

Палеонтологический журнал, 2022, № 2, стр. 21-32
Фораминиферы семейства Elphidiidae кайнозоя Дальнего Востока: палеонтология, детальная стратиграфия и палеобиогеография
С. И. Бордунов a, b, *, Д. М. Ольшанецкий a, **
a Геологический институт РАН
119017 Москва, Россия
b Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: sib-msu@mail.ru
** E-mail: o.mitia@gmail.com
Поступила в редакцию 08.08.2021
После доработки 27.09.2021
Принята к публикации 28.09.2021
- EDN: OIJLRO
- DOI: 10.31857/S0031031X22020052
Аннотация
Представители фораминифер семейства Elphidiidae Galloway, 1933 широко распространены в кайнозойских отложениях Дальнего Востока, в которых сосредоточены основные месторождения нефти и газа региона. Виды данного семейства являются важным стратиграфическим и коррелятивным репером и используются в качестве видов-индексов для местных и региональных биостратиграфических зон, а также для слоев с фауной. Приведено систематическое описание характерных видов.
ВВЕДЕНИЕ
Представители фораминифер семейства Elphidiidae Galloway, 1933 широко распространены в кайнозойских отложениях Дальнего Востока (Asano, 1958; Волошинова и др., 1970; Серова, 1978, 2001; Серова, Спирина, 1984; Тузов и др., 1997; Фрегатова, Дмитриева, 2004; Дмитриева, 2007; Бордунов, 2013, 2015; Жаров и др., 2013; Фрегатова, Беньямовский, 2013; Ольшанецкий, 2018а и др.). Виды данного семейства бентосных фораминифер практически повсеместно распространены и в современных шельфовых бассейнах Арктики и Северной Пацифики (Cushman, Grant, 1927; Cushman, 1936; Щедрина, 1953; Гудина, 1966, 1969; Саидова, 1975; Фурсенко и др., 1979; Хорева, 1988; Lukina, 2001 и др.). Изучение кайнозойских бассейнов Дальнего Востока представляет большой научный и практический интерес, так как именно в осадочном чехле кайнозойского возраста сосредоточены основные месторождения нефти и газа региона.
Широкому расселению и развитию эльфидиид способствовал ряд факторов. В первую очередь, это цикличное развитие дальневосточных бассейнов в кайнозое, когда этапы трансгрессий сменялись регрессиями, в результате которых бассейны испытывали обмеление. Глубина компенсации растворения кальцита (лизоклин), возможно, увеличилась в связи с некоторым общим потеплением водных масс, что благоприятно сказалось на сохранении в осадке известковых раковин. Во-вторых, этому также способствовал относительно теплый климат палеоцена и эоцена. Фораминиферы с известковой раковиной вновь стали важными компонентами бентосной фауны того времени, расселившись в широком ареале в палеоген–неогеновых морях. Климатические оптимумы начала эоцена, конца раннего–начала среднего миоцена вызывали миграцию теплолюбивой фауны в более северные, сравнительно холодноводные бассейны (Гладенков, Синельникова, 1990; Гладенков, Шанцер, 1993; Савельева, 2006). В-третьих, относительно толстая известковая раковина видов этого семейства могла долго сохраняться в осадках, не разрушаясь в процессе литификации и диагенеза осадков. Большое значение в геологической истории имели и тектонические движения, которые приводили к глубоким перестройкам структурного плана региона. Открывались и закрывались проливы, нарушалась и возобновлялась связь с океаном, что оказывало непосредственное влияние на пути миграции и состав планктонных и бентосных сообществ фауны, в т.ч. и фораминифер (Gladenkov et al., 2002; Гладенков, Гладенков, 2004; Плетнев, 2007; Гладенков, 2015). И именно бентосная фауна, к которой относятся и эльфидииды, наиболее быстро реагировала на подобные изменения палеообстановок (Гладенков, 2015).
Представители эльфидиид относительно быстро эволюционировали и поэтому имеют достаточно высокое значение для разработки зональных шкал региона. Виды данного семейства являются важным стратиграфическим и коррелятивным репером и часто используются в качестве видов-индексов для местных, региональных и субглобальных стратиграфических зон, а также для слоев с фауной (Серова, Спирина, 1984; Дмитриева, 2007; Фрегатова, Беньямовский, 2013; Бордунов, 2015; Бордунов и др., 2018; Ольшанецкий, 2018б).
Морфология раковин фораминифер семейства Elphidiidae и их систематическое положение хорошо и детально изучены отечественными (Волошинова, Дайн, 1952; Волошинова, 1958; Крашенинников, 1960; Волошинова, Кузнецова, 1964; Волошинова и др., 1970; Гудина, Левчук, 1983 и др.) и зарубежными микропалеонтологами (Cushman, 1930, 1936, 1939; Galloway, 1933; Asano, 1937, 1958; Wade, 1957; Buzas et al., 1985; Hayward et al., 1997). Поэтому основное внимание в настоящей статье уделено важной биостратиграфической роли данного семейства в построении местных и региональных стратиграфических шкал Дальневосточного региона.
МАТЕРИАЛ И МЕТОДИКА
Материал для исследований раковин фораминифер семейства Elphidiidae получен в результате изучения разрезов кайнозоя Западной и Восточной Камчатки, Сахалина и скважин на шельфе Охотского моря. При их изучении применен комплекс как традиционных, так и новых методов изучения морфологии раковин. Выявление особенностей строения раковин этого семейства составляло одну из задач проводимых исследований. Для изучения морфологического строения был использован метод фотографирования раковин фораминифер в сканирующем микроскопе FEI Tecnai G2 200k V Transmission Electron Microscope (НИЦ Курчатовского ин-та). Применение микроскопа данного типа позволяет осуществлять съемку раковин без напыления их поверхности золотом или графитом. Это позволило избежать артефактов, довольно часто появляющихся при напылении образцов золотом и придающих изображениям характерный вид снимков на СЭМ. Также важно, что ненапыленные образцы остаются пригодными для дальнейшего изучения и съемки под оптическим микроскопом. Изображения одних и тех же экземпляров, сделанные с разных ракурсов при помощи оптического микроскопа и СЭМ, помогают выявлять наибольшее количество диагностических признаков и значительно повышают узнаваемость видов фораминифер, изображений которых на сегодняшний день вообще недостаточно. В результате применения данного метода был получен важный материал для установления родовой и видовой диагностики представителей изучаемого семейства бентосных фораминифер. При описании видов использована система высших таксонов фораминифер, предложенная A. Лебликом и Х. Таппан (Loeblich, Tappan, 1987) с уточнением по “Всемирной базе данных по современным фораминиферам”, представленной на сайте www.marinespecies.org/foraminifera (Gross, 2001; Hayward, Gross, 2021). Экземпляры, изображенные на табл. IV , хранятся в коллекции палеонтологических образцов Геологического ин-та РАН, Москва.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Фораминиферы семейства Elphidiidae широко распространены в отложениях кайнозоя Дальнего Востока и давно используются при стратиграфическом расчленении и датировании разрезов этих отложений. Смена бентосных сообществ соответствует, с одной стороны, эволюционному развитию фораминифер, а с другой − изменению палеообстановок (глубины бассейна, температуры воды, подводных течений водных масс, колебаний уровня моря). Отмечаемые перестройки состава и структуры зональных сообществ находят отражение в палеобиогеографической провинциальности данных сообществ, что помогает в проведении более обоснованных корреляций кайнозойских отложений по бентосным фораминиферам.
Анализ ареалов распространения зональных видов имеет важное значение в стратиграфической корреляции. Например, изучение ареалов крупных представителей семейства Elphidiidae, таких как Pseudoelphidiella subcarinata (Voloshinova, 1958) (табл. IV , фиг. 1, 2 ) и Elphidiella oregonensis (Cushman et Grant, 1927) (табл. IV , фиг. 3, 4 ), которые используются в качестве видов-индексов для местных и региональных стратиграфических зон, а также для слоев с фауной (Тузов и др., 1997; Фрегатова, Дмитриева, 2004; Дмитриева, 2007; Бордунов, 2015), показало их широкое распространение в Северной Пацифике в достаточно узком стратиграфическом интервале, что и определило их биостратиграфическое значение. Данные виды маркируют продвижение относительно тепловодных и мелководных сообществ к северу на фоне региональных регрессий, что обусловило миграцию и расселение этих видов вдоль шельфов Северной Пацифики (рис. 1).
Рис. 1.
Изученные разрезы кайнозойских отложений и ареалы распространения: a – Psedoelphidiella subcarinata; б – Elphidiella oregonensis. Разрезы: 1 – Северо-Восточный Сахалин; 2 – Пограничный прогиб Сахалина; 3 – Южный Сахалин; 4 – Северная Япония; 5 – скв. Магаданская в Охотском море; 6 – Точилинский разрез Западной Камчатки; 7 – Колпаковский прогиб Западной Камчатки; 8 – о. Карагинский; 9 – Хатырский прогиб Корякского нагорья; 10 – Анадырский прогиб Корякского нагорья; 11 – скв. Центральная; 12 – скв. Нортон; 13 – залив Аляска; 14 – залив Тоффино.
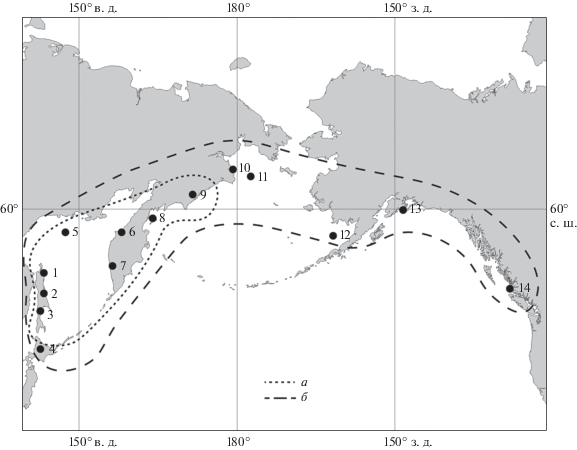
Анализ палеобиогеографической провинциальности зональных сообществ показал, что в течение кайнозоя в сообществах бентосных фораминифер Дальнего Востока доминировали бореальные виды с включением космополитных и южнобореальных видов. Изменения в составе сообществ связаны с прогрессирующим похолоданием на протяжении кайнозоя, а также рядом хорошо выраженных периодов потепления (климатическими оптимумами) во время формирования отложений на границе палеоцена и нижнего эоцена, верхнего эоцена, пограничных отложений нижнего и среднего миоцена, нижнего плиоцена (Гладенков, Шанцер, 1993; Плетнев, 2007; Гладенков, 2015).
На основе смены сообществ бентосных фораминифер в разрезе кайнозоя региона выделяются стратоны, которые по палеонтологическому и стратиграфическому критериям соответствуют биостратиграфическим комплексным зонам, а по масштабу их распространения – региональным или местным зонам.
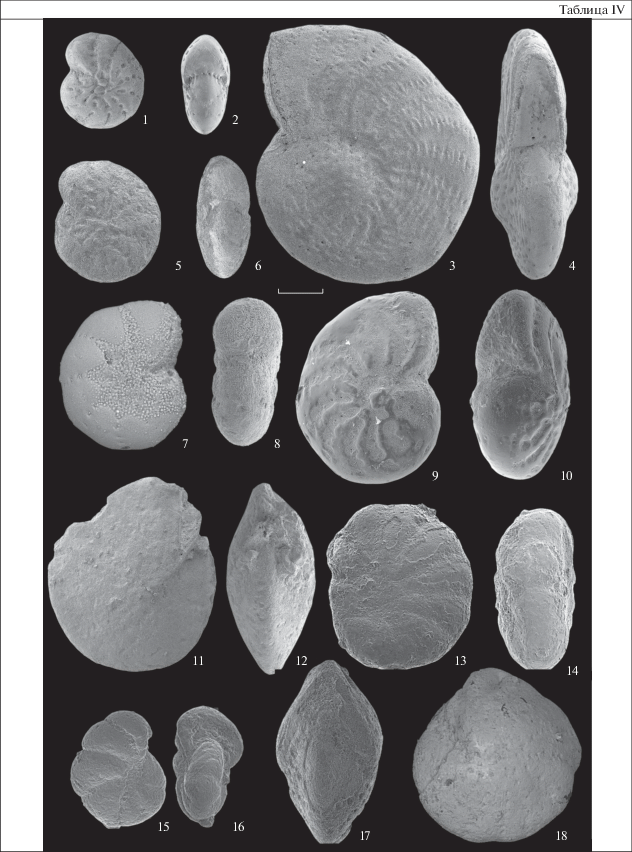
Объяснение к таблице IV
Фиг. 1, 2 . Elphidium clavatum Cushman, 1930, экз. ГИН, № 5312/15951: 1 – вид сбоку, 2 – вид со стороны устья; Восточная Камчатка; плиоцен–плейстоцен, ольховская свита.
Фиг. 3, 4 . Elphidiella oregonensis (Cushman et Grant, 1927), экз. ГИН, № 5318/42113: 3 – вид сбоку, 4 – вид со стороны устья; Восточная Камчатка; плиоцен–плейстоцен, ольховская свита.
Фиг. 5, 6 . Elphidium hughesi (Cushman et Grant, 1927), экз. ГИН, № 5318/4202: 5 – вид сбоку, 6 – вид со стороны устья; Восточная Камчатка; плиоцен–плейстоцен, ольховская свита.
Фиг. 7, 8 . Cribroelphidium vulgare (Voloshinova, 1952), экз. ГИН, № 5289/1102: 7 – вид сбоку, 8 – вид со стороны устья; Восточная Камчатка; верхний миоцен, юнюньваямская свита.
Фиг. 9, 10 . Elphidiella simplex Voloshinova, 1958, экз. ГИН, № 5289/1121: 9 – вид сбоку, 10 – вид со стороны устья; Восточная Камчатка; нижний плиоцен; лимитеваямская свита.
Фиг.11, 12 . Pseudoelphidiella subcarinata (Voloshinova, 1958), экз. ГИН, № om08/1544: 11– вид сбоку, 12 – вид со стороны устья); Западная Камчатка; нижний миоцен, кулувенская свита.
Фиг. 13, 14 . Cribroelphidium crassum (V. Kuznetsova, 1970), экз. ГИН, №om08/1513: 13 – вид сбоку, 14 – вид со стороны устья; Западная Камчатка; нижний миоцен, кулувенская свита.
Фиг. 15, 16 . Elphidium sumitomoi Asano et Murata, 1948, экз. ГИН, № om08/1519: 15 – вид сбоку, 16 – вид со стороны устья; Западная Камчатка; верхний эоцен – олигоцен; аманинская свита.
Фиг. 17, 18 . Elphidium californicum Cook, 1959, экз. ГИН, № om08/1503: 17 – вид сбоку, 18 – вид со стороны устья; Западная Камчатка; верхний эоцен – олигоцен, аманинская свита.
Длина мерной линейки 100 мкм.
В нижних горизонтах осадочного чехла кайнозойского возраста (палеоцен?–средний эоцен) на Дальнем Востоке выделяют комплекс с фораминиферами Elphidium californicum Cook, 1950 (Серова, Спирина, 1984; Жаров и др., 2013, Дмитриева и др., 2015). Этот вид широко распространен на Камчатке (табл. IV , фиг. 17, 18 ). В верхнем эоцене–нижнем олигоцене важное стратиграфическое значение имеют находки Cribroelphidium sumitomoi (Asano et Murata, 1958) (Серова, 2001; Жаров и др., 2013) (табл. IV , фиг. 15, 16 ). В нижнем миоцене региона выделяется региональная зона по фораминиферам Pseudoelphidiella subcarinata (Дмитриева, 2007; Бордунов, 2015; Бордунов и др., 2018). Данная зона прослеживается во всех регионах Охотоморского, включая Камчатку (табл. IV , фиг. 11, 12 ), Беринговоморского и Япономорского бассейнов. Высокий стратиграфический и коррелятивный потенциал этого вида обусловлен его частой встречаемостью, хорошей сохранностью, легко отличимой морфологией. В нижнем миоцене также выделяют слои с фораминиферами с индексом из другого рода – Cribroelphidium crassum V. Kuznetsova, 1970, хорошо распознаваемым видом с рядом характерных внешних признаков (Волошинова и др., 1970; Большаков и др., 1989), которые нами показаны на примере экземпляра с Камчатки (табл. IV , фиг. 13, 14 ). С видами-индексами из этого семейства М.Я. Серовой (1978) выделены слои с Pseudoelphidiella hannai (Cushman et Grant, 1927) на Западной Камчатке и слои с Elphidiella tumiensis V. Kuznetsova, 1970 на Северном Сахалине.
В среднем миоцене Берингова моря выделяют местные стратиграфические подразделения в ранге слоев с фауной и местных зон с видом-индексом Pseudoelphidiella problematica (Voloshinova, 1958), имеющим некоторое сходство с P. subcarinata, но отличимым по размерам раковины, строению межсептальных мостиков, характеру устьевых борозд (Волошинова и др., 1970). В Точилинском опорном разрезе Западной Камчатки выделены слои с Cribroelphidium micrum Voloshinova, 1958 (Серова, 1978).
В верхнем миоцене Камчатки выделяют слои с Elphidiella nutovoensis (Borovleva, 1952) (Серова, 1978), слои с Elphidiella simplex Voloshinova, 1958 (Дмитриева, 2007), слои с Elphidiella nabilensis (Voloshinova et Borovleva, 1952) (Бордунов, 2013). В комплексах бентосных фораминифер региона часто присутствует Cribroelphidium vulgare (Voloshinova, 1952) (табл. IV , фиг. 7, 8 ), принятый в качестве индекса зоны в верхнем миоцене Восточной Камчатки (Бордунов, 2015). Вид имеет более крупную раковину с более грубой скульптурой, чем у его вероятного потомка C. bartletti (Cushman, 1933), широко распространенного в регионе в плиоцен–четвертичное время.
В плиоцене Сахалина, Западной и Восточной Камчатки, Берингова моря выделяют слои с фораминиферами, местную, региональную зону с индексом Elphidiella oregonensis (табл. IV , фиг. 3, 4 ). Вид имеет крупную для рода раковину, уплощенную с боков, c крупными выпуклыми шишками в пупочной области, что позволяет легко его диагностировать (Волошинова и др., 1970; Бордунов, 2015). Этот характернейший вид нижнего плиоцена Дальневосточного региона встречается в мелководно-морских отложениях внутреннего шельфа. Другим характерным видом шельфовых сообществ плиоцена является E. hughesi (Cushman et Grant, 1927), используемый в региональной биостратиграфии Северной Пацифики (табл. IV , фиг. 5, 6 ) (Бордунов, 2013).
В позднем плиоцене появляется вид Elphidium clavatum Cushman (табл. IV , фиг. 1, 2 ), который широко распространен в четвертичное и современное время в арктических и дальневосточных морях. Это относительно мелководный шельфовый эвригалинный вид, который используется в качестве индекс-вида для выделения комплексов бентосных фораминифер, зональных подразделений в ранге местных зон плейстоценовых отложений Арктики и Дальнего Востока (Бордунов, 2015). К тому же, этот оппортунистический вид, свидетельствующий об усилении влияния пресноводного стока, сопровождавшегося поступлением большого количества терригенного материала и органики (Овсепян и др., 2015), является хорошим палеоэкологическим репером при изучении кайнозойских отложений Дальнего Востока.
ЗАКЛЮЧЕНИЕ
Таким образом, анализ стратиграфического распространения представителей семейства Elphidiidae в разрезах кайнозойских отложений Дальнего Востока показал, что они имеют важное биостратиграфическое значение и широко используются при детальном стратиграфическом расчленении и региональной корреляции, что позволяет выделять на их основе дробные стратиграфические подразделения в ранге местных и региональных зон или слоев с фауной. Смена бентосных сообществ соответствует, с одной стороны, эволюционному развитию фораминифер, а с другой − изменению палеообстановок (глубины бассейна, температуры воды, подводных течений, колебаний уровня моря). Отмечаемые перестройки состава и структуры зональных сообществ находят отражение в палеобиогеографической провинциальности данных сообществ, что помогает в проведении более обоснованных корреляций кайнозойских отложений. Описанные виды маркируют продвижение относительно тепловодных и мелководных сообществ к северу на фоне региональных регрессий, что обусловило их миграцию и расселение вдоль шельфов Северной Пацифики.
Работа выполнена по теме госзадания ГИН РАН № 0114-2021-0003 и при поддержке проекта РФФИ № 19-05-00361.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
СЕМЕЙСТВО ELPHIDIIDAE GALLOWAY, 1933
ПОДСЕМЕЙСТВО ELPHIDIINAE GALLOWAY, 1933
Род Elphidium Montfort, 1808
Elphidium clavatum (Cushman, 1930)
Табл. IV , фиг. 1, 2
Elphidium incertum var. clavatum: Cushman, 1930, с. 20, табл. 7 , фиг. 10a, 10b ; 1939, с. 57, табл. 16 , фиг. 1, 2 .
Elphidium incertum: Phleger, 1952, с. 83, табл. 14 , фиг. 7 .
Elphidium clavatum: Loeblich, Tappan, 1953, с. 98, табл. 19 , фиг. 8–10 ; Lei, Li, 2016, с. 359, рис. 83 .
Retroelphidium clavatum: Волошинова и др., 1970, с. 157, табл. XXXIX , фиг. 9–13 ; Бордунов, 2015, табл. XII , фиг. 6–8, 11 .
Retroelphidium subclavatum: Гудина, 1966, с. 45, табл. IV , фиг. 4–10 , табл. IX , фиг. 3 , табл. Х, фиг. 3 ; Фурсенко и др., 1979, c. 166, табл. 16 , фиг. 5 , табл. 43 , фиг. 1, табл. 42 , фиг. 5, 6 ; Хорева, 1988, с. 61, табл. VIII , фиг. 2 .
Cribroelphidium excavatum clavatum: Narayan et al., 2005, с. 128, табл. 3 , фиг. 25, 26 .
Elphidium excavatum subsp. clavatum: Hayward, Gross, 2021, http://marinespecies.org/aphia.php?p=taxdetails&id=466569.
Голотип – Cushman Collection, № 10400; Северная Атлантика, залив Мэн; современный.
Описание. Раковина планиспиральная, округлая, контур округлый, ровный, иногда немного волнистый в конце последнего оборота. Со стороны устья раковина с прямыми боковыми сторонами со скругленными краями. Септальные швы слегка изогнутые, плоские, обычно заглубляются к периферическому краю. В последнем обороте насчитывается от восьми до 12 камер. Размеры камер постепенно увеличиваются по мере роста раковины. Пупочная область обычно прикрыта карбонатной шишкой различной формы, вокруг которой располагаются пустулы. Устье находится в основании последней камеры. Форамен представляет собой серию отверстий, расположенных в основании септальной поверхности.
Размеры в мкм. Диаметр 150–300, толщина 100–150.
Сравнение. От близкого вида E. incertum отличается в первую очередь крупной монолитной пупочной шишкой и менее выраженными септальными мостиками.
Замечания. Для E. clavatum характерна широкая изменчивость, форма раковины бывает разной степени выпуклости, периферический край – от закругленного до суженного. Пупочная шишка может быть выражена в разной степени, состоять из одной или нескольких частей.
Распространение. Вид широко распространен в плиоцене–плейстоцене Северной Пацифики, Северной Атлантики, а также субглобально в современных морях на глубинах от 25 до 200 м.
Материал. 120 экз.; Россия, Восточная Камчатка, п-ов Камчатский мыс; плиоцен–плейстоцен, ольховская свита.
Elphidium hughesi Cushman et Grant, 1927
Табл. IV , фиг. 5, 6
Elphidium hughesi: Cushman, Grant, 1927, с. 75, табл. 7 , фиг. 1; Cushman et al., 1930, c. 61, табл. 3 , фиг. 15 ; Cushman, 1939, с. 49, табл. 13 , фиг. 7, 8 ; Волошинова и др., 1970, c. 156, табл. XXXIX , фиг. 1–3 .
Elphidium hughesi hughesi: Rau, 1970, c. 24, табл. 1 , фиг. 11 .
Elphidiella hughesi: Osterman, 1996, табл. 1 , фиг. 6 .
Retroelphidium hughesi: Хорева, 1988, с. 62, табл. VIII , фиг. 3 ; Бордунов, 2015, табл. XII , фиг. 9, 10 .
Голотип – Invert. Coll. Lel and Stanford Junior Univ., № 412; Калифорния; нижний плиоцен.
Описание. Раковина округлая, с волнистым контуром. Периферический край закругленный. В последнем обороте 10–12 камер, размеры которых увеличиваются по мере роста раковины. Швы изогнутые, хорошо заметные, с отчетливыми септальными мостиками, чередующиеся с септальными отверстиями округлой формы. Пупочная область плоская, прикрытая карбонатной шишкой, часто разделенной на несколько частей. Устье дырчатое в основании септальной поверхности.
Размеры в мкм. Диаметр 200–350, толщина 100–150.
Сравнение. Вид близок к E. clavatum, от которого отличается в первую очередь более крупными и отчетливыми септальными мостиками, находящимися в большем числе на каждой стороне камеры, а также более крупными размерами.
Замечания. Для E. hughesi характерна сильная изменчивость, контур раковины может быть округлым или волнистым, сильно меняется форма и размер пупочной шишки и форма периферического края.
Распространение. Плиоцен–плейстоцен Сахалина (маруямская, верхненутовская, помырская свиты), Камчатки (лимимтеваямская, усть-лимимтеваямская, ольховская свиты), Арктики (скважина 910 ODP), Японии (формация Сетана) и Калифорнии.
Материал. 54 экз.; Россия, Восточная Камчатка, п-ов Камчатский мыс; плиоцен–плейстоцен, ольховская свита.
Elphidium californicum Cook, 1959
Табл. IV , фиг. 17, 18
Elphidium californicum: Cook in Mallory, 1959, с. 184, табл. 15 , фиг. 10a, 10b .
Голотип – UC A-5360-8, № 41,589; Калифорния; верхний эоцен.
Описание. Раковина большая, планиспиральная, двояковыпуклая, билатерально-симметричная. Периферический край заостренный, но без киля. Наибольшая толщина раковины приходится на пупок. В наружном обороте насчитывается до 18–20 камер, постепенно увеличивающихся в размере по мере нарастания. Швы совсем немного вдавлены и чуть S-образно изогнуты, вперед по линии навивания у пупочной области выступает меньшая дуга и назад ближе к центру камер – большая. Септальные мостики короткие, не приподнятые и слабо различимые. Стенка гладкая. Пупок закрыт пупочной шишкой, иногда сливающейся со стенкой раковины. Устье состоит из цепочки пор в основании устьевой поверхности.
Размеры в мкм. Диаметр 220–400, толщина 100–200.
Сравнение. Отличается от E. charlottensis более широкой дисковидной формой с приподнятым и в большей степени выраженным пупком. Септальные отверствия выражены слабее.
Замечания. Раковины могут сильно отличаться по степени развития пупочной шишки и ее выраженности на раковине. Вид (табл. IV , фиг. 17, 18 ) имеет большее сходство с гипотипом, чем с голотипом, выделенным автором вида.
Распространение. Средний–верхний эоцен Сахалина (люкаминская свита), Западной Камчатки (снатольская, ковачинская, аманинская свиты) и верхний эоцен Северной Америки.
Материал. Четыре экз.; Россия, Западная Камчатка, бухта Квачина; верхний эоцен, аманинская свита.
Род Cribroelphidium Cushman et Brönnimann, 1948
Cribroelphidium sumitomoi (Asano et Murata, 1958)
Табл. IV , фиг. 15, 16
Elphidium sumitomoi: Asano, 1958, с. 51, табл. 8 , фиг. 20a, 20b .
Cribroelphidium sumitomoi: Волошинова и др., 1970, c. 165, табл. XLIII , фиг. 1a, 1б; Серова, Спирина, 1984, c. 127, табл. XVII , фиг. 7, 8 .
Голотип – IGPS coll. Cat., № 77177; Япония; олигоцен, формация Кисима.
Описание. Раковина округлая, периферический край в целом закругленный, контур слегка волнистый. Пупочная область вдавленная, обычно не прикрытая пупочным бугорком. Камеры немногочисленные, немного вздутые; в среднем семь или восемь у взрослых форм. Швы углубленные, септальные ямки и мостики на швах отчетливые. Устье состоит из округлых пор в основании устьевой поверхности.
Размеры в мкм. Диаметр 220–450, толщина 140–350.
Сравнение. C. sumitomoi ближе всего по морфологии к С. subglobosum, от которого отличается в первую очередь закругленным периферическим краем и меньшим количеством камер, а также менее выраженной пупочной областью.
Замечания. Характерны четкие границы между камерами за счет их выпуклости; межсептальные швы тонкие, отсутствуют пупочные наросты.
Распространение. Верхний эоцен–нижний миоцен Японии, олигоцен–нижний миоцен Сахалина (мачигарская, холмская свиты); эоцен–олигоцен Камчатки (снатольская, ковачинская, аманинская свиты).
Материал. Шесть экз.; Россия, Западная Камчатка, бухта Квачина; верхний эоцен, аманинская свита.
Cribroelphidium vulgare (Voloshinova, 1952)
Табл. IV , фиг. 7, 8
Elphidium vulgare: Волошинова, Дайн, 1952, с. 53, табл. VIII , фиг. 4, 6, 7 .
Cribroelphidium vulgare: Волошинова, 1958, с. 174, табл. VII , фиг. 2–10 ; Волошинова и др., 1970, с. 169, табл. XLII , фиг. 5, 6 ; Серова, 1978, с. 115, табл. VIII , фиг. 6 а, 6в; Серова, Спирина, 1984, с. 126, табл. XVII , фиг. 5 а, 5б; Бордунов, 2015, табл. XIII , фиг. 5, 6 .
Голотип – ВНИГРИ, № 442/97; Сахалин, Катангли; верхний миоцен, окобыкайская свита.
Описание. Раковина от средних до крупных размеров для рода, с прямыми боковыми сторонами, иногда чуть лопастными. Контур округлый, ровный, периферический край округлый. В наружном обороте восемь–десять камер, слегка выпуклых, очень равномерно и незначительно увеличивающихся в размерах по мере роста. Швы между камерами углубленные, слабоизогнутые или, чаще, прямые; у периферического края плоские, поверхностные; в пупочной области швы становятся более широкими, образуя звездчатую форму в центре раковины. Пупок достаточно широкий, слегка углубленный, многолучевой. Устье и форамен типичные для рода. Стенка гладкая, матовая, фарфоровидная.
Размеры в мкм. Диаметр 250–450, толщина 120–250.
Сравнение. От близкого вида C. bartletti (Cushman, 1933) отличается менее вздутой формой раковины, более широкой пупочной областью, более крупными размерами.
Замечания. Изменчивость выражается в увеличении числа камер в наружном обороте (от четырех до пяти–восьми), расширении пупочной области, увеличении степени вздутости в процессе роста.
Распространение. Вид имеет широкое распространение в средне–верхнемиоценовых отложениях Сахалина – в верхах дагинской, в окобыкайской и нутовской свитах Восточного побережья острова, в сертунайской и александровской свитах Западного побережья, маруямской свите Южного Сахалина. Средний–верхний миоцен Западной Камчатки; ильинская, какертская и этолонская свиты. Верхний миоцен–плейстоцен Восточной Камчатки (юнюньваямская, усть-лимимтеваямская и ольховская свиты).
Материал. 28 экз.; Россия, Восточная Камчатка, о. Карагинский; верхний миоцен, юнюньваямская свита.
Cribroelphidium crassum (V. Kuznetsova, 1970)
Табл. IV , фиг. 13, 14
Cribroelphidium crassum: Волошинова и др., 1970, с. 168, табл. 43 , фиг. 6–8 ; Бордунов, 2015, табл. XIII , фиг. 12, 13 ; Бордунов и др., 2018, табл. VII , фиг. 7, 8 .
Голотип – ВНИГРИ, № 534/11; Сахалин; нижний миоцен, тумская свита.
Описание. Раковина крупная, инволютная, округлая в профиль. В наружном обороте насчитываются восемь–десять довольно широких камер. Камеры немного выпуклые, что создает слегка волнистый контур. Периферический край широкий, округлый, выровненный с внешней стороны. Пупочная область вдавленная, пупочная шишка состоит из крупных гранул. Швы очень отчетливые, заглубленные, широкие, уплощенные, немного изогнутые. Септальные мостики и ямки отчетливые. Устьевая поверхность широкая, выпуклая, с круглыми сравнительно большими устьевыми отверстиями и фораменом в основании.
Размеры в мкм. Диаметр 300–600, толщина 150–300.
Сравнение. Имеет определенное сходство с более молодым видом C. subglobosum (Voloshinova). Отличается большими размерами, более плоскими камерами, менее углубленной пупочной областью.
Распространение. Олигоцен–нижний миоцен Сахалина (холмская свита) и Камчатки (вивентекская, кулувенская, ильинская свиты).
Материал. Семь экз.; Россия, Западная Камчатка, бухта Квачина; нижний миоцен, кулувенская свита.
ПОДСЕМЕЙСТВО ELPHIDIELLIDAE HOLZMANN ET PAWLOWSKI, 2017
Род Elphidiella Cushman, 1936
Elphidiella oregonensis (Cushman et Grant, 1927)
Табл. IV , фиг. 3, 4
Elphidium oregonense: Cushman, Grant, 1927, с. 79, табл. 8 , фиг. 3 ; Cushman, 1939, с. 50, табл. 13 , фиг. 14–16 ; Волошинова, Дайн, 1952, с. 50, табл. 6 , фиг. 9 , табл. 7 , фиг. 6 ; Loeblich, Tappan, 1953, с. 103, табл. 18 , фиг. 1–3 .
Elphidium yezoense: Asano, 1937, с. 589, табл. 14 (3), фиг. 11 .
Elphidiella oregonensis: Волошинова и др., 1970, с. 176, табл. XLVI , фиг. 1, 2 ; Серова, 1978, с. 117, табл. IX , фиг. 6 а, 6в; Серова, Спирина, 1984, с. 128, табл. XVIII , фиг. 2, 3 ; Бугрова и др., 2005, табл. 62 , фиг. 2 ; Narayan et al., 2005, с. 128, табл. 3 , фиг. 29, 30 ; Бордунов, 2015, табл. XI , фиг. 1, 5 .
Голотип – Mus. Calif. Acad. Sci., № 2809; Калифорния; плейстоцен.
Описание. Раковина крупная, бывает более 1 мм в диаметре, сплюснутая с боков, инволютная, иногда полуэволютная, с округлым контуром. Пупочная область ровная, не вдавленная, закрытая крупными пупочными шишками, покрытыми крупными отверстиями пупочных каналов. Последний оборот раковины состоит из большого числа, до 20, немного вздутых камер. Периферический край ровный, слегка волнистый в области последних камер. Швы хорошо различимые, слегка вдавленные, ограниченные двумя рядами крупных пор, наполненными мелкими бугорками. Стенки раковины гладкие, полупрозрачные, толстые и покрытые мелкими порами. Устье состоит из щели в основании устьевой поверхности с круглыми порами, нерегулярно расположенными посередине.
Размеры в мкм. Диаметр 550–1500, толщина 150–300.
Сравнение. Имеет внешнее сходство с E. nabilensis, однако отличается от последнего отсутствием вдавленного пупка и поверхностной пупочной шишкой.
Замечания. Elphidium yezoense Asano, 1937 из плиоценовых отложений севера Японии, вероятно, является младшим синонимом описываемого вида.
Распространение. Плиоцен–плейстоцен Дальнего Востока, Калифорнии, Японии (формация Такикава, слои с Fortipecten takahashii); верхний миоцен–плиоцен Сахалина (верхненутовская свита); плиоцен–плейстоцен Северного Сахалина (помырская свита); нижний плиоцен Южного Сахалина (маруямская свита, слои с Fortipecten takahashii); в нижнем плиоцене Камчатки образуют монотаксонные комплексы в основании энемтенской и лимимтеваямской свит.
Материал. 45 экз.; Россия, Восточная Камчатка, о. Карагинский; нижний плиоцен, лимимтеваямская свита.
Elphidiella simplex Voloshinova, 1958
Табл. IV , фиг. 9, 10
Elphidiella simplex: Волошинова, 1958, с. 179, табл. VIII , фиг. 1–4 , 6, 7 ; Волошинова и др., 1970, с. 173, табл. XLV , фиг. 3–5 ; Бугрова и др., 2005, табл. 62 , фиг. 1а, 1б; Бордунов, 2015, табл. IX , фиг. 8 .
Голотип – ВНИГРИ, № 442/103; Сахалин, Катангли; нижний миоцен, дагинская свита.
Описание. Раковина относительно небольших размеров для рода, с овальным контуром. Периферический край узкозакругленный, ровный. Наружный оборот состоит из 10–12, до 15 камер. Камеры слабоизогнутые, плоские. Швы широкие, слабоуглубленные, слабоизогнутые; довольно отчетливо видны просвечивающие меридиональные каналы. Пупочная область плоская или слегка углубленная. Иногда формируется небольшая пупочная шишка. Септальные швы до центра ее не доходят, оставляя узкое пространство неправильной формы. Устьевая поверхность высокая и сравнительно узкая. Форамен в виде очень мелких многочисленных отверстий в основании устьевой поверхности, обычно разрывающихся и образующих подобие длинной неровной щели. На септах два ряда отверстий отчетливо видны на последних камерах наружного оборота.
Размеры в мкм. Диаметр 350–600, толщина 150–250.
Сравнение. Данный вид имеет определенное сходство с E. tenera, от которого отличается в первую очередь строением швов и контуром раковины, вследствие более быстрого нарастания камер в ширину.
Замечания. Изменчивость выражается, главным образом, в большей или меньшей степени уплощенности раковины.
Распространение. Миоцен Сахалина (уйнинская, дагинская, окобыкайская свиты), верхний миоцен–плиоцен Камчатки (этолонская, лимимтеваямская свиты).
Материал. Пять экз.; Россия, Восточная Камчатка, о. Карагинский; нижний плиоцен, лимимтеваямская свита.
Род Pseudoelphidiella Voloshinova et V. Kuznetsova, 1970
Pseudoelphidiella subcarinata (Voloshinova, 1958)
Табл. IV , фиг. 11, 12
Cribrononion subcarinatus: Волошинова, 1958, с. 137, табл. I , фиг. 13–15 .
Pseudoelphidiella subcarinatа: Волошинова и др., 1970, с. 180, табл. LI , фиг. 8, 9 ; Серова, 1978, с. 120, табл. X , фиг. 1а, 1в; Серова, Спирина, 1984, с. 121, табл. XIX , фиг. 5 а, 5б; Бугрова и др., 2005, табл. 62 , фиг. 3, 4 ; Бордунов, 2015, с. 115, табл. XI , фиг. 1, 3 ; Бордунов и др., 2018, табл. XII , фиг. 1, 2 , табл. XIV, фиг. 9, 10 .
Голотип – ВНИГРИ, № 442/79; Сахалин, Катангли; нижний миоцен, дагинская свита.
Описание. Раковины средних и крупных для рода размеров, дисковидной формы, с узким периферическим краем и выпуклой пупочной областью. Контур округлый. Наружный оборот состоит из 15–20 камер, незначительно увеличивающихся в размерах по мере нарастания. Камеры соединены плоскими двухконтурными швами с очень мелкими, часто неясными септальными ямками в два ряда. Пупочная область покрыта прозрачным скелетным веществом. Устье дырчатое, в основании септальной поверхности.
Размеры в мкм. Диаметр 400–1000, толщина 150–350.
Сравнение. Наблюдается сходство с более молодой формой P. hannai (Cushman et Grant, 1927) по дисковидной форме раковины, но имеет большие размеры раковины.
Замечания. Значительное сходство имеет с Elphidium satoi Asano et Murata, 1958 из олигоцена Японии (формация Кисима) (Asano, 1958), от которого отличается большим числом камер в последнем обороте и более узкой раковиной.
Распространение. Олигоцен–нижний миоцен Сахалина (дагинская, борская, тумская, уйнинская, мачигарская свиты), нижний–средний миоцен Камчатки (кулувенская, пестроцветная, пахачинская, ильинская свиты), олигоцен Японии (формации Кисимаи, Асагай).
Материал. Восемь экз.; Россия, Западная Камчатка, бухта Квачина; нижний миоцен, кулувенская свита.
Список литературы
Большаков А.К., Большакова Р.А., Шаинян С.Х. О возрасте осадочных образований Северо-Охотского прогиба // Возраст геологических образований Охотоморского региона и прилегающих территорий. Владивосток: ДВО АН СССР, 1989. С. 16–26.
Бордунов С.И. Опыт расчленения неогена Восточной Камчатки по бентосным фораминиферам. Изменение структуры древних сообществ // Стратиграфия в начале XXI века – тенденции и новые идеи. М.: Геокарт, ГЕОС, 2013. С. 29–39.
Бордунов С.И. Стратиграфия и фораминиферы неогена Восточной Камчатки. М.: ГЕОС, 2015. 148 с.
Бордунов С.И., Дмитриева Т.В., Фрегатова Н.А. Значение фораминифер для расчленения разреза и выделения региональных биостратиграфических зон // Опорный разрез кайнозоя Западно-Камчатской структурно-формационной зоны. М.: Геокарт, ГЕОС, 2018. С. 42–53.
Бугрова Э.М., Гладкова В.И., Дмитриева Т.В. и др. Практическое руководство по микрофауне. Т. 8. Фораминиферы кайнозоя. СПб.: Изд-во ВСЕГЕИ, 2005. 324 с.
Волошинова Н.А. О новой систематике нонионид // Микрофауна CCCР. Л., 1958. С. 117–223. (Тр. ВНИГРИ. Сб. 9. Вып. 115).
Волошинова Н.А., Дайн Л.Г. Нониониды, кассидулиниды и хилостомеллиды // Ископаемые фораминиферы СССР. Л., М.: Гостоптехиздат, 1952. С. 1–114. (Тр. ВНИГРИ. Нов. Сер. Вып. 63).
Волошинова Н.А., Кузнецова В.Н. Новые данные по морфологии и эволюционному развитию некоторых представителей семейства Elphidiidae // Вопросы микропалеонтологии. Вып. 8. М.: Наука, 1964. С. 138–153.
Волошинова Н.А., Кузнецова В.Н., Леоненко Л.С. Фораминиферы неогеновых отложений Сахалина. Л.: Недра, 1970. 304 с.
Гладенков Ю.Б., Гладенков А.Ю. Начало формирования межокеанических связей Пацифики и Арктики через Берингов пролив в неогене // Стратигр. Геол. корреляция. 2004. Т. 12. № 2. С. 72–89.
Гладенков Ю.Б. Северотихоокеанская область в палеогене и неогене как пример полузамкнутой морской экосистемы // Стратигр. Геол. корреляция. 2015. Т. 23. № 4. С. 91–105.
Гладенков Ю.Б., Синельникова В.Н. Моллюски и климатические оптимумы миоцена Камчатки. М.: Наука, 1990. 171 с.
Гладенков Ю.Б., Шанцер А.Е. Геологические события палеогена Камчатки // Стратигр. Геол. корреляция. 1993. Т. 1. № 1. С. 97–108.
Гудина В.И. Фораминиферы и стратиграфия четвертичных отложений северо-запада Сибири. М.: Наука, 1966. 146 с.
Гудина В.И. Морской плейстоцен сибирских равнин. Фораминиферы Енисейского севера. М.: Наука, 1969. 80 с.
Гудина В.И., Левчук Л.К. Морфология и классификация эльфидиид (Foraminifera) // Морфология и систематика беспозвоночных фанерозоя. Новосибирск, 1983. С. 28–37 (Тр. Ин-та геол. и геофиз. СО АН СССР. Вып. 538).
Дмитриева Т.В. Зональная шкала по бентосным фораминиферам продуктивных отложений палеогена и неогена Охотской нефтегазоносной провинции // Стратиграфия и ее роль в развитии нефтегазового комплекса России. СПб.: ВНИГРИ, 2007. С. 376–395.
Дмитриева Т.В., Мещерякова Е.Ю., Фрегатова Н.А. и др. Новые данные по биостратиграфии палеоценовых отложений Западной Камчатки (фораминиферы, диноцисты, споры и пыльца) // Современная микропалеонтология. Сб. трудов XVI Всеросс. микропалеонтол. совещ. Калининград, 2015. С. 15–19.
Жаров А.Э., Митрофанова Л.И., Тузов В.П. Стратиграфия кайнозойских отложений шельфа Северного Сахалина // Стратигр. Геол. корреляция. 2013. Т. 21. № 5. С. 72–93.
Крашенинников В.А. Эльфидииды миоценовых отложений Подолии. М.: Изд-во АН СССР, 1960. 165 c. (Тр. Геол. ин-та АН СССР. Вып. 21).
Овсепян Я.С., Талденкова Е.Е., Баух Х.А. и др. Реконструкция событий позднего плейстоцена–голоцена на континентальном склоне моря Лаптевых по комплексам бентосных и планктонных фораминифер // Стратигр. Геол. корреляция, 2015. Т. 23. № 6. С. 96–112.
Ольшанецкий Д.М. Нониониды снатольской свиты в Увучинском разрезе Охотоморского побережья Западной Камчатки // Тр. XVII Всеросс. микропалеонтол. совещ. “Современная микропалеонтология – проблемы и перспективы” (24–29 сентября 2018 г., Казань). М.: ПИН РАН, 2018а. С. 101–105.
Ольшанецкий Д.М. Стратиграфическая схема нижнего палеогена Северной Пацифики по глубоководным бентосным фораминиферам // Тихоокеанск. геол. 2018б. Т. 37. № 2. С. 17–42.
Плетнев С.П. Изменение экоструктуры кайнозойских фораминифер как основа для реконструкции глубин Охотоморского седиментационного бассейна // Геол. и полезн. ископ. мирового океана. 2007. №1(7). С. 99–109.
Савельева О.Л. Сопоставление климатических и биотических событий с вулканической активностью в кайнозое Камчатки // Проблемы эксплозивного вулканизма (к 50-летию катастрофического извержения вулкана Безымянный). Матер. первого междунар. симп. Петропавловск-Камчатский, 2006. С. 170–177.
Саидова Х.М. Бентосные фораминиферы Тихого океана. М.: Изд-во ИО АН СССР, 1975. 160 с.
Серова М.Я. Стратиграфия и фораминиферы неогена Камчатки. М.: Наука, 1978. 176 c. (Тр. Геол. ин-та АН СССР. Вып. 323).
Серова М.Я. Фораминиферы и биостратиграфия верхнего палеогена Северной Пацифики. М.: Наука, 2001. 215 с.
Серова М.Я., Спирина Л.В. Биостратиграфический анализ. Фораминиферы // Атлас фауны и флоры неогеновых отложений Дальнего Востока. Точилинский опорный разрез Западной Камчатки. М., 1984. С. 61–67 (Тр. Геол. ин-та АН СССР. Вып. 385).
Тузов В.П., Митрофанова Л.И., Данченко Р.В. и др. Стратиграфия палеогеновых отложений Колпаковского прогиба Западной Камчатки) // Стратигр. Геол. корреляция. 1997. Т. 5. № 3. С. 66–82.
Фрегатова Н.А., Беньямовский В.Н. Слои с бентосными фораминиферами // Стратиграфия морского палеоцена и нижнего эоцена Западной Камчатки. М.: ГЕОС, 2013. С. 68–85.
Фрегатова Н.А., Дмитриева Т.В. Разработка зонального стандарта верхнего олигоцена–миоцена Западной Камчатки по бентосным фораминиферам // Стратиграфия нефтегазоносных бассейнов. СПб.: Недра, 2004. С. 177–198.
Фурсенко А.В., Троицкая Т.С., Левчук Л.К. и др. Фораминиферы Дальневосточных морей СССР. Новосибирск, 1979. 398 с. (Тр. ИГиГ СО АН СССР. Вып. 387).
Хорева И.М. Фораминиферы антропогена северо-западного обрамления Тихого океана. М.: Наука, 1988. 104 с.
Щедрина З.Г. Новые данные по фауне фораминифер Охотского моря и ее распространение // Тр. Зоол. ин-та АН СССР. 1953. Т. 13. С. 12–32.
Asano K. On the Japanese species of Elphidium and its allied genera // Trans. Paleontol. Soc. Japan. 1937. V. 45. № 538. P. 581–591.
Asano K., Murata S. Some Paleogene smaller Foraminifera from Japan // Sci. Rep. Toh. Univ. 2nd ser. Geol. 1958. V. 29. P. 43–75.
Buzas M.A., Culvers S.J., Isham L.B. A comparison of fourteen elphidiid (Foraminiferida) taxa // J. Paleontol. 1985. V. 59. № 5. P. 1075–1090.
Cushman J.A. The Foraminifera of the Atlantic Ocean. Part 7. Nonionidae, Camerinidae, Peneroplidae and Alveolinellidae // Bull. U.S. Nat. Mus. 1930. V. 104. № 7. P. 1–79.
Cushman J.A. Some new species of Elphidium and related genera. // Contrib. Cushman lab. for foraminiferal res. 1936. V. 12. № 4. P. 78–89.
Cushman J.A. A Monograph of the Foraminiferal Family Nonionidae // U.S. Dep. Inter. Geol. Surv. 1939. Prof. Paper 191. 124 p.
Cushman J.A., Grant U.S. Late Tertiary and Quaternary Elphidiums of the west coast of North America // Trans. San Diego Soc. Natur. Hist. 1927. V. 5. № 6. P. 69–82.
Cushman J.A., Stewart R., Stewart K. Tertiary Foraminifera from Humholdt County. A preliminarv survey of the fauna // Trans. San Diego Soc. Natur. Hist. 1930. V. 6. P. 41–94.
Galloway J.J. A Manual of Foraminifera. Bloomington: Principia Press, 1933. 483 p.
Gladenkov A.Y., Oleinik A.E., Marincovich L. Jr., Barinov K.B. A refined age for the earliest opening of Bering Strait // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2002. V. 183. P. 321–328.
Gross O. Foraminifera // European register of marine species: a check-list of the marine species in Europe and a bibliography of guides to their identification // Collection Patrimoines Naturels. 2001. № 50. P. 60–75.
Hayward B.W., Gross O. Elphidiinae Galloway, 1933. World Foraminifera Database. 2021. http://marinespecies.org/ aphia.php?p=taxdetails&id=721061
Hayward B.W., Hollis C.J., Grenfell H.R. Recent Elphidiidae (Foraminiferida) of the South-West Pacific and Fossil Elphidiidae of New Zealand // Monogr. Inst. Geol. and Nuclear Sci. 1997. № 16. 166 p.
Kaiho K. Paleogene Foraminifera from Hokkaido, Japan. Part 1. Lithostratigraphy and Biostratigraphy Including Description of New Species // Sci. Rep. Toh. Univ. 2nd ser. 1984. V. 55. № 1. 74 p.
Lei Y., Li T. Atlas of Benthic Foraminifera from China Seas the Bohai Sea and the Yellow Sea. Beijing: Springer Geol. and Sci. Press, 2016. 399 p.
Loeblich A.R., Tappan H. Studies of Arctic Foraminifera // Smithson. Misc. Coll. 1953. V. 121. № 7. 150 p.
Loeblich A.R., Tappan H. Foraminiferal Genera and Their Classification. N.Y.: Van Nostrand Reinhold Company, 1987. 970 p.
Lukina T.G. Foraminifera of the Laptev Sea // Protistology. 2001. V. 2. № 2. P. 105–122.
Mallory V.S. Lower Tertiary Biostratigraphy of the California Coast Ranges. Tulsa, Oklahoma: Amer. Assoc. Petrol. Geol., 1959. 416 p.
Narayan Y.B., Barnes C.R., Johns M.J. Taxonomy and biostratigraphy of Cenozoic foraminifers from Shell Canada wells, offshore Vancouver Island, Toffino Bay, B. C. // Micropaleontol. 2005. V. 51. № 2. P. 101–167.
Osterman L. Pliocene and Quaternary benthic foraminifers from Site 910, Yermak Plateau // Proc. Ocean Drilling Program, Sci. Results. 1996. V. 151. P. 187–195. https://doi.org/10.2973/odp.proc.sr.151.107.1996
Phleger F.B. Foraminifera distribution in some sediment samples from the Canadian and Greenland Arctic // Cushman Found. Foram. Res. 1952. Contrib. 3. P. 80–89.
Rau W. Foraminifera, Stratigraphy, and Paleoecology of the Quinault Formation, Point Grenville-Raft River Coastal Area, Washington // Bull. Wash. Dep. Natur. Res. 1970. № 62. 48 p.
Wade M. Morphology and taxonomy of the foraminiferal family Elphidiidae // J. Wash. Acad. Sci. 1957. V. 47. № 10. P. 330–339.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал