du Plio-Pléistocène dʼEurope: systématique, évolution et biochronologie. Thèse Doct. dʼEtat ès Sciences, Université de Lyon I, 2002. 386 p.
Crégut-Bonnoure E. Nouvelles données paléogéographiques et chronologiques sur les Caprinae (Mammalia, Bovidae) du Pléistocène moyen et supérieur d’Europe // Munibe Antropol. Arkeol. 2005. V. 57. P. 205–219.
Crégut-Bonnoure E. European Ovibovini, Ovini, Caprini (Caprinae, Mammalia) from the Plio-Pleistocene: new interpretations // Proc. 18th Intern. Senckenberg Conf., VI Intern. Paleontological Colloquium in Weimar. P. 139–158 (Cour. Forsch.-Inst. Senckenb. 2006. V. 256).
Crégut-Bonnoure E., Dimitrijević V. Megalovis balcanicus sp. nov. and Soergelia intermedia sp. nov. (Mammalia, Bovidae, Caprinae), new Ovibovini from the Early Pleistocene of Europe // Rev. Paléobiol. 2006. V. 25. № 2. P. 723–773.
De Giuli C., Masini F. A new element of the Late Villafranchian (Tasso Unit) faunas of Italy: occurrence of Ovibovini (Bovidae, Artiodactyla, Mammalia) in the fauna of Casa Frata (Upper Valdarno, Tuscany) // Boll. Soc. Paleontol. Ital. 1983. V. 22. P. 271–280.
Fernandez P., Crégut E. Les Caprinae (Rupicaprini, Ovibovini, Ovini et Caprini) de la séquence pléistocène de Kozarnika (Bulgarie du Nord): morphométrie, biochronologie et implications phylogéniques // Rev. Paléobiol. 2007. V. 26. № 2. P. 425–503.
Gentry A.W. An ovibovine (Mammalia, Bovidae) from the Neogene of Stratzing, Austria // Ann. Naturhist. Mus. Wien. 2001. Bd 102A. P. 189–199.
Harington C.R. Faunal exchanges between Siberia and North America: Evidence from Quaternary land mammal remains in Siberia, Alaska and Yukon Territory // Canad. J. Anthropol. 1980. V. 1. № 1. P. 45–49.
Harington C.R. Soergelia: an indicator of Holarctic Middle Pleistocene deposits? // Nat. Canad. J. Zool. 1987. V. 67. № 5. P. A1–A10.
Kahlke H.-D. Ovibos aus den Kiesen von Süssenborn. Ein Beitrag zur Systematik und Phylogenie der Ovibovini und zur Stratigraphie des Pleistozäns // Geologie. 1963. Bd 12. H. 8. S. 942–972.
Kahlke H.-D. Die Soergelia–Reste aus den Kiesen von Süssenborn bei Weimar // Paläontol. Abh. Apt. A. 1969. Bd 3. H. 3/4. S. 531–544.
Kahlke R.D., García N., Kostopoulos D.S. et al. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe // Quatern. Sci. Rev. 2011. V. 30. P. 1368–1395.
Kahlke R.-D., Lacombat F. The earliest immigration of woolly rhinoceros (Coelodonta tologoijensis, Rhinocerotidae, Mammalia) into Europe and its adaptive evolution in Palaearctic cold stage mammal faunas // Quatern. Sci. Rev. 2008. V. 27. P. 1951–1961.
Kostopoulos D.S. The Plio-Pleistocene Artiodactyls (Vertebrata, Mammalia) of Macedonia. 1. The fossiliferous site “Apollonia–1”, Mygdonia basin of Greece // Geodiversitas. 1997. V. 19. № 4. P.845–875.
Martinez-Navarro B., Sardella R., Rook L. et al. First occurrence of Soergelia (Ovibovini, Bovidae, Mammalia) in the Early Pleistocene of Italy // Quatern. Intern. 2012. V. 267. P. 98–102.
Moullé P.-E., Echassoux A., Martinez-Navarro B. Ammotragus europaeus: une nouvelle espèce de Caprini (Bovidae, Mammalia) du Pléistocène inférieur à la grotte du Vallonnet (France) // C. R. Palevol. 2004. V. 3. № 8. P. 663–673.
Moyá-Solá S. Los bovidos (Artiodactyla, Mammalia) del yacimiento del Pleistoceno inferior de Venta Micena (Orce, Granada, Espaňa) // Paleontol. Evol. 1987. № 1. P. 181–236.
Radulesco C., Samson P. Soergelia elisabethae Schaub dans le Pleistocene moyen de l’Oltenie (Roumanie) // Eclog. Geol. Helv. 1965. V. 58. № 2. P. 1107–1110.
Radulesco C., Samson P. Biochronology and evolution of the Early Pliocene to the Early Pleistocene mammalian faunas of Roumania // Boll. Soc. Paleontol. Ital. 2001. V. 40. P. 285–291.
Schaub S. Soergelia n. gen., ein Caprine aus dem thuringischen Altpleistocaen // Eclog. Geol. Helv. 1951. V. 44. № 2. S. 375–381.
Sher A.V. Olyorian land mammal age of Northeastern Siberia // Palaeontogr. Ital. 1986. V. 74. P. 97–112.
Van der Made J., Rosell J., Blasco R. Faunas from Atapuerca at the Early-Middle Pleistocene limit: the ungulates from level TD8 in the context of climatic change // Quatern. Intern. 2017. V. 433A. P. 296–346.
Van Kolfschoten T., Vervoort-Kerkhoff Y. The Pleistocene and Holocene mammalian assemblages from the Maasvlakte near Rotterdam (the Netherlands): with special reference to the Ovibovini Soergelia minor and Praeovibos cf. priscus // Deinsea. 1999. V. 7. P. 369–381.
Vekua A. Die Wirbeltierfauna des Villafranchian von Dmanisi und ihre biostratigraphische Bedeuting // Jb. Rom.-German. Zentralmus. Mainz. 1995. Bd 42. S. 77–180.
Vislobokova I.A., Agadzhanyan A.K., Lopatin A.V. The case of Trlica TRL11–10 (Montenegro): Implications for possible early hominin dispersals into the Balkans in the middle of the Early Pleistocene // Quatern. Intern. 2020. V. 554. P. 15–35.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал